Prevention of Recurrent Acute Otitis Media in Children Through the Use of Lactobacillus salivarius PS7, a Target-Specific Probiotic Strain


Abstract
:1. Introduction
2. Materials and Methods
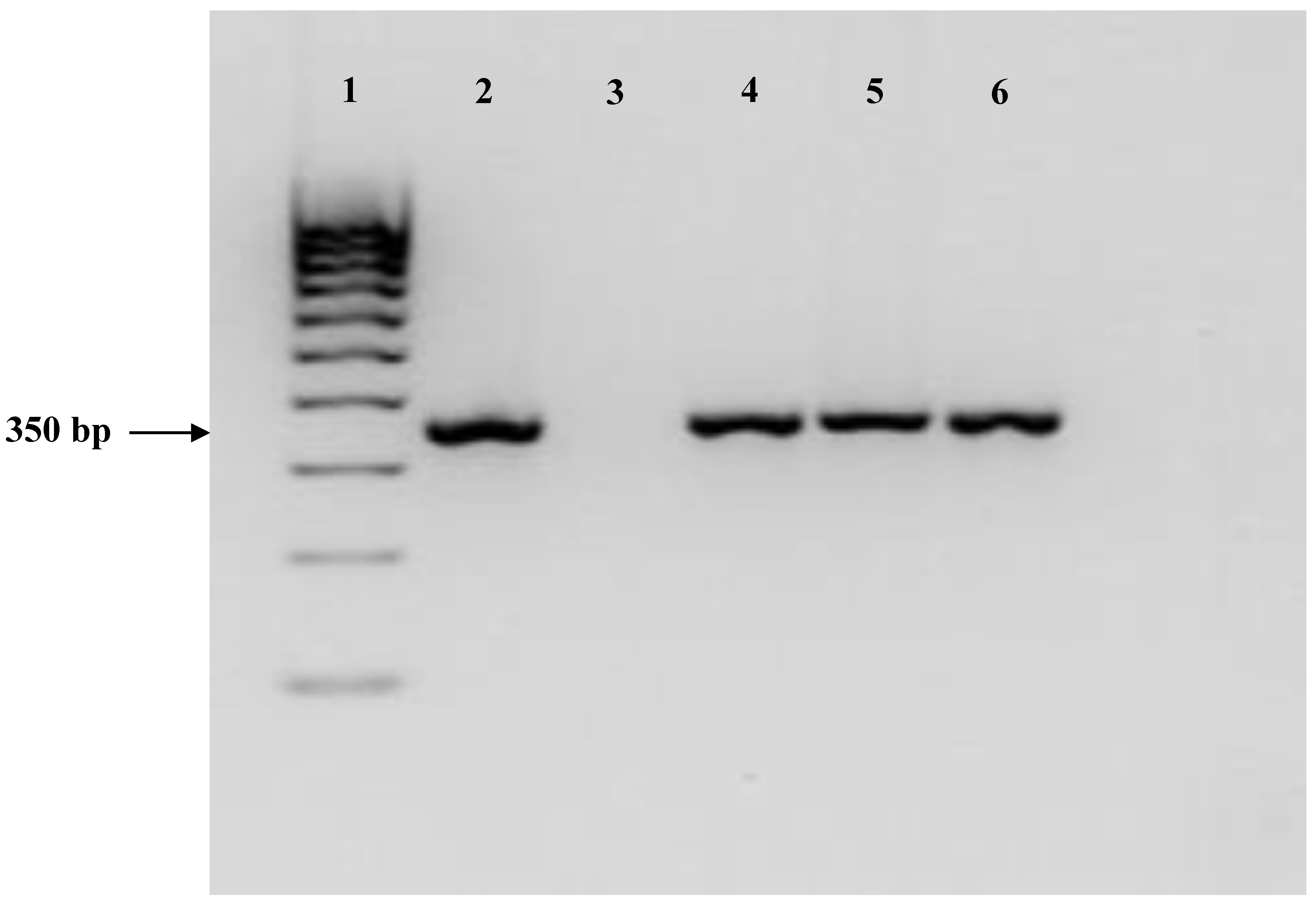
2.1. Isolation and Identification of the Strain
2.2. Survival after In Vitro Exposure to Saliva and Gastrointestinal-Like Conditions
2.3. Adhesion to Caco-2/HT-29 Cells
2.4. Production of Riboflavin, Folate and Cyanocobalamin
2.5. Antimicrobial Activity of Strain PS7 against AOM-Related Pathogens
2.6. Production of Specific Antimicrobials (Bacteriocins, Lactic Acid, Acetic Acid, Hydrogen Peroxide) by Strain PS7
2.7. Coagregation Assays
2.8. Co-Culture Studies
2.9. Sensitivity to Antibiotics
2.10. Formation of Biogenic Amines and Degradation of Mucin
2.11. Acute and Repeated Dose (4-Weeks) Oral Toxicity Studies in a Rat Model
2.12. Pilot Clinical Trial: Prevention of rAOM in Children
2.13. Statistical Analysis
3. Results
3.1. Identification and In Vitro Characterization of the Strain
3.2. Acute and Repeated Dose (4-Weeks) oral Toxicity Studies in a Rat Model
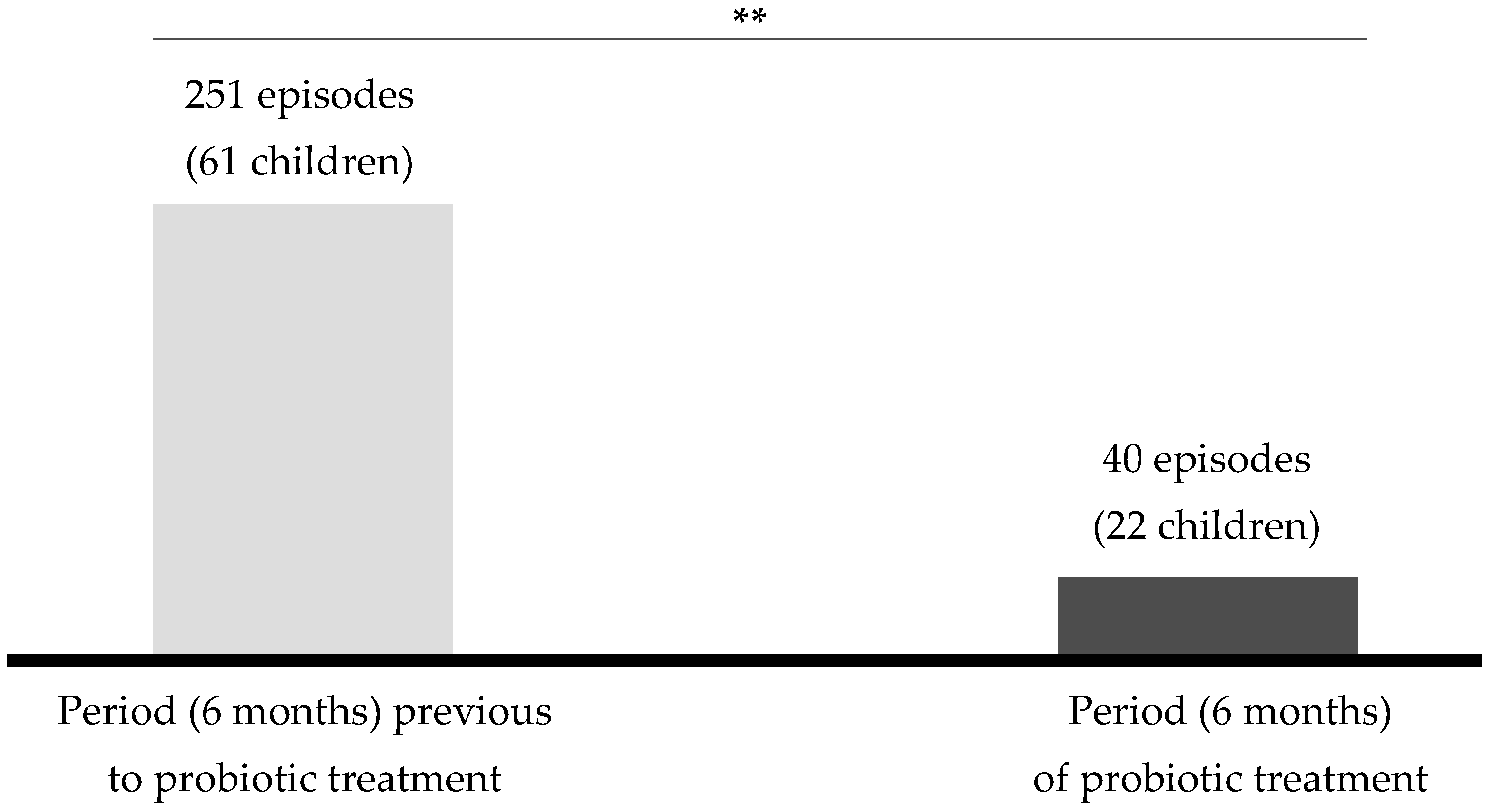
3.3. Pilot Clinical Trial: Prevention of AOM in Children
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Rovers, M.M. The burden of otitis media. Vaccine 2008, 26, G2–G4. [Google Scholar] [CrossRef] [PubMed]
- Venekamp, R.P.; Sanders, S.L.; Glasziou, P.P.; Del Mar, C.B.; Rovers, M.M. Antibiotics for acute otitis media in children. Cochrane Database Syst. Rev. 2015, CD000219. [Google Scholar] [CrossRef]
- Klein, J.O. The burden of otitis media. Vaccine 2000, 19, S2–S8. [Google Scholar] [CrossRef]
- Pelton, S.I.; Leibovitz, E. Recent advances in otitis media. Pediatr. Infect. Dis. J. 2009, 28, S133–S137. [Google Scholar] [CrossRef] [PubMed]
- Pichichero, M.E. Ten-year study of acute otitis media in Rochester, NY. Pediatr. Infect. Dis. J. 2016, 35, 1027–1032. [Google Scholar] [CrossRef] [PubMed]
- Jervis-Bardy, J.; Rogers, G.B.; Morris, P.S.; Smith-Vaughan, H.C.; Nosworthy, E.; Leong, L.E.; Smith, R.J.; Weyrich, L.S.; de Haan, J.; Carney, A.S.; et al. The microbiome of otitis media with effusion in Indigenous Australian children. Int. J. Pediatr. Otorhinolaryngol. 2015, 79, 1548–1555. [Google Scholar] [CrossRef] [PubMed]
- Chan, C.L.; Wabnitz, D.; Bardy, J.J.; Bassiouni, A.; Wormald, P.J.; Vreugde, S.; Psaltis, A.J. The microbiome of otitis media with effusion. Laryngoscope 2016, 126, 2844–2851. [Google Scholar] [CrossRef]
