Categorizing Wetland Vegetation by Airborne Laser Scanning on Lake Balaton and Kis-Balaton, Hungary

Abstract
:
1. Introduction
1.1. The Conservation Status of Shore Wetlands
1.2. Objective
2. State of the Art
2.1. Passive Remote Sensing of Wetland Vegetation
2.2. Airborne Laser scanning as a Method for Vegetation Surveys
2.3. Wetland Vegetation Mapping Based on ALS as a Standalone Tool
2.4. Wetland Vegetation Mapping Based on the Fusion of ALS-Derived Data with Other Data
2.5. Enhancing the Information Contained in ALS Point Datasets
3 Data and Methods
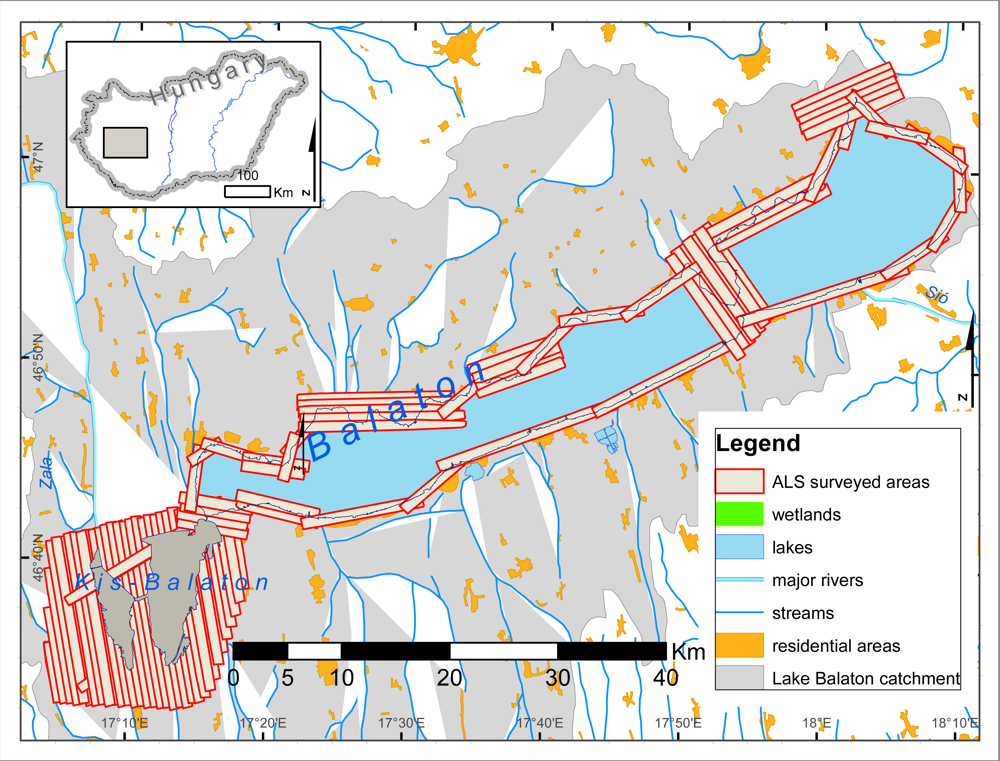
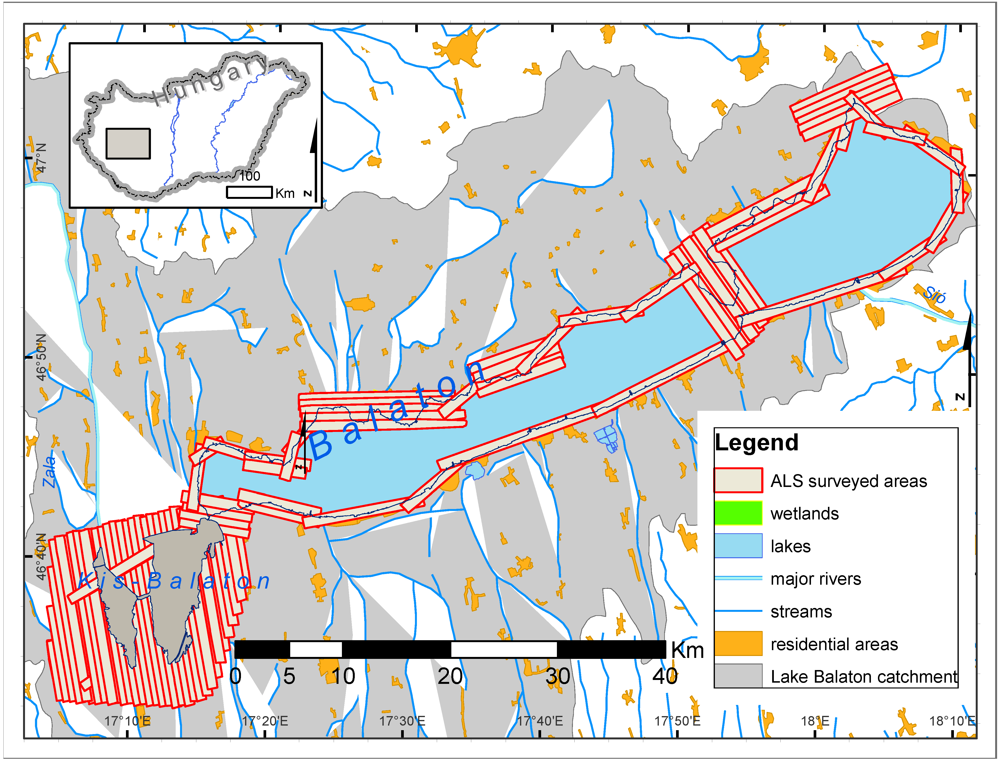
3.1. Study Area
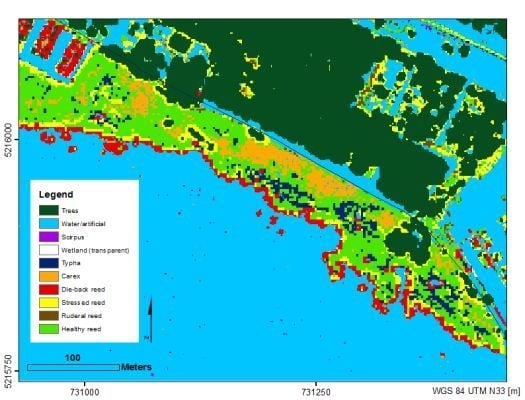
3.2. Categories Used for Vegetation Classification
3.3. Data and Processing
3.3.1. Airborne Laser Scanning Data
3.3.2. Ground Truth Data
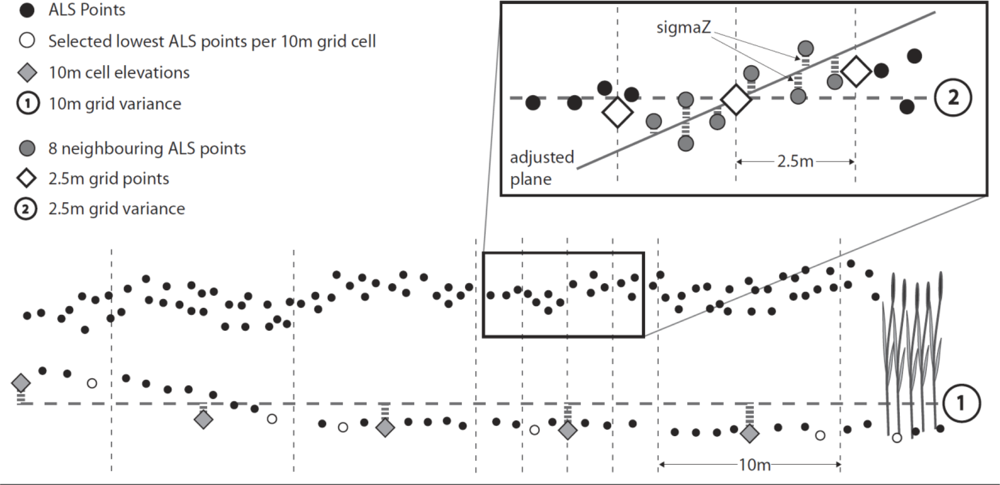
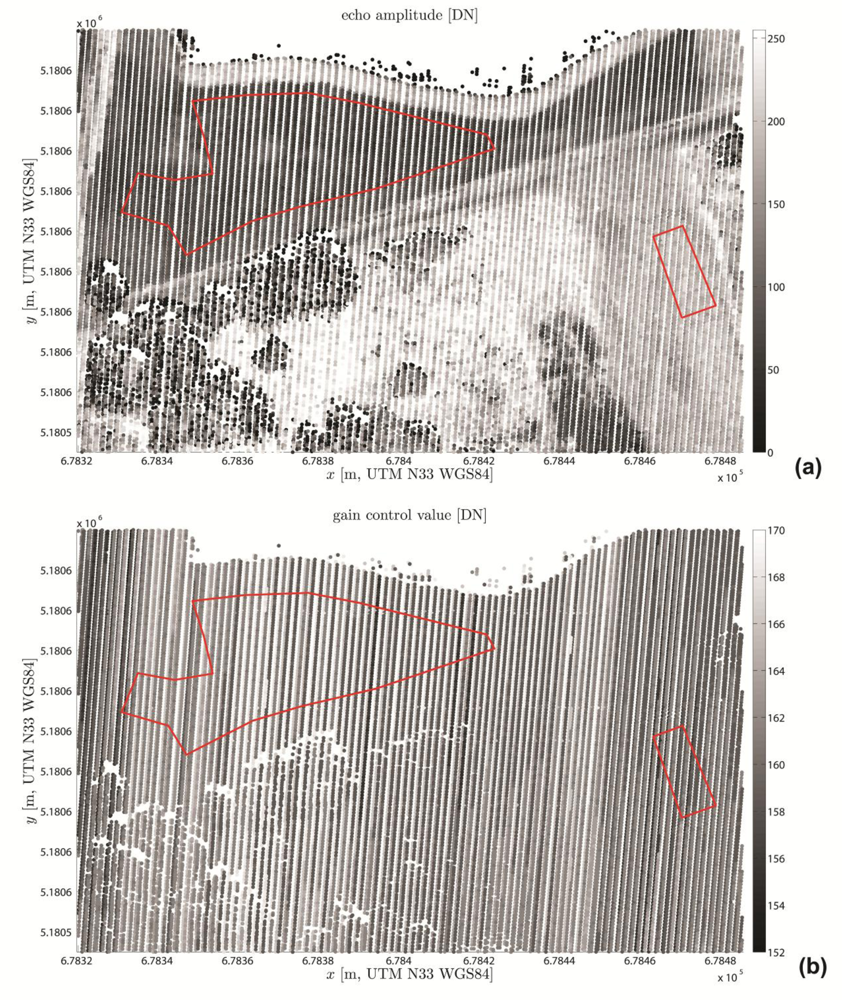
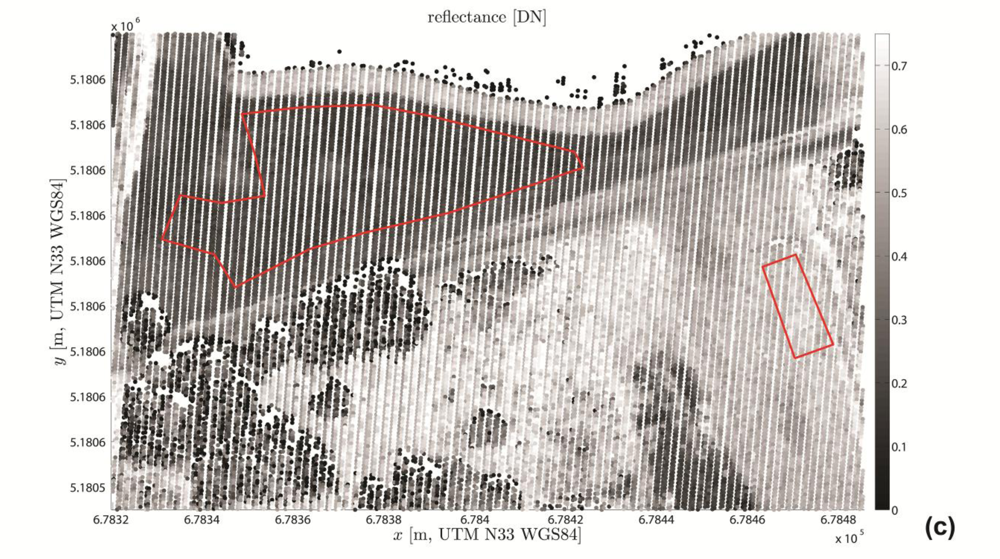
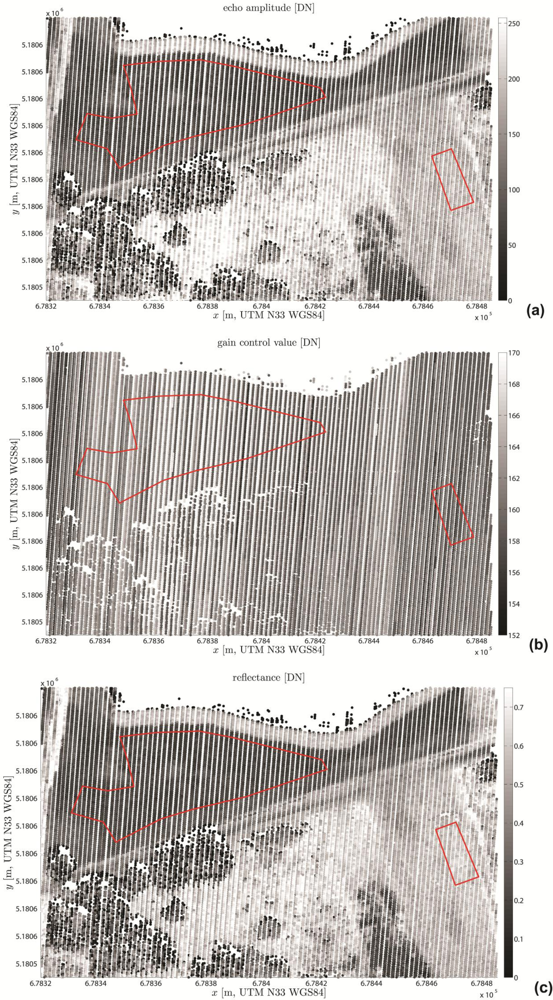
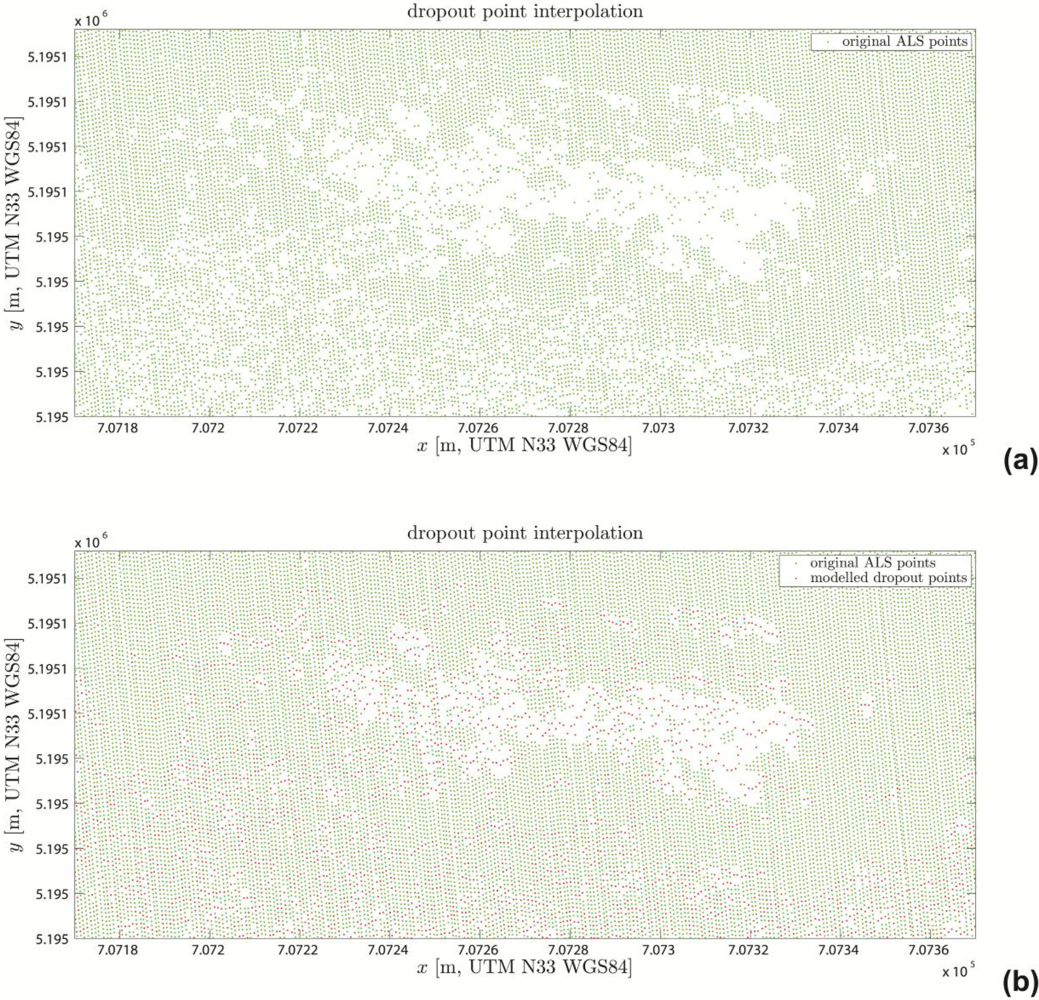
3.3.3. Visualization and Quality Control
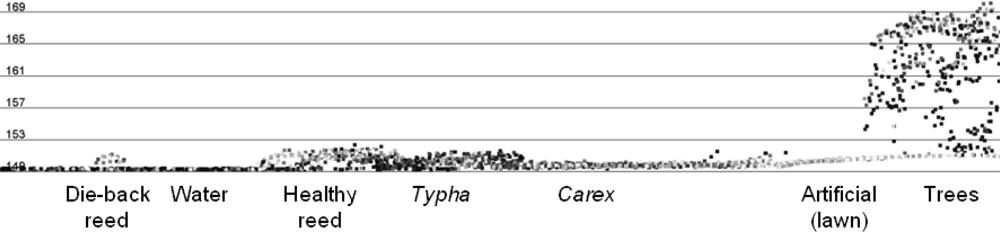
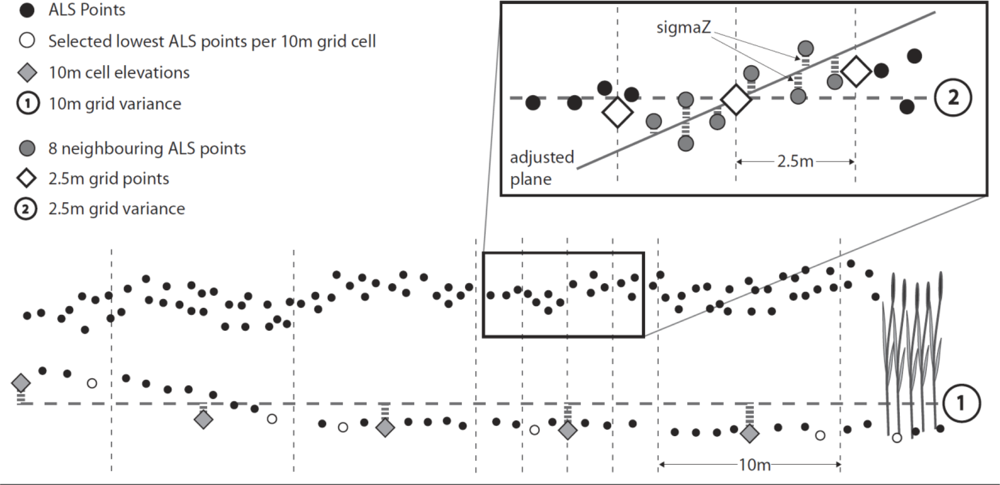
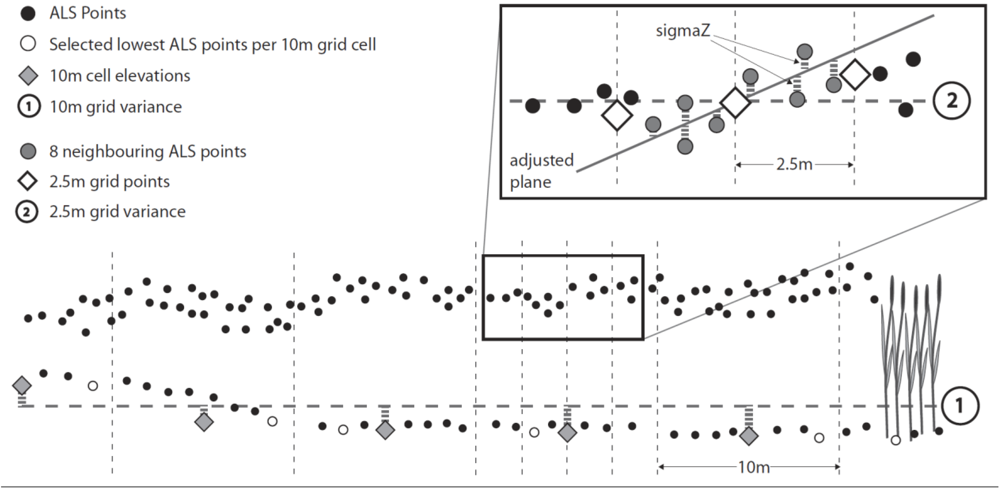
3.3.4. Input Parameters and Calculations
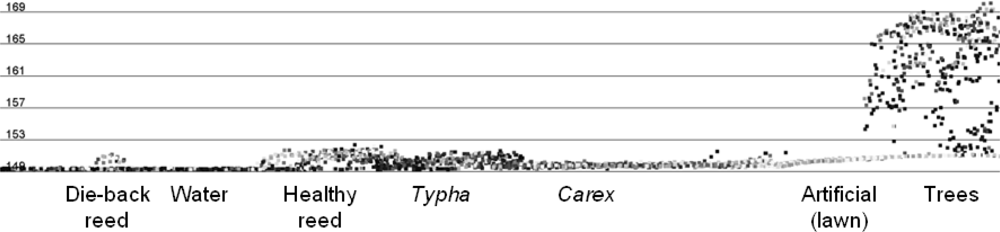
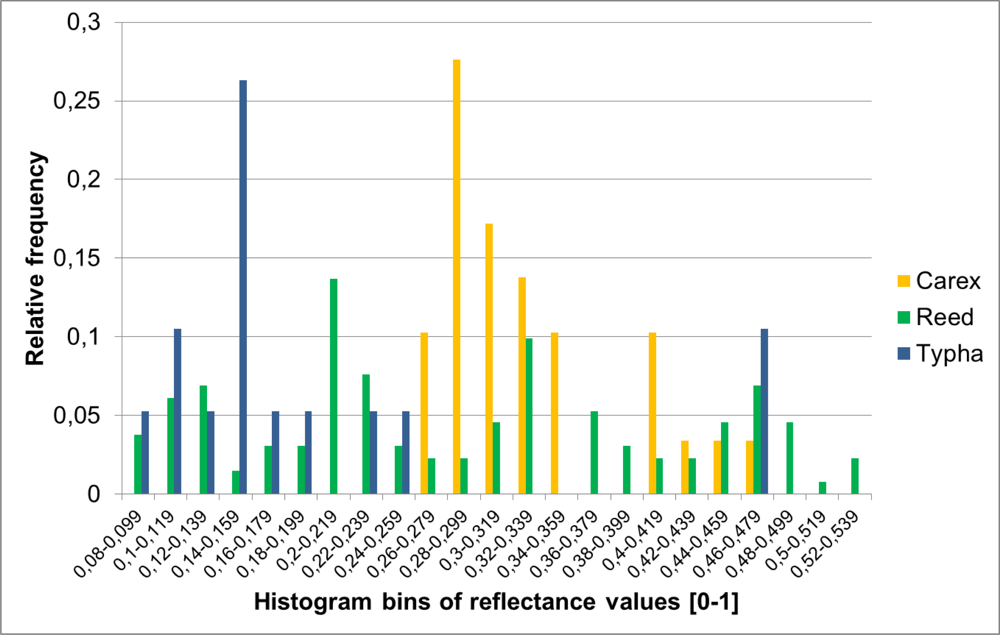
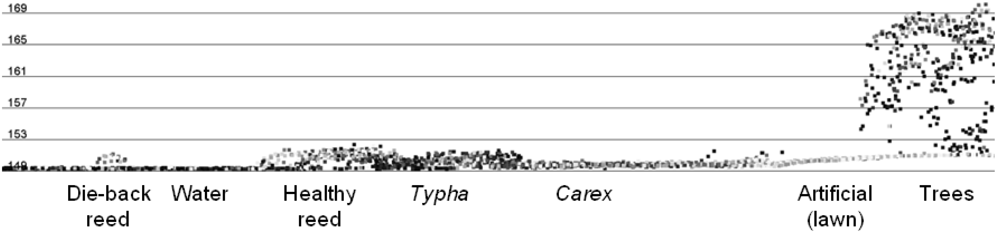
3.3.5. Signature Analysis
3.3.6. Classification Algorithms Applied for Wetland Masking and Classification (Table 1)
3.4. Validation
4. Results
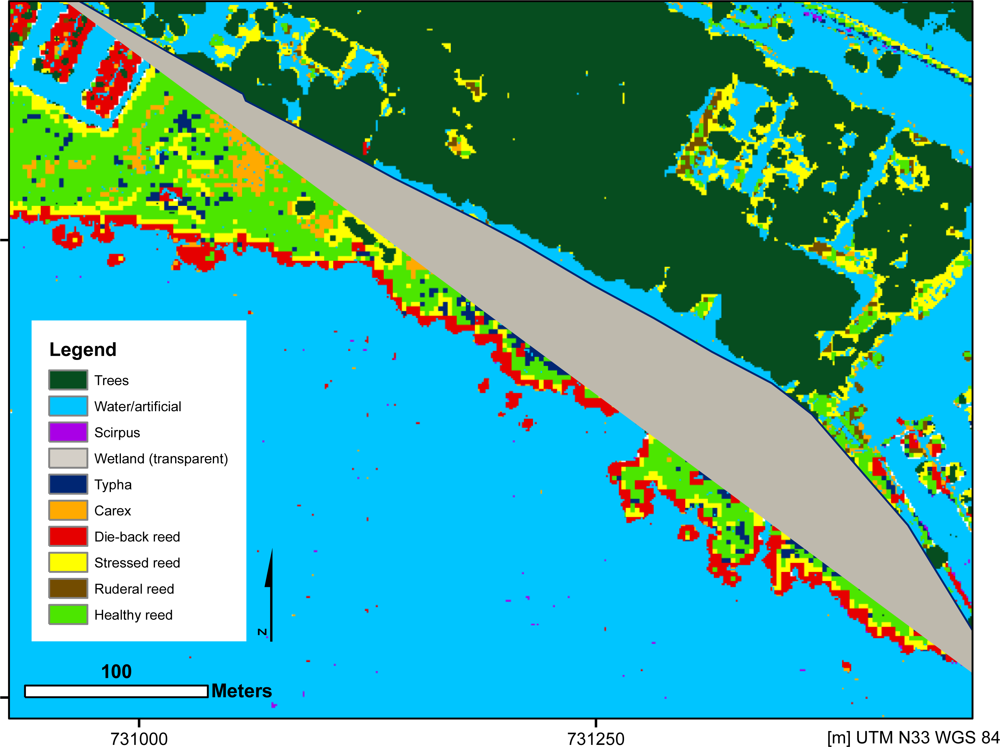
4.1. Visual Quality Control Results
4.2. Numeric Quality Control (Tables 2, 3)
5. Discussion
5.1. Discussion of the Survey Flight
5.2. Discussion of the Processing Methodology
5.2.1. Parameter Calculation and Algorithm
5.2.2. Selection of Categories
5.2.3. Accuracy of Classification Categories
5.3. Discussion of Quality Control Method
5.4. Comparing Classification Accuracy with Other Studies
5.4.1. Multispectral and Hyperspectral Surveys
5.4.2. Combined ALS-Multispectral Surveys
5.4.3. ALS-Based Vegetation Surveys
5.5. Applicability of the New Method for Regional and Local Scale Wetland Vegetation Mapping
6. Conclusions
Acknowledgments
References
- Strayer, D.L.; Findlay, S.E.G. Ecology of freshwater shore zones. Aquat. Sci 2010, 72, 127–163. [Google Scholar]
- Ostendorp, W. Schilf ALS Lebensraum. In Artenschutzsymposium Teichrohrsänger; Landesanstalt für Umweltschutz Baden-Württemberg: Karlsruhe, Germany, 1993; Volume 68, pp. 173–280. [Google Scholar]
- Vymazal, J. Enhancing ecosystem services on the landscape with created, constructed and restored wetlands. Ecol. Eng 2011, 37, 1–5. [Google Scholar]
- Schmieder, K. European lake shores in danger—Concepts for a sustainable development. Limnologica 2004, 34, 3–14. [Google Scholar]
- Wetzel, R.G. Limnology, 3rd ed; Academic Press: London, UK, 2001; p. 1066. [Google Scholar]
- Segal, S. Principles on structure, zonation and succession of aquatic macrophytes. Hidrobiologia 1971, 12, 89–97. [Google Scholar]
