Differential Modulation of Cancellous and Cortical Distal Femur by Fructose and Natural Mineral-Rich Water Consumption in Ovariectomized Female Sprague Dawley Rats

 ,
,  , ,
, ,
Abstract
1. Introduction
2. Material and Methods
2.1. In Vivo Animal Experimentation and Treatments
2.2. Assessment of Bone Microstructural Properties and Volumetric Bone Mineral Density by Microcomputed Tomography
2.2.1. Scan, Reconstruction, and Image Processing
2.2.2. Microstructural Parameters of Cancellous Bone
2.2.3. Microstructural Parameters of Cortical Bone
2.2.4. Volumetric Bone Mineral Density
2.3. Assessment of Circulating Markers of Bone Metabolism
2.4. Statistical Analysis
3. Results
3.1. Microstructural Properties and Volumetric Bone Mineral Density by Micro-CT
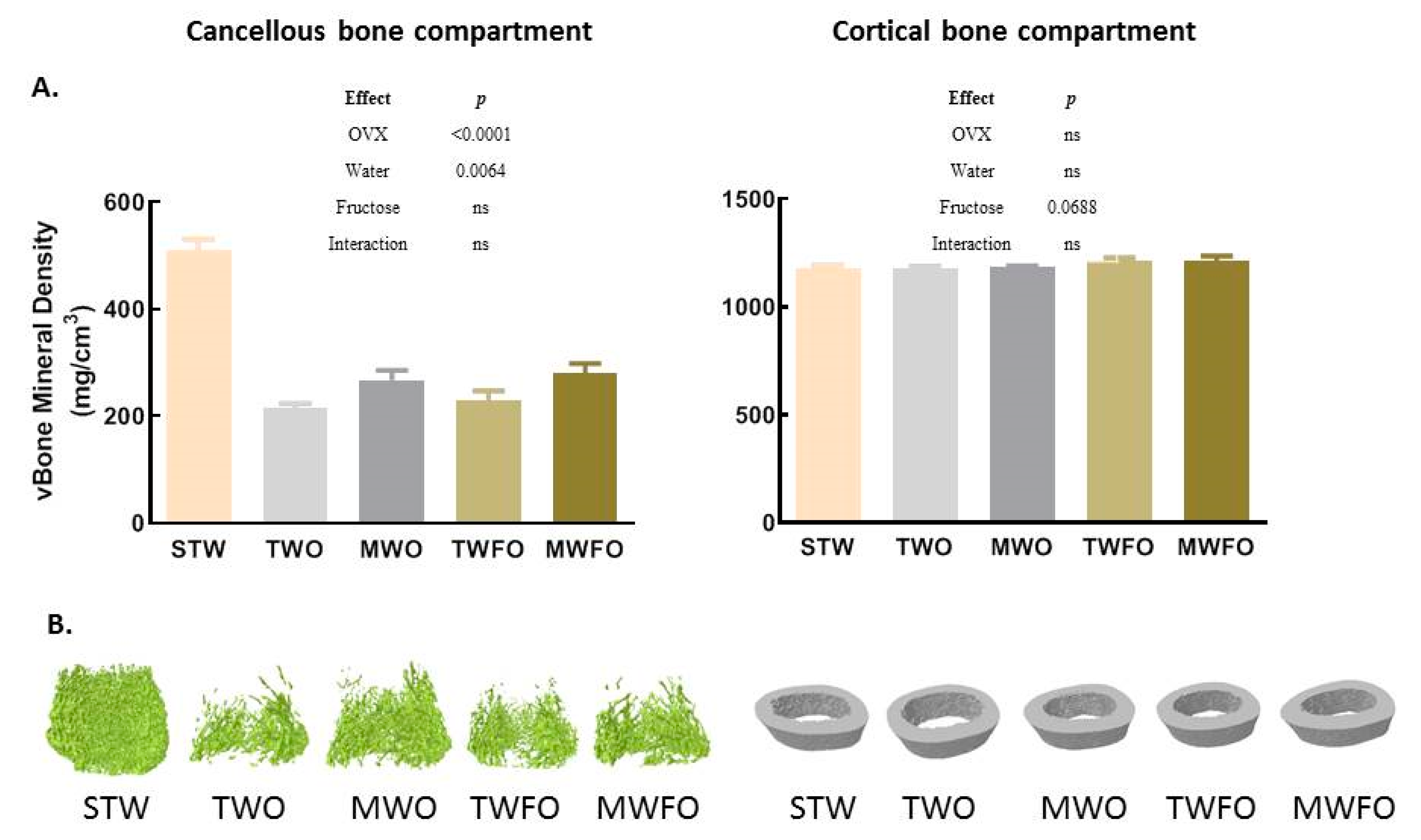
3.1.1. Cancellous Bone Compartment
3.1.2. Cortical Bone Compartment
3.2. Circulating Markers of Bone Metabolism
4. Discussion
4.1. Ovariectomy
4.2. Fructose
4.3. Natural Mineral-Rich Water
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Osterhoff, G.; Morgan, E.F.; Shefelbine, S.J.; Karim, L.; McNamara, L.M.; Augat, P. Bone mechanical properties and changes with osteoporosis. Injury 2016, 47 (Suppl. S2), S11–S20. [Google Scholar] [CrossRef]
- Vaananen, H.K.; Harkonen, P.L. Estrogen and bone metabolism. Maturitas 1996, 23, S65–S69. [Google Scholar] [CrossRef]
- Zebaze, R.M.; Ghasem-Zadeh, A.; Bohte, A.; Iuliano-Burns, S.; Mirams, M.; Price, R.I.; Mackie, E.J.; Seeman, E. Intracortical remodelling and porosity in the distal radius and post-mortem femurs of women: A cross-sectional study. Lancet 2010, 375, 1729–1736. [Google Scholar] [CrossRef]
- Cheung, A.M.; Frame, H.; Ho, M.; Mackinnon, E.S.; Brown, J.P. Bone strength and management of postmenopausal fracture risk with antiresorptive therapies: Considerations for women’s health practice. Int. J. Womens Health 2016, 8, 537–547. [Google Scholar] [PubMed]
- Iolascon, G.; Napolano, R.; Gioia, M.; Moretti, A.; Riccio, I.; Gimigliano, F. The contribution of cortical and trabecular tissues to bone strength: Insights from denosumab studies. Clin. Cases Miner. Bone Metab. 2013, 10, 47–51. [Google Scholar] [CrossRef] [PubMed]
- Levin, V.A.; Jiang, X.; Kagan, R. Estrogen therapy for osteoporosis in the modern era. Osteoporos. Int. 2018, 29, 1049–1055. [Google Scholar] [CrossRef]
- Liu, W.; Zhang, X. Receptor activator of nuclear factor-kappaB ligand (RANKL)/RANK/osteoprotegerin system in bone and other tissues (review). Mol. Med. Rep. 2015, 11, 3212–3218. [Google Scholar] [CrossRef]
- Kalaiselvi, V.; Prabhu, K.; Ramesh, M.; Venkatesan, V. The association of serum osteocalcin with the bone mineral density in post menopausal women. J. Clin. Diagn. Res. 2013, 7, 814–816. [Google Scholar]