- Ngo, C.C.; Massa, H.M.; Thornton, R.B.; Cripps, A.W. Predominant bacteria detected from the middle ear fluid of children experiencing otitis media: A systematic review. PLoS ONE 2016, 11, e0150949. [Google Scholar] [CrossRef]
- Granath, A. Recurrent acute otitis media: What are the options for treatment and prevention? Curr. Otorrhinolatyngol. Rep. 2017, 5, 93–100. [Google Scholar] [CrossRef]
- Karlidağ, T.; Demirdağ, K.; Kaygusuz, I.; Ozden, M.; Yalçin, S.; Oztürk, L. Resistant bacteria in the adenoid tissues of children with otitis media with effusion. Int. J. Pediatr. Otorhinolaryngol. 2002, 64, 35–40. [Google Scholar] [CrossRef]
- Brook, I.; Gober, A.E. In vitro bacterial interference in the nasopharynx of otitis media-prone and non-otitis media-prone children. Arch. Otolaryngol. Head Neck Surg. 2000, 126, 1011–1013. [Google Scholar] [CrossRef] [PubMed]
- Leibovitz, E.; Greenberg, D.; Piglansky, L.; Raiz, S.; Porat, N.; Press, J.; Leiberman, A.; Dagan, R. Recurrent acute otitis media occurring within one month from completion of antibiotictherapy: Relationship to the original pathogen. Pediatr. Infect. Dis. J. 2003, 22, 209–216. [Google Scholar] [CrossRef] [PubMed]
- Marchisio, P.; Claut, L.; Rognoni, A.; Esposito, S.; Passali, D.; Bellussi, L.; Drago, L.; Pozzi, G.; Mannelli, S.; Schito, G.; et al. Differences in nasopharyngeal bacterial flora in children with nonsevere recurrent acute otitis media and chronic otitis media with effusion: Implications for management. Pediatr. Infect. Dis. J. 2003, 22, 262–268. [Google Scholar] [CrossRef]
- Klein, J.O. Bacterial resistance and antimicrobial drug selection. In Evidence Based Otitis Media; BC Decker: Hamilton, ON, Canada, 2003; pp. 429–437. [Google Scholar]
- Hall-Stoodley, L.; Hu, F.Z.; Gieseke, A.; Nistico, L.; Nguyen, D.; Hayes, J. Direct detection of bacterial biofilms on the middle-ear mucosa of children with chronic otitis media. JAMA 2006, 296, 202–211. [Google Scholar] [CrossRef] [PubMed]
- Thornton, R.B.; Rigby, P.J.; Wiertsema, S.P.; Filion, P.; Langlands, J.; Coates, H.L.; Vijayasekaran, S.; Keil, A.D.; Richmond, P.C. Multi-species bacterial biofilm and intracellular infection in otitis media. BMC Pediatr. 2011, 11, 94. [Google Scholar] [CrossRef] [PubMed]
- Chan, C.L.; Richter, K.; Wormald, P.J.; Psaltis, A.J.; Vreugde, S. Alloiococcus otitidis forms multispecies biofilm with Haemophilus influenzae: Effects on antibiotic susceptibility and growth in adverse conditions. Front. Cell Infect. Microbiol. 2017, 7, 344. [Google Scholar] [CrossRef] [PubMed]