- van der Putten, W.H. Die-back of Phragmites australis in European wetlands: An overview of the European Research Programme on Reed Die-Back and Progression (1993–1994). Aquat. Bot 1997, 59, 263–275. [Google Scholar]
- Cizkova, H.; Brix, H.; Kopecky, J.; Lukavska, J. Organic acids in the sediments of wetlands dominated by Phragmites australis: Evidence of phytotoxic concentrations. Aquat. Bot 1999, 64, 303–315. [Google Scholar]
- Weisner, S.E.B. Effects of an organic sediment on performance of young Phragmites australis clones at different water depth treatments. Hydrobiologia 1996, 330, 189–194. [Google Scholar]
- Zlinszky, A. A Balatoni Nádpusztulás Légifelvételes Vizsgálata; Eötvös Loránd University: Budapest, Hungary, 2007. [Google Scholar]
- Fogli, S.; Marchesini, R.; Gerdol, R. Reed (Phragmites australis) decline in a brackish wetland in Italy. Mar. Environ. Res 2002, 53, 465–479. [Google Scholar]
- Kovács, M.; Turcsányi, G.; Tuba, Z.; Wolcsánszky, S.E.; Vásárhelyi, T.; Dely-Draskovics, Á.; Tóth, S.; Koltay, A.; Kaszab, L.; Szőke, P.; et al. The Decay of Reed in Hungarian Lakes. In Conservation and Management of Lakes; Salánki, J., Herodek, S., Eds.; Akadémiai Kiadó: Budapest, Hungary, 1989; pp. 461–470. [Google Scholar]
- Hunter, P.D.; Gilvear, D.J.; Tyler, A.N.; Willby, N.J.; Kelly, A. Mapping macrophytic vegetation in shallow lakes using the Compact Airborne Spectrographic Imager (CASI). Aquat. Conserv 2010, 20, 717–727. [Google Scholar]
- van der Putten, W.H. Assessing Ecological Change in European Wetlands: How to Know What Parameters Should Be Monitored to Evaluate the Die-Back of Common Reed (Phragmites Australis). In Monitoring of Ecological Change in Wetlands of Middle Europe; Aubrecht, G., Dick, G., Prentice, C., Eds.; Botanischer Arbeitsgemeinschaft am Oberösterreichisen Landesmuseum: Linz, Austria, 1993; pp. 61–68. [Google Scholar]
- Ostendorp, W. Dieback of reeds in Europe—A critical review of literature. Aquat. Bot 1989, 35, 5–26. [Google Scholar]
- Hortobágyi, T.; Simon, T. Növényföldrajz, Társulástan, Ökológia, 3 ed.; Nemzeti Tankönyvkiadó Rt: Budapest, Hungary, 1981. [Google Scholar]
- Kelly, M.; Tuxen, K.A.; Stralberg, D. Mapping changes to vegetation pattern in a restoring wetland: Finding pattern metrics that are consistent across spatial scale and time. Ecol. Indic 2011, 11, 263–273. [Google Scholar]
- Weilhoefer, C.L. A review of indicators of estuarine tidal wetland condition. Ecol. Indic 2011, 11, 514–525. [Google Scholar]
- Alberdi, I.; Condes, S.; Martinez-Millan, J. Review of monitoring and assessing ground vegetation biodiversity in national forest inventories. Environ. Monit. Assess 2010, 164, 649–676. [Google Scholar]
- Rücker, A.; Grosser, S.; Melzer, A. Geschichte und Ursachen des Röhrichtrückgangs am Ammersee (Deutschland). Limnologica 1999, 29, 11–20. [Google Scholar]
- Hellsten, S.; Keto, A.; Suoramiemi, M.; Partanen, S. Long-term changes in the aquatic vegetation of Lake Päijänne, Southern Finland. Verh. Internat. Verein Limnol 2006, 29, 1167–1173. [Google Scholar]
