1. Introduction
Osteoporosis is a metabolic bone disease that increases fracture risk in the elderly and poses significant social and economic burden to the world’s population. The worldwide estimated osteoporosis-related fracture rate is around 40% in aged women and about 13% in men [
1]. Aging [
2] and estrogen deficiency [
3] are the two major leading causes of osteoporosis. The age-related changes in calcium homeostasis may contribute to the chronic depletion of calcium from bone [
4]. Indeed, the impaired intestinal calcium absorption during aging is believed to be the consequence of the age-related decline in renal 1,25(OH)
2D
3 production and intestinal vitamin D receptor (VDR) expression that lead to reduced 1,25(OH)
2D
3 sensitivity [
5]. The reduction of estrogen level during menopausal transition contributes to rapid bone loss in women, with an average decline of 10% bone mineral density (BMD) in the five years around menopause [
3]. The estrogen deficiency-induced bone loss in postmenopausal women is complicated by the age-induced abnormalities in the vitamin D endocrine system [
6]. Vitamin D deficiency-induced secondary hyperparathyroidism leads to an increase in the rate of bone loss [
2]. Calcium and vitamin D supplements are commonly used by adults for the management of bone health [
7]. However, concerns have been raised regarding the potential increase in the risk of cardiovascular events by the use of Ca supplements in healthy postmenopausal women [
8,
9]. In addition, the safety data concerning long term use of high-dose vitamin D3 supplementation in elderly, especially those with compromised renal function, is limited [
10]. Moreover, recent cohort studies and systematic reviews suggested a weak association between increased dietary intake of calcium or calcium and/or vitamin D supplements and a reduced fracture risk in the older population [
11,
12,
13,
14]. Thus, alternative approaches for the improvement of calcium balance and long-term management of bone health are needed.
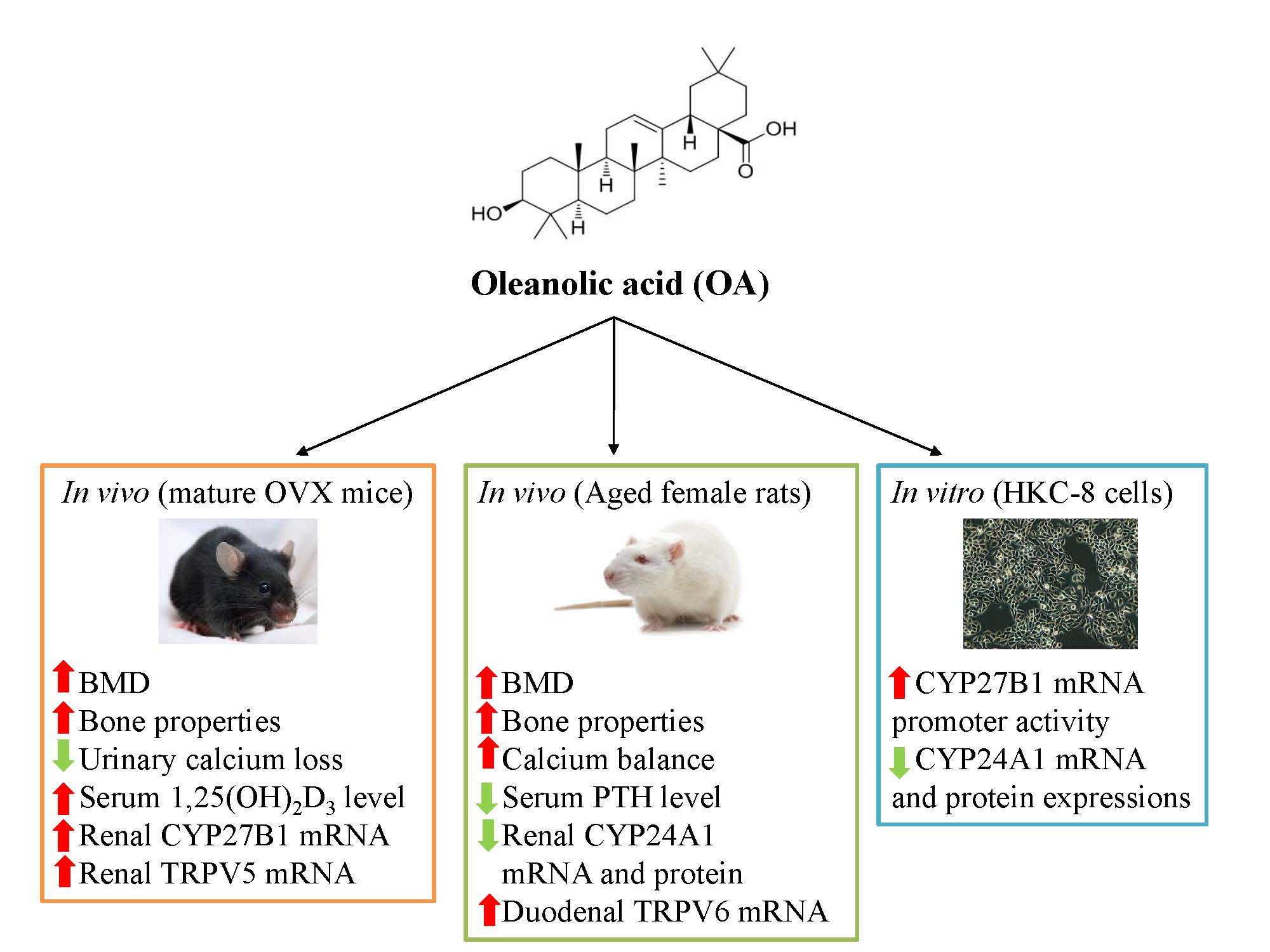
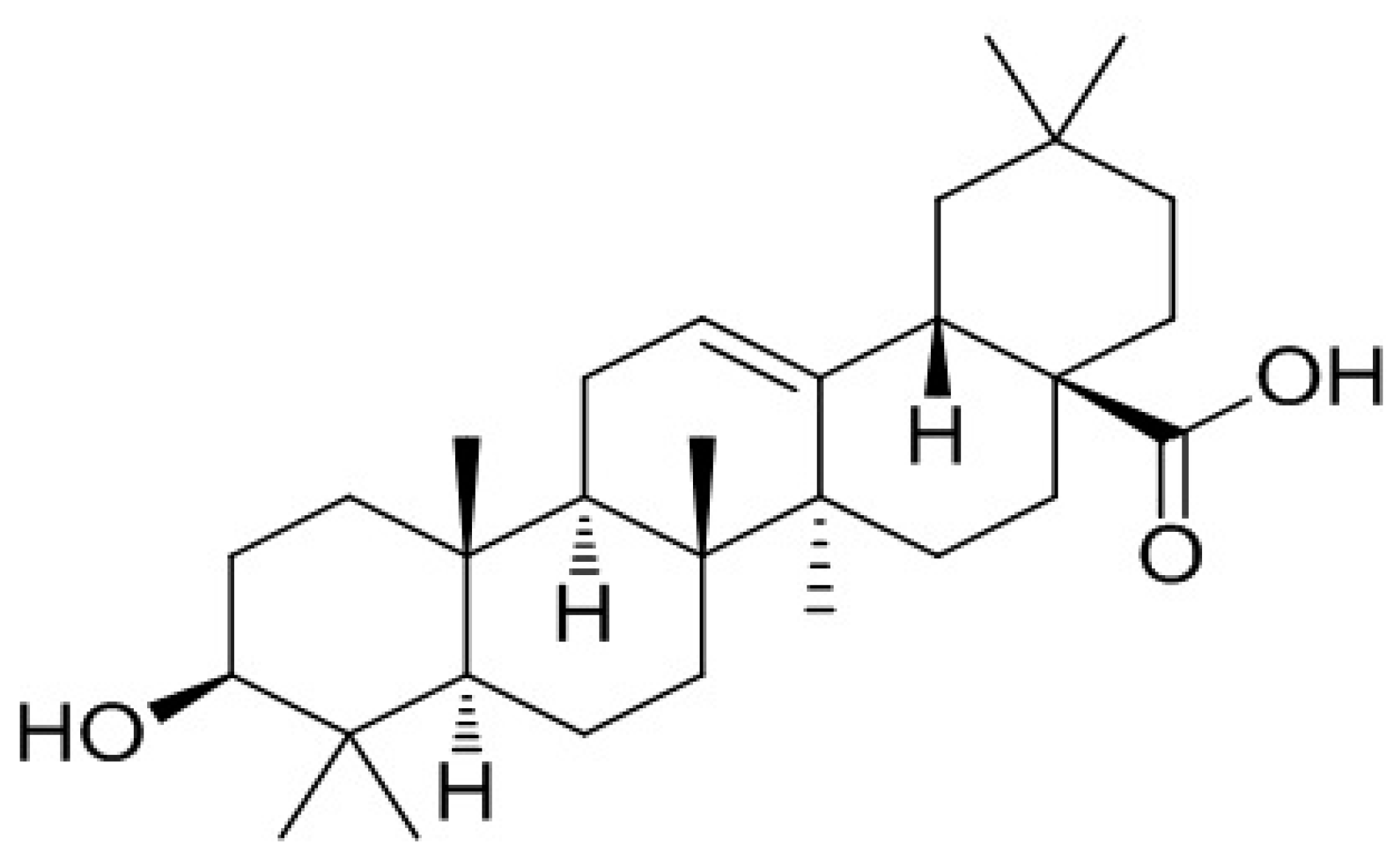
Oleanolic acid (3β-hydroxyolean-12-en-28-oic acid) (
Figure 1) is a bone protective pentacyclic triterpenoid compound [
15] that has been identified in over 1620 dietary plants and medicinal herbs [
16]. Oleanolic acid (OA) glucosides and its derivatives have been shown to reduce the formation of osteoclast-like multinucleated cells in a primary co-culture system [
17,
18,
19]. Oral administration of 0.1–10 mg/kg/day quinoxaline derivative of OA for 5 weeks significantly improved BMD in ovariectomised (OVX) mice [
