Effects of Tempeh Fermentation with Lactobacillus plantarum and Rhizopus oligosporus on Streptozotocin-Induced Type II Diabetes Mellitus in Rats


Abstract
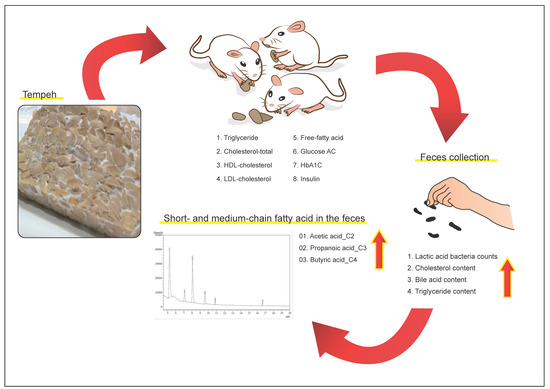
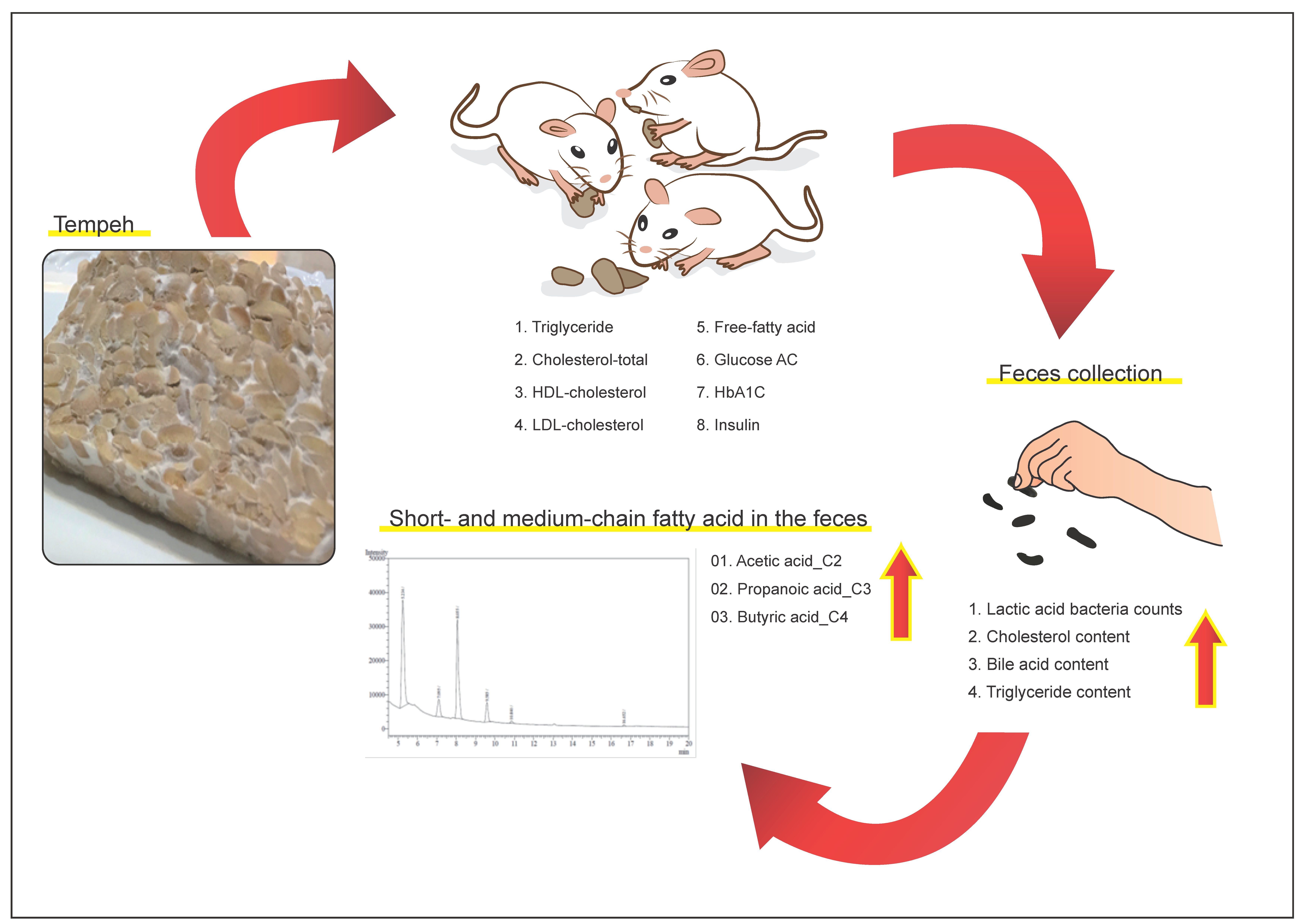
:
1. Introduction
2. Materials and Methods
2.1. Sample Preparation
2.2. Animals and Diets
2.3. Serum Samples
2.4. Fasting Serum Glucose
2.5. Oral Glucose Tolerance Test (OGTT)
2.6. Biochemical Measurements
2.7. Homeostasis Model Assessment-Insulin Resistance (HOMA-IR)
2.8. Stool Assay
2.9. Next-Generation Sequencing Analysis of Stool Samples
2.9.1. Amplicon Library Construction and Sequencing
2.9.2. Bioinformatic Analysis
2.10. Statistical Analysis
3. Results
3.1. Hyperglycemic Rat Model
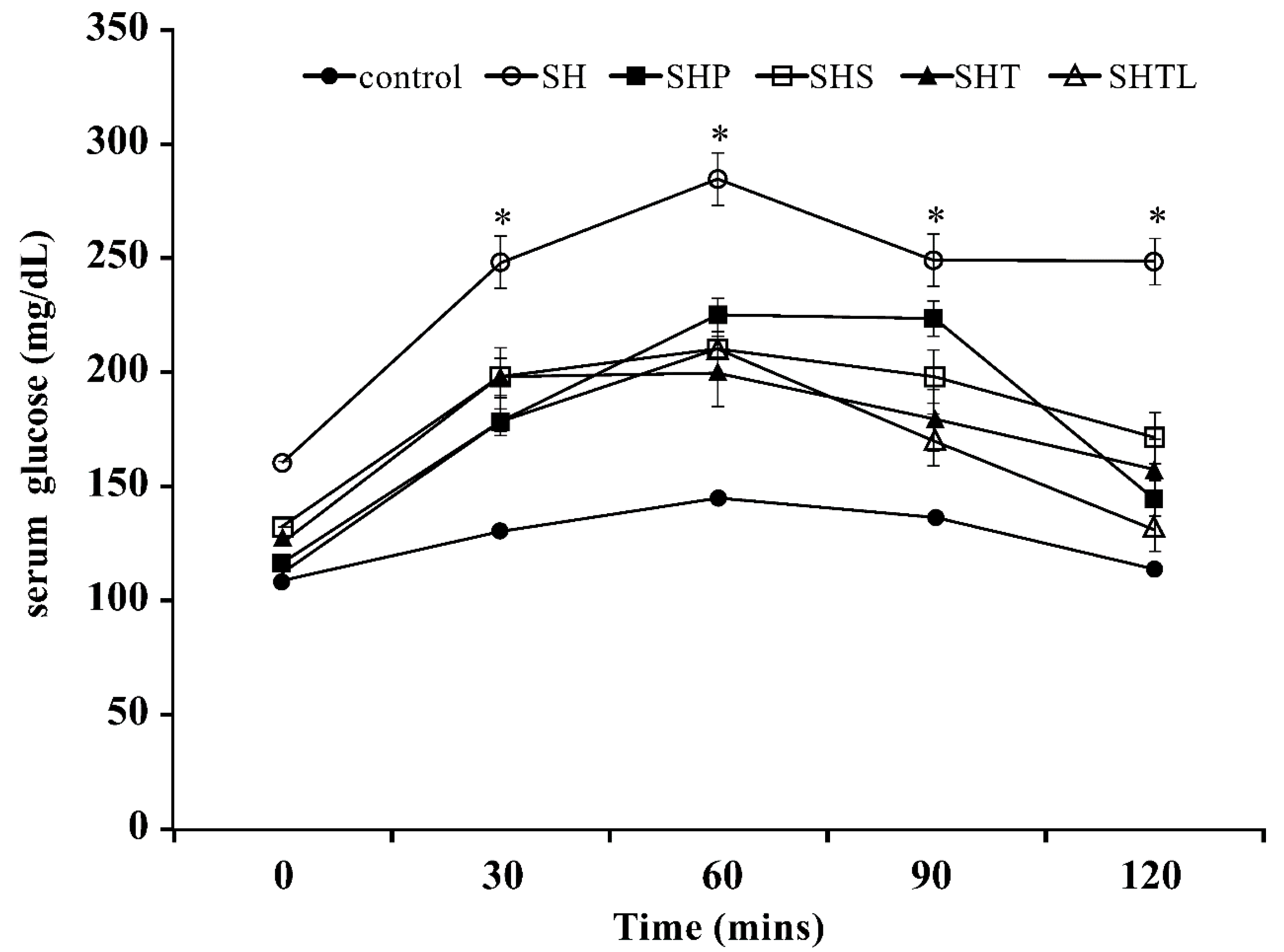
3.2. Oral Glucose Tolerance Test
3.3. Effects of Various Treatments on Serum Biochemistry in DM Rats
3.4. Changes in Total Lactic Acid Bacteria in Diabetes Mellitus (DM) Rat Stools
3.5. Changes in Short Chain Fatty Acids (SCFAs) in DM Rat Stools
3.6. Changes in Total Cholesterol, Bile Acid, and TG in DM Rat Stools
3.7. Microbiota Analysis of DM Rats
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Guariguata, L.; Whiting, D.R.; Hambleton, I.; Beagley, J.; Linnenkamp, U.; Shaw, J.E. Global estimates of diabetes prevalence for 2013 and projections for 2035. Diabetes Res. Clin. Pract. 2014, 103, 137–149. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Wang, X.; Wang, J.; Wu, F.; Sui, Y.; Yang, L.; Wang, Z. Lactobacillus plantarum strains as potential probiotic cultures with cholesterol-lowering activity. J. Dairy Sci. 2013, 96, 2746–2753. [Google Scholar] [CrossRef] [PubMed]
- Panwar, H.; Calderwood, D.; Gillespie, A.L.; Wylie, A.R.; Graham, S.F.; Grant, I.R.; Green, B.D. Identification of lactic acid bacteria strains modulating incretin hormone secretion and gene expression in enteroendocrine cells. J. Funct. Foods 2016, 23, 348–358. [Google Scholar] [CrossRef] [Green Version]
- Barus, T.; Wati, L.; Suwanto, A. Diversity of protease-producing Bacillus spp. from fresh Indonesian tempeh based on 16S rRNA gene sequence. HAYATI J. Biosci. 2017, 24, 35–40. [Google Scholar] [CrossRef]
- Behloul, N.; Wu, G. Genistein: A promising therapeutic agent for obesity and diabetes treatment. Eur. J. Pharmacol. 2013, 698, 31–38. [Google Scholar] [CrossRef] [PubMed]
- Jeleń, H.; Majcher, M.; Ginja, A.; Kuligowski, M. Determination of compounds responsible for tempeh aroma. Food Chem. 2013, 141, 459–465. [Google Scholar] [CrossRef] [PubMed]
- Haron, H.; Ismail, A.; Shahar, S.; Azlan, A.; Peng, L.S. Apparent bioavailability of isoflavones in urinary excretions of postmenopausal Malay women consuming tempeh compared with milk. Int. J. Food Sci. Nutr. 2011, 62, 642–650. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahmad, A.; Ramasamy, K.; Majeed, A.B.A.; Mani, V. Enhancement of β-secretase inhibition and antioxidant activities of tempeh, a fermented soybean cake through enrichment of bioactive aglycones. Pharm. Biol. 2015, 53, 758–766. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.M.; Zhang, Y.B.; Chi, M.H. Soy protein supplementation reduces clinical indices in type 2 diabetes and metabolic syndrome. Yonsei Med. J. 2016, 57, 681–689. [Google Scholar] [CrossRef] [PubMed]