- Fergie, N.; Bayston, R.; Pearson, J.P.; Birchall, J.P. Is otitis media with effusion a biofilm infection? Clin. Otolaryngol. Allied Sci. 2004, 29, 38–46. [Google Scholar] [CrossRef]
- Tanaka, S.; Kobayashi, T.; Songjinda, P.; Tateyama, A.; Tsubouchi, M.; Kiyohara, C.; Shirakawa, T.; Sonomoto, K.; Nakayama, J. Influence of antibiotic exposure in the early postnatal period on the development of intestinal microbiota. FEMS Immunol. Med. Microbiol. 2009, 56, 80–87. [Google Scholar] [CrossRef] [Green Version]
- Blaser, M.J.; Falkow, S. What are the consequences of the disappearing human microbiota? Nat. Rev. Microbiol. 2009, 7, 887–894. [Google Scholar] [CrossRef]
- Rutten, N.B.; Rijkers, G.T.; Meijssen, C.B.; Crijns, C.E.; Oudshoorn, J.H.; van der Ent, C.K.; Vlieger, A.M. Intestinal microbiota composition after antibiotic treatment in early life: The INCA study. BMC Pediatr. 2015, 15, 204. [Google Scholar] [CrossRef]
- Azad, M.B.; Konya, T.; Persaud, R.R.; Guttman, D.S.; Chari, R.S.; Field, C.J.; Sears, M.R.; Mandhane, P.J.; Turvey, S.E.; Subbarao, P.; et al. Impact of maternal intrapartum antibiotics, method of birth and breastfeeding on gut microbiota during the first year of life: A prospective cohort study. BJOG 2016, 123, 983–993. [Google Scholar] [CrossRef] [PubMed]
- Lewis, J.D.; Chen, E.Z.; Baldassano, R.N.; Otley, A.R.; Griffiths, A.M.; Lee, D.; Bittinger, K.; Bailey, A.; Friedman, E.S.; Hoffmann, C.; et al. Inflammation, antibiotics, and diet as environmental stressors of the gut microbiome in pediatric Crohn’s disease. Cell Host Microbe 2015, 18, 489–500. [Google Scholar] [CrossRef] [PubMed]
- Zeissig, S.; Blumberg, R.S. Life at the beginning: Perturbation of the microbiota by antibiotics in early life and its role in health and disease. Nat. Immunol. 2014, 15, 307–310. [Google Scholar] [CrossRef] [PubMed]
- Lappan, R.; Imbrogno, K.; Sikazwe, C.; Anderson, D.; Mok, D.; Coates, H.; Vijayasekaran, S.; Bumbak, P.; Blyth, C.C.; Jamieson, S.E.; et al. A microbiome case-control study of recurrent acute otitis media identified potentially protective bacterial genera. BMC Microbiol. 2018, 18, 13. [Google Scholar] [CrossRef] [PubMed]
- Niittynen, L.; Pitkäranta, A.; Korpela, R. Probiotics and otitis media in children. Int. J. Pediatr. Otorhinolaryngol. 2012, 76, 465–470. [Google Scholar] [CrossRef] [PubMed]