20]. Similarly, oral feeding with 20 mg/kg/day OA for 3 months was shown to protect against bone loss in OVX rats, and OA was also shown to induce osteoblastic differentiation of bone mesenchymal stem cells (MSCs) [
21]. In addition, treatment of male mice with OA acetate for eight days effectively prevented lipopolysaccharide-induced inflammatory bone loss [
22]. Mechanistic studies demonstrated that the inhibitory effects of OA on osteoclastogeneis were mediated by the receptor activator of NF-κB ligand (RANKL) signaling pathway in RAW264.7 cells [
23], while the stimulatory effects of OA on human MSCs towards osteoprogenitor cells involved the inhibition of Notch signaling pathways [
24].
Our group was the first to report that
Fructus Ligustri Lucidi (FLL), an OA-rich Chinese herb, could regulate bone turnover markers and enhance calcium balance in mature OVX rats [
25]. Indeed, OA is being used as the authentication marker [
26] of FLL. Our subsequent studies showed that FLL significantly improved calcium balance and bone properties in both aged ovary-intact [
27] and aged OVX rats [
28,
29]. The improvement of bone properties in rats by FLL appeared to be associated with its actions on the vitamin D–parathyroid hormone (PTH) axis [
30,
31]. Recent studies further reported that FLL exerted stimulatory effects on bone and calcium balance in growing male and female rats [
32,
33]. These studies clearly show that FLL exerts bone protective effects through regulating calcium and vitamin D metabolism in growing, mature, and aged animal models. Thus, it is of interest to determine whether OA, as the major bioactive identified in FLL, could also exert its bone protective effects via modulation of the calcium–vitamin D axis.
1α,25-dihydroxyvitamin D
3 (1,25(OH)
2D
3) is essential for normal bone mineralization and calcium homeostasis in the body [
34]. The circulating concentration of 1,25(OH)
2D
3 is tightly controlled by the biosynthetic enzyme, 25-hydroxyvitamin D 1-α-hydroxylase (CYP27B1), as well as the catalytic enzyme, 25-hydroxyvitamin D 24-hydroxylase (CYP24A1) [
35]. CYP27B1 is primarily located in the proximal tubule of the nephron [
36] and is tightly regulated by many hormonal factors, including parathyroid hormone (PTH) [
37], 1,25(OH)
2D
3 itself [
38], fibroblast growth factor 23 (FGF23) [
39], as well as minerals, such as Ca and phosphate (P) ions [
40,
41]. CYP24A1 is induced by 1,25(OH)
2D
3 [
42] and FGF-23 [
43] and suppressed by PTH [
42]. A previous study showed that FLL could increase circulating 1,25(OH)
2D
3 by inducing renal CYP27B1 activity in rat renal proximal tubule cells [
30]. Thus, it is of interest to determine if OA could also increase circulating 1,25(OH)
2D
3 and alter vitamin D metabolism.