- Csaplovics, E. Interpretation von Farbinfrarotbildern, Kartierung von Vegetationsschäden in Brixlegg, Schilfkartierung Neusiedler See. Ph.D. Thesis, Technische Universität Wien, Vienna, Austria. 1982; 23, 1–192. [Google Scholar]
- Whyte, R.S.; Trexel-Kroll, D.; Klarer, D.M.; Shields, R.; Francko, D.A. The invasion and spread of Phragmites australis during a period of low water in a Lake Erie coastal wetland. J. Coastal Res 2008, 55, 111–120. [Google Scholar]
- Dienst, M.; Schmieder, K.; Ostendorp, W. Effects of water level variations on the dynamics of the reed belts of Lake Constance. Limnologica 2004, 34, 29–36. [Google Scholar]
- Laba, M.; Downs, R.; Smith, S.; Welsh, S.; Neider, C.; White, S.; Richmond, M.; Philpot, W.; Baveye, P. Mapping invasive wetland plants in the Hudson River National Estuarine Research Reserve using Quickbird satellite imagery. Remote Sens. Environ 2008, 112, 286–300. [Google Scholar]
- Belluco, E.; Camuffo, M.; Ferrari, S.; Modenese, L.; Silvestri, S.; Marani, A.; Marani, M. Mapping salt-marsh vegetation by multispectral and hyperspectral remote sensing. Remote Sens. Environ 2006, 105, 54–67. [Google Scholar]
- Pengra, B.W.; Johnston, C.A.; Loveland, T.R. Mapping an invasive plant, Phragmites australis, in coastal wetlands using the EO-1 Hyperion hyperspectral sensor. Remote Sens. Environ 2007, 108, 74–81. [Google Scholar]
- Yang, X.J. Remote sensing and GIS applications for estuarine ecosystem analysis: An overview. Int. J. Remote Sens 2005, 26, 5347–5356. [Google Scholar]
- Burai, P.; Lövei, G.Z.; Lénárt, C.; Nagy, I.; Enyedi, P. Mapping aquatic vegetation of the Rakamaz-Tiszanagyfalui Nagy-morotva using Hyperspectral Imagery. Acta Geographica Debrecina Landscape and Environment 2010, 4, 1–10. [Google Scholar]
- Artigas, F.; Pechmann, I.C. Balloon imagery verification of remotely sensed Phragmites australis expansion in an urban estuary of New Jersey, USA. Landscape Urban Plan 2010, 95, 105–112. [Google Scholar]
- Siciliano, D.; Wasson, K.; Potts, D.C.; Olsen, R.C. Evaluating hyperspectral imaging of wetland vegetation as a tool for detecting estuarine nutrient enrichment. Remote Sens. Environ 2008, 112, 4020–4033. [Google Scholar] [Green Version]
- Schmidt, K.S.; Skidmore, A.K. Spectral discrimination of vegetation types in a coastal wetland. Remote Sens. Environ 2003, 85, 92–108. [Google Scholar]
- Wehr, A.; Lohr, U. Airborne laser scanning—An introduction and overview. ISPRS J. Photogramm 1999, 54, 68–82. [Google Scholar]
- Kobler, A.; Pfeifer, N.; Ogrinc, P.; Todorovski, L.; Ostir, K.; Dzeroski, S. Repetitive interpolation: A robust algorithm for DTM generation from aerial laser scanner data in forested terrain. Remote Sens. Environ 2007, 108, 9–23. [Google Scholar]
- Kraus, K.; Pfeifer, N. Determination of terrain models in wooded areas with airborne laser scanner data. ISPRS J. Photogramm 1998, 53, 193–203. [Google Scholar]
- Fricker, G.A.; Saatchi, S.S.; Meyer, V.; Gillespie, T.W.; Sheng, Y. Application of semi-automated filter to improve waveform lidar sub-canopy elevation model. Remote Sens 2012, 4, 1494–1518. [Google Scholar]
