Hantavirus Immunology of Rodent Reservoirs: Current Status and Future Directions

Abstract
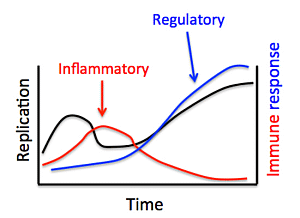
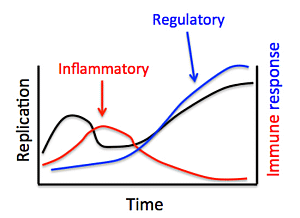
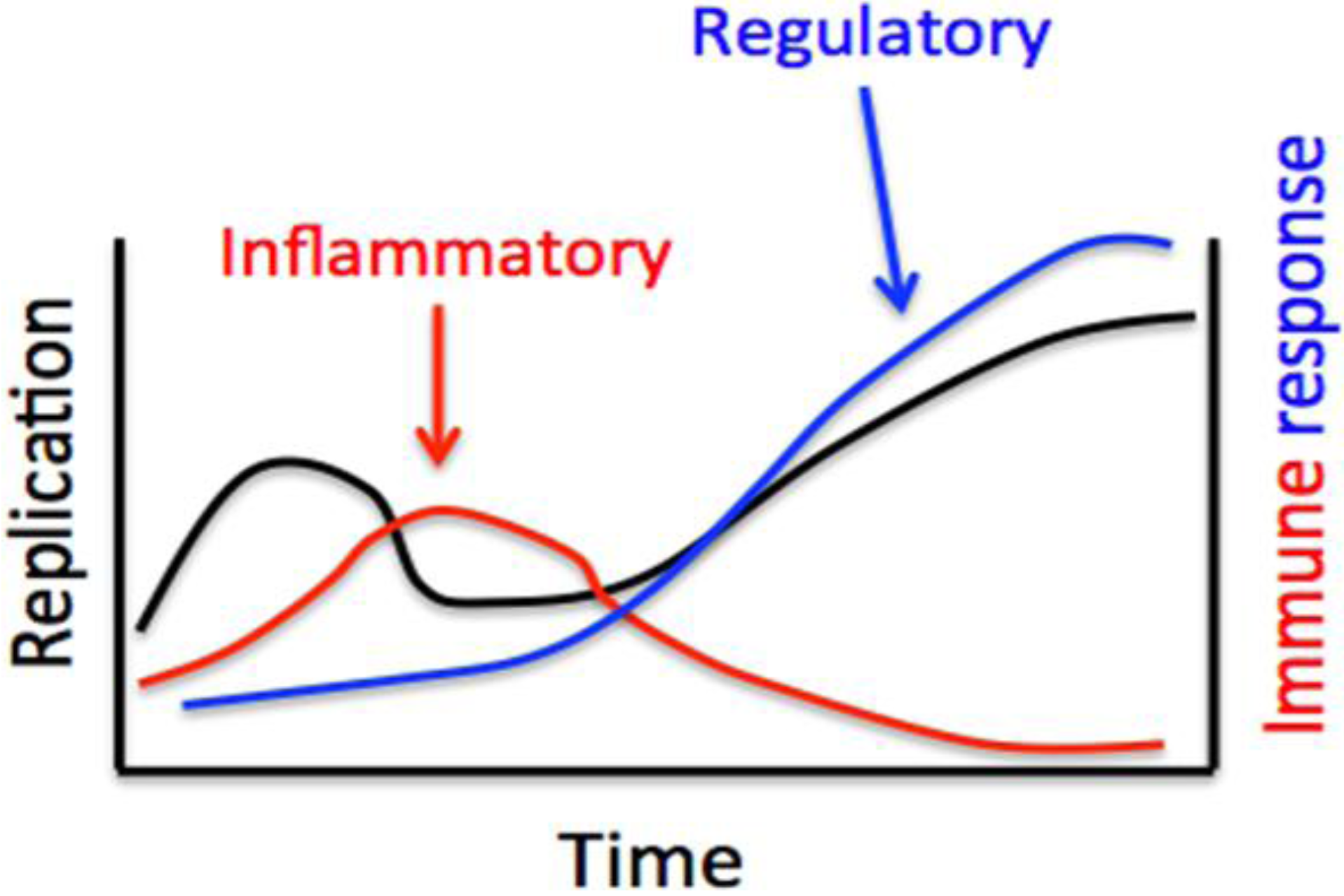
:
1. Introduction

{kind=link}
{kind=link}
| Virus | Reservoir | Disease * |
|---|---|---|
| Hantaan virus | Apodemus agrarius | HFRS |
| Dobrava virus | Apodemus flavicollis | HFRS |
| Seoul virus | Rattus norvegicus | HFRS |
| Saaremaa virus | Apodemus agrarius | HFRS |
| Amur virus | Apodemus peninsulae | HFRS |
| Puumala virus | Myodes glareolus | HFRS |
| Sin Nombre virus | Peromyscus maniculatus | HCPS |
| New York-1 virus | Peromyscus leucopus | HCPS |
| Monongahela virus | Peromyscus leucopus | HCPS |
| Bayou virus | Otyzomys palustris | HCPS |
| Black Creek Canal virus | Sