Brain Fatty Acid Composition and Inflammation in Mice Fed with High-Carbohydrate Diet or High-Fat Diet

 ,
,
Abstract
:
1. Introduction
2. Materials and Methods
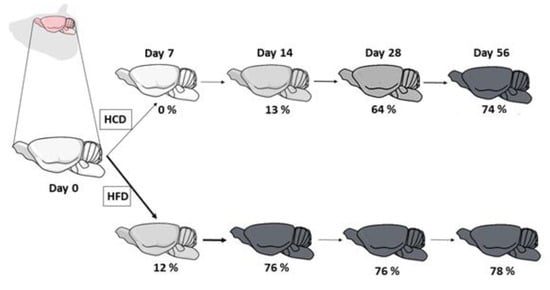
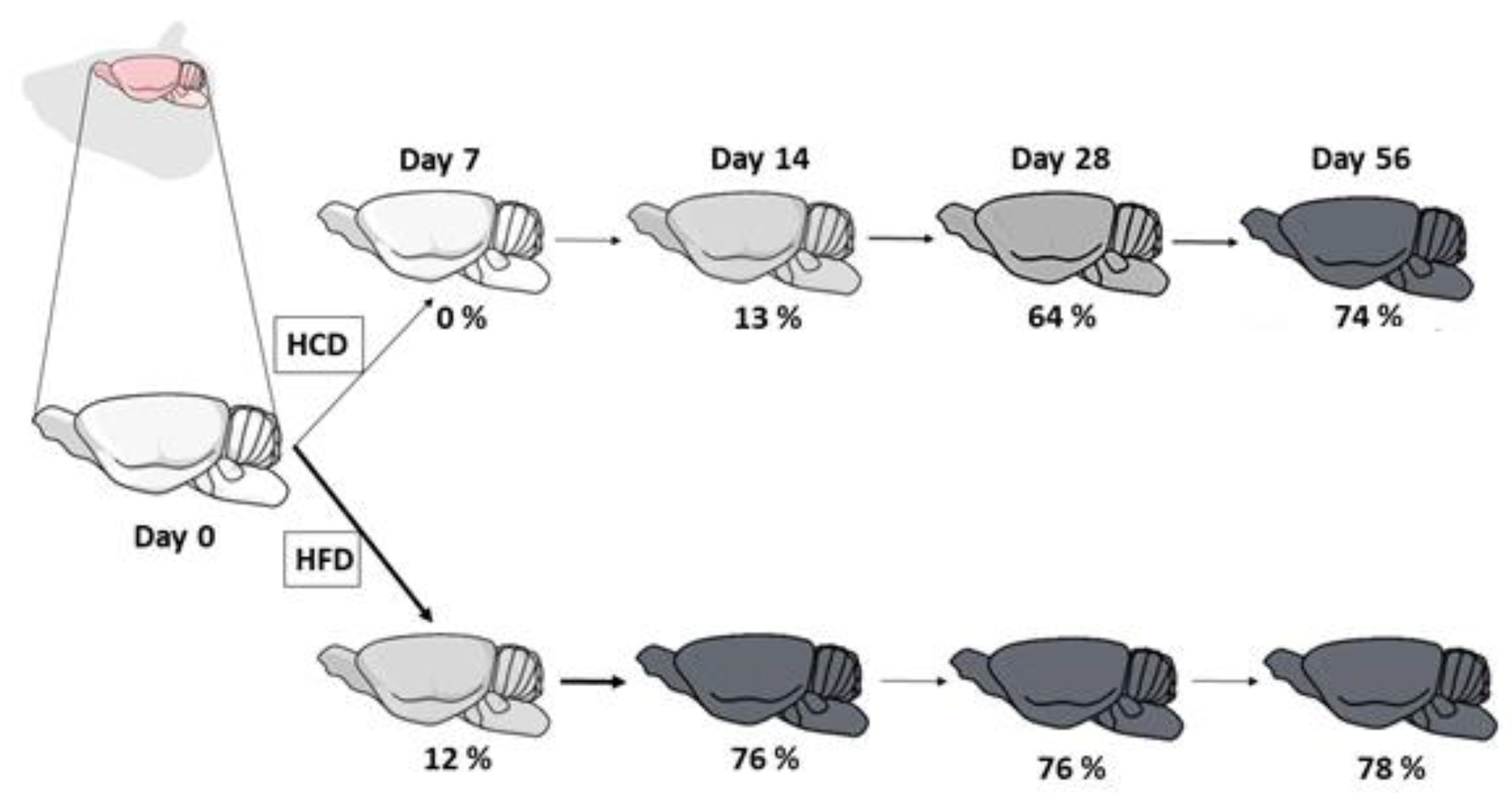
2.1. Animals and Treatments
2.2. Fatty Acid Composition Analysis
2.3. Expressions of Inflammatory Genes and Estimation of the Inflammatory Marker Index (IMI)
2.4. Statistical Analysis
3. Results
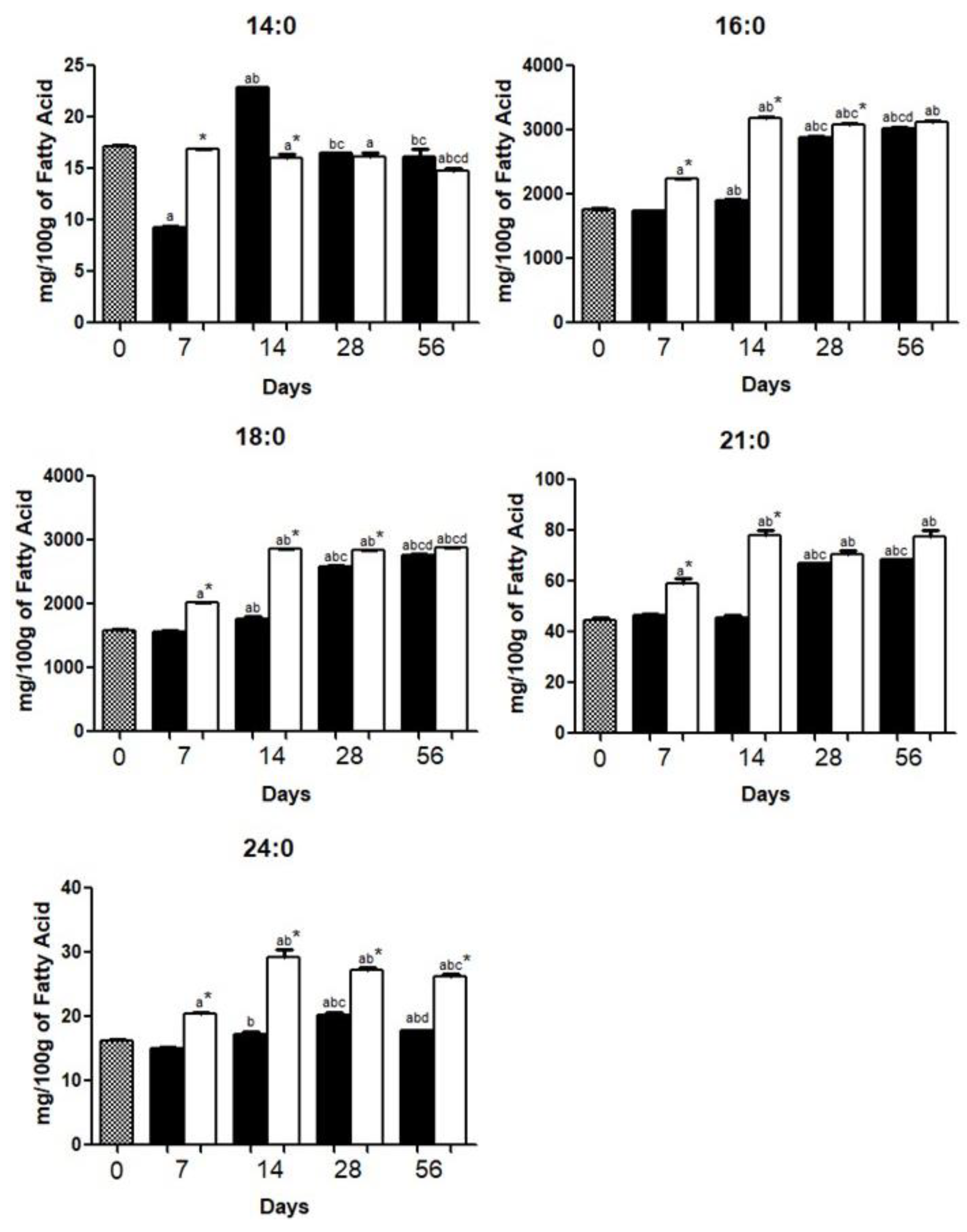
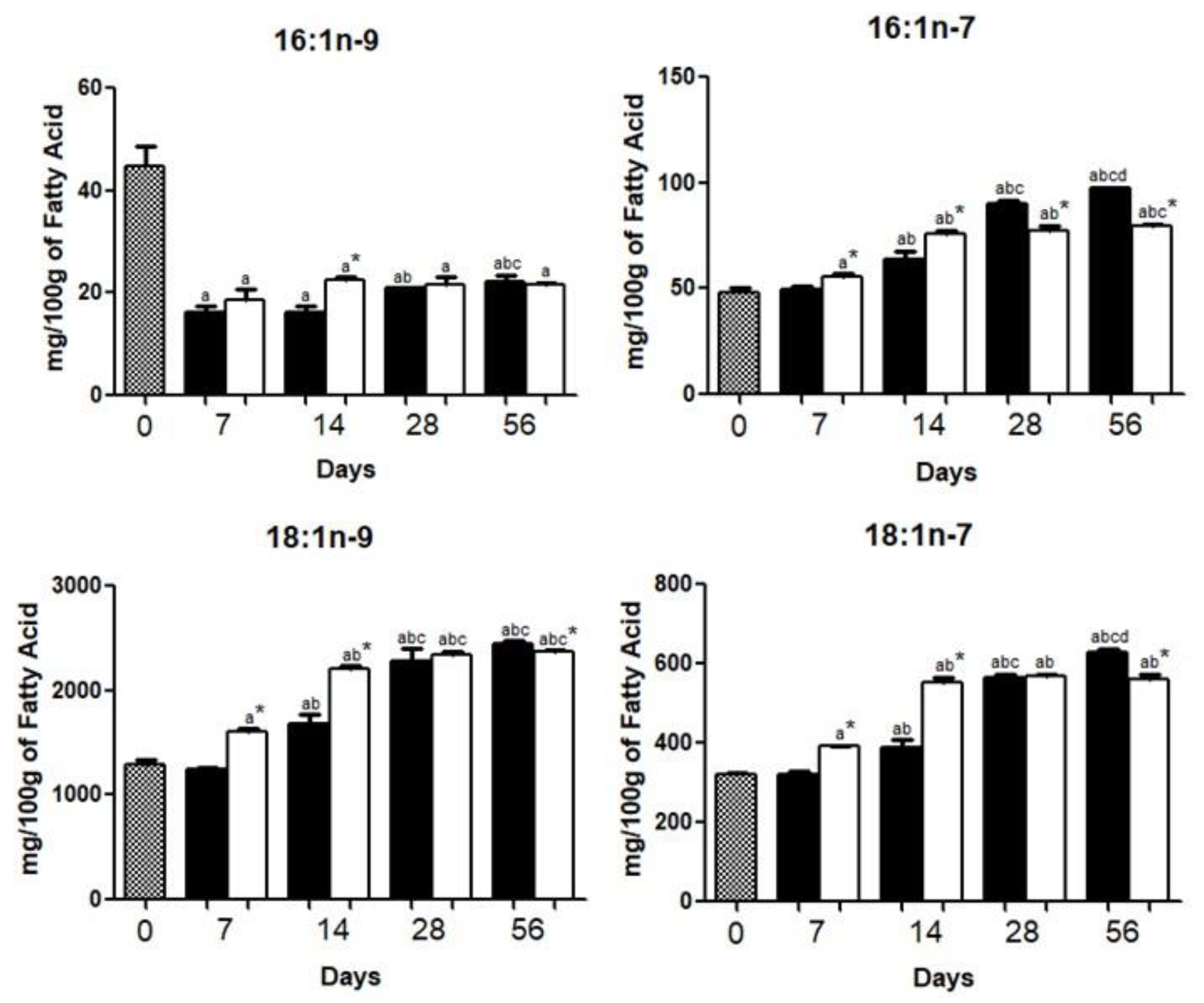
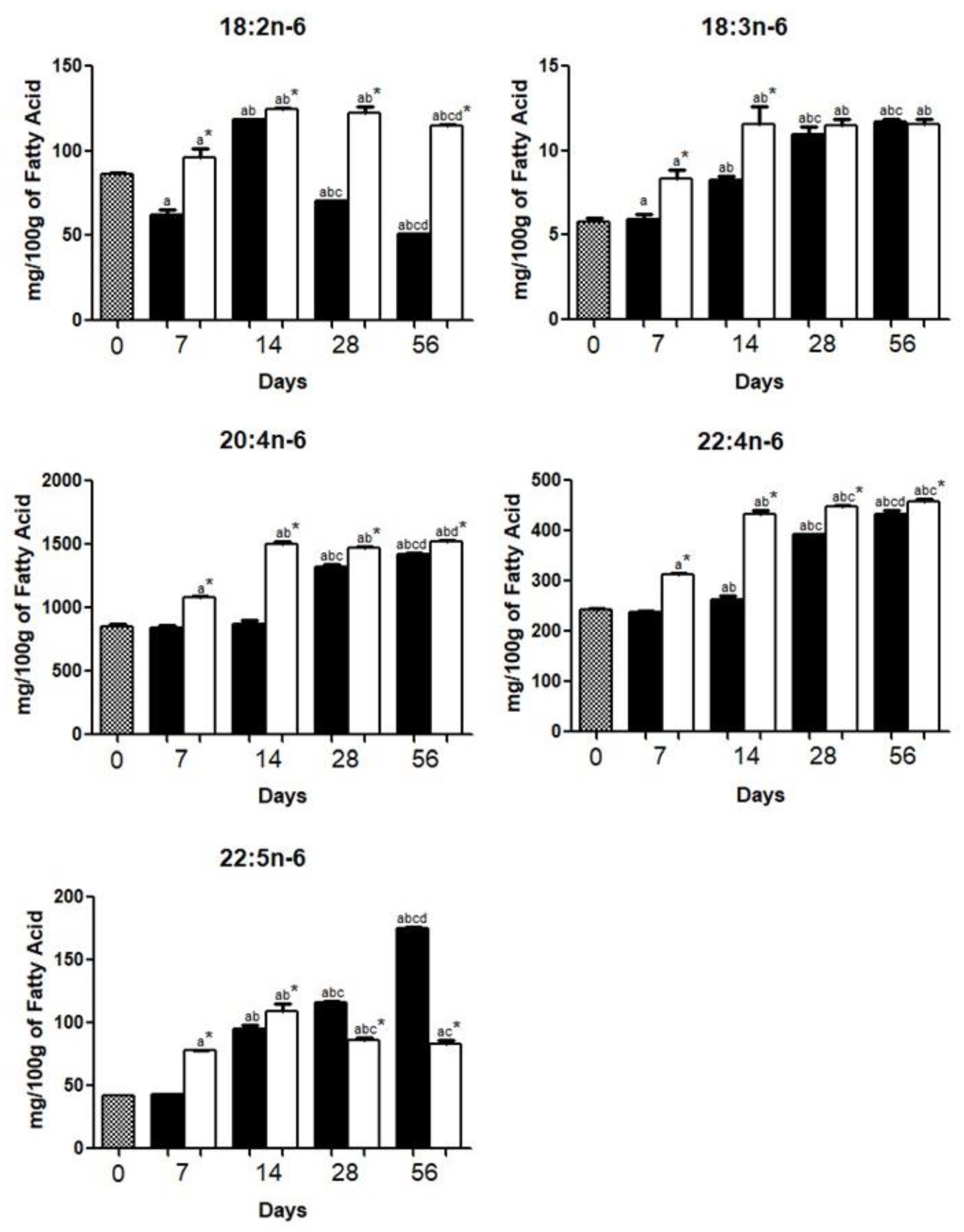
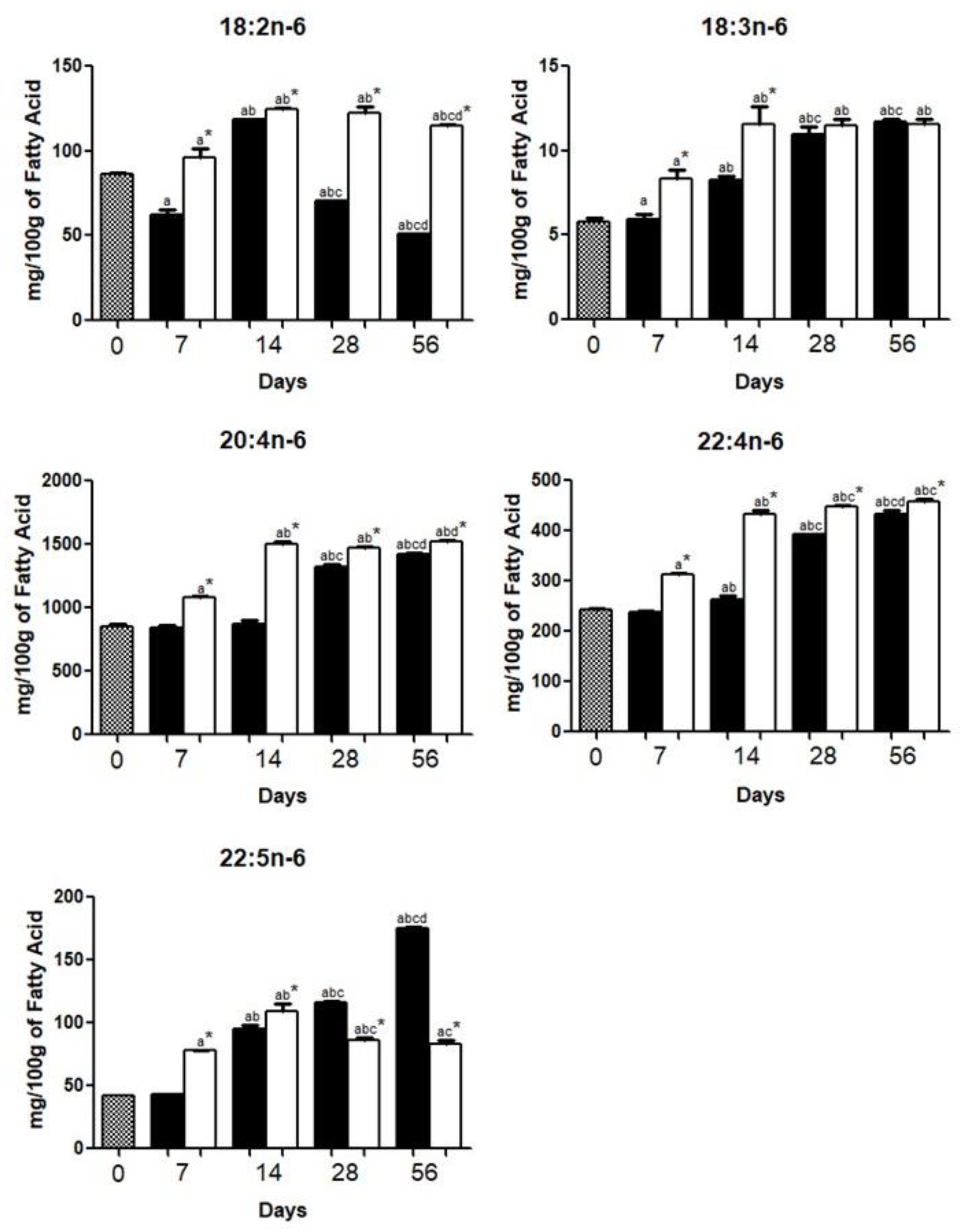
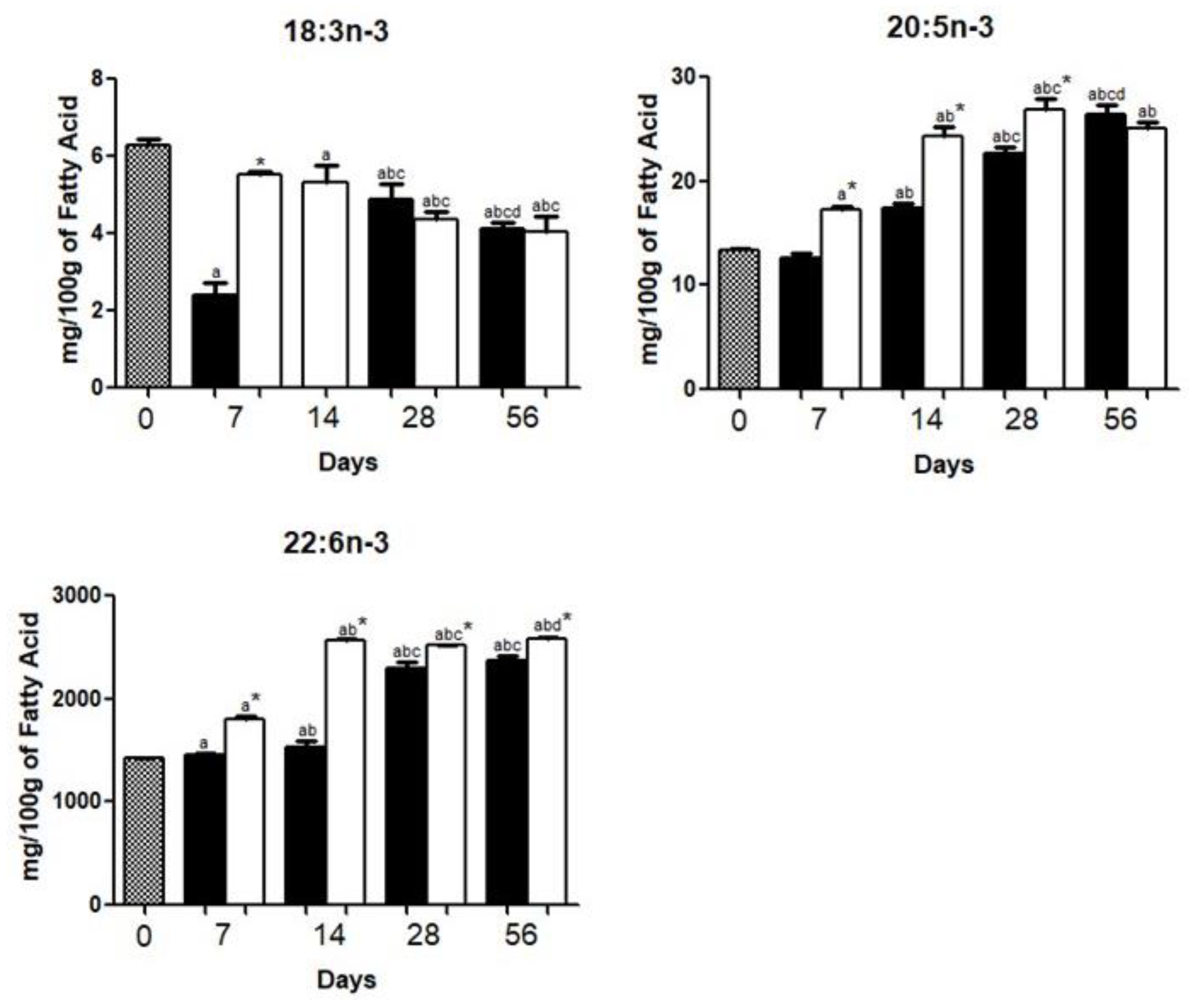
3.1. Brain Fatty Acid Deposition
3.2. Expression of Inflammatory Genes
4. Discussion
4.1. Brain Fatty Acid Deposition
4.2. Expression of Inflammatory Genes
5. Conclusions
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Freitas, H.R.; Ferreira, G.D.C.; Trevenzoli, I.H.; Oliveira, K.D.; de Melo Reis, R.A. Fatty acids, antioxidants and physical activity in brain aging. Nutrients 2017, 9, 1263. [Google Scholar] [CrossRef] [PubMed]
- Molteni, R.; Barnard, R.J.; Ying, Z.; Roberts, C.K.; Gómez-Pinilla, F. A high-fat, refined sugar diet reduces hippocampal brain-derived neurotrophic factor, neuronal plasticity, and learning. Neuroscience 2002, 112, 803–814. [Google Scholar] [CrossRef] [Green Version]
- Denver, P.; Gault, V.A.; McClean, P.L. Sustained high fat diet modulates inflammation, insulin signaling and cognition in mice and a modified xenin peptide ameliorates neuropathology in a chronic high fat model. Diabetes Obes. Metab. 2018, 20, 1166–1175. [Google Scholar] [CrossRef] [PubMed]
- Henderson, S.T. High carbohydrate diets and Alzheimer’s disease. Med. Hypotheses 2004, 62, 689–700. [Google Scholar] [CrossRef] [PubMed]
- Cai, D. Neuroinflammation and neurodegeneration in overnutrition-induced diseases. Trends Endocrinol. Metab. 2013, 24, 40–47. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ross, B.M. ω-3 Fatty acid deficiency in major depressive disorder is caused by the interaction between diet and a genetically determined abnormality in phospholipid metabolism. Med. Hypotheses 2007, 68, 515–524. [Google Scholar] [CrossRef] [PubMed]