- Jochem, A.; Hollaus, M.; Rutzinger, M.; Höfle, B. Estimation of aboveground biomass in Alpine forests: A semi-empirical approach considering canopy transparency derived from airborne lidar data. Sensors 2011, 11, 278–295. [Google Scholar]
- Eysn, L.; Hollaus, M.; Schadauer, K.; Pfeifer, N. Forest delineation based on airborne LIDAR data. Remote Sens 2012, 4, 762–783. [Google Scholar]
- Lindberg, E.; Hollaus, M. Comparison of methods for estimation of stem volume, stem number and basal area from Airborne Laser Scanning data in a hemi-boreal forest. Remote Sens 2012, 4, 1004–1023. [Google Scholar]
- Garcia, M.; Riano, D.; Chuvieco, E.; Salas, J.; Danson, F.M. Multispectral and LiDAR data fusion for fuel type mapping using Support Vector Machine and decision rules. Remote Sens. Environ 2011, 115, 1369–1379. [Google Scholar]
- Riano, D.; Chuvieco, E.; Ustin, S.L.; Salas, J.; Rodriguez-Perez, J.R.; Ribeiro, L.M.; Viegas, D.X.; Moreno, J.M.; Fernandez, H. Estimation of shrub height for fuel-type mapping combining airborne LiDAR and simultaneous color infrared ortho imaging. Int. J. Wildland Fire 2007, 16, 341–348. [Google Scholar]
- Mitchell, J.J.; Glenn, N.F.; Sankey, T.T.; Derryberry, D.R.; Anderson, M.O.; Hruska, R.C. Small-footprint Lidar estimations of sagebrush canopy characteristics. Photogramm. Eng. Remote Sensing 2011, 77, 521–530. [Google Scholar]
- Streutker, D.R.; Glenn, N.F. LiDAR measurement of sagebrush steppe vegetation heights. Remote Sens. Environ 2006, 102, 135–145. [Google Scholar]
- Sankey, T.T.; Bond, P. LiDAR-based classification of sagebrush community types. Rangeland Ecol. Manag 2011, 64, 92–98. [Google Scholar]
- Nayegandhi, A.; Brock, J.C.; Wright, C.W.; O’Connell, M.J. Evaluating a small footprint, waveform-resolving lidar over coastal vegetation communities. Photogramm. Eng. Remote Sensing 2006, 72, 1407–1417. [Google Scholar]
- Mücke, W.; Hollaus, M.; Briese, C. Application and Analysis of Airborne Laser Scanning Data on Reed Beds. In International Symposium on Advanced Methods of Monitoring Reed Habitats in Europe; Csaplovics, E., Schimdt, J., Eds.; Rhombos-Verlag: Berlin, Germany, 2010. [Google Scholar]
- Cobby, D.M.; Mason, D.C.; Horritt, M.S.; Bates, P.D. Two-dimensional hydraulic flood modelling using a finite-element mesh decomposed according to vegetation and topographic features derived from airborne scanning laser altimetry. Hydrol. Process 2003, 17, 1979–2000. [Google Scholar]
- Straatsma, M.W.; Baptist, M. Floodplain roughness parameterization using airborne laser scanning and spectral remote sensing. Remote Sens. Environ 2008, 112, 1062–1080. [Google Scholar]
- Johansen, K.; Tiede, D.; Blaschke, T.; Arroyo, L.A.; Phinn, S. Automatic geographic object based mapping of streambed and riparian zone extent from LiDAR data in a temperate rural urban environment, Australia. Remote Sens 2011, 3, 1139–1156. [Google Scholar]