- Wynn, E.; Krieg, M.A.; Lanham-New, S.A.; Burckhardt, P. Postgraduate Symposium: Positive influence of nutritional alkalinity on bone health. Proc. Nutr. Soc. 2010, 69, 166–173. [Google Scholar] [CrossRef]
- Gambacciani, M.; Levancini, M. Hormone replacement therapy and the prevention of postmenopausal osteoporosis. Prz. Menopauzalny 2014, 13, 213–220. [Google Scholar]
- Komori, T. Animal models for osteoporosis. Eur. J. Pharmacol. 2015, 759, 287–294. [Google Scholar] [CrossRef] [PubMed]
- Migliaccio, S.; Marocco, C.; Mocini, E.; Lenzi, A.; Greco, E. Role of Mediterranean diet in bone health. Clin. Cases Miner. Bone Metab. 2018, 15, 16–18. [Google Scholar]
- Buclin, T.; Cosma, M.; Appenzeller, M.; Jacquet, A.F.; Decosterd, L.A.; Biollaz, J.; Burckhardt, P. Diet acids and alkalis influence calcium retention in bone. Osteoporos. Int. 2001, 12, 493–499. [Google Scholar] [CrossRef] [PubMed]
- Kelly, O.J.; Gilman, J.C.; Kim, Y.; Ilich, J.Z. Micronutrient Intake in the Etiology, Prevention and Treatment of Osteosarcopenic Obesity. Curr. Aging Sci. 2016, 9, 260–278. [Google Scholar] [CrossRef]
- Cepollaro, C.; Orlandi, G.; Gonnelli, S.; Ferrucci, G.; Arditti, J.C.; Borracelli, D.; Toti, E.; Gennari, C. Effect of calcium supplementation as a high-calcium mineral water on bone loss in early postmenopausal women. Calcif. Tissue Int. 1996, 59, 238–239. [Google Scholar] [CrossRef] [PubMed]
- Costi, D.; Calcaterra, P.G.; Iori, N.; Vourna, S.; Nappi, G.; Passeri, M. Importance of bioavailable calcium drinking water for the maintenance of bone mass in post-menopausal women. J. Endocrinol. Investig. 1999, 22, 852–856. [Google Scholar] [CrossRef]
- Guillemant, J.; Le, H.T.; Accarie, C.; du Montcel, S.T.; Delabroise, A.M.; Arnaud, M.J.; Guillemant, S. Mineral water as a source of dietary calcium: Acute effects on parathyroid function and bone resorption in young men. Am. J. Clin. Nutr. 2000, 71, 999–1002. [Google Scholar] [CrossRef]
- Roux, S.; Baudoin, C.; Boute, D.; Brazier, M.; De La Gueronniere, V.; De Vernejoul, M.C. Biological effects of drinking-water mineral composition on calcium balance and bone remodeling markers. J. Nutr. Health Aging 2004, 8, 380–384. [Google Scholar]
- Meunier, P.J.; Jenvrin, C.; Munoz, F.; de la Gueronniere, V.; Garnero, P.; Menz, M. Consumption of a high calcium mineral water lowers biochemical indices of bone remodeling in postmenopausal women with low calcium intake. Osteoporos. Int. 2005, 16, 1203–1209. [Google Scholar] [CrossRef]
- Wynn, E.; Krieg, M.A.; Aeschlimann, J.M.; Burckhardt, P. Alkaline mineral water lowers bone resorption even in calcium sufficiency: Alkaline mineral water and bone metabolism. Bone 2009, 44, 120–124. [Google Scholar] [CrossRef]
- Burckhardt, P. The effect of the alkali load of mineral water on bone metabolism: Interventional studies. J. Nutr. 2008, 138, 435S–437S. [Google Scholar] [CrossRef] [PubMed]