- Martín, V.; Fabelo, N.; Santpere, G.; Puig, B.; Marín, R.; Ferrer, I.; Díaz, M. Lipid alterations in lipid rafts from Alzheimer’s disease human brain cortex. J. Alzheimers Dis. 2010, 19, 489–502. [Google Scholar] [CrossRef] [PubMed]
- Chang, C.Y.; Ke, D.S.; Chen, J.Y. Essential fatty acids and human brain. Acta Neurol. Taiwan 2009, 18, 231–241. [Google Scholar] [PubMed]
- Beilharz, J.E.; Maniam, J.; Morris, M.J. Diet-induced cognitive deficits: The role of fat and sugar, potential mechanisms and nutritional interventions. Nutrients 2015, 7, 6719–6738. [Google Scholar] [CrossRef] [PubMed]
- Farooqui, A.A.; Horrocks, L.A.; Farooqui, T. Modulation of inflammation in brain: A matter of fat. J. Neurochem. 2007, 101, 577–599. [Google Scholar] [CrossRef] [PubMed]
- Guillemot-Legris, O.; Muccioli, G.G. Obesity-induced neuroinflammation: Beyond the hypothalamus. Trends Neurosci. 2017, 40, 237–257. [Google Scholar] [CrossRef] [PubMed]
- Da Silva-Santi, L.; Antunes, M.; Caparroz-Assef, S.; Carbonera, F.; Masi, L.; Curi, R.; Visentainer, J.; Bazotte, R. Liver fatty acid composition and inflammation in mice fed with high-carbohydrate diet or high-fat diet. Nutrients 2016, 8, 682. [Google Scholar] [CrossRef] [PubMed]
- Reeves, P.G.; Nielsen, F.H.; Fahey, G.C. AIN-93 Purified diets for laboratory rodents: Final report of the American Institute of Nutrition ad hoc writing committee on the reformulation of the AIN-76A rodent diet. J. Nutr. 1993, 123, 1939–1951. [Google Scholar] [CrossRef] [PubMed]
- Santos, O.O.; Montanher, P.F.; Bonafé, E.G.; Prado, I.N.; Maruyama, S.A.; Matsushita, M.; Visentainer, J.V. A Simple, fast and efficient method for transesterification of fatty acids in foods assisted by ultrasound energy. J. Braz. Chem. Soc. 2014, 25, 1712–1719. [Google Scholar] [CrossRef]
- Liu, W.; Saint, D. A new quantitative method of real time reverse transcription polymerase chain reaction assay based on simulation of polymerase chain reaction kinetics. Anal. Biochem. 2002, 302, 52–59. [Google Scholar] [CrossRef] [PubMed]