- Rosso, P.H.; Ustin, S.L.; Hastings, A. Use of lidar to study changes associated with Spartina invasion in San Francisco Bay marshes. Remote Sens. Environ 2006, 100, 295–306. [Google Scholar]
- Collin, A.; Long, B.; Archambault, P. Salt-marsh characterization, zonation assessment and mapping through a dual-wavelength LiDAR. Remote Sens. Environ 2010, 114, 520–530. [Google Scholar]
- Knight, J.M.; Dale, P.E.R.; Spencer, J.; Griffin, L. Exploring LiDAR data for mapping the micro-topography and tidal hydro-dynamics of mangrove systems: An example from southeast Queensland, Australia. Estuar. Coast. Shelf Sci 2009, 85, 593–600. [Google Scholar]
- Jenkins, R.B.; Frazier, P.S. High-resolution remote sensing of upland swamp boundaries and vegetation for baseline mapping and monitoring. Wetlands 2010, 30, 531–540. [Google Scholar]
- Morris, J.T.; Porter, D.; Neet, M.; Noble, P.A.; Schmidt, L.; Lapine, L.A.; Jensen, J.R. Integrating LIDAR elevation data, multi-spectral imagery and neural network modelling for marsh characterization. Int. J. Remote Sens 2005, 26, 5221–5234. [Google Scholar]
- Gilvear, D.; Tyler, A.; Davids, C. Detection of estuarine and tidal river hydromorphology using hyper-spectral and LiDAR data: Forth estuary, Scotland. Estuar. Coast. Shelf Sci 2004, 61, 379–392. [Google Scholar]
- Mather, P.M. Computer Processing of Remotely Sensed Images; John Wiley & Sons Ltd: Chichester, UK, 2006; p. 324. [Google Scholar]
- Onojeghuo, A.O.; Blackburn, G.A. Optimising the use of hyperspectral and LiDAR data for mapping reedbed habitats. Remote Sens. Environ 2011, 115, 2025–2034. [Google Scholar]
- Geerling, G.W.; Labrador-Garcia, M.; Clevers, J.; Ragas, A.M.J.; Smits, A.J.M. Classification of floodplain vegetation by data fusion of spectral (CASI) and LiDAR data. Int. J. Remote Sens 2007, 28, 4263–4284. [Google Scholar]
- Arroyo, L.A.; Johansen, K.; Armston, J.; Phinn, S. Integration of LiDAR and QuickBird imagery for mapping riparian biophysical parameters and land cover types in Australian tropical savannas. For. Ecol. Manage 2010, 259, 598–606. [Google Scholar]
- Höfle, B.; Pfeifer, N. Correction of laser scanning intensity data: Data and model-driven approaches. ISPRS J. Photogramm 2007, 62, 415–433. [Google Scholar]
- Lehner, H.; Briese, C. Radiometric calibration of full-waveform airborne laser scanning data based on natural surfaces. Int. Arch. Photogramm. Remote Sens. Spat. Inf. Sci 2010, 38, 360–365. [Google Scholar]
- Habib, A.F.; Kersting, A.P.; Shaker, A.; Yan, W.Y. Geometric calibration and radiometric correction of LiDAR data and their impact on the quality of derived products. Sensors 2011, 11, 9069–9097. [Google Scholar]
- Höfle, B.; Vetter, M.; Pfeifer, N.; Mandlburger, G.; Stotter, J. Water surface mapping from airborne laser scanning using signal intensity and elevation data. Earth Surf. Process. Landf 2009, 34, 1635–1649. [Google Scholar]
- Moffiet, T.; Mengersen, K.; Witte, C.; King, R.; Denham, R. Airborne laser scanning: Exploratory data analysis indicates potential variables for classification of individual trees or forest stands according to species. ISPRS J. Photogramm 2005, 59, 289–309. [Google Scholar]