- Feizollahzadeh, S.; Ghiasvand, R.; Rezaei, A.; Khanahmad, H.; Hariri, M. Effect of probiotic soy milk on serum levels of adiponectin, inflammatory mediators, lipid profile, and fasting blood glucose among patients with type II diabetes mellitus. Probiotics Antimicrob. Proteins 2017, 9, 41–47. [Google Scholar] [CrossRef] [PubMed]
- Gandhi, G.R.; Stalin, A.; Balakrishna, K.; Ignacimuthu, S.; Paulraj, M.G.; Vishal, R. Insulin sensitization via partial agonism of PPARγ and glucose uptake through translocation and activation of GLUT4 in PI3K/p-Akt signaling pathway by embelin in type 2 diabetic rats. Biochim. Biophys. Acta 2013, 1830, 2243–2255. [Google Scholar] [CrossRef] [PubMed]
- Matthews, D.R.; Hosker, J.P.; Rudenski, A.S.; Naylor, B.A.; Treacher, D.F.; Turner, R.C. Homeostasis model assessment: Insulin resistance and β-cell function from fasting plasma glucose and insulin concentrations in man. Diabetologia 1985, 28, 412–419. [Google Scholar] [CrossRef] [PubMed]
- Holben, W.E.; Williams, P.; Saarinen, M.; Särkilahti, L.K.; Apajalahti, J.H.A. Phylogenetic analysis of intestinal microflora indicates a novel Mycoplasma phylotype in farmed and wild salmon. Microb. Ecol. 2002, 44, 175–185. [Google Scholar] [CrossRef] [PubMed]
- Ramu, R.; Shirahatti, P.S.; Zameer, F.; Dhananjaya, B.L.; Prasad, N. Assessment of in vivo antidiabetic properties of umbelliferone and lupeol constituents of banana (Musa sp. var. Nanjangud Rasa Bale) flower in hyperglycaemic rodent model. PLoS ONE 2016, 11, e0151135. [Google Scholar]
- Lebovitz, H.E. Postprandial hyperglycaemic state: Importance and consequences. Diabetes Res. Clin. Pract. 1998, 40, S27–S28. [Google Scholar] [PubMed]
- Ali, A.A.; Velasquez, M.T.; Hansen, C.T.; Mohamed, A.I.; Bhathena, S.J. Modulation of carbohydrate metabolism and peptide hormones by soybean isoflavones and probiotics in obesity and diabetes. J. Nutr. Biochem. 2005, 16, 6993–6999. [Google Scholar] [CrossRef] [PubMed]
- Park, S.; Kim, D.S.; Kim, J.H.; Kim, J.S.; Kim, H.J. Glyceollin-containing fermented soybeans improve glucose homeostasis in diabetic mice. Nutrition 2012, 28, 204–211. [Google Scholar] [CrossRef] [PubMed]
- Himsworth, H.P. Diabetes mellitus: Its differentiation into insulin-sensitive and insulin-insensitive types. Lancet 1936, 227, 127–130. [Google Scholar] [CrossRef]
- Yalow, R.S.; Berson, S.A. Plasma insulin concentrations in nondiabetic and early diabetic subjects: Determinations by a new sensitive immuno-assay technic. Diabetes 1960, 9, 254–260. [Google Scholar] [CrossRef] [PubMed]
- Kahn, C.R.; Flier, J.S.; Bar, R.S.; Archer, J.A.; Gorden, P.; Martin, M.M. The syndromes of insulin resistance and acanthosis nigricans: Insulin-receptor disorders in man. N. Engl. J. Med. 1976, 294, 739–745. [Google Scholar] [CrossRef] [PubMed]
- Olefsky, J.; Farquhar, J.W.; Reaven, G. Relationship between fasting plasma insulin level and resistance to insulin-mediated glucose uptake in normal and diabetic subjects. Diabetes 1973, 22, 507–513. [Google Scholar] [CrossRef] [PubMed]
- Kolterman, O.G.; Insel, J.; Saekow, M.; Olefsky, J.M. Mechanisms of insulin resistance in human obesity: Evidence for receptor and postreceptor defects. J. Clin. Investig. 1980, 65, 1272–1284. [Google Scholar] [CrossRef] [PubMed]