- Hussain, G.; Schmitt, F.; Loeffler, J.P.; de Aguilar, J.L.G. Fatting the brain: A brief of recent research. Front. Cell. Neurosci. 2013, 7, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Bazotte, R.B.; Silva, L.G.; Schiavon, F.P.M. Insulin Resistance in the liver: Deficiency or excess of insulin? Cell Cycle 2014, 13, 2494–2500. [Google Scholar] [CrossRef] [PubMed]
- Shaw, C.S.; Clark, J.; Wagenmakers, A.J. The effect of exercise and nutrition on intramuscular fat metabolism and insulin sensitivity. Annu. Rev. Nutr. 2010, 30, 13–34. [Google Scholar] [CrossRef] [PubMed]
- Saponaro, C.; Gaggini, M.; Carli, F.; Gastaldelli, A. The subtle balance between lipolysis and lipogenesis: A critical point in metabolic homeostasis. Nutrients 2015, 7, 9453–9474. [Google Scholar] [CrossRef] [PubMed]
- Abbott, S.K.; Else, P.L.; Atkins, T.A.; Hulbert, A.J. Fatty acid composition of membrane bilayers: Importance of diet polyunsaturated fat balance. Biochim. Biophys. Acta Biomembr. 2012, 1818, 1309–1317. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elsherbiny, M.E.; Goruk, S.; Monckton, E.A.; Richard, C.; Brun, M.; Emara, M.; Field, C.J.; Godbout, R. Long-term effect of docosahexaenoic acid feeding on lipid composition and brain fatty acid-binding protein expression in rats. Nutrients 2015, 7, 8802–8817. [Google Scholar] [CrossRef] [PubMed]
- Calder, P.C. n-3 Fatty acids, inflammation and immunity: New mechanisms to explain old actions. Proc. Nutr. Soc. 2013, 72, 326–336. [Google Scholar] [CrossRef] [PubMed]
- De Luigi, A.; Pizzimenti, S.; Quadri, P.; Lucca, U.; Tettamanti, M.; Fragiacomo, C.; De Simoni, M.G. Peripheral inflammatory response in alzheimer’s disease and multiinfarct dementia. Neurobiol. Dis. 2002, 11, 308–314. [Google Scholar] [CrossRef] [PubMed]
- Bowers, W.J.; Federoff, H.J. Amyloid immunotherapy-engendered CNS inflammation. Neurobiol. Aging 2002, 23, 675–676. [Google Scholar] [CrossRef]