- Ogata, F.; Nagai, N.; Ito, Y.; Kawasaki, N. Fundamental study on effect of high-mineral drinking water for osteogenesis in calciprivia ovariectomized rats. Yakugaku Zasshi J. Pharm. Soc. Jpn. 2014, 134, 679–685. [Google Scholar] [CrossRef] [PubMed]
- Dahl, C.; Sogaard, A.J.; Tell, G.S.; Forsen, L.; Flaten, T.P.; Hongve, D.; Omsland, T.K.; Holvik, K.; Meyer, H.E.; Aamodt, G. Population data on calcium in drinking water and hip fracture: An association may depend on other minerals in water. A NOREPOS study. Bone 2015, 81, 292–299. [Google Scholar] [PubMed]
- Vannucci, L.; Fossi, C.; Quattrini, S.; Guasti, L.; Pampaloni, B.; Gronchi, G.; Giusti, F.; Romagnoli, C.; Cianferotti, L.; Marcucci, G.; et al. Calcium Intake in Bone Health: A Focus on Calcium-Rich Mineral Waters. Nutrients 2018, 10, 1930. [Google Scholar] [CrossRef] [PubMed]
- Qiu, Z.; Tan, Y.; Zeng, H.; Wang, L.; Wang, D.; Luo, J.; Zhang, L.; Huang, Y.; Chen, J.A.; Shu, W. Multi-generational drinking of bottled low mineral water impairs bone quality in female rats. PLoS ONE 2015, 10, e0121995. [Google Scholar] [CrossRef]
- Huang, Y.; Ma, X.; Tan, Y.; Wang, L.; Wang, J.; Lan, L.; Qiu, Z.; Luo, J.; Zeng, H.; Shu, W. Consumption of Very Low Mineral Water Is Associated with Lower Bone Mineral Content in Children. J. Nutr. 2019. [Google Scholar] [CrossRef] [PubMed]
- Bagi, C.M.; Edwards, K.; Berryman, E. Metabolic Syndrome and Bone: Pharmacologically Induced Diabetes has Deleterious Effect on Bone in Growing Obese Rats. Calcif. Tissue Int. 2018, 102, 683–694. [Google Scholar] [CrossRef]
- Das, J.K.; Severo, M.; Pereira, C.D.; Patricio, E.; Magalhaes, J.; Monteiro, R.; Neves, D.; Martins, M.J. Natural mineral-rich water ingestion by ovariectomized fructose-fed Sprague-Dawley rats: Effects on sirtuin 1 and glucocorticoid signaling pathways. Menopause 2017, 24, 563–573. [Google Scholar] [CrossRef]
- Dornas, W.C.; de Lima, W.G.; Pedrosa, M.L.; Silva, M.E. Health implications of high-fructose intake and current research. Adv. Nutr. 2015, 6, 729–737. [Google Scholar] [CrossRef]
- Zhang, D.M.; Jiao, R.Q.; Kong, L.D. High Dietary Fructose: Direct or Indirect Dangerous Factors Disturbing Tissue and Organ Functions. Nutrients 2017, 9, 335. [Google Scholar] [CrossRef]
- Hanayama, R.; Shimizu, H.; Nakagami, H.; Osako, M.K.; Makino, H.; Kunugiza, Y.; Tomita, T.; Tsukamoto, I.; Yoshikawa, H.; Rakugi, H.; et al. Fluvastatin improves osteoporosis in fructose-fed insulin resistant model rats through blockade of the classical mevalonate pathway and antioxidant action. Int. J. Mol. Med. 2009, 23, 581–588. [Google Scholar] [PubMed]
- Koh, E.T.; Bourdeau, J.E.; Tae, W.C. A magnesium-deficient high fructose diet augments bone-sparing action of exogenous oestrogen in ovariectomized rats. Magnes. Res. 1993, 6, 333–342. [Google Scholar] [PubMed]