- Howard, G.O.; Leary, D.H.; Zaccaro, D.; Haffner, S.; Rewers, M.; Hamman, R. Insulin sensitivity and atherosclerosis. Circulation 1996, 93, 1809–1817. [Google Scholar] [CrossRef] [PubMed]
- Yip, J.; Facchini, F.S.; Reaven, G.M. Resistance to insulin-mediated glucose disposal as a predictor of cardiovascular disease. J. Clin. Endocrinol. MeTab. 1998, 83, 2773–2776. [Google Scholar] [CrossRef] [PubMed]
- Park, S.C.; Hwang, M.H.; Kim, Y.H.; Kim, J.C.; Song, J.C.; Lee, K.W. Comparison of pH and bile resistance of Lactobacillus acidophilus strains isolated from rat, pig, chicken, and human sources. World J. Microbiol. Biotechnol. 2006, 22, 35–37. [Google Scholar] [CrossRef]
- Danielson, A.D.; Peo, E.R., Jr.; Shahani, K.M.; Lewis, A.J.; Whalen, P.J.; Amer, M.A. Anticholesteremic property of Lactobacillus acidophilus yogurt fed to mature boars. J. Anim. Sci. Technol. 1989, 67, 966–974. [Google Scholar] [CrossRef]
- Liong, M.T.; Shah, N.P. Effects of a Lactobacillus casei synbiotic on serum lipoprotein, intestinal microflora, and organic acids in rats. J. Dairy Sci. 2006, 89, 1390–1399. [Google Scholar] [CrossRef]
- Choi, M.S.; Jung, U.J.; Yeo, J.K.; Kim, M.J.; Lee, M.K. Genistein and daidzein prevent diabetes onset by elevating insulin level and altering hepatic gluconeogenic and lipogenic enzyme activities in non-obese diabetic (NOD) mice. Diabetes Metab. Res. Rev. 2008, 24, 74–81. [Google Scholar] [CrossRef] [PubMed]
- Ding, M.; Pan, A.; Manson, J.E.; Willett, W.C.; Malik, V.; Rosner, B. Consumption of soy foods and isoflavones and risk of type 2 diabetes: A pooled analysis of three US cohorts. Eur. J. Clin. Nutr. 2016, 70, 1381. [Google Scholar] [CrossRef] [PubMed]
- Mezei, O.; Banz, W.J.; Steger, R.W.; Peluso, M.R.; Winters, T.A.; Shay, N. Soy isoflavones exert antidiabetic and hypolipidemic effects through the PPAR pathways in obese Zucker rats and murine RAW 264.7 cells. J. Nutr. 2003, 133, 1238–1243. [Google Scholar] [CrossRef] [PubMed]
- Park, S.A.; Choi, M.S.; Cho, S.Y.; Seo, J.S.; Jung, U.J.; Kim, M.J. Genistein and daidzein modulate hepatic glucose and lipid regulating enzyme activities in C57BL/KsJ-db/db mice. Life Sci. 2006, 79, 1207–1213. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.S. Effects of soy protein and genistein on blood glucose, antioxidant enzyme activities, and lipid profile in streptozotocin-induced diabetic rats. Life Sci. 2006, 79, 1578–1584. [Google Scholar] [CrossRef] [PubMed]
- Kwon, D.Y.; Daily, J.W.; Kim, H.J.; Park, S. Antidiabetic effects of fermented soybean products on type 2 diabetes. Nutr. Res. 2010, 30, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Anandharaj, M.; Sivasankari, B.; Santhanakaruppu, R.; Manimaran, M.; Rani, R.P.; Sivakumar, S. Determining the probiotic potential of cholesterol-reducing Lactobacillus and Weissella strains isolated from gherkins (fermented cucumber) and south Indian fermented koozh. Res. Microbiol. 2015, 166, 428–439. [Google Scholar] [CrossRef] [PubMed]
- Erkkilä, S.; Petäjä, E. Screening of commercial meat starter cultures at low pH and in the presence of bile salts for potential probiotic use. Meat Sci. 2000, 55, 297–300. [Google Scholar] [CrossRef]