- Schönfeld, P.; Reiser, G. Brain lipotoxicity of phytanic acid and very long-chain fatty acids. Harmful cellular/mitochondrial activities in refsum disease and X-linked adrenoleukodystrophy. Aging Dis. 2016, 7, 136–149. [Google Scholar]
- Udenwobele, D.I.; Su, R.C.; Good, S.V.; Ball, T.B. Myristoylation: An important protein modification in the immune response. Front. Immunol. 2017, 8, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Polo-Hernández, E.; De Castro, F.; García-García, A.G.; Tabernero, A.; Medina, J.M. Oleic acid synthesized in the periventricular zone promotes axonogenesis in the striatum during brain development. J. Neurochem. 2010, 114, 1756–1766. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bossi, E.; Siegrist, H.P.; Zuppinger, K.; Wiesmann, U.; Herschkowitz, N. Utilization of oleic acid and D-beta-OH-butyrate (D-BOHB) as energy substrates by brain cell cultures of newborn mice at different glucose concentrations. Pediatr. Res. 1980, 14, 1417. [Google Scholar] [CrossRef]
- Bentsen, H. Dietary polyunsaturated fatty acids, brain function and mental health. Microb. Ecol. Health Dis. 2017, 28, 1281916. [Google Scholar] [CrossRef]
- Avellini, L.; Terracina, L.; Gaiti, A. Linoleic acid passage through the blood-brain barrier and a possible effect of age. Neurochem. Res. 1994, 19, 129–133. [Google Scholar] [CrossRef] [PubMed]
- Rao, J.S.; Ertley, R.N.; Lee, H.J.; DeMar, J.C.; Arnold, J.T.; Rapoport, S.I.; Bazinet, R.P. n-3 polyunsaturated fatty acid deprivation in rats decreases frontal cortex BDNF via a p38 MAPK-dependent mechanism. Mol. Psychiatry 2007, 12, 36–46. [Google Scholar] [CrossRef] [PubMed]
- Anderson, G.J.; Connor, W.E. Uptake of fatty acids by the developing rat brain. Lipids 1988, 23, 286–290. [Google Scholar] [CrossRef] [PubMed]
- Kitajka, K.; Sinclair, A.J.; Weisinger, R.S.; Weisinger, H.S.; Mathai, M.; Jayasooriya, A.P.; Halver, J.E.; Puskás, L.G. Effects of dietary omega-3 polyunsaturated fatty acids on brain gene expression. Proc. Natl. Acad. Sci. USA 2004, 101, 10931–10936. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiao, Y.; Huang, Y.; Chen, Z.Y. Distribution, depletion and recovery of docosahexaenoic acid are region-specific in rat brain. Br. J. Nutr. 2005, 94, 544. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.T.; Bazinet, R.P. β-oxidation and rapid metabolism, but not uptake regulate brain eicosapentaenoic acid levels. Prostag. Leuk. Essent. Fat. Acids 2015, 92, 33–40. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, S.; Dyer, R.A.; Beasley, C.L. Evidence for altered cell membrane lipid composition in postmortem prefrontal white matter in bipolar disorder and schizophrenia. J. Psychiatr. Res. 2017, 95, 135–142. [Google Scholar] [CrossRef] [PubMed]
- Bazinet, R.P.; Layé, S. Polyunsaturated fatty acids and their metabolites in brain function and disease. Nat. Rev. Neurosci. 2014, 15, 771–785. [Google Scholar] [CrossRef] [PubMed]
- Martínez, M.; Mougan, I. Fatty acid composition of human brain phospholipids during normal development. J. Neurochem. 1998, 71, 2528–2533. [Google Scholar] [CrossRef] [PubMed]
- Haag, M. Essential fatty acids and the brain. Can. J. Psychiatry 2003, 48, 195–203. [Google Scholar] [CrossRef] [PubMed]
- Bousquet, M.; Calon, F.; Cicchetti, F. Impact of ω-3 fatty acids in Parkinson’s disease. Ageing Res. Rev. 2011, 10, 453–463. [Google Scholar] [CrossRef] [PubMed]
- Gupta, S.; Knight, A.G.; Gupta, S.; Keller, J.N.; Bruce-Keller, A.J. Saturated long-chain fatty acids activate inflammatory signaling in astrocytes. J. Neurochem. 2012, 120, 1060–1071. [Google Scholar] [CrossRef] [PubMed]
- Wada, R.; Tifft, C.J.; Proia, R.L. Microglial activation precedes acute neurodegeneration in Sandhoff disease and is suppessed by bone marrow transplantation. Proc. Natl. Acad. Sci. USA 2000, 97, 10954–10959. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Zhang, J.; Jiang, W.; Cao, Z.; Zhao, F.; Cai, T.; Aschner, M.; Luo, W. The role of NLRP3-CASP1 in inflammasome-mediated neuroinflammation and autophagy dysfunction in manganese induced hippocampal-dependent impairment of learning and memoryability. Autophagy 2017, 13, 914–927. [Google Scholar] [CrossRef] [PubMed]


 .
.
.
. .
.
.
. .
.
.
. .
.
.
.