The present study was designed to test the hypothesis that OA could protect against estrogen deficiency- and age-induced bone loss via its actions involving the promotion of calcium absorption and regulation of vitamin D metabolism. The effects of OA on bone properties and calcium–vitamin D metabolism in both mature OVX mice and aged female rats as well as its effects on vitamin D metabolic enzymes in human proximal tubule (HKC-8) cells were determined. The results provide evidence of the osteoprotective effects of OA in OVX mice and aged rats.
2. Materials and Methods
2.1. Animal Study Design
2.1.1. Experiment 1: Bone Protective Effect of OA in OVX Mouse Model
Forty-two four-month-old C57BL/6J mice were purchased from The Chinese University of Hong Kong. Mice were randomly subjected to either sham-operated or ovariectomy (OVX) and were orally administrated by daily gavage for 6 weeks, as follows: sham + vehicle (Sham, n = 9), OVX + vehicle (OVX, n = 9), OVX + 17β-oestradiol (E2, 200 μg/kg/day, n = 8), OVX + OAL (low dose of OA, 50 mg/kg/day, n = 8), and OVX + OAH (high dose of OA, 100 mg/kg/day, n = 8). The animals were pair-fed with 3 g, the minimum daily average food intake, phytoestrogen-free AIN-93M rodent diet (Research diets, New Brunswick, NJ, USA). The 17β-Oestradiol (E2) was from Sigma–Aldrich (Sigma, St. Louis, MO, USA) and OA (purity > 98%) was from Shanghai Winherb Medical Technology Co. (Shanghai, China). The body weights of the animals were monitored on a weekly basis. One day before sacrifice, mice were individually housed in metabolic cages for urine collection. The mice were then sacrificed by cardiac stick exsanguinations under anaesthesia. Serum was prepared in aliquots and stored at −80 °C for biochemical measurements. The uterine index was calculated from the wet weight of uterus over the body weight. The left tibia and intact lumbar vertebra were collected, with soft tissue cleared, wrapped in saline-soaked gauze and stored at −20 °C for micro-computed tomography (μCT) analysis.
2.1.2. Experiment 2: Effect of OA on Bone and Calcium Balance in an Aged Female Rat Model
Twenty-four retired breeder Sprague–Dawley (SD) rats, aged nine-months-old, were purchased from Beijing Vital River Laboratories (Beijing, China). The retired breeders were raised to thirteen-months-old to establish the aged rat model. Before the treatment regimen, rats were pair-fed with a normal calcium diet (NCD, TD98005, 0.6% calcium, 0.65% phosphorus) for five days as acclimation. Aged female rats were randomly assigned to three groups (n = 8/group): control diets with oral administration of OA (25 mg/kg/day) or its vehicle by daily gavage, and a high calcium diet (HCD, 1.2% calcium, 0.65% phosphorus) served as a positive control group. All animals were pair-fed with 15 g/day of a control diet with water supply ad libitum for 12 weeks. Diets were purchased from Harlan Teklad (Madison, WI, USA). Before sacrifice, rats were individually placed into metabolic cages for 24 h urine and feces collection. Upon sacrifice, blood, duodenal mucosa, and cortex of the left kidney were collected. Serum in aliquots and tissue samples were stored at −80 °C for further analysis. The left tibia, femur, and the intact lumbar vertebra were collected with soft tissues cleaned, wrapped by phosphate buffered saline (PBS)-soaked gauze, and stored at −20 °C for micro-CT scanning and bone calcium content measurement.
All animals were housed in a room at 22 °C and provided with a 12 h light and dark cycle. All the experimental procedures were approved by the Animal Ethics Committee of The Hong Kong Polytechnic University (ASESC No.: 14-15/04-ABCT-R-STUDENT (mice study) and ASESC No.: 130602 (rat study)).
2.2. Biochemical Assays of Serum and Urine Samples
The concentrations of calcium and phosphorous in serum and urine samples were measured by standard colorimetric methods, using commercial kits, following the manufacturers’ instructions (STANBIO laboratory, Boerne, TX, USA). Mice serum osteocalcin (OCN) level was measured by a mouse osteocalcin ELISA kit (Alfa Aesar, Lancashire, UK). Mice urinary deoxypyridinoline (DPD) level was measured using a METRATM DPD EIA kit (Quidel Corporation, San Diego, CA, USA). Serum 1,25(OH)2D3 level was determined using a mouse 1,25-dihydroxyvitamin D3 (1,25(OH)2D3) ELISA kit (BlueGene Biotech, Shanghai, China) for OVX mice and a 1,25-(OH)2-Vitamin D ELISA Kit (Immundiagnostik AG, Bensheim, Germany) for aged rats. Rat serum PTH level was determined by Rat Bio Active Intact PTH ELISA Kit (Immutopics, Inc., San Clemente, CA, USA). Urinary calcium, phosphorous, and DPD were corrected by urinary creatinine (Cr) levels, which were determined by picric acid methods, using commercial kits (Zhongsheng Beikong Bio-technology and Science Inc., Beijing, China).
2.3. Microcomputed Tomography (μCT)
The left tibias, left femurs and fourth lumbar vertebrae (L4) of OVX mice and aged rats were scanned at voxel sizes of 10.5 μm3 and 21 μm3, respectively, by the μCT system (viva-CT40; Scanco Medical, Bassersdorf, Switzerland). Scans were performed at a medium resolution and using energy of 70 kVp, intensity of 114 μA, with an integration time of 300 ms. Distal femurs and proximal tibias were scanned in 100 slices for OVX mice and 210 slices for aged rats, from the metaphyseal growth plates. The volume of interest was contoured from 50 or 100 serials of slices (corresponding to a 0.525 mm region and 2.1 mm region), starting from the disappearance of the condyle, for evaluation. For the lumbar vertebra (L4), 100 slices for OVX mice and 150 slices centered in L4 for aged rats were scanned and evaluated. Morphometric parameters included bone mineral density (BMD, mg HA/cm3), bone volume/tissue volume (BV/TV, %), trabecular number (Tb.N, 1/mm), trabecular thickness (Tb.Th, mm), trabecular separation (Tb.Sp, mm), and connectivity density (Conn.D, 1/mm3) were calculated using a three-dimensional direct model with a constant threshold of 300.