- Felice, J.I.; Gangoiti, M.V.; Molinuevo, M.S.; McCarthy, A.D.; Cortizo, A.M. Effects of a metabolic syndrome induced by a fructose-rich diet on bone metabolism in rats. Metabolism 2014, 63, 296–305. [Google Scholar] [CrossRef] [PubMed]
- Jatkar, A.; Kurland, I.J.; Judex, S. Diets High in Fat or Fructose Differentially Modulate Bone Health and Lipid Metabolism. Calcif. Tissue Int. 2017, 100, 20–28. [Google Scholar] [CrossRef] [PubMed]
- Nuche-Berenguer, B.; Moreno, P.; Esbrit, P.; Dapia, S.; Caeiro, J.R.; Cancelas, J.; Haro-Mora, J.J.; Villanueva-Penacarrillo, M.L. Effect of GLP-1 treatment on bone turnover in normal, type 2 diabetic, and insulin-resistant states. Calcif. Tissue Int. 2009, 84, 453–461. [Google Scholar] [CrossRef] [PubMed]
- Yarrow, J.F.; Toklu, H.Z.; Balaez, A.; Phillips, E.G.; Otzel, D.M.; Chen, C.; Wronski, T.J.; Aguirre, J.I.; Sakarya, Y.; Tumer, N.; et al. Fructose consumption does not worsen bone deficits resulting from high-fat feeding in young male rats. Bone 2016, 85, 99–106. [Google Scholar] [CrossRef] [PubMed]
- Tian, L.; Yu, X. Fat, Sugar, and Bone Health: A Complex Relationship. Nutrients 2017, 9, 506. [Google Scholar] [CrossRef]
- Tian, L.; Wang, C.; Xie, Y.; Wan, S.; Zhang, K.; Yu, X. High Fructose and High Fat Exert Different Effects on Changes in Trabecular Bone Micro-structure. J. Nutr. Health Aging 2018, 22, 361–370. [Google Scholar] [CrossRef] [PubMed]
- Campos, V.C.; Tappy, L. Physiological handling of dietary fructose-containing sugars: Implications for health. Int. J. Obes. (Lond.) 2016, 40 (Suppl. S1), S6–S11. [Google Scholar] [CrossRef]
- Pereira, C.D.; Passos, E.; Severo, M.; Vito, I.; Wen, X.; Carneiro, F.; Gomes, P.; Monteiro, R.; Martins, M.J. Ingestion of a natural mineral-rich water in an animal model of metabolic syndrome: Effects in insulin signalling and endoplasmic reticulum stress. Horm. Mol. Biol. Clin. Investig. 2016, 26, 135–150. [Google Scholar] [CrossRef]
- Pereira, C.D.; Severo, M.; Araujo, J.R.; Guimaraes, J.T.; Pestana, D.; Santos, A.; Ferreira, R.; Ascensao, A.; Magalhaes, J.; Azevedo, I.; et al. Relevance of a Hypersaline Sodium-Rich Naturally Sparkling Mineral Water to the Protection against Metabolic Syndrome Induction in Fructose-Fed Sprague-Dawley Rats: A Biochemical, Metabolic, and Redox Approach. Int. J. Endocrinol. 2014, 2014, 384583. [Google Scholar] [CrossRef] [PubMed]
- Pereira, C.D.; Severo, M.; Neves, D.; Ascensao, A.; Magalhaes, J.; Guimaraes, J.T.; Monteiro, R.; Martins, M.J. Natural mineral-rich water ingestion improves hepatic and fat glucocorticoid-signaling and increases sirtuin 1 in an animal model of metabolic syndrome. Horm. Mol. Biol. Clin. Investig. 2015, 21, 149–157. [Google Scholar] [CrossRef] [PubMed]