- Tsai, Y.T.; Cheng, P.C.; Fan, C.K.; Pan, T.M. Time-dependent persistence of enhanced immune response by a potential probiotic strain Lactobacillus paracasei subsp. paracasei NTU 101. Int. J. Food Microbiol. 2008, 128, 219–225. [Google Scholar] [CrossRef] [PubMed]
- Panasevich, M.R.; Schuster, C.M.; Phillips, K.E.; Meers, G.M.; Chintapalli, S.V.; Wankhade, U. Soy compared with milk protein in a Western diet changes fecal microbiota and decreases hepatic steatosis in obese OLETF rats. J. Nutr. Biochem. 2017, 46, 125–136. [Google Scholar] [CrossRef] [PubMed]
- Shankardass, K.; Chuchmach, S.; Chelswick, K.; Stefanovich, C.; Spurr, S.; Brooks, J. Bowel function of long-term tube-fed patients consuming formulae with and without dietary fiber. J. Parenter. Enteral. Nutr. 1990, 14, 508–512. [Google Scholar] [CrossRef] [PubMed]
- Schneider, S.M.; Pouget, I.; Staccini, P.; Rampal, P.; Hebuterne, X. Quality of life in long-term home enteral nutrition patients. Clin. Nutr. 2000, 19, 23–28. [Google Scholar] [CrossRef] [PubMed]
- Cabré, E. Fibre supplementation of enteral formula-diets: A look to the evidence. Clin. Nutr. 2004, 1, 63–71. [Google Scholar] [CrossRef]
- Meier, R.; Gassull, M.A. Consensus recommendations on the effects and benefits of fibre in clinical practice. Clin. Nutr. 2004, 1, 73–80. [Google Scholar] [CrossRef]
- Hofman, Z.; Van Drunen, J.D.E.; De Later, C.; Kuipers, H. The effect of different nutritional feeds on the postprandial glucose response in healthy volunteers and patients with type II diabetes. Eur. J. Clin. Nutr. 2004, 58, 1553. [Google Scholar] [CrossRef] [PubMed]
- Read, N.W. Diarrhee motrice. Clin. Gastroenterol. 1986, 15, 657–686. [Google Scholar] [PubMed]
- Russell, W.R.; Hoyles, L.; Flint, H.J.; Dumas, M.E. Colonic bacterial metabolites and human health. Curr. Opin. Microbiol. 2013, 16, 246–254. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Topping, D.L.; Clifton, P.M. Short-chain fatty acids and human colonic function: Roles of resistant starch and nonstarch polysaccharides. Physiol. Rev. 2001, 81, 1031–1064. [Google Scholar] [CrossRef] [PubMed]
- Fukuda, S.; Toh, H.; Hase, K.; Oshima, K.; Nakanishi, Y.; Yoshimura, K.; Taylor, T.D. Bifidobacteria can protect from enteropathogenic infection through production of acetate. Nature 2011, 469, 543–547. [Google Scholar] [CrossRef] [PubMed]
- Maslowski, K.M.; Vieira, A.T.; Ng, A.; Kranich, J.; Sierro, F.; Yu, D. Regulation of inflammatory responses by gut microbiota and chemoattractant receptor GPR43. Nature 2009, 461, 1282. [Google Scholar] [CrossRef] [PubMed]
- Hara, H.; Haga, S.; Aoyama, Y.; Kiriyama, S. Short-chain fatty acids suppress cholesterol synthesis in rat liver and intestine. J. Nutr. 1999, 129, 942–948. [Google Scholar] [CrossRef] [PubMed]
- Hughes, S.A.; Shewry, P.R.; Gibson, G.R.; McCleary, B.V.; Rastall, R.A. In vitro fermentation of oat and barley derived β-glucans by human faecal microbiota. FEMS Microbiol. Ecol. 2008, 64, 482–493. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arora, T.; Sharma, R.; Frost, G. Propionate. Anti-obesity and satiety enhancing factor? Appetite 2011, 56, 511–515. [Google Scholar] [CrossRef] [PubMed]