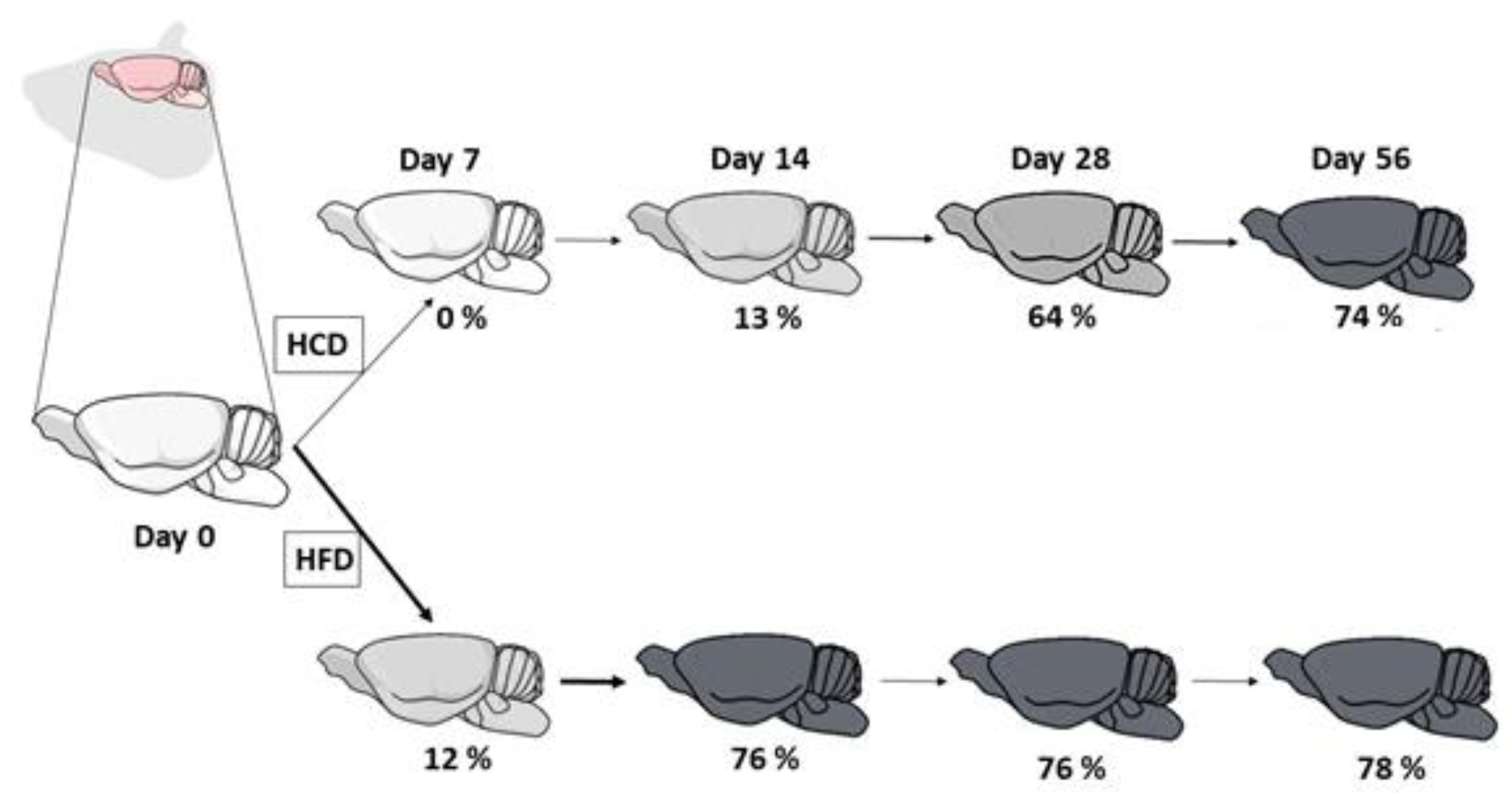

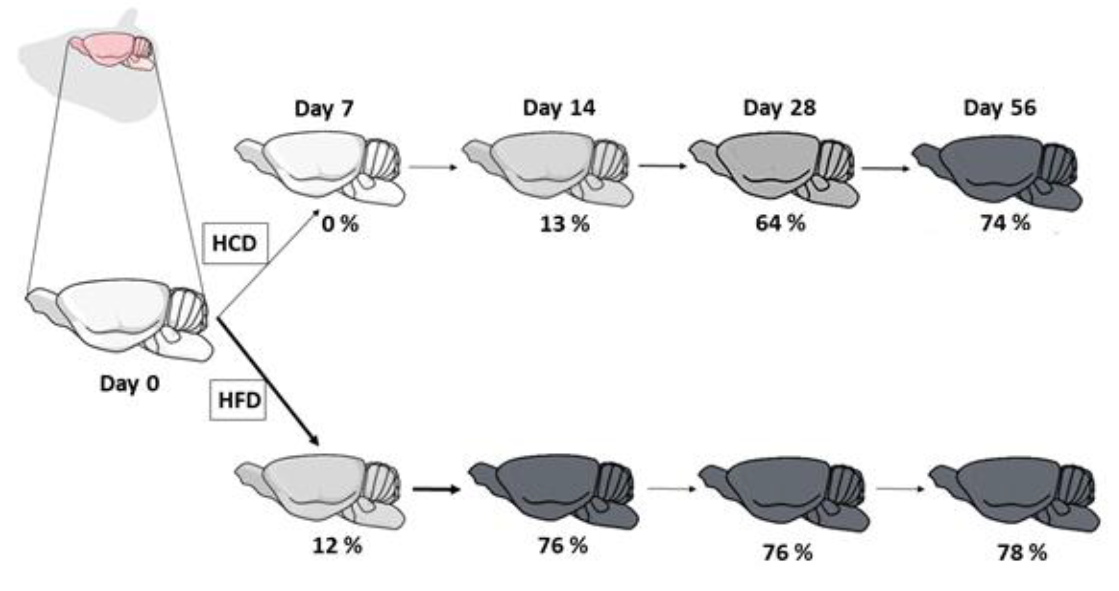{kind=link}
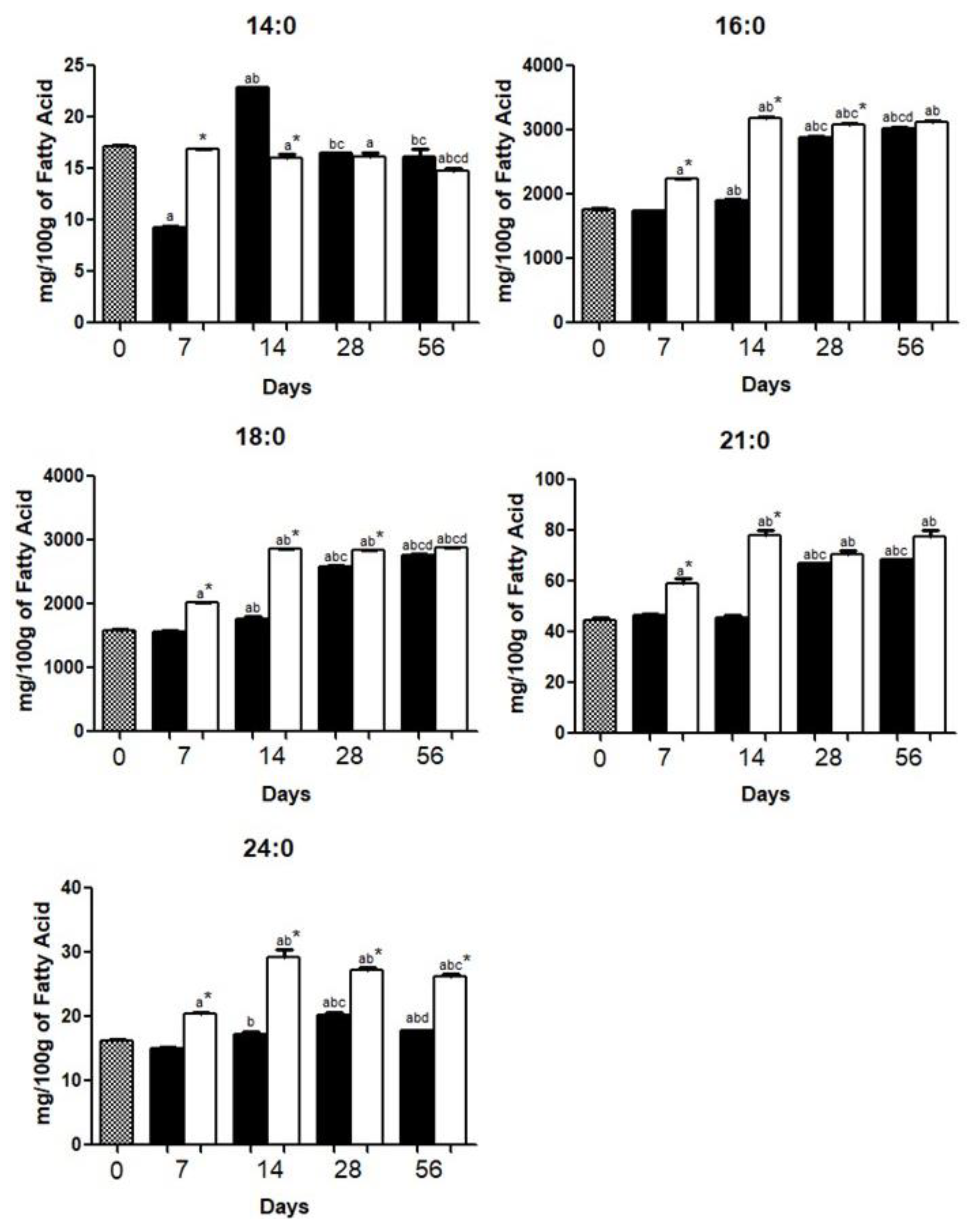{kind=link}
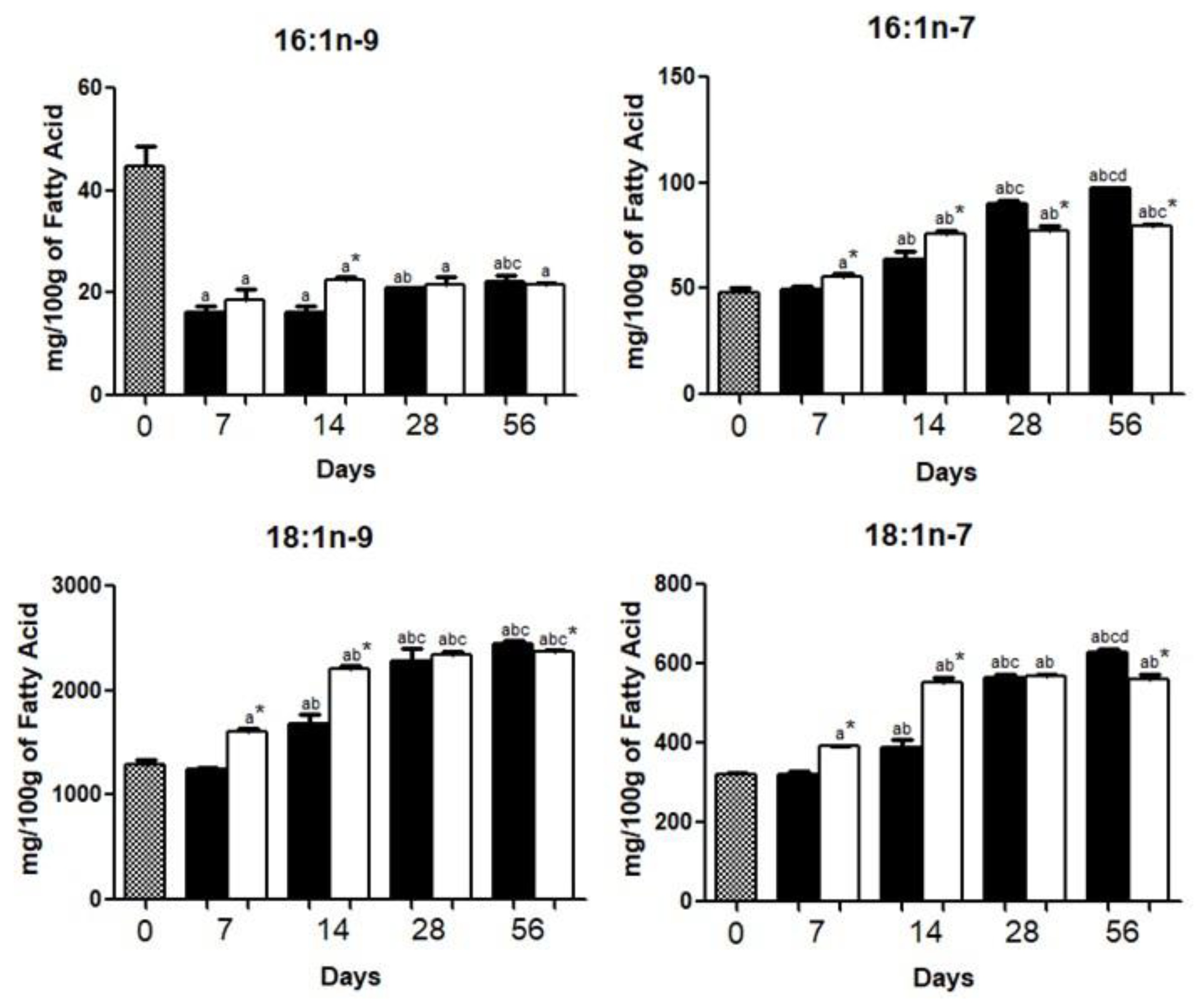{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | RefSeq | Sense | Antisense |
|---|---|---|---|
| Cox 2 | YP_001686701.1 | AACATCCCCTT CCTGCGAAG | AAGTCC ACTCCATGGCCCAG |
| F4/80 | NM_010130.4 | CCTGAACATGCAACCTGCCAC | GGGCATGAGCAGBCTGTAGGATC |
| iNOS | NM_001313921.1 | CGGCAAACCCAAGGTCTACG | CACCTGCTCCTCGCTCAAGTTC |
| Il-6 | NM_001314054.1 | GGTAGCATCCATCATTTCTTTG | CGGAGAGGAGACTTCACAAGAG |
| Il-1β | NM_008361.4 | GGCAGCTACCTGTGTCTTTCCC | ATATGGGTCCG ACAGCACGAG |
| TNF-α | NM_001278601.1 | TCTTCTCATTCCTGCTTGTGGC | CACTTGGTGGTTTGCTACGACG |
| Il-10 | NM_010548.2 | TGCCAAGCCTTATCGGAAATG | AAATCGATGACAGCGCCTCAG |
| Itgam | NM_001082960.1 | TAATGACTCTGCGTTTGCCCTG | ATTGGAGCTGCCCACAATGAG |
| Aif1 | NM_019467.2 | CCTGAGGAGATTTCAACAGAAGC | GGACCGTTCTCACACTTCCC |
| Fatty Acids | 0 (Day) | 7 (Day) | 14 (Day) | 28 (Day) | 56 (Day) | ∆% | |
|---|---|---|---|---|---|---|---|
| SFA | HCD | 3438.6 ± 13.1 | 3382.6 ± 9.7 | 3761.6 ± 34.9 a,b | 5577.5 ± 47.2 a,b,c | 5905.9 ± 23.9 a,b,c,d | 71.7% |
| HFD | 4363.6 ± 15.0 a,* | 6157.7 ± 41.7 a,b,* | 6047.2 ± 26.0 a,b,c,* | 6122.4 ± 34.4 a,b,c,d,* | 78.0% | ||
| MUFA | HCD | 1709.0 ± 41.6 | 1632.3 ± 17.3 | 2147.9 ± 101.9 a,b | 2956.7 ± 119.1 a,b,c | 3202.5 ± 31.1a,b,c,d | 87.3% |
| HFD | 2072.2 ± 29.7 a,* | 2859.4 ± 36.2 a,b,* | 3012.9 ± 22.8 a,b,c | 3035.1 ± 24.8 a,b,c,* | 77.5% | ||