2.4. Bone Calcium Content
The left tibia of both OVX mice and aged rats were first dried at 110 °C in muffle furnace for 12 h and then incinerated at 800 °C for 20 h. The ash weight was recorded. 100 mg of bone ash was weighed and dissolved in 2 mL of 37% HCl and diluted by Milli-Q water for atomization. The bone calcium content was determined by atomic absorption spectrophotometer (PerkinElmer, AAnalyst 100 Spectrometer, Norwalk, CT, USA).
2.5. Calcium Balance Study
The calcium content in 24 h fecal samples from aged rats was determined by the same method as for bone calcium, described previously [
25]. The calcium absorption rate and calcium balance were calculated using the following equations: Ca absorption rate (%) = (Ca intake − fecal Ca)/Ca intake × 100; Ca balance = Ca intake − (urinary Ca + fecal Ca).
2.6. Cell Culture Study
Human proximal kidney tubule cells (HKC-8) were a kind gift from Dr. Racusen of Johns Hopkins University [
44]. Cells were cultured in DMEM/F12 medium (Life Technologies, Carlsbad, CA, USA), supplemented with 1X penicillin/streptomycin and 5% Fetal Bovine Serum (FBS) (Life Technologies, Carlsbad, CA, USA). The cell culture was maintained on 100 mm culture dishes in the incubator, in a 5% CO
2–95% air atmosphere, at 37 °C. Forskolin (FSK, #F6886, purity ≥ 98%, Sigma–Aldrich, St. Louis, MO, USA) and 1,25(OH)
2D
3 (1,25D, #D1530, purity ≥ 99%, Sigma–Aldrich, St. Louis, MO, USA) were applied as positive controls for determining CYP27B1 and CYP24A1 expressions, respectively. For assessment of promoter activities, mRNA, and protein expressions, the medium was changed to a chemically–defined, serum-free medium containing the following supplements for 24 h before drug treatment: insulin (5 μg/mL), transferrin (5 μg/L), Na
2SeO
3 (5 ng/mL), tri-iodothyronine (0.37 nmol/L), epidermal growth factor (2.5 ng/mL), and hydrocortisone (1 nmol/L).
2.7. Real-Time Polymerase Chain Reaction (PCR) Analysis
RNA from animal tissues and HKC-8 cells was isolated using TRIzol
® Reagent (Invitrogen, Carlsbad, CA, USA). Total RNA (4 μg) was reverse-transcribed by M-MLV Reverse Transcriptase (Invitrogen, Carlsbad, California, USA) following the manufacturer’s instructions. Real-time PCR was performed in a total of 20 μL of reaction mixture, containing 10 μL of SYBR Green Mastermix (Applied Biosystems, Carlsbad, CA, USA) and 0.5 μL of cDNA product by 7900HT Fast Real-Time PCR System (Applied Biosystems, Carlsbad, CA, USA) using the two-step program: initial denaturation at 95 °C for 10 min, 40 cycles of denaturation at 95 °C for 15 s and 60 °C for 1 min. The sequences of primers for target genes and the housekeeping gene glyceraldehyde-3-phosphate dehydrogenase (GAPDH) are listed in
Table 1. The relative quantity of mRNA was calculated by fitting the Ct value to the standard curve using the SDS software package (Applied Biosystems, Carlsbad, CA, USA), and each gene expression was normalized using its own GAPDH expression level.
2.8. Western Blot Analysis
Cortexes of kidneys and HKC-8 cells were homogenized and lysed in Nonidet P-40 Lysis Buffer (20 mM Tris-HCl, pH 7.5; 150 mM NaCl, 1 mM MgCl2, 10% glycerol, 1% Nonidet P-40) supplemented with protease inhibitors: 1 mM PMSF, 2 μg/mL aprotinin, 2 μg/mL leupeptin, 10 mM NaF, and 1 mM sodium orthovanadate (Sigma, St. Louis, MO, USA). Protein concentrations were determined using the Bradford protein assay (Bio-Rad, Philadelphia, PA, USA). The proteins were separated by sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE) and transblotted to polyvinylidene difluoride (PVDF) membranes (Immobilin-P, Millipore Corp., Bedford, MA, USA) and probed with the primary antibodies: rabbit anti-CYP27B1 (1:500, Santa Cruz Biotechnology, Santa Cruz, CA, USA), rabbit anti-CYP27B1 (1:500, Santa Cruz Biotechnology, Santa Cruz, CA, USA), or mouse anti-β actin (1:5000, Abcam, Cambridge, MA, USA), followed by IgG-HRP-conjugated secondary antibodies anti-rabbit (1:2000, Santa Cruz Biotechnology, Santa Cruz, CA, USA) or anti-mouse (1:3000, Cell Signaling Technology, Beverly, MA, USA). Membranes were incubated with enhanced chemiluminescence (ECL) substrate (ClarityTM Western ECL Substrate, Bio-Rad, Philadelphia, PA, USA) for 5 min. The bound antibodies were visualized and quantified by Lumi-Imager (Roche, Manheim, Germany). The signal intensity of the bands was presented as Biochemical Light Units (BLU).
2.9. Transient Transfection
HKC-8 cells were transiently transfected with CYP27- or CYP24-promoter construct. The CYP27-promoter construct, a reporter plasmid containing a full-length insert of 1576 bp CYP27B1 promoter region, was a kind gift from Dr. Farzana Perwad from University of California, San Francisco [
45]. CYP24-promoter construct, a reporter plasmid containing a 300-bp vitamin D responsive region of the CYP24 promoter, was generated by the late Dr. Jack Omdahl [
46] and was kindly provided by Dr. JoEllen Welsh from University at Albany-SUNY [
47]. HKC-8 cells were co-transfected in serum-free medium with 0.4 μg of CYP27- or CYP24-promoter plasmid, together with 0.1 μg constitutive thymidine kinase promoter plasmid (pRL-TK, Promega, Madison, WI, USA) using FuGene HD transfection reagent (Promega). After 4 h of incubation, cells were treated with vehicle (0.1% ethanol), FSK (10
−5 M), 1,25D (10
−8 M) or OA (10
−8–10
−5 M) for 24 h. A dual luciferase reporter assay was conducted using reagents from Promega. The promoter activities of CYP27 or CYP24 were normalized to pRL-TK. Data were presented as relative luciferase units (RLU).