- Kullen, M.J.; Sanozky-Dawes, R.B.; Crowell, D.C.; Klaenhammer, T.R. Use of DNA sequence of variable regions of the 16SrRNA gene for rapid and accurate identification of bacteria in the Lactobacillus acidophilus complex. J. Appl. Microbiol. 2000, 89, 511–518. [Google Scholar] [CrossRef] [PubMed]
- Marín, M.; Arroyo, R.; Espinosa-Martos, I.; Fernández, L.; Rodríguez, J.M. Identification of emerging human mastitis pathogens by MALDI-TOF and assessment of their antibiotic resistance patterns. Front. Microbiol. 2017, 8, 1258. [Google Scholar] [CrossRef] [PubMed]
- Martín, V.; Maldonado-Barragán, A.; Moles, L.; Rodriguez-Baños, M.; Campo, R.D.; Fernández, L.; Rodríguez, J.M.; Jiménez, E. Sharing of bacterial strains between breast milk and infant feces. J. Hum. Lact. 2012, 28, 36–44. [Google Scholar] [CrossRef]
- Marteau, P.; Minekus, M.; Havenaar, R.; Huis In’t Veld, J.H.J. Survival of lactic acid bacteria in a dynamic model of the stomach and small intestine: Validation and the effects of bile. J. Dairy Sci. 1997, 80, 1031–1037. [Google Scholar] [CrossRef]
- Martín, R.; Olivares, M.; Marín, M.L.; Fernández, L.; Xaus, J.; Rodríguez, J.M. Probiotic potential of 3 Lactobacilli strains isolated from breast milk. J. Hum. Lact. 2005, 21, 8–17. [Google Scholar] [CrossRef]
- Conway, P.L.; Gorbach, S.L.; Goldin, B.R. Survival of lactic acid bacteria in the human stomach and adhesion to intestinal cells. J. Dairy Sci. 1987, 70, 1–12. [Google Scholar] [CrossRef]
- Coconnier, M.H.; Klaenhammer, T.R.; Kernéis, S.; Bernet, M.F.; Servin, A.L. Protein-mediated adhesion of Lactobacillus acidophilus BG2FO4 on human enterocyte and mucus-secreting cell lines in culture. Appl. Environ. Microbiol. 1992, 58, 2034–2039. [Google Scholar] [PubMed]
- Juarez del Valle, M.; Laiño, J.; Savoy de Giori, G.; LeBlanc, J.G. Riboflavin producing lactic acid bacteria as a biotechnological strategy to obtain bio-enriched soymilk. Food Res. Int. 2014, 62, 1015–1019. [Google Scholar] [CrossRef]
- Laiño, J.E.; LeBlanc, J.G.; Savoy de Giori, G. Production of natural folates by lactic acid bacteria starter cultures isolated from artisanal Argentinean yogurts. Can. J. Microbiol. 2012, 58, 581–588. [Google Scholar] [CrossRef] [PubMed]
- Horwitz, W. Official Methods of Analysis of AOAC International; AOAC International: Gaithersburg, MD, USA, 2000. [Google Scholar]