- Pereira, C.D.; Severo, M.; Rafael, L.; Martins, M.J.; Neves, D. Effects of natural mineral-rich water consumption on the expression of sirtuin 1 and angiogenic factors in the erectile tissue of rats with fructose-induced metabolic syndrome. Asian J. Androl. 2014, 16, 631–638. [Google Scholar] [CrossRef] [PubMed]
- Pereira, C.D.; Monteiro, R.; Martins, M.J. Further insights into the metabolically healthy obese phenotype: The role of magnesium. Eur. J. Intern. Med. 2014, 25, e105–e106. [Google Scholar] [CrossRef] [PubMed]
- Pereira, C.D.; Collado, M.C.; Passos, E.; Azevedo, I.; Monteiro, R.; Martins, M.J. Comment to: Luo et al. (2013). Int. J. Cardiol. 2014, 172, 512–514. [Google Scholar] [CrossRef] [PubMed]
- Costa-Vieira, D.; Monteiro, R.; Martins, M.J. Metabolic Syndrome Features: Is There a Modulation Role by Mineral Water Consumption? A Review. Nutrients 2019, 11, 1141. [Google Scholar] [CrossRef]
- Wronski, T.J.; Cintron, M.; Dann, L.M. Temporal relationship between bone loss and increased bone turnover in ovariectomized rats. Calcif. Tissue Int. 1988, 43, 179–183. [Google Scholar] [CrossRef]
- Hunt, J.R.; Hunt, C.D.; Zito, C.A.; Idso, J.P.; Johnson, L.K. Calcium requirements of growing rats based on bone mass, structure, or biomechanical strength are similar. J. Nutr. 2008, 138, 1462–1468. [Google Scholar] [CrossRef]
- Martin-Fernandez, M.; Garzon-Marquez, F.M.; Diaz-Curiel, M.; Prieto-Potin, I.; Alvarez-Galovich, L.; Guede, D.; Caeiro-Rey, J.R.; De la Piedra, C. Comparative study of the effects of osteoprotegerin and testosterone on bone quality in male orchidectomised rats. Aging Male 2018, 1–13. [Google Scholar] [CrossRef]
- Bouxsein, M.L.; Boyd, S.K.; Christiansen, B.A.; Guldberg, R.E.; Jepsen, K.J.; Muller, R. Guidelines for assessment of bone microstructure in rodents using micro-computed tomography. J. Bone Miner. Res. 2010, 25, 1468–1486. [Google Scholar] [CrossRef]
- Cao, J.J.; Gregoire, B.R.; Gao, H. High-fat diet decreases cancellous bone mass but has no effect on cortical bone mass in the tibia in mice. Bone 2009, 44, 1097–1104. [Google Scholar] [CrossRef] [PubMed]
- Fujita, Y.; Watanabe, K.; Maki, K. Serum leptin levels negatively correlate with trabecular bone mineral density in high-fat diet-induced obesity mice. J. Musculoskelet Neuronal. Interact. 2012, 12, 84–94. [Google Scholar] [PubMed]
- Ma, C.H.; Chiua, Y.C.; Wu, C.H.; Jou, I.M.; Tu, Y.K.; Hung, C.H.; Hsieh, P.L.; Tsai, K.L. Homocysteine causes dysfunction of chondrocytes and oxidative stress through repression of SIRT1/AMPK pathway: A possible link between hyperhomocysteinemia and osteoarthritis. Redox. Biol. 2018, 15, 504–512. [Google Scholar] [CrossRef]