2.10. Statistical Analysis
All data obtained from the experiments were presented in the form of mean ± SEM (standard error of mean). The differences between groups of data were analyzed by GraphPad Prism Version 6.00 (GraphPad Software, La Jolla, CA, USA). The significances between different groups of means were evaluated by one-way analysis of variance (ANOVA). Post-test analysis of multiple comparisons was carried by Tukey’s test at a confidence level of 95%. p-values less than 0.05 were considered to be statistically significant.
4. Discussion
The present study demonstrates that OA could improve estrogen deficiency-induced and age-related bone loss and deterioration of bone properties in mature OVX mice and aged female rats, respectively. The positive action of OA on bone is partially associated with its suppression of urinary calcium loss and increase in serum 1,25(OH)2D3 levels in OVX mice. The improvement of bone health by OA in aged rats might involve its actions to increase calcium balance. Our results also indicated that OA bestows beneficial effects on bone health, calcium balance and vitamin D metabolism. The mechanism by which OA regulates vitamin D metabolism might be, at least in part, associated with its actions on vitamin D metabolic enzymes, in vivo.
OA increased BMD and improved bone microarchitecture at three bone sites (proximal tibia, distal femur and lumbar vertebra L4) in both OVX mice and aged rats. Our results agree with previous reports in which a quinoxaline derivative of OA was shown to increase the BMD in femurs of OVX mice [
20], and 3-months of treatment with OA increased BV/TV in OVX rats [
21]. An increase in connectivity density (a crucial structural property of cancellous bone) was shown to improve cancellous bone strength and reduce the risk of fractures by others [
48]. Thus, the significant increase in connectivity density at all three sites of OVX mice and aged rats by OA suggests that it might also strengthen cancellous bone. However, the increased BMD and bone microarchitecture may not result in a reduced fracture risk [
49,
50]. It would be of interest to further investigate whether OA increases biomechanical strength at different bone sites of relevant animal models, especially in a larger number of aged rats, in our future study.
OA behaves like E2, which exerts bone protective actions in an OVX model. E2 as well as OA, at 50 and 100 mg/kg/body weights, was shown to improve BMD and bone properties at three bone sites of OVX mice. However, unlike E2, OA did not suppress the OVX-induced increases in serum OCN and urinary DPD levels in mice. Thus, the bone protective effects exerted by OA in OVX mice might be different from E2 and mediated by distinct mechanisms of actions. In addition, OA increased BMD and improved bone properties in an aged rat model. Such protective actions of OA might be mediated by its reported bone anti-resorption activities [