- Magnusson, J.; Schnürer, J. Lactobacillus coryniformis subsp. coryniformis strain Si3 produces a broad-spectrum proteinaceous antifungal compound. Appl. Environ. Microbiol. 2001, 67, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Dodd, H.M.; Horn, N.; Zhang, H.; Gasson, M.J. A lactococcal expression system for engineered nisins. Appl. Environ. Microbiol. 1992, 58, 3683–3693. [Google Scholar] [PubMed]
- Çataloluk, O. Molecular characterization of the gene encoding for salivaricin B activity and its flanking sequences. Turk. J. Biol. 2001, 25, 379–386. [Google Scholar]
- Flynn, S.; van Sinderen, D.; Thornton, G.M.; Holo, H.; Nes, I.F.; Collins, J.K. Characterization of the genetic locus responsible for the production of ABP-118, a novel bacteriocin produced by the probiotic bacterium Lactobacillus salivarius subsp. salivarius UCC118. Microbiology 2002, 148, 973–984. [Google Scholar] [CrossRef]
- Song, Y.L.; Kato, N.; Matsumiy, Y.; Lu, C.X.; Kato, H.; Watanabe, K. Identification of an hydrogen peroxide production by fecal and vaginal lactobacilli isolated from Japanese women and newborn infants. J. Clin. Microbiol. 1999, 37, 3062–3064. [Google Scholar]
- Yap, P.S.; Gilliland, S.E. Comparison of newly isolated strains of Lactobacillus delbrueckii susp. lactis for hydrogen peroxide production at 5 °C. J. Dairy Sci. 2000, 83, 628–632. [Google Scholar] [CrossRef]
- Reid, G.; McGroarty, J.A.; Gil Domingue, P.A.; Chow, A.W.; Bruce, A.W.; Eisen, A.; Costerton, J.W. Coaggregation of urogenital bacterial in vitro and in vivo. Curr. Microbiol. 1990, 20, 47–52. [Google Scholar] [CrossRef]
- Younes, J.A.; van der Mei, H.C.; van den Heuvel, E.; Busscher, H.J.; Reid, G. Adhesion forces and coaggregation between vaginal staphylococci and lactobacilli. PLoS ONE 2012, 7, e36917. [Google Scholar] [CrossRef] [PubMed]
- Klare, I.; Konstabel, C.; Müller-Bertling, S.; Reissbrodt, R.; Huys, G.; Vancanneyt, M.; Swings, J.; Goossens, H.; Witte, W. Evaluation of new broth media for microdilution antibiotic susceptibility testing of lactobacilli, lactococci, pediococci, and bifidobacteria. Appl. Environ. Microbiol. 2005, 71, 8982–8986. [Google Scholar] [CrossRef] [PubMed]
- Langa, S.; Maldonado-Barragán, A.; Delgado, S.; Martín, R.; Martín, V.; Jiménez, E.; Ruíz-Barba, J.L.; Mayo, B.; Connor, R.I.; Suárez, J.E.; et al. Characterization of Lactobacillus salivarius CECT 5713, a strain isolated from human milk: From genotype to phenotype. Appl. Microbiol. Biotechnol. 2012, 94, 1279–1287. [Google Scholar] [CrossRef] [PubMed]
- EFSA Guidance on the characterisation of microorganisms used as feed additives or as production organisms. EFSA J. 2018, 16, e05206.