- Lin, H.V.; Frassetto, A.; Kowalik, E.J.; Nawrocki, A.R.; Lu, M.M.; Kosinski, J.R. Butyrate and propionate protect against diet-induced obesity and regulate gut hormones via free fatty acid receptor 3-independent mechanisms. PLoS ONE 2012, 7, e35240. [Google Scholar] [CrossRef] [PubMed]
- Schneider, S.M.; Girard-Pipau, F.; Anty, R.; van der Linde, E.G.; Philipsen-Geerling, B.J.; Knol, J. Effects of total enteral nutrition supplemented with a multi-fibre mix on faecal short-chain fatty acids and microbiota. Clin. Nutr. 2006, 25, 82–90. [Google Scholar] [CrossRef] [PubMed]
- Clarkson, T.B. Soy, soy phytoestrogens and cardiovascular disease. J. Nutr. 2002, 132, 566S–569S. [Google Scholar] [CrossRef] [PubMed]
- Lichtenstein, A.H.; Jalbert, S.M.; Adlercreutz, H.; Goldin, B.R.; Rasmussen, H.; Schaefer, E.J. Lipoprotein response to diets high in soy or animal protein with and without isoflavones in moderately hypercholesterolemic subjects. Arterioscler. Thromb. Vasc. Biol. 2002, 22, 1852–1858. [Google Scholar] [CrossRef] [PubMed]
- Matsumoto, K.; Watanabe, Y.; Yokoyama, S.I. Okara, soybean residue, prevents obesity in a diet-induced murine obesity model. Biosci. Biotechnol. Biochem. 2007, 71, 720–727. [Google Scholar] [CrossRef] [PubMed]
- Otieno, D.O.; Shah, N.P. Endogenous β-glucosidase and β-galactosidase activities from selected probiotic micro-organisms and their role in isoflavone biotransformation in soymilk. J. Appl. Microbiol. 2007, 103, 910–917. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suzuki, T.; Hara, H. Role of flavonoids in intestinal tight junction regulation. J. Nutr. Biochem. 2011, 22, 401–408. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.H.; Park, J.S.; Jung, J.W.; Byun, K.W.; Kang, K.S.; Lee, Y.S. Daidzein supplementation prevents non-alcoholic fatty liver disease through alternation of hepatic gene expression profiles and adipocyte metabolism. Int. J. Obes. 2011, 35, 1019. [Google Scholar] [CrossRef] [PubMed]
- Begley, M.; Gahan, C.G.; Hill, C. The interaction between bacteria and bile. FEMS Microbiol. Rev. 2005, 29, 625–651. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kakiyama, G.; Pandak, W.M.; Gillevet, P.M.; Hylemon, P.B.; Heuman, D.M.; Daita, K. Modulation of the fecal bile acid profile by gut microbiota in cirrhosis. J. Hepatol. 2013, 58, 949–955. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ridlon, J.M.; Kang, D.J.; Hylemon, P.B.; Bajaj, J.S. Bile acids and the gut microbiome. Curr. Opin. Gastroenterol. 2014, 30, 332. [Google Scholar] [CrossRef] [PubMed]
- Islam, K.S.; Fukiya, S.; Hagio, M.; Fujii, N.; Ishizuka, S.; Ooka, T. Bile acid is a host factor that regulates the composition of the cecal microbiota in rats. Gastroenterology 2011, 141, 1773–1781. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Hang, X.; Fan, X.; Li, D.; Yang, H. Characterization and selection of Lactobacillus strains for their effect on bile tolerance, taurocholate deconjugation and cholesterol removal. World J. Microbiol. Biotechnol. 2008, 24, 7–14. [Google Scholar] [CrossRef]
- Begley, M.; Hill, C.; Gahan, C.G. Bile salt hydrolase activity in probiotics. Appl. Environ. Microbiol. 2006, 72, 1729–1738. [Google Scholar] [CrossRef] [PubMed]