17,
18,
19,
23] and/or the reported stimulatory effects on bone formation [
21,
24]. In addition, the bone protective actions of OA in vivo might also be mediated indirectly by other hormones, such as 1,25(OH)
2D
3 and PTH.
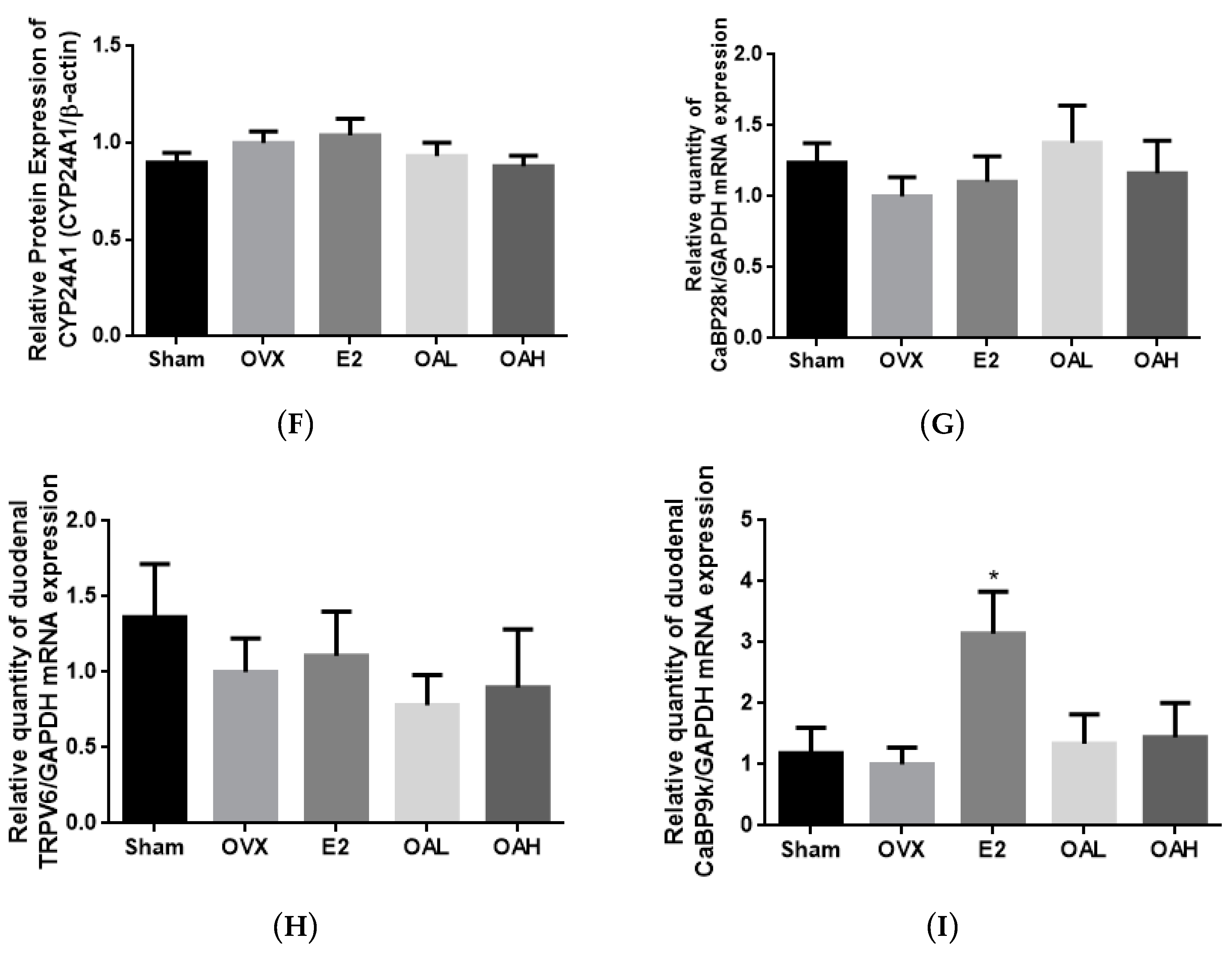
OA mimicked E2 in suppressing urinary calcium loss and inducing renal TRPV5 mRNA expression in OVX mice. However, renal TRPV5 expression could be independently induced by 1,25(OH)
2D
3 [
51] and estrogen [
52] in OVX mice. Since serum 1,25(OH)
2D
3 levels were also increased in OVX mice, it is unclear if the induction of renal TRPV5 mRNA by OA is mediated via its E2-like actions or via its actions on 1,25(OH)
2D
3. Indeed, unlike E2, OA did not increase duodenal CaBP9K mRNA in OVX mice, suggesting that its actions on Ca transport protein expression might be distinct from estrogen. Using an
in silico target identification tool [
53], OA was predicted to interact with estrogen receptors (ER), among the hundreds of predicted targets for osteoporosis [
54]. However, the results of the competitive receptor binding assay from our laboratory indicated that OA did not bind to estrogen receptor α (ERα) or β (ERβ) (
Supplementary Materials Figure S1), suggesting that OA did not activate classical estrogen signaling pathway. Thus, the actions of OA in improving calcium re-absorption might be mediated by non-classical estrogen signaling pathways or indirectly via an increase in serum 1,25(OH)
2D
3 level.
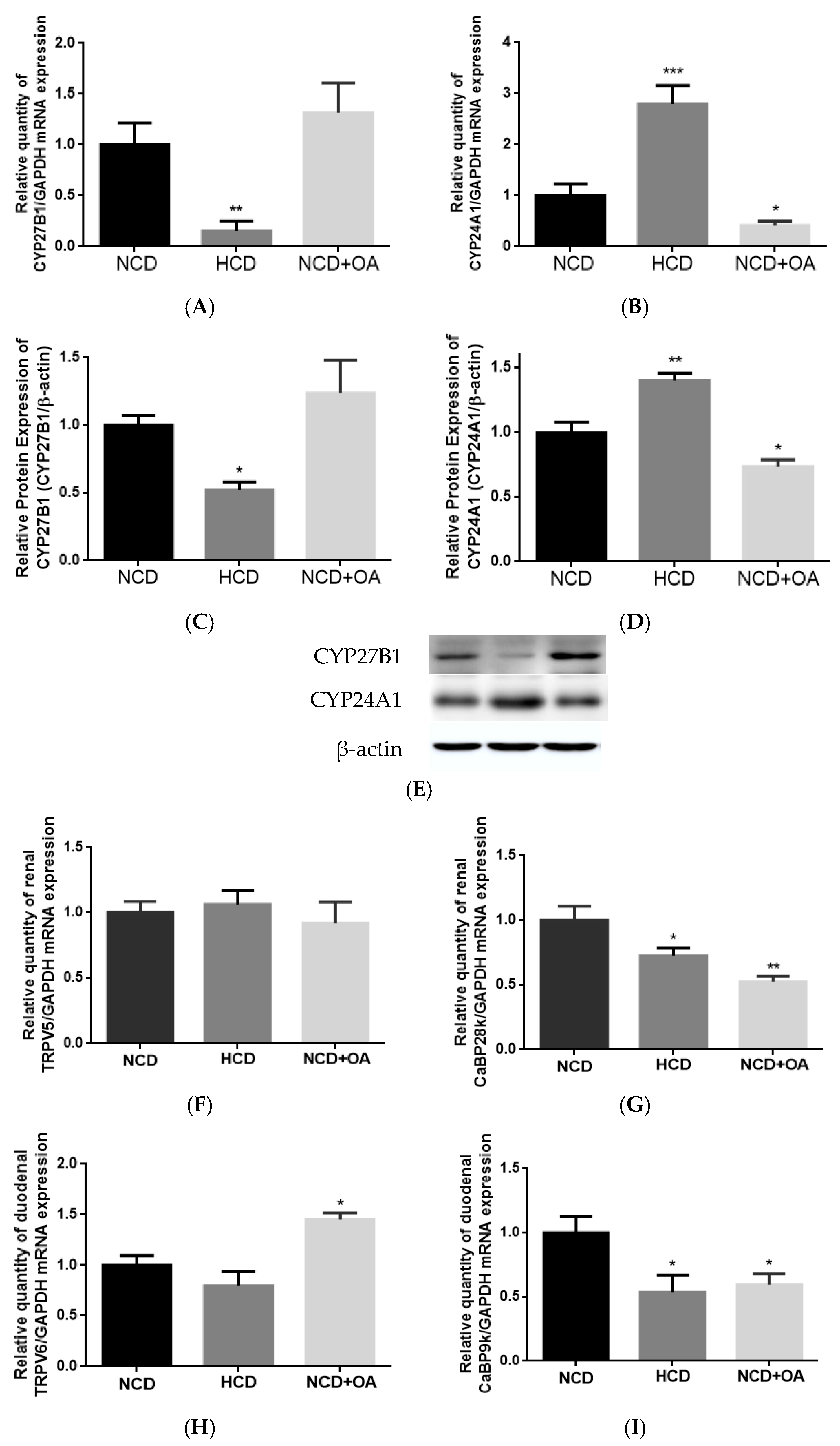
OA mimicked HCD in improving calcium balance in aged female rats. The improvement in Ca balance by OA was associated with an increase in duodenal TRPV6 mRNA expression in aged female rats. Thus, apart from the direct effects on bone, our results indicated that OA might protect bone through enhancing calcium balance in aged rats, which, in turn, via its actions in regard to increasing calcium entry across brush border membranes [
34]. However, duodenal CaBP9K and renal CaBP28K mRNA expression in aged female rats was unexpectedly suppressed by OA in a similar action to that of HCD. The decrease in duodenal CaBP9K and renal CaBP28K by HCD was likely due to the decrease in serum 1,25(OH)
2D
3 in aged rats with excess dietary Ca intake [
55]. However, as OA did not alter serum 1,25(OH)
2D
3 in aged rats, a vitamin D-independent mechanism might be involved in this action. As the activities and protein expression of the calcium transport proteins were not been measured in the present study, future studies will be needed to characterize the effects of OA as well as its mechanism of actions involved in regulation of intestinal and renal Ca transport in aged rats.