- Bover-Cid, S.; Holzapfel, W.H. Improved screening procedure for biogenic amine production by lactic acid bacteria. Int. J. Food Microbiol. 1999, 53, 33–41. [Google Scholar] [CrossRef]
- Zhou, J.S.; Gopal, P.K.; Hill, H.S. Potential probiotic lactic acid bacteria Lactobacillus rhamnosus (HN001), Lactobacillus acidophilus (HN017) and Bifidobacterium lactis (HN019) do not degrade gastric mucin in vitro. Int. J. Food Microbiol. 2001, 63, 81–90. [Google Scholar] [CrossRef]
- Anadón, A.; Martínez, M.A.; Ares, I.; Ramos, E.; Martínez-Larrañaga, M.R.; Contreras, M.M.; Ramos, M.; Recio, I. Acute and repeated dose (4 weeks) oral toxicity studies of two antihypertensive peptides, RYLGY and AYFYPEL, that correspond to fragments (90-94) and (143-149) from alpha(s1)-casein. Food Chem. Toxicol. 2010, 48, 1836–1845. [Google Scholar] [CrossRef]
- Lara-Villoslada, F.; Sierra, S.; Martín, R.; Delgado, S.; Rodríguez, J.M.; Olivares, M.; Xaus, J. Safety assessment of two probiotic strains, Lactobacillus coryniformis CECT5711 and Lactobacillus gasseri CECT5714. J. Appl. Microbiol. 2007, 103, 175–184. [Google Scholar] [CrossRef]
- Del Castillo, F.; Baquero, F.; de la Calle, T.; López, M.V.; Ruiz-Canela, J.; Alfayate, S.; Llop, F.M.; Ortega, M.J.C.; Rey, C.C. Documento de consenso sobre etiología, diagnóstico y tratamiento de la otitis media aguda. Rev. Pediatr. Aten. Primaria 2012, 14, 195–205. [Google Scholar] [CrossRef]
- Díez, O.; Batista, N.; Bordes, A.; Lecuona, M.; Lara, M. Microbiological diagnosis of upper respiratory tract infections. Enferm. Infecc. Microbiol. Clin. 2007, 25, 387–393. [Google Scholar] [CrossRef] [PubMed]
- The R Project for Statistical Computing. Available online: http: www.R-project.org (accessed on 29 July 2016).
- FAO; WHO. Guidelines for the Evaluation of Probiotics in Food; Report of a Joint FAO/WHO Working Group on Drafting Guidelines for the Evaluation of Probiotics in Food; Joint FAO/WHO Working Group: London, ON, Canada, 2002. [Google Scholar]
- Martín, R.; Jiménez, E.; Olivares, M.; Marín, M.; Fernández, L.; Xaus, J. Lactobacillus salivarius CECT 5713, a potential probiotic strain isolated from infant feces and breast milk of a mother-child pair. Int. J. Food Microbiol. 2006, 112, 35–43. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.K.; Puong, K.Y.; Ouwehand, A.C.; Salminen, S. Displacement of bacterial pathogens from mucus and Caco-2 cell surface by lactobacilli. J. Med. Microbiol. 2003, 52, 925–930. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bresnick, S.D.; Ryan, A.F. Modulation of middle ear immune response by gut immunization. Laryngoscope 1989, 99, 1293–1298. [Google Scholar] [CrossRef] [PubMed]
- Claesson, M.J.; Li, Y.; Leahy, S.; Canchaya, C.; van Pijkeren, J.P.; Cerdeño-Tárraga, A.M.; Parkhill, J.; Flynn, S.; O’Sullivan, G.C.; Collins, J.K.; et al. Multireplicon genome architecture of Lactobacillus salivarius. Proc. Natl. Acad. Sci. USA 2006, 103, 6718–6723. [Google Scholar] [CrossRef] [PubMed]
- Corr, S.C.; Li, Y.; Riedel, C.U.; O’Toole, P.W.; Hill, C.; Gahan, C.G. Bacteriocin production as a mechanism for the antiinfective activity of Lactobacillus salivarius UCC118. Proc. Natl. Acad. Sci. USA 2007, 104, 7617–7621. [Google Scholar] [CrossRef] [PubMed]
- Younes, J.A.; Reid, G.; van der Mei, H.C.; Busscher, H.J. Lactobacilli require physical contact to reduce staphylococcal TSST-1 secretion and vaginal epithelial inflammatory response. Pathog. Dis. 2016, 74, ftw029. [Google Scholar] [CrossRef]
- Rosas-Salazar, C.; Shilts, M.H.; Tovchigrechko, A.; Schobel, S.; Chappell, J.D.; Larkin, E.K.; Gebretsadik, T.; Halpin, R.A.; Nelson, K.E.; Moore, M.L.; et al. Nasopharyngeal Lactobacillus is associated with a reduced risk of childhood wheezing illnesses following acute respiratory syncytial virus infection in infancy. J. Allergy Clin. Immunol. 2018, 142, 1447–1456. [Google Scholar] [CrossRef]