| PUFA | HCD | 2665.6 ± 26.8 | 2660.1 ± 21.5 | 2938.0 ± 60.4 a,b | 4245.7 ± 57.2 a,b,c | 4504.6 ± 36.8 a,b,c,d | 68.9% |
| HFD | 3401.8 ± 31.1a,* | 4784.2 ± 25.9 a,b,* | 4693.0 ± 11.5 a,b,c,* | 4810.7 ± 9.0 a,b,d,* | 80.4% | ||
| n-6 | HCD | 1224.8 ± 23.1 | 1189.1 ± 18.3 | 1360.3 ± 29.8 a,b | 1914.9 ± 20.8 a,b,c | 2094.9 ± 8.5 a,b,c,d | 71.0% |
| HFD | 1574.3 ± 13.2 a,* | 2182.3 ± 22.9 a,b,* | 2141.6 ± 8.0 a,b,c,* | 2192.5 ± 8.0 a,b,d,* | 79.0% | ||
| n-3 | HCD | 1440.8 ± 13.5 | 1471.0 ± 11.3 | 1577.7 ± 52.5 a,b | 2330.8 ± 53.3 a,b,c | 2409.8 ± 35.8 a,b,c | 67.2% |
| HFD | 1827.5 ± 28.2 a,* | 2601.9 ± 11.9 a,b,* | 2551.4 ± 8.2 a,b,c,* | 2618.2 ± 4.1 a,b,d,* | 81.7% | ||
| PUFA/SFA | HCD | 0.78 ± 0.01 | 0.79 ± 0.01 | 0.78 ± 0.02 | 0.76 ± 0.02 | 0.76 ± 0.01 | −2.6% |
| HFD | 0.78 ± 0.01 | 0.78 ± 0.01 | 0.78 ± 0.01 | 0.79 ± 0.01 | 1.2% | ||
| MUFA/SFA | HCD | 0.50 ± 0.03 | 0.48 ± 0.01 | 0.57 ± 0.05 a,b | 0.53 ± 0.04 | 0.54 ± 0.01 b | 8.0% |
| HFD | 0.47 ± 0.02 | 0.46 ± 0.02 a | 0.50 ± 0.01b | 0.50 ± 0.01 b,c | 0.0% | ||
| n-6/n-3 | HCD | 0.85 ± 0.02 | 0.81 ± 0.02 a | 0.86 ± 0.04 b | 0.82 ± 0.03 a,c | 0.87 ± 0.02 b,d | 2.3% |
| HFD | 0.86 ± 0.02 | 0.84 ± 0.01 b | 0.84 ± 0.01 b | 0.84 ± 0.01 b | −1.1% | ||
| SUM | HCD | 7813.3 | 7675.1 | 8847.5 | 12792.5 | 13613.1 | 74.2% |
| HFD | 9837.7 | 13801.3 | 13753.1 | 13968.2 | 78.7% |
| Genes | Total Brain Tissue | |
|---|---|---|
| HCD | HFD | |
| F4/80 | 1.41 ± 0.21 | 0.75 ± 0.11 * |
| IL-6 | 1.12 ± 0.14 | 0.92 ± 0.08 |