- Zainabadi, K.; Liu, C.J.; Caldwell, A.L.M.; Guarente, L. SIRT1 is a positive regulator of in vivo bone mass and a therapeutic target for osteoporosis. PLoS ONE 2017, 12, e0185236. [Google Scholar] [CrossRef] [PubMed]
- Martin, E.A.; Ritman, E.L.; Turner, R.T. Time course of epiphyseal growth plate fusion in rat tibiae. Bone 2003, 32, 261–267. [Google Scholar] [CrossRef]
- Galea, G.L.; Hannuna, S.; Meakin, L.B.; Delisser, P.J.; Lanyon, L.E.; Price, J.S. Quantification of Alterations in Cortical Bone Geometry Using Site Specificity Software in Mouse models of Aging and the Responses to Ovariectomy and Altered Loading. Front. Endocrinol. (Lausanne) 2015, 6, 52. [Google Scholar] [CrossRef]
- Wagner, D.; Fahrleitner-Pammer, A. Levels of osteoprotegerin (OPG) and receptor activator for nuclear factor kappa B ligand (RANKL) in serum: Are they of any help? Wien Med. Wochenschr. 2010, 160, 452–457. [Google Scholar] [CrossRef]
- Singh, S.; Kumar, D.; Lal, A.K. Serum Osteocalcin as a Diagnostic Biomarker for Primary Osteoporosis in Women. J. Clin. Diagn. Res. 2015, 9, RC04–RC07. [Google Scholar] [CrossRef]
- Sanchis-Gimeno, J.A.; Llido, S.; Guede, D.; Nalla, S.; Martinez-Soriano, F.; Blanco-Perez, E.; Caeiro, J.R. Atlases with Arcuate Foramen present cortical bone thickening that may contribute to a lower fracture risk. World Neurosurg. 2018, 117, e162–e166. [Google Scholar] [CrossRef]
- Tarantino, U.; Rao, C.; Tempesta, V.; Gasbarra, E.; Feola, M. Hip fractures in the elderly: The role of cortical bone. Injury 2016, 47 (Suppl. S4), S107–S111. [Google Scholar] [CrossRef]
- Vranken, L.; Wyers, C.E.; van Rietbergen, B.; Driessen, J.H.M.; Geusens, P.; Janzing, H.M.J.; van der Velde, R.Y.; van den Bergh, J.P.W. The association between prevalent vertebral fractures and bone quality of the distal radius and distal tibia as measured with HR-pQCT in postmenopausal women with a recent non-vertebral fracture at the Fracture Liaison Service. Osteoporos. Int. 2019, 30, 1789–1797. [Google Scholar] [CrossRef] [PubMed]
- Ohlsson, C.; Sundh, D.; Wallerek, A.; Nilsson, M.; Karlsson, M.; Johansson, H.; Mellstrom, D.; Lorentzon, M. Cortical Bone Area Predicts Incident Fractures Independently of Areal Bone Mineral Density in Older Men. J. Clin. Endocrinol. Metab. 2017, 102, 516–524. [Google Scholar] [PubMed]
- Stacy, B.D.; Wilson, B.W. Acidosis and hypercalciuria: Renal mechanisms affecting calcium, magnesium and sodium excretion in the sheep. J. Physiol. 1970, 210, 549–564. [Google Scholar] [CrossRef]
- Levin, M.; Winaver, J. Effects of systemic alkalosis on urinary magnesium excretion in the rat. Miner. Electrolyte Metab. 1989, 15, 214–220. [Google Scholar] [PubMed]


{kind=link}
| STW | TWO | MWO | TWFO | MWFO | p OVX Effect | p Water Effect | p Fructose Effect | p Interaction | |
|---|---|---|---|---|---|---|---|---|---|
| Bone volumetric fraction (BV/TV, %) | 30.63 ± 6.69 | 6.23 ± 0.37 | 11.33 ± 2.83 | 7.07 ± 1.41 | 10.65 ± 1.50 | <0.0001 | <0.0001 (TWO vs. MWO 0.0009; TWFO vs. MWFO 0.0138) | ns | ns |
| Specific bone surface (BS/BV, mm−1) | 34.53 ± 3.81 | 41.39 ± 4.61 | 36.70 ± 3.02 | 39.30 ± 4.37 | 37.25 ± 1.36 | 0.0185 | 0.0405 | ns | ns |
| Bone surface density (BS/TV, mm−1) | 10.37 ± 1.30 | 2.62 ± 0.19 | 4.09 ± 0.73 | 2.85 ± 0.75 | 3.89 ± 0.63 | <0.0001 | 0.0001 (TWO vs. MWO 0.0055; TWFO vs. MWFO 0.0465) | ns | ns |
| Trabecular thickness (Tb.Th, µm) | 110.91 ± 6.61 | 99.82 ± 7.78 | 107.77 ± 7.46 | 100.13 ± 5.62 | 106.37 ± 2.36 | 0.0239 | 0.0181 | ns | ns |
| Trabecular spacing (Tb.Sp, µm) | 213.32 ± 29.88 | 972.04 ± 173.12 | 807.39 ± 170.42 | 1046.67 ± 247.39 | 838.22 ± 64.15 | <0.0001 | 0.0230 | ns | ns |
| Trabecular number (Tb.N, mm−1) | 2.74 ± 0.46 | 0.63 ± 0.03 | 1.04 ± 0.19 | 0.70 ± 0.18 | 0.98 ± 0.16 | <0.0001 | <0.0001 (TWO vs. MWO 0.0024; TWFO vs. MWFO 0.0298) | ns | ns |
| Trabecular bone pattern factor (Tb.Pf, mm−1) | 6.05 ± 3.94 | 15.73 ± 2.74 | 12.81 ± 1.07 | 14.60 ± 1.94 | 12.85 ± 0.86 | 0.0006 | 0.0069 (TWO vs. MWO 0.0735) | ns | ns |
| Structural model índex (SMI) | 1.43 ± 0.44 | 2.38 ± 0.15 | 2.13 ± 0.13 | 2.31 ± 0.07 | 2.20 ± 0.12 | 0.0005 | 0.0016 (TWO vs. MWO 0.0101) | ns | ns |
| Degree of anisotropy (DA) | 0.34 ± 0.14 | 0.50 ± 0.14 | 0.43 ± 0.04 | 0.45 ± 0.17 | 0.46 ± 0.11 | 0.0656 | ns | ns | ns |
| STW | TWO | MWO | TWFO | MWFO | p OVX Effect | p Water Effect | p Fructose Effect | p Interaction | |
|---|---|---|---|---|---|---|---|---|---|
| Cortical thickness (Ct.Th, µm) | 496.13 ± 43.84 | 457.67 ± 56.96 | 489.40 ± 21.65 | 539.79 ± 61.06 | 535.48 ± 24.52 | ns | ns | 0.0049 (TWO vs. TWFO 0.0318) | ns |
| Cortical volume (Ct.V, mm3) | 10.51 ± 0.77 | 10.92 ± 0.57 | 11.20 ± 1.15 | 11.24 ± 1.02 | 11.29 ± 0.40 | ns | ns | ns | ns |
| Medullary volume (MV, mm3) | 7.76 ± 1.54 | 9.32 ± 1.44 | 6.93 ± 0.63 | 7.12 ± 1.52 | 6.24 ± 0.68 | ns | 0.0033 (TWO vs. MWO 0.0150) | 0.0080 (TWO vs. TWFO 0.0195) | ns |
| Cross-sectional thickness (Cs.Th, µm) | 440.96 ± 62.50 | 404.95 ± 59.33 | 455.86 ± 42.76 | 502.25 ± 53.71 | 497.16 ± 17.56 | ns | ns | 0.0024 (TWO vs. TWFO 0.0101) | ns |
| Mean total cross-sectional bone area (B.Ar, mm2) | 5.35 ± 0.48 | 5.46 ± 0.48 | 5.45 ± 0.54 | 5.50 ± 0.48 | 5.53 ± 0.21 | ns | ns | ns | ns |
| Cortical periosteal perimeter (Ct.Pe.Pm, mm) | 11.27 ± 0.77 | 11.92 ± 0.44 | 11.25 ± 0.68 | 11.18 ± 0.73 | 10.96 ± 0.08 | ns | 0.0708 | 0.0399 | ns |
| Cortical endosteal perimeter (Ct.En.Pm, mm) | 7.25 ± 0.73 | 7.99 ± 0.59 | 7.14 ± 0.47 | 7.06 ± 0.76 | 6.62 ± 0.20 | 0.0799 | 0.0126 (TWO vs. MWO 0.0704) | 0.0058 (TWO vs. TWFO 0.0419) | ns |
| Mean polar moment of inertia (polar, MMI, mm4) | 11.97 ± 2.64 | 13.96 ± 1.93 | 12.08 ± 2.52 | 11.99 ± 2.82 | 11.12 ± 0.34 | ns | ns | ns | ns |