- Brandtberg, T. Classifying individual tree species under leaf-off and leaf-on conditions using airborne lidar. ISPRS J. Photogramm 2007, 61, 325–340. [Google Scholar]
- Chust, G.; Galparsoro, I.; Borja, A.; Franco, J.; Uriarte, A. Coastal and estuarine habitat mapping, using LIDAR height and intensity and multi-spectral imagery. Estuar. Coast. Shelf Sci 2008, 78, 633–643. [Google Scholar]
- Brzank, A.; Heipke, C.; Goepfert, J.; Soergel, U. Aspects of generating precise digital terrain models in the Wadden Sea from lidar-water classification and structure line extraction. ISPRS J. Photogramm 2008, 63, 510–528. [Google Scholar]
- Virág, Á. A Balaton Múltja és Jelene, 1st ed.; Egri nyomda: Eger, Hungary, 1998; p. 904. [Google Scholar]
- Dömötörfy, Z.; Reeder, D.; Pomogyi, P. Changes in the macro-vegetation of the Kis-Balaton Wetlands over the last two centuries: a GIS perspective. Hydrobiologia 2003, 506, 671–679. [Google Scholar]
- Kárpáti, I.; Szeglet, P.; Tóth, I. Die Vegetationskarte der Bozsauer-Bucht. BFB-Bericht 1987, 63, 63–68. [Google Scholar]
- Kárpáti, I.; Szeglet, P.; Kárpáti, V. Vegetationskarte des Naturschutzgebietes Kisbalaton und seine Primärproduktion. BFB-Bericht 1983, 47, 183–196. [Google Scholar]
- Lang, M.W.; McCarty, G.W. LIDAR intensity for improved detection of inundation below the forest canopy. Wetlands 2009, 29, 1166–1178. [Google Scholar]
- Clevering, O.A.; Lissner, J. Taxonomy, chromosome numbers, clonal diversity and population dynamics of Phragmites australis. Aquat. Bot 1999, 64, 185–208. [Google Scholar]
- Engloner, A.I. Structure, growth dynamics and biomass of reed (Phragmites australis)—A review. Flora 2009, 204, 331–346. [Google Scholar]
- Poulin, B.; Lefebvre, G.; Mauchamp, A. Habitat requirements of passerines and reedbed management in southern France. Biol. Conserv 2002, 107, 315–325. [Google Scholar]
- Zlinszky, A.; Tóth, V.; Pomogyi, P.; Timár, G. Initial report of the AIMWETLAB project: Simultaneous airborne hyperspectral, LIDAR and photogrammetric survey of the full shoreline of Lake Balaton, Hungary. Geographia Technica 2011, 11, 101–117. [Google Scholar]
- Mandlburger, G.; Otepka, J.; Karel, W.; Wagner, W.; Pfeifer, N. Orientation and processing of airborne laser scanning data (OPALS)—Concept and first results of a comprehensive ALS software. Int. Arch. Photogramm. Remote Sens. Spat. Inf. Sci 2009, 38, 55–60. [Google Scholar]
- Wagner, W.; Ullrich, A.; Ducic, V.; Melzer, T.; Studnicka, N. Gaussian decomposition and calibration of a novel small-footprint full-waveform digitising airborne laser scanner. ISPRS J. Photogramm 2006, 60, 100–112. [Google Scholar]
- Wagner, W.; Hollaus, M.; Briese, C.; Ducic, V. 3D vegetation mapping using small-footprint full-waveform airborne laser scanners. Int. J. Remote Sens 2008, 29, 1433–1452. [Google Scholar]
