1. Introduction
Currently, there are many researchers studying species distribution models (SDMs) in terms of the theory [
1,
2,
3], model [
4,
5,
6,
7], and model evaluation method [
8,
9] in academic circles. For the detection of marginal peripheral populations, provenance distribution modelling may represent a valuable step forward in spatial analysis [
10]. To improve the accuracy of the model and deal with the uncertainty, ensemble models which are based on different consensus approaches could be used to balance the accuracy and enhance the robustness of SDM models [
11,
12]. In particular, under the climate regime shifts, bioclimate envelope models, which are a kind of SDM, were developed for north America to guide assisted migration and successfully predict the transformation of Douglas-fir provenance to Europe [
13]. Besides, researchers have tried to measure the climatic requirements of species using the data that is beyond their realized niches, according to the advent of large biodiversity databases and some revised SDM analysis approaches [
14]. Furthermore, to evaluate habitat suitability for plants that have no specific habitat requirement, the fuzzy matter element model (FME) which is established on the basis of Fuzzy and GIS, could provide a quantified and effective analysis. Researchers have applied FME to predict the suitability distribution of
Schisandra sphenanthera Rehd. et Wils. in the Qinling Mountains of China [
15].
The traditional SDMs, such as the generalized linear model (GLM) [
16], generalized additive model (GAM) [
17], genetic algorithm for rule-set production (GARP) [
18], Maxent [
19], and random forest (RF) [
20], are based on the location of sample sites of species. However, the FME fits the relationship between environmental variables data of sample sites and species characteristics information, such as the content of marker compounds which are extracted from the species [
21]. Therefore, the model could determine the appropriate range of various ecological environmental variables (EEVs), which provides a more accurate result for research and has a certain guiding significance for the protection of plant resources.
Over the years, researchers have explored the FME, and applied this method to predict species distribution. From 2012 to the present, our team applied the FME to study the suitability distribution of many medicinal plant species (