| Mean eccentricity (Ecc) | 0.75 ± 0.03 | 0.74 ± 0.02 | 0.73 ± 0.04 | 0.73 ± 0.02 | 0.77 ± 0.03 | ns | ns | ns (MWO vs. MWFO 0.0738) | 0.0346 |
| STW | TWO | MWO | TWFO | MWFO | p OVX Effect | p Water Effect | p Fructose Effect | p Interaction | |
|---|---|---|---|---|---|---|---|---|---|
| N-terminal propeptide of type I procollagen (ng/mL) | 6.87 ± 4.40 | 5.75 ± 3.88 | 4.24 ± 1.78 | 3.33 ± 2.87 | 3.61 ± 2.59 | ns | ns | ns | ns |
| Osteopontin (ng/mL) | 40.32 ± 7.02 | 41.98 ± 5.48 | 40.41 ± 1.87 | 35.90 ± 5.11 | 41.21 ± 2.72 | ns | ns | ns (TWO vs. TWFO 0.0959) | 0.0738 |
| Osteocalcin (ng/mL) | 156.73 ± 43.53 | 204.14 ± 30.74 | 169.30 ± 17.07 | 175.18 ± 3.76 | 172.50 ± 24.97 | 0.0541 | 0.0779 (TWO vs. MWO 0.0935) | ns | ns |
| CTX-1 (ng/mL) | 16.27 ± 4.47 | 24.23 ± 3.21 | 22.90 ± 6.76 | 22.09 ± 5.60 | 23.74 ± 0.60 | 0.0090 | ns | ns | ns |
| OPG (pg/mL) | 280.24 ± 54.54 | 208.72 ± 17.61 | 243.63 ± 18.16 | 286.51 ± 46.01 | 238.64 ± 9.56 | 0.0415 | ns (TWFO vs. MWFO 0.0954) | 0.0160 (TWO vs. TWFO 0.0049) | 0.0076 |
| RANKL (pg/mL) | 13.15 ± 0.79 | 17.04 ± 0.95 | 16.10 ± 3.32 | 22.15 ± 1.25 | 17.93 ± 6.33 | 0.0003 | ns | 0.0780 | ns |
| OPG/RANKL | 21.38 ± 4.29 | 12.40 ± 1.28 | 13.91 ± 3.87 | 11.82 ± 0.86 | 15.05 ± 4.94 | 0.0020 | ns | ns | ns |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pereira, C.; Guede, D.; Durães, C.; Brandão, I.; Silva, N.; Passos, E.; Bernardes, M.; Monteiro, R.; Martins, M.J. Differential Modulation of Cancellous and Cortical Distal Femur by Fructose and Natural Mineral-Rich Water Consumption in Ovariectomized Female Sprague Dawley Rats. Nutrients 2019, 11, 2316. https://doi.org/10.3390/nu11102316
Pereira C, Guede D, Durães C, Brandão I, Silva N, Passos E, Bernardes M, Monteiro R, Martins MJ. Differential Modulation of Cancellous and Cortical Distal Femur by Fructose and Natural Mineral-Rich Water Consumption in Ovariectomized Female Sprague Dawley Rats. Nutrients. 2019; 11(10):2316. https://doi.org/10.3390/nu11102316
Chicago/Turabian StylePereira, Cidália, David Guede, Cecília Durães, Inês Brandão, Nuno Silva, Emanuel Passos, Miguel Bernardes, Rosário Monteiro, and Maria João Martins. 2019. "Differential Modulation of Cancellous and Cortical Distal Femur by Fructose and Natural Mineral-Rich Water Consumption in Ovariectomized Female Sprague Dawley Rats" Nutrients 11, no. 10: 2316. https://doi.org/10.3390/nu11102316
APA StylePereira, C., Guede, D., Durães, C., Brandão, I., Silva, N., Passos, E., Bernardes, M., Monteiro, R., & Martins, M. J. (2019). Differential Modulation of Cancellous and Cortical Distal Femur by Fructose and Natural Mineral-Rich Water Consumption in Ovariectomized Female Sprague Dawley Rats. Nutrients, 11(10), 2316. https://doi.org/10.3390/nu11102316