- Lehner, H.; Kager, H.; Roncat, A.; Zlinszky, A. Consideration of Laser Pulse Fluctuations and Automatic Gain Control in Radiometric Calibration of Airborne Laser Scanning Data. Proceedings of 6th ISPRS Student Consortium and WG VI/5 Summer School, Fayetteville State University, NC, USA, 30 July–6 August 2011.
- Hollaus, M.; Mandlburger, G.; Pfeifer, N.; Mücke, W. Land cover dependent derivation of digital surface models from airborne laser scanning data. Int. Arch. Photogramm. Remote Sens. Spat. Inf. Sci 2010, 38, 222–226. [Google Scholar]
- Lillesand, T.M.; Kiefer, R.W.; Chipman, J.W. Remote Sensing and Image Interpretation: International Edition, 5th ed.; John Wiley & Sons: New York, NY, USA, 2003; p. 704. [Google Scholar]
- Ressl, C.; Pfeifer, N.; Mandlburger, G. Applying 3D Affine Transformation and Least Squares Matching for Airborne Laser Scanning Strips Adjustment without GNSS/IMU Trajectory Data. Proceedings of ISPRS Workshop Laser Scanning 2011, Calgary, AB, Canada, 29–31 August 2011.
- Briese, C.; Höfle, B.; Lehner, H.; Wagner, W.; Pfenningbauer, M.; Ullrich, A. Calibration of full-waveform airborne laser scanning data for object classification. Proc. SPIE 2008, 6950, 6950H. [Google Scholar]
- Stevens, J.P.; Blackstock, T.H.; Howe, E.A.; Stevens, D.P. Repeatability of Phase I habitat survey. J. Environ. Manage 2004, 73, 53–59. [Google Scholar]
- Vanden Borre, J. User Requirements towards the Integration of Remote Sensing in Natura 2000 Monitoring. Results of the Work Package 2200; Belgian Science Policy: Brussels, Belgium, 2009. [Google Scholar]
- Gibbs, J.P. Importance of small wetlands for the persistance of local-populations of wetland-associated animals. Wetlands 1993, 13, 25–31. [Google Scholar]
- Standovár, T.; Primack, R. A Természetvédelmi Biológia Alapjai, 1st ed.; Nemzeti Tankönyvkiadó Rt: Budapest, Hungary, 2001; p. 541. [Google Scholar]
- Wang, C.; Menenti, M.; Stoll, M.P.; Belluco, E.; Marani, M. Mapping mixed vegetation communities in salt marshes using airborne spectral data. Remote Sens. Environ 2007, 107, 559–570. [Google Scholar]
- Pal, M.; Mather, P.M. An assessment of the effectiveness of decision tree methods for land cover classification. Remote Sens. Environ 2003, 86, 554–565. [Google Scholar]
- Verrelst, J.; Geerling, G.W.; Sykora, K.V.; Clevers, J. Mapping of aggregated floodplain plant communities using image fusion of CASI and LiDAR data. Int. J. Appl. Earth Obs. Geoinf 2009, 11, 83–94. [Google Scholar]
- Holmgren, J.; Persson, A.; Soderman, U. Species identification of individual trees by combining high resolution LIDAR data with multi-spectral images. Int. J. Remote Sens 2008, 29, 1537–1552. [Google Scholar]
- Hartfield, K.A.; Landau, K.I.; van Leeuwen, W.J.D. Fusion of high resolution aerial multispectral and LiDAR data: land cover in the context of urban mosquito habitat. Remote Sens 2011, 3, 2364–2383. [Google Scholar]
- Hollaus, M.; Mücke, W.; Höfle, B.; Dorigo, W.; Pfeifer, N.; Wagner, W.; Bauerhansl, C.; Regner, B. Tree Species Classification Based on Full-Waveform Airborne Laser Scanning Data. Proceedings of Silvilaser 2009, College Station, TX, USA, 14–16 October 2009.