Table 1). The methodology of using FME is based on the following process: (1) According to previous research and the specific conditions of the plant growth environment, the EEVs are selected to construct its habitat suitability index (HSI) system; (2) EEVs, together with the characteristic values (such as the content of marker compounds of medicinal plant species), could be used to construct the membership function, which also represents the membership degree of EEVs on the habitat suitability of species; (3) the weights of EEVs for habitat suitability are calculated by using the maximum entropy theory; and (4) the spatial distribution of habitat suitability range for various EEVs are obtained by GIS layer calculations. Thus, the model adds the membership degrees of all variables and combines them with the weight, and finally obtains the average value of the membership degree.
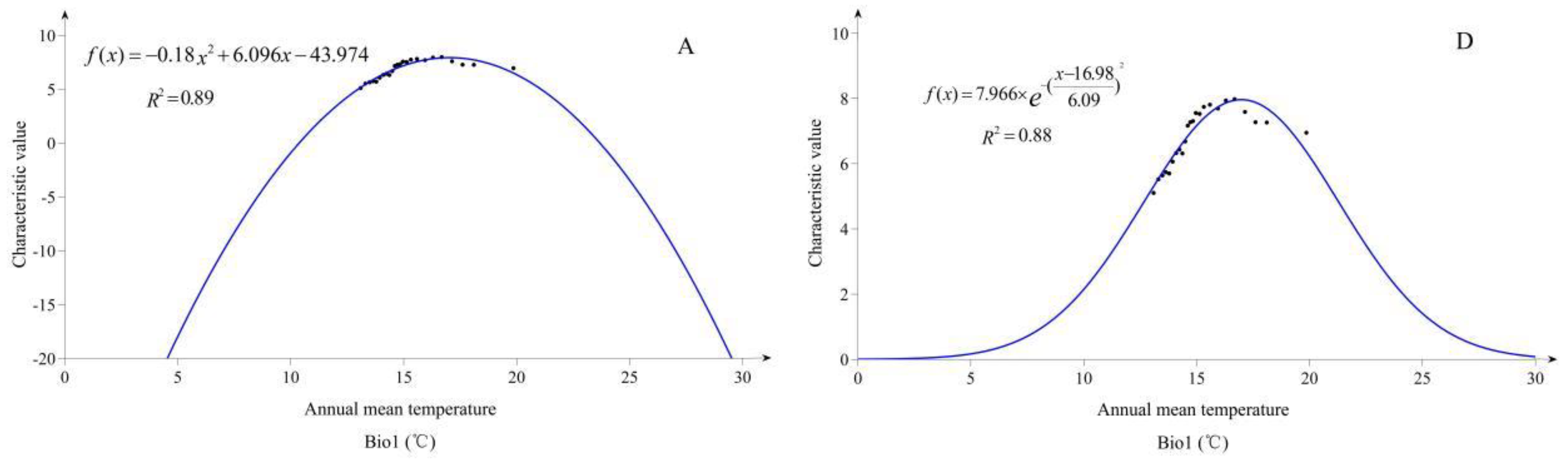
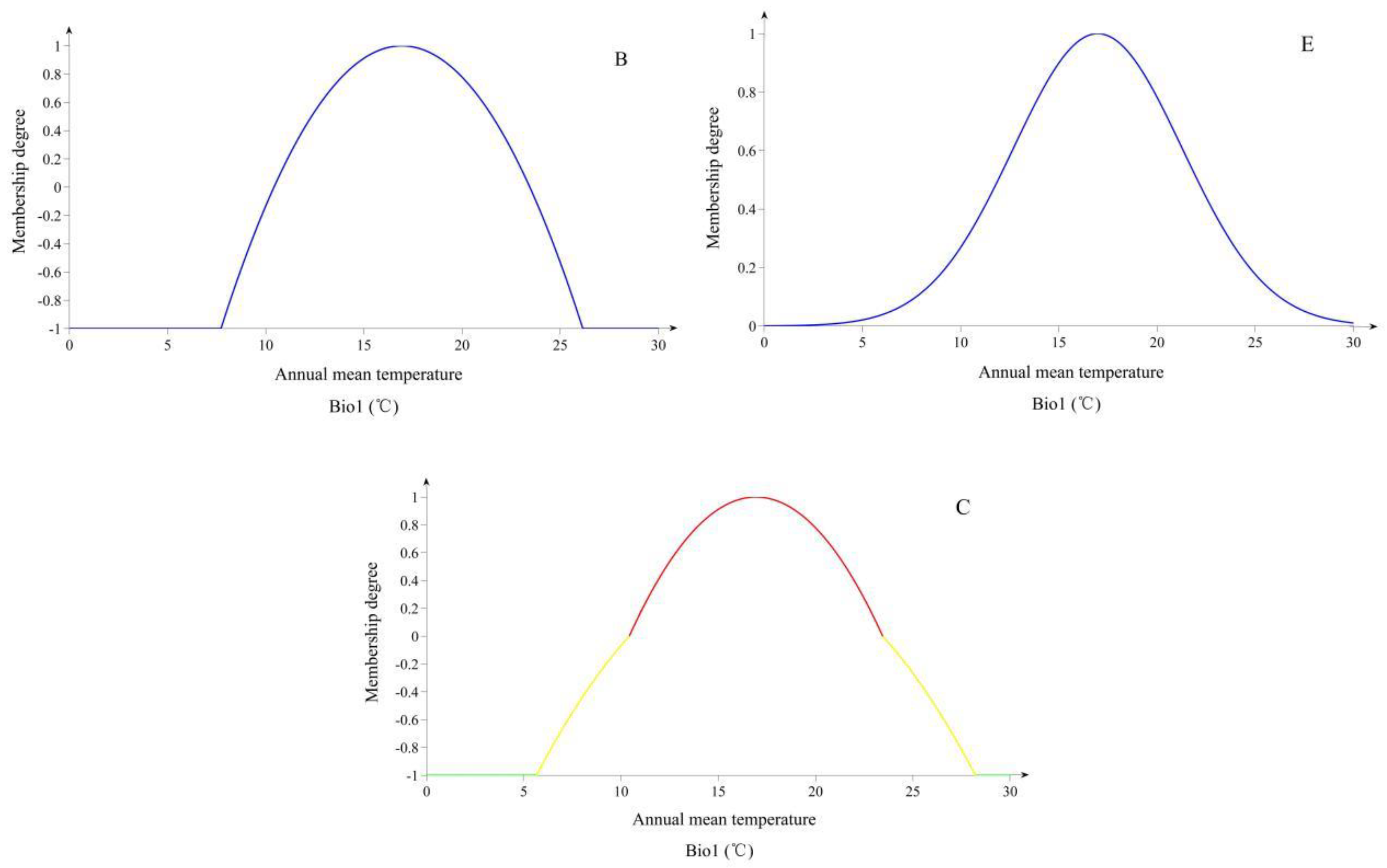
Based on a large number of actual operations of FME, we found that FME needs to be optimized and improved in the following aspects: (1) The Gaussian membership function often used by researchers might not reasonably describe the relationship between characteristic values of species and EEVs of sample sites in the low membership range. For the Gaussian membership function, since the range of the single-variable’s membership value is 0 to 1, it is possible that the HSI may present the result of the accumulated comprehensive error. It is also possible that the suitable habitat which is predicted from the model is not actually appropriated for the survival of species because of the accumulated comprehensive error; (2) any EEVs we choose for modelling have two effects on the survival of species, promotion and inhibition. For inhibition, the previous FME might not be adequately considered. Therefore, a better way is needed to describe the inhibitive effects of EEVs on the survival of species; (3) for classification of the suitability index, previous research on SDMs emphasizes the probabilistic interpretation from 0 to 1, and almost all models have limited the suitability index interval to [0, 1] [
30,
31]. However, such a classification is not very conducive to unsuitable area analysis and research; (4) we also find that the FME has the disadvantages of containing too much EEV data, and redundancy, which leads to a huge workload for researchers.