- EFSA. Scientific opinion on the update of the list of QPS-recommended biological agents intentionally added to food or feed as notified to EFSA. EFSA J. 2017, 15, 4664. [Google Scholar]
- Danielsen, M.; Wind, A. Susceptibility of Lactobacillus spp. to antimicrobial agents. Int. J. Food Microbiol. 2003, 26, 1–11. [Google Scholar] [CrossRef]
- Zhou, J.S.; Pillidge, C.J.; Gopal, P.K.; Gill, H.S. Antibiotic susceptibility profiles of new probiotic Lactobacillus and Bifidobacterium strains. Int. J. Food Microbiol. 2005, 98, 211–217. [Google Scholar] [CrossRef] [PubMed]
- Klare, I.; Konstabel, C.; Werner, G.; Huys, G.; Vankerckhoven, V.; Kahlmeter, G.; Hildebrandt, B.; Müller-Bertling, S.; Witte, W.; Goossens, H. Antimicrobial susceptibilities of Lactobacillus; Pediococcus and Lactococcus human isolates and cultures intended for probiotic or nutritional use. J. Antimicrob. Chemother. 2007, 59, 900–912. [Google Scholar] [CrossRef] [PubMed]
- Tapiovaara, L.; Lehtoranta, L.; Swanljung, E.; Mäkivuokko, H.; Laakso, S.; Roivainen, M.; Korpela, R.; Pitkäranta, A. Lactobacillus rhamnosus GG in the middle ear after randomized, double-blind, placebo-controlled oral administration. Int. J. Pediatr. Otorhinolaryngol. 2014, 78, 1637–1641. [Google Scholar] [CrossRef] [PubMed]
- Hatakka, K.; Blomgren, K.; Pohjavuori, S.; Kaijalainen, T.; Poussa, T.; Leinonen, M.; Korpela, R.; Pitkäranta, A. Treatment of acute otitis media with probiotics in otitis-prone children-a double-blind, placebo-controlled randomised study. Clin. Nutr. 2007, 26, 314–321. [Google Scholar] [CrossRef] [PubMed]
- Marchisio, P.; Santagati, M.; Scillato, M.; Baggi, E.; Fattizzo, M.; Rosazza, C. Streptococcus salivarius 24SMB administered by nasal spray for the prevention of acute otitis media in otitis-prone children. Eur. J. Clin. Microbiol. Infect. Dis. 2015, 34, 2377–2383. [Google Scholar] [CrossRef] [PubMed]
- La Mantia, I.; Varricchio, A.; Ciprandi, G. Bacteriotherapy with Streptococcus salivarius 24SMB and Streptococcus oralis 89a nasal spray for preventing recurrent acute otitis media in children: A real-life clinical experience. Int. J. Gen. Med. 2017, 10, 171–175. [Google Scholar] [CrossRef] [PubMed]
- Chan, C.L.; Wabnitz, D.; Bassiouni, A.; Wormald, P.J.; Vreugde, S.; Psaltis, A.J. Identification of the bacterial reservoirs for the middle ear using phylogenic analysis. JAMA Otolaryngol. Head Neck Surg. 2017, 143, 155–161. [Google Scholar] [CrossRef] [PubMed]
- Torretta, S.; Marchisio, P.; Drago, L.; Baggi, E.; De Vecchi, E.; Garavello, W.; Nazzari, E.; Pignataro, L.; Esposito, S. Nasopharyngeal biofilm-producing otopathogens in children with nonsevere recurrent acute otitis media. Otolaryngol. Head Neck Surg. 2012, 146, 991–996. [Google Scholar] [CrossRef] [PubMed]
- Buzatto, G.P.; Tamashiro, E.; Proenca-Modena, J.L.; Saturno, T.H.; Prates, M.C.; Gagliardi, T.B.; Carenzi, L.R.; Massuda, E.T.; Hyppolito, M.A.; Valera, F.C.P.; et al. The pathogens profile in children with otitis media with effusion and adenoid hypertrophy. PLoS ONE 2017, 12, e0171049. [Google Scholar] [CrossRef] [PubMed]
- Cohen, R.; Martin, E.; de La Rocque, F.; Thollot, F.; Pecquet, S.; Werner, A.; Boucherat, M.; Varon, E.; Bingen, E.; Levy, C. Probiotics and prebiotics in preventing episodes of acute otitis media in high-risk children: A randomized, double-blind, placebo-controlled study. Pediatr. Infect. Dis. J. 2013, 32, 810–814. [Google Scholar] [CrossRef] [PubMed]
- Popova, M.; Molimard, P.; Courau, S.; Crociani, J.; Dufour, C.; Le Vacon, F.; Carton, T. Beneficial effects of probiotics in upperrespiratory tract infections and their mechanical actions to antagonize pathogens. J. Appl. Microbiol. 2012, 113, 1305–1318. [Google Scholar] [CrossRef] [PubMed]
- Levi, J.R.; Brody, R.M.; McKee-Cole, K.; Pribitkin, E.; O’Reilly, R. Complementary and alternative medicine for pediatric otitis media. Int. J. Pediatr. Otorhinolaryngol. 2013, 77, 926–931. [Google Scholar] [CrossRef] [PubMed]