| IL-1β | 1.52 ± 0.23 | 1.28 ± 0.16 |
| TNF-α | 0.49 ± 0.06 | 0.26 ± 0.05 |
| iNOS | 1.10 ± 0.11 | 1.23 ± 0.33 |
| IL-10 | 2.09 ± 0.55 | 1.23 ± 0.33 |
| COX-2 | 1.12 ± 0.11 | 1.33 ± 0.17 |
| Itgam | 1.31 ± 0.18 | 0.83 ± 0.08 * |
| Aif1 | 1.63 ± 0.22 | 0.89 ± 0.20 |
| IMI | 10.38 | 15.20 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gimenez da Silva-Santi, L.; Masetto Antunes, M.; Mori, M.A.; Biesdorf de Almeida-Souza, C.; Vergílio Visentainer, J.; Carbonera, F.; Rabello Crisma, A.; Nunes Masi, L.; Massao Hirabara, S.; Curi, R.; et al. Brain Fatty Acid Composition and Inflammation in Mice Fed with High-Carbohydrate Diet or High-Fat Diet. Nutrients 2018, 10, 1277. https://doi.org/10.3390/nu10091277
Gimenez da Silva-Santi L, Masetto Antunes M, Mori MA, Biesdorf de Almeida-Souza C, Vergílio Visentainer J, Carbonera F, Rabello Crisma A, Nunes Masi L, Massao Hirabara S, Curi R, et al. Brain Fatty Acid Composition and Inflammation in Mice Fed with High-Carbohydrate Diet or High-Fat Diet. Nutrients. 2018; 10(9):1277. https://doi.org/10.3390/nu10091277
Chicago/Turabian StyleGimenez da Silva-Santi, Lorena, Marina Masetto Antunes, Marco Aurélio Mori, Camila Biesdorf de Almeida-Souza, Jesuí Vergílio Visentainer, Fabiana Carbonera, Amanda Rabello Crisma, Laureane Nunes Masi, Sandro Massao Hirabara, Rui Curi, and et al. 2018. "Brain Fatty Acid Composition and Inflammation in Mice Fed with High-Carbohydrate Diet or High-Fat Diet" Nutrients 10, no. 9: 1277. https://doi.org/10.3390/nu10091277