To cope with the above issues, this research tries to provide the following optimization. Firstly, we propose a quadratic function instead of a Gaussian function to fit the relationship between species characteristic values and the EEVs, because the quadratic function could describe the symmetry, as well as the Gaussian function. Secondly, extending the numerical range of membership from [0, 1] to [−1, 1], and a new criterion of habitat suitability, are also proposed. Thirdly, EEVs are filtered and key EEVs are retained to participate in modeling. Lastly, we take bio-climatic variables, topographic variables, and soil variables as EEVs in FME model predictions of future scenarios.
This research aims to optimize the key steps of the FME method; improve the fuzzy membership function, making it more conform to the biological and ecological characteristics; and avoid the accumulation of errors due to function fitting. We used
Gynostemma pentaphyllum (Thunb.) Makino as an example, which was studied by FME before [
28].
G. pentaphyllum is a herbaceous climbing plant [
32]. The gypenosides extracted from the
G. pentaphyllum exhibit anti-tumor and anti-aging properties, lower cholesterol, and enhance immunity [
33,
34]. We selected key EEVs to be incorporated in the modeling, which could change the previous demand for a large number of environmental variables and reduced the heavy workload. After screening, we tried to use six kinds of EEVs, and gypenosides data to show the optimized FME. BCC-CSM1-1 (BC) is an atmospheric-sea-land and sea-ice global coupling model of climate. This system is also used by China’s national climate center for the historical test and interdecadal test, which makes it more reliable for participation in the coupled model of the CMIP5 [
35]. Therefore, it is reasonable to select the data under BC for future forecasting in China. As for representative concentration pathways (RCPs), RCP4.5 and RCP6.0 are both intermediate stable paths, but RCP4.5 has a higher priority than RCP6.0 [
36]. Hence, in this research, we chose RCP4.5 as a representative concentration pathway. Finally, we used bio-climatic data for the 2050s (average for 2041–2060) and 2070s (average for 2061–2080) of RCP4.5 scenarios to show the advantages of the FME in predicting the future climate suitability of species.