- Nagata, Y.; Ishiwaki, N.; Sugano, M. Studies on the mechanism of antihypercholesterolemic action of soy protein and soy protein-type amino acid mixtures in relation to the casein counterparts in rats. J. Nutr. 1982, 112, 1614–1625. [Google Scholar] [CrossRef] [PubMed]
- El-Gawad, I.A.A.; El-Sayed, E.M.; Hafez, S.A.; El-Zeini, H.M.; Saleh, F.A. The hypocholesterolaemic effect of milk yoghurt and soy-yoghurt containing bifidobacteria in rats fed on a cholesterol-enriched diet. Int. Dairy J. 2005, 15, 37–44. [Google Scholar] [CrossRef]
- Imaizumi, K.; Hirata, K.; Zommara, M.; Sugano, M.; Suzuki, Y. Effects of cultured milk products by Lactobacillus and Bifidobacterium species on the secretion of bile acids in hepatocytes and in rats. J. Nutr. Sci. Vitaminol. 1992, 38, 343–351. [Google Scholar] [CrossRef] [PubMed]
- Gudbrandsen, O.A.; Wergedahl, H.; Liaset, B.; Espe, M.; Berge, R.K. Dietary proteins with high isoflavone content or low methionine-glycine and lysine-arginine ratios are hypocholesterolaemic and lower the plasma homocysteine level in male Zucker fa/fa rats. Br. J. Nutr. 2005, 94, 321–330. [Google Scholar] [CrossRef] [PubMed]
- Ni, W.; Yoshida, S.; Tsuda, Y.; Nagao, K.; Sato, M.; Imaizumi, K. Ethanol-extracted soy protein isolate results in elevation of serum cholesterol in exogenously hypercholesterolemic rats. Lipids 1999, 34, 713–716. [Google Scholar] [CrossRef] [PubMed]
- Cao, Y.; Ma, Z.; Zhang, H.; Jin, Y.; Zhang, Y.; Hayford, F. Phytochemical properties and nutrigenomic implications of yacon as a potential source of prebiotic: Current evidence and future directions. Foods 2018, 7, 59. [Google Scholar] [CrossRef] [PubMed]
- Okarter, N.; Liu, R.H. Health benefits of whole grain phytochemicals. Crit. Rev. Food Sci. Nutr. 2010, 50, 193–208. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.Y.; Wu, S.J.; Fang, J.Y.; Wang, Y.P.; Shyu, Y.T. Cardiovascular and intestinal protection of cereal pastes fermented with lactic acid bacteria in hyperlipidemic hamsters. Food Res. Int. 2012, 48, 428–434. [Google Scholar] [CrossRef]
- Lazaridou, A.; Biliaderis, C.G. Molecular aspects of cereal β-glucan functionality: Physical properties, technological applications and physiological effects. J. Cereal Sci. 2007, 46, 101–118. [Google Scholar] [CrossRef]
- Ooi, L.G.; Liong, M.T. Cholesterol-lowering effects of probiotics and prebiotics: A review of in vivo and in vitro findings. Int. J. Mol. Sci. 2010, 11, 2499–2522. [Google Scholar] [CrossRef] [PubMed]
- Bäckhed, F.; Ley, R.E.; Sonnenburg, J.L.; Peterson, D.A.; Gordon, J.I. Host-bacterial mutualism in the human intestine. Science 2005, 307, 1915–1920. [Google Scholar] [CrossRef] [PubMed]
- Yusof, N.; Hamid, N.; Ma, Z.F.; Lawenko, R.M.; Mohammad, W.M.Z.W.; Collins, D.A.; Lee, Y.Y. Exposure to environmental microbiota explains persistent abdominal pain and irritable bowel syndrome after a major flood. Gut Pathog. 2017, 9, 75. [Google Scholar] [CrossRef] [PubMed]
- Sáez, C. Gut Microbiota May Improve Sugar Metabolism in Humans. 2016. Available online: http://www.gutmicrobiotaforhealth.com/en/gut-microbiota-may-improve-sugar-metabolism-in-humans/ (accessed on 11 February 2016).