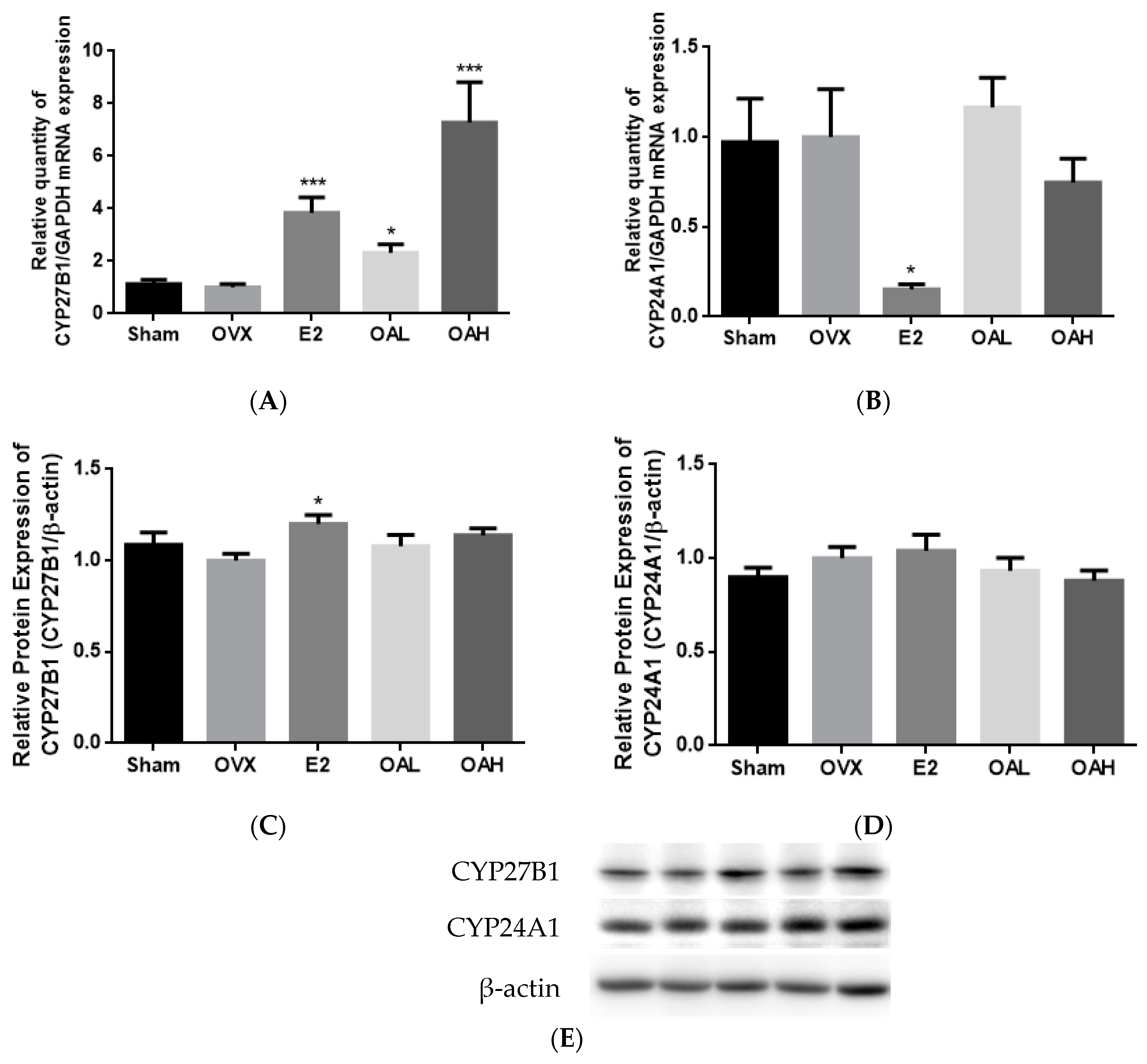
Our results are the first to report that OA at 100 mg/kg/day could significantly increase serum 1,25(OH)
2D
3 levels and renal CYP27B1 mRNA expression in OVX mice following treatment for 6 weeks. The actions of OA on vitamin D metabolism in OVX mice appeared to be different from those of E2. The present study as well as others [
56,
57] showed that E2 did not raise serum 1,25(OH)
2D
3 levels in OVX mice. In addition, E2 could induce CYP27B1 and suppress CYP24A1 expression in our study using mice and in other studies using avian species [
58] models. The mechanisms involved in the alteration of vitamin D metabolism by OA in OVX mice remains to be determined. In contrast, OA at 25 mg/kg/day did not alter serum 1,25(OH)
2D
3 levels nor renal CYP27B1 expression in aged female rats following treatment for 3 months. This observation agrees with those reported for aged rats in which the induction of renal CYP27B1 by stimuli, such as PTH, were blunted [
59]. In contrast, OA could significantly suppress renal CYP24A1 mRNA and protein expressions in aged rats. In fact, renal CYP24A1 appeared to be more sensitive to the stimuli in aged rats than renal CYP27B1 [
2]. Thus, it is possible that OA could alter renal vitamin D metabolism in aged rats via its actions on renal CYP24A1 expression. Recent studies suggest that extra-renal CYP27B1 plays an important role in paracrine and autocrine control of 1,25(OH)
2D
3 production [
60,
61]. As OA significantly improved bone properties and induced duodenum TRPV6 protein expression without altering serum 1,25(OH)
2D
3 levels, future study will be needed to study the mechanism of OA on regulating the local production of 1,25(OH)
2D
3 in bone and duodenum in aged rats.
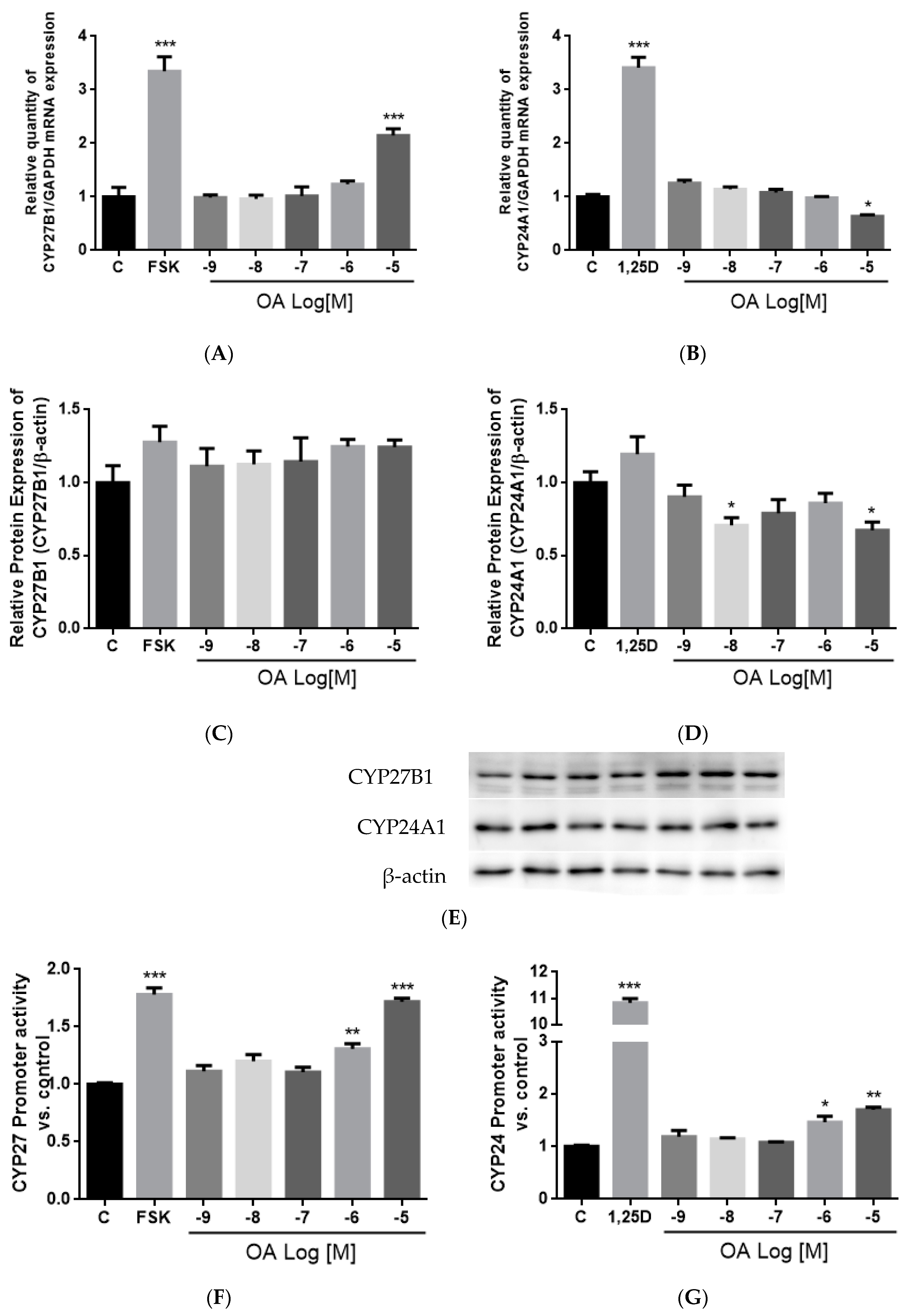
OA significantly induced CYP27B1 and mRNA and stimulated CYP27B1 promoter activity in a dose-dependent manner, suggesting that OA regulated CYP27B1 expression transcriptionally. The induction of OA on CYP27B1 mRNA and promoter activity were of similar levels as those induced by PTH and forskolin in HKC-8, as reported by previous studies [
62,
63]. In addition, OA suppressed mRNA and protein expressions of CYP24A1 in HKC-8 cells, and this result agrees with the action of OA in aged rats. However, a high dose of OA unexpectedly increased CYP24A1 promoter activity in HKC-8 cells. Such unexpected CYP24A1 regulation was reported for the actions of PTH in porcine proximal tubule cell AOK-B50 cells transfected with a full length CYP24A1 promoter [
64]. In that study, PTH was shown to work synergistically with 1,25(OH)
2D
3 to further induce, rather than inhibit, CYP24A1 promoter activity. The discrepancies between the responses of endogenous CYP24A1 expression and the transfected CYP24A1 promoter activities could be accounted for by the composition of the promoter region in the plasmid constructs. Indeed, the promoter employed in the present study is a partial promoter with a 5′-flanking region and partial exon 1 of CYP24A1 promoter which contains a vitamin D receptor (VDR) responsive region [
46,
47], and a more distal promoter sequence or 3′ untranslated region of the CYP24A1 promoter might be required for the inhibitory effects of OA.
One limitation of our study was that we only measured the renal CYP27B1 and CYP24A1 expression levels and mRNA expressions of the Ca-transporter in the intestine and kidney. Although we observed beneficial effects of OA on bone, Ca balance, and modulatory effects on vitamin D metabolism, the molecular targets and mechanisms involved in OA regulating Ca and vitamin D metabolism remain unclear. Further studies will be needed to investigate if OA could alter local production of 1,25(OH)2D3 via its actions on extra-renal CYP27B1 in bone and duodenum in both animal models. Future studies of the actions of OA on intestinal Ca absorption in OVX animal models as well as the proteins and activities of intestinal and renal Ca-transporters would have provided more evidence on the mechanisms involved.

 ,
, 






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}