3. Results
The test results show that the matching degree between the estimated result and the actual data is as high as 87.18%, indicating that the model operation is very reasonable [
40]. After testing, we confirmed the reliability of the relationship between current EEV data and the suitability of
G. pentaphyllum. Therefore, with this relationship, we predicted the probably extent of
G. pentaphyllum habitat suitability in the 2050s and 2070s periods (
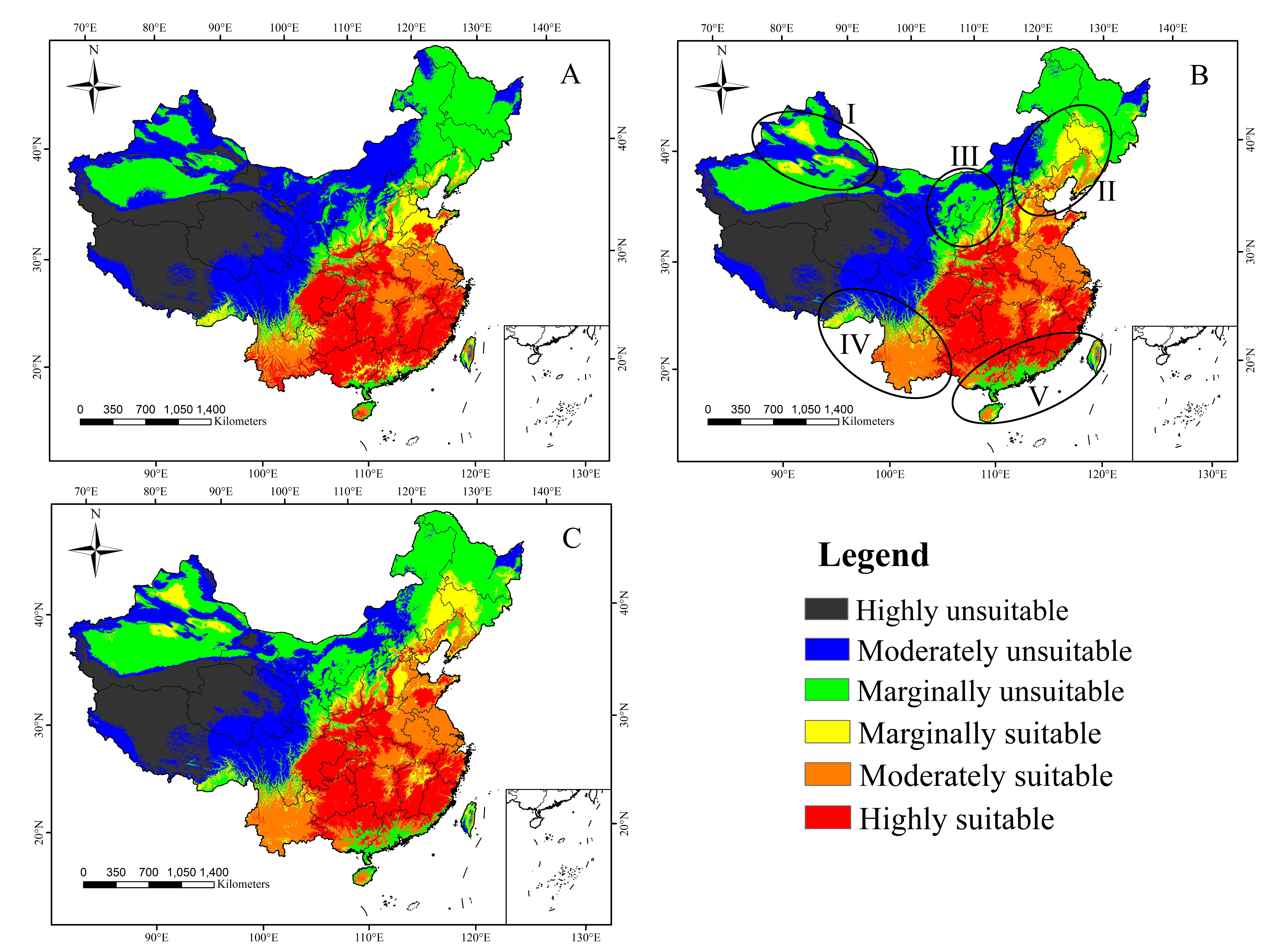
Figure 3).