- De Filippo, C.; Cavalieri, D.; Di Paola, M.; Ramazzotti, M.; Poullet, J.B.; Massart, S. Impact of diet in shaping gut microbiota revealed by a comparative study in children from Europe and rural Africa. Proc. Natl. Acad. Sci. USA 2010, 107, 14691–14696. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pan, X.D.; Chen, F.Q.; Wu, T.X.; Tang, H.G.; Zhao, Z.Y. Prebiotic oligosaccharides change the concentrations of short-chain fatty acids and the microbial population of mouse bowel. J. Zhejiang Univ. Sci. B 2009, 10, 258–263. [Google Scholar] [CrossRef] [PubMed] [Green Version]



{kind=link}
{kind=link}
{kind=link}
| Items/Groups | Control | SH | SHP | SHS | SHT | SHTL |
|---|---|---|---|---|---|---|
| Triglyceride (mg/dL) | 55.11 ± 20.0 bcd | 118.1 ± 35.8 a | 49.30 ± 8.52 cd | 71.50 ± 17.2 bc | 76.40 ± 24.7 b | 47.90 ± 9.95 d |
| Cholesterol-total (mg/dL) | 53.50 ± 6.86 c | 90.33 ± 11.1 a | 66.50 ± 13.4 bc | 79.67 ± 14.4 ab | 69.67 ± 14.4 bc | 65.50 ± 9.98 bc |
| HDL-cholesterol (mg/dL) | 40.56 ± 7.78 ab | 35.71 ± 4.59 b | 34.20 ± 6.16 b | 45.13 ± 10.3 a | 40.29 ± 4.08 ab | 40.14 ± 3.42 ab |
| Cholesterol/HDL-C | 1.41 ± 0.07 b | 2.12 ± 0.35 a | 2.04 ± 0.36 a | 2.02 ± 0.15 a | 1.94 ± 0.16 a | 2.01 ± 0.17 a |
| LDL-cholesterol (mg/dL) | 7.89 ± 2.23 c | 36.00 ± 8.68 a | 23.63 ± 7.20 b | 28.75 ± 9.77 b | 24.78 ± 6.29b | 25.00 ± 5.24 b |
| Free-fatty acid (mmol/L) | 1.43 ± 0.61 b | 2.31 ± 0.25 a | 1.16 ± 0.06 b | 1.55 ± 0.23 b | 1.36 ± 0.31 b | 1.41 ± 0.24 b |
| Glucose AC (mg/dL) | 100 ± 8.4 c | 199 ± 42.3 a | 125 ± 34.6 bc | 151 ± 25.5 b | 141 ± 24.8 b | 109 ± 17.3 c |
| HbA1C (%) | 4.02 ± 0.13 d | 6.96 ± 1.05 a | 5.17 ± 0.97 bc | 5.58 ± 1.42 b | 5.51 ± 1.25 b | 4.42 ± 0.32 cd |
| Insulin (ng/mL) | 2.48 ± 2.11 b | 9.99 ± 5.46 a | 1.61 ± 0.81 b | 2.11 ± 0.67 b | 2.61 ± 0.53 b | 1.65 ± 0.53 b |
| HOMA-IR | 0.55 ± 0.18 c | 4.46 ± 0.95 a | 0.54 ± 0.19 c | 0.89 ± 0.17 bc | 1.07 ± 0.36 b | 0.59 ± 0.16 c |
| Items/Groups | Control | SH | SHP | SHS | SHT | SHTL |
|---|---|---|---|---|---|---|
| Week 0 | 7.66 ± 0.04 a | 7.65 ± 0.09 a | 7.64 ± 0.01 a | 7.59 ± 0.05 a | 7.75 ± 0.08 ab | 7.67 ± 0.05 a |
| Week 4 | 8.91 ± 0.07 a | 8.09 ± 0.06 c | 7.71 ± 0.27 d | 8.04 ± 0.16 c | 8.31 ± 0.04 bc | 8.44 ± 0.05 b |
| Week | Items | Groups | |||||
|---|---|---|---|---|---|---|---|
| Control | SH | SHP | SHS | SHT | SHTL | ||
| 4 | Acetic acid_C2 | 4.16 ± 0.41 d | 5.21 ±0.11 c | 5.30 ± 0.29 c | 5.93 ± 0.31 c | 6.86 ± 0.28 b | 7.86 ±0.64 a |
| Propanoic acid_C3 | 0.55 ± 0.11 c | 0.70 ± 0.17 bc | 0.84 ± 0.19 abc | 1.01 ± 0.16 ab | 0.87 ± 0.07 ab | 1.13 ± 0.07 a | |