- Surveying and Mapping Authority of the Republic of Slovenia. In Technical Documentation: Laser Scanning and Aerial Photography; Ministry of the Environment and Spatial Planning: Ljubljana, Slovenia, 2010; p. 23.
- Flood, M. Laser altimetry: From science to commercial lidar mapping. Photogramm. Eng. Remote Sensing 2001, 67, 1209–1211. [Google Scholar]
- Briese, C.; Mandlburger, G.; Pfeifer, N. Airborne Laser Scanning—High Quality Digital Terrain Modelling. Proceedings of Geo-Sibir, III International Scientific Congress, Novosibirsk, Russia, 23–27 April 2007; pp. 79–92.
- Fritzmann, P.; Hoefle, B.; Vetter, M.; Sailer, R.; Stoetter, J.; Bollmann, E. Surface classification based on multi-temporal airborne LiDAR intensity data in high mountain environments: A case study from Hintereisferner, Austria. Zeitschrift Fur Geomorphologie 2011, 55, 105–126. [Google Scholar]
- Oberleitner, I.; Dick, G. Feuchtgebietsinventar Österreich-Grundlagenerhebung, 1st ed.; Umweltbundesamt: Vienna, Austria, 1996; pp. 1–41. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Wetland Vegetation Classification | Variables | Logical Operator | |||
| Category | Reflectance [0−1] | NDSM Height [m] | Dropount Point Count | Grid Variance [m] | |
| Carex | >0.22 | <0.95 | <0.04 | AND | |
| die-back reed | >0.07 and <0.34 | >3 | AND | ||
| Typha | <0.155 | <0.3 | AND | ||
| ruderal reed | >0.4 | <0.2 | AND | ||
| stressed reed | >0.2 | AND | |||
| healthy reed | <0.4 | <0.2 | AND | ||
| input raster cell size [m] | 2.5 | 2.5 | 2.5 | 1 | |
| output raster cell size [m] | 2.5 | ||||
| Wetland/Non-wetland Identification | Variables | Logical operator | |||
| Category | Reflectance [0−1] | NDSM Height [m] | DTM Variance (“Slope”) [m] | Sigma Z [m] | |
| Scirpus | >0.02 and <0.07 | >0.28 and <0.6 | >0.01 and <0.1 | AND | |
| tree | >3.5 | >3 | >1 | OR | |
| water/artificial | <0.045 or >0.55 | <0.4 | >0.9 | <0.02 | OR |
| wetland | >0.4 | <0.9 | >0.02 and <1 | AND | |
| input raster cell size [m] | 2.5 | 2.5 | 10 | 1 | |
| output raster cell size [m] | 1 | ||||
| Reference Field Photographs | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| classified as | Typha | Carex | Die-Back Reed | Stressed Reed | Ruderal Reed | Healthy Reed | Tree | Water/Artificial | Scirpus | Totals | User’s accuracy |
| Typha | 78 | 7 | 6 | 7 | 0 | 8 | 0 | 1 | 0 | 107 | 72.9 |
| Carex | 1 | 29 | 0 | 1 | 1 | 0 | 0 | 3 | 0 | 35 | 82.9 |
| Die-back reed | 7 | 0 | 75 | 16 | 2 | 13 | 0 | 6 | 1 | 120 | 62.5 |
| Stressed reed | 0 | 3 | 6 | 78 | 1 | 5 | 2 | 2 | 0 | 97 | 80.4 |
| Ruderal reed | 0 | 5 | 0 | 1 | 33 | 0 | 0 | 0 | 0 | 39 | 84.6 |
| Healthy reed | 2 | 4 | 11 | 4 | 5 | 109 | 0 | 0 | 1 | 136 | 80.2 |
| Tree | 0 | 0 | 0 | 0 | 0 | 0 | 99 | 0 | 0 | 99 | 100.0 |
| Water/artificial | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 104 | 1 | 105 | 99.1 |
| Scirpus | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 36 | 37 | 97.3 |
| Totals | 88 | 48 | 98 | 107 | 42 | 135 | 101 | 117 | 39 | 775 | |
| Producer’s accuracy | 88.6 | 60.4 | 76.5 | 72.9 | 78.6 | 80.7 | 98.0 | 88.9 | 92.3 | ||
| Total accuracy | Cohen’s Kappa | ||||||||||
| 82.71% | K | 0.80 | |||||||||
| Number of Reference Points | Correctly Classified | Ommission Errors | Commission Errors | User’s Accuracy [%] | Producer’s Accuracy [%] | Grouped from Original Classes | |
|---|---|---|---|---|---|---|---|
| Wetland Class | 518 | 506 | 0 | 0 | 100.0 | 97.1 | Typha, Carex, Healthy reed, Stressed reed, Die-back reed, Ruderal reed |
| Reed Class | 382 | 359 | 23 | 10 | 91.6 | 94.0 | Healthy reed, Stressed reed, Die-back reed, Ruderal reed |
| Unhealty Reed | 205 | 175 | 30 | 12 | 80.7 | 85.4 | Stressed reed, Die-back reed |
| Wetland Not Reed | 136 | 115 | 21 | 6 | 81.0 | 84.6 | Typha, Carex |
Share and Cite
Zlinszky, A.; Mücke, W.; Lehner, H.; Briese, C.; Pfeifer, N. Categorizing Wetland Vegetation by Airborne Laser Scanning on Lake Balaton and Kis-Balaton, Hungary. Remote Sens. 2012, 4, 1617-1650. https://doi.org/10.3390/rs4061617
Zlinszky A, Mücke W, Lehner H, Briese C, Pfeifer N. Categorizing Wetland Vegetation by Airborne Laser Scanning on Lake Balaton and Kis-Balaton, Hungary. Remote Sensing. 2012; 4(6):1617-1650. https://doi.org/10.3390/rs4061617
Chicago/Turabian StyleZlinszky, András, Werner Mücke, Hubert Lehner, Christian Briese, and Norbert Pfeifer. 2012. "Categorizing Wetland Vegetation by Airborne Laser Scanning on Lake Balaton and Kis-Balaton, Hungary" Remote Sensing 4, no. 6: 1617-1650. https://doi.org/10.3390/rs4061617