Under the current EEVs, we predicted six different habitat suitabilities of
G. pentaphyllum in China. The growth suitability areas of
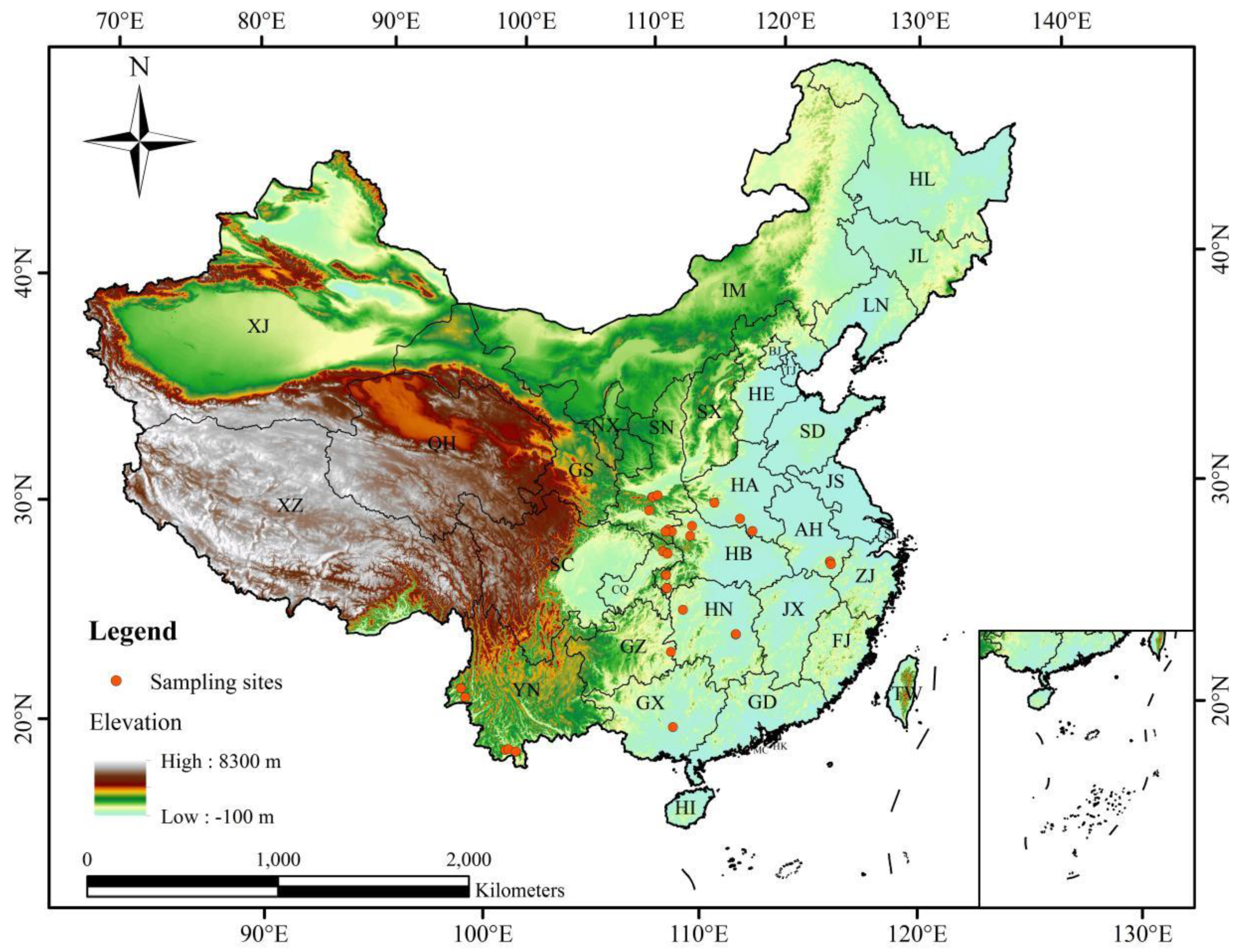
G. pentaphyllum are mainly located in the hilly and mountain terrain areas in the monsoon region of eastern China (
Figure 3A). The highly suitable habitat is mainly distributed in the provinces of Yunnan, Guizhou, Sichuan, Shaanxi, Henan, Hunan, Guangdong, Jiangxi, Fujian, Zhejiang, and Guangxi Zhuang Autonomous Region. In addition, a small number of highly suitable regions are located in Shandong, Hubei, Hebei, and Hainan. In regards to climate type, highly suitable areas are mainly affected by the subtropical monsoon and monsoon humid climate. The moderately suitable habitat is mainly distributed in Yunnan, Hubei, Hunan, Anhui, and Jiangsu provinces. The marginally suitable habitat is mainly distributed in Shandong, Hebei, and southern Tibet.
For the unsuitable habitats of G. pentaphyllum, they are mainly located in the plateau of western and northern China, and a small part of the area is located in the plains of eastern China. The highly unsuitable habitat of G. pentaphyllum is mainly located in the western and northern parts of the Qinghai-Tibet Plateau, as well as in the desert areas in Gansu province, Inner Mongolia Autonomous Region, and Xinjiang Uighur Autonomous Region. The moderately unsuitable areas of G. pentaphyllum are mainly located in the southern and eastern parts of the Qinghai-Tibet Plateau, Inner Mongolia Autonomous Region, Ningxia Hui Autonomous Region, Xinjiang Uygur Autonomous Region, and Gansu Province, with a small portion located in the western part of the Tibet Autonomous Region. The marginally unsuitable regions of G. pentaphyllum are mainly located in parts of the Xinjiang Uygur Autonomous Region and northeast of China. A few marginally unsuitable regions are located in Gansu, Shaanxi, Shanxi, and Hebei provinces, as well as the areas where Sichuan and Yunnan provinces border the Qinghai-Tibet Plateau and the coastal areas of Guangdong province.