| Butyric acid_C4 | 0.51 ± 0.06 abc | 0.27 ± 0.02 c | 0.45 ± 0.06 bc | 0.70 ± 0.21 ab | 0.57 ± 0.28 abc | 0.83 ± 0.11 a | |
| Isobutyic acid_C4t | 0.00 ± 0.01 a | 0.04 ± 0.05 a | 0.02 ± 0.01 a | 0.01 ± 0.02 a | 0.04 ± 0.02 a | 0.05 ± 0.01 a | |
| Valeric acid_C5 | 0.03 ± 0.01 ab | 0.00 ± 0.00 b | 0.01 ± 0.02 b | 0.09 ± 0.07 ab | 0.05 ± 0.03 ab | 0.11 ± 0.04 a | |
| Isovaleric acid_C5t | 0.02 ± 0.01 a | 0.05 ± 0.06 a | 0.05 ± 0.01 a | 0.05 ± 0.01 a | 0.06 ± 0.03 a | 0.08 ± 0.02 a | |
| Caproic acid_C6 | 0.00 ± 0.00 a | 0.00 ± 0.00 a | 0.00 ± 0.00 a | 0.00 ± 0.00 a | 0.01 ± 0.01 a | 0.01 ± 0.02 a | |
| Week | Items | Groups | |||||
|---|---|---|---|---|---|---|---|
| Control | SH | SHP | SHS | SHT | SHTL | ||
| Week 0 | |||||||
| Feces weight (g) | 81.7 ± 1.75 | 81.0 ± 5.00 | 81.7 ± 3.06 | 80.7 ± 3.73 | 80.3 ± 4.30 | 81.7 ± 4.16 | |
| cholesterol content (mg/g) | 1.56 ± 0.31 | 1.47 ± 0.57 | 1.67 ± 0.16 | 1.66 ± 0.34 | 1.60 ± 0.26 | 1.43 ± 0.18 | |
| Bile acid content (µg/g) | 6.35 ± 0.51 | 6.22 ± 0.49 | 6.54 ± 0.41 | 6.84 ± 0.59 | 6.48 ± 0.35 | 6.56 ± 0.36 | |
| Triglyceride content (µg/g) | 57.52 ± 2.85 | 57.14 ± 3.48 | 58.12 ± 4.98 | 58.28 ± 2.78 | 57.04 ± 4.62 | 57.28 ± 2.14 | |
| Week 4 | |||||||
| Feces weight (g) | 81.7 ± 3.80 c | 79.7 ± 3.06 c | 83.0 ± 9.8 bc | 96.7 ± 4.16 ab | 100.0 ± 6.00 a | 104.0 ± 6.27 a | |
| cholesterol content (mg/g) | 4.90 ± 1.32 d | 27.5 ± 0.93 c | 29.2 ± 2.62 bc | 29.0 ± 3.13 bc | 32.1 ± 2.44 b | 35.6 ± 1.34 a | |
| Bile acid content (µg/g) | 4.63 ± 0.55 d | 176.4 ± 0.44 b | 247.7 ± 3.73 a | 115.9 ± 2.76 c | 173.0 ± 6.78 b | 248.2 ± 3.86 a | |
| Triglyceride content (µg/g) | 68.97 ± 1.76 a | 47.63 ± 3.45 c | 57.48 ± 2.01 b | 43.11 ± 0.24 c | 67.16 ± 3.15 a | 72.29 ± 8.87 a | |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huang, Y.-C.; Wu, B.-H.; Chu, Y.-L.; Chang, W.-C.; Wu, M.-C. Effects of Tempeh Fermentation with Lactobacillus plantarum and Rhizopus oligosporus on Streptozotocin-Induced Type II Diabetes Mellitus in Rats. Nutrients 2018, 10, 1143. https://doi.org/10.3390/nu10091143
Huang Y-C, Wu B-H, Chu Y-L, Chang W-C, Wu M-C. Effects of Tempeh Fermentation with Lactobacillus plantarum and Rhizopus oligosporus on Streptozotocin-Induced Type II Diabetes Mellitus in Rats. Nutrients. 2018; 10(9):1143. https://doi.org/10.3390/nu10091143
Chicago/Turabian StyleHuang, Ying-Che, Bo-Hua Wu, Yung-Lin Chu, Wen-Chang Chang, and Ming-Chang Wu. 2018. "Effects of Tempeh Fermentation with Lactobacillus plantarum and Rhizopus oligosporus on Streptozotocin-Induced Type II Diabetes Mellitus in Rats" Nutrients 10, no. 9: 1143. https://doi.org/10.3390/nu10091143