- Tano, K.; Grahn Håkansson, E.; Holm, S.E.; Hellström, S. A nasal spray with alpha haemolytic streptococci as long term prophylaxis against recurrent otitis media. Int. J. Pediatr. Otorhinolaryngol. 2002, 62, 17–23. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Indicator Strain | Overlay Method (cm) | Well Diffusion Assay (cm) | Co-Cultures (CFU/mL) | ||
|---|---|---|---|---|---|
| Non-Adjusted pH | pH 6.2 | Initial Load | Final Load | ||
| S. pneumoniae MP07 | 3.6 | 1.1 | 1.1 | 7.40 | Nd a |
| S. pyogenes MP03 | 2.0 | 1.2 | 0.7 | 7.74 | Nd a |
| S. aureus MP29 | 1.6 | 1.3 | 1.2 | 7.52 | 4.81 a |
| S. epidermidis MP33 | 2.3 | 1.1 | 1.0 | 7.53 | 5.90 a |
| A. otitidis MP02 | 1.0 | 1.2 | 1.2 | – | – |
| E. faecalis MP64 | 0.5 | 0.4 | 0.4 | 8.02 | 8.78 b |
| H. influenzae MP04 | 2.2 | – | – | – | – |
| M. catarrhalis MP08 | 2.1 | – | – | – | – |
| P. aeruginosa MP24 | 1.5 | – | – | – | – |
| E. coli MP69 | 1.4 | – | – | 7.70 | 5.74 a |
| pH | L–Lactic Acid | D-Lactic Acid | Acetic Acid |
|---|---|---|---|
| 4.01± 0.04 | 10.29 ± 0.70 | Nd | 0.68 ± 0.17 |
| Antibiotics | Cut-Off Values * | MIC (L. salivarius PS7) |
|---|---|---|
| Ampicillin | 4 | 0.5 |
| Clindamycin | 4 | 0.5 |
| Chloramphenicol | 4 | 2 |
| Erythromycin | 1 | 0.25 |
| Streptomycin | 64 | 32 |
| Gentamicin | 16 | 2 |
| Kanamycin | 64 | 128 |
| Tetracyclin | 8 | 2 |
| Vancomycin | n.r. | >128 |
| Linezolid | 2 | 1 |
| Penicillin | 1 | 0.25 |
| Characteristic | Sex | n | Mean ± SD or Median * | p-Value |
|---|---|---|---|---|
| Age (Years) and Gender | 3.31 ± 1.7 | |||
| <3 years (n = 30) | Males | 16 | ||
| Females | 14 | |||
| ≥3 years (n = 31) | Males | 15 | ||
| Females | 16 | |||
| Inclusion months: | ||||
| September | 7 | |||
| October | 5 | |||
| November | 6 | |||
| December | 7 | |||
| January | 6 | |||
| February | 10 | |||
| March | 9 | |||
| April | 6 | |||
| May | 3 | |||
| June | 2 | |||
| History of AOM episodes/child | ||||
| Preceding 6 months | 4 (3–4) * | <0.001 ¥ | ||
| Preceding 12 months | 5 (5–6) * | <0.001 ¥ | ||
| During the study | 0 (0–1) * |
| Time 0 | After 6 Months | p-Value | |
|---|---|---|---|
| Number of positive samples (bacterial growth) | 61 | 17 | < 0.001 ¥ |
| Median log10 CFU (per swab) | 4 (3.5–5) | 2 (1.5–2) | 0.012 * |
| Number of samples with the presence of: | |||
| Actinomyces europaeus | 2 | 0 | |
| Alloiococcus otitidis | 12 | 2 | |
| Coagulase-negative staphylococci | 14 | 15 | |
| Haemophilus haemolyticus | 11 | 1 | |
| Haemophilus influenza | 7 | 1 | |
| Moraxella catharralis | 14 | 3 | |
| Neisseria spp. | 6 | 0 | |
| Pseudomonas aeruginosa | 4 | 0 | |
| Rhodococcus ruber | 2 | 0 | |
| Staphylococcus aureus | 11 | 3 | |
| Streptococcus pneumoniae | 9 | 3 | |
| Streptococcus pyogenes | 10 | 6 | |
| Group viridans streptococci | 9 | 11 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cárdenas, N.; Martín, V.; Arroyo, R.; López, M.; Carrera, M.; Badiola, C.; Jiménez, E.; Rodríguez, J.M. Prevention of Recurrent Acute Otitis Media in Children Through the Use of Lactobacillus salivarius PS7, a Target-Specific Probiotic Strain. Nutrients 2019, 11, 376. https://doi.org/10.3390/nu11020376
Cárdenas N, Martín V, Arroyo R, López M, Carrera M, Badiola C, Jiménez E, Rodríguez JM. Prevention of Recurrent Acute Otitis Media in Children Through the Use of Lactobacillus salivarius PS7, a Target-Specific Probiotic Strain. Nutrients. 2019; 11(2):376. https://doi.org/10.3390/nu11020376
Chicago/Turabian StyleCárdenas, Nivia, Virginia Martín, Rebeca Arroyo, Mario López, Marta Carrera, Carlos Badiola, Esther Jiménez, and Juan M. Rodríguez. 2019. "Prevention of Recurrent Acute Otitis Media in Children Through the Use of Lactobacillus salivarius PS7, a Target-Specific Probiotic Strain" Nutrients 11, no. 2: 376. https://doi.org/10.3390/nu11020376