Under the selected climate change scenarios, the suitable region of
G. pentaphyllum will undergo some changes in the 2050s and 2070s periods (
Table 7), and the overall habitat suitability of
G. pentaphyllum will move to the north (
Figure 3B,C). In general, the habitat suitability of
G. pentaphyllum in China will gradually shift to high latitudes during 2050s and 2070s periods, mainly occurring in the five regions (
Figure 3B).
In the 2050s period, the marginally suitable areas for the growth of
G. pentaphyllum will first appear in the northern and southern regions of the Tianshan Mountains in Xinjiang Uighur Autonomous Region (I), and these regions will show signs of expansion during the 2070s period (
Figure 3C). For the border between the North China Plain and the Northeast Plain (II), from the current to the 2050s, some areas will change from marginally unsuitable to marginally suitable habitats, and some areas will change from marginally suitable to moderately suitable habitats. Furthermore, in the 2070s, the marginally and moderately suitable areas will display the trend of expanding to the northeast. For the region III where the Loess Plateau and the Inner Mongolian Plateau border, from the current to the 2050s and 2070s, the moderately unsuitable habitat of
G. pentaphyllum will gradually turn into marginally unsuitable habitat, and the marginally unsuitable areas will expand northwards. For region IV, in Yunnan province, the current highly suitable areas will gradually become moderately suitable areas. In the foothills of the southern Himalayas, the current marginally suitable areas will gradually become marginally unsuitable areas. In the coastal areas of Guangdong, Fujian, and parts of Hainan Island (V), there are clear signs of a decline in habitat suitability.







{kind=link}
{kind=link}
{kind=link}
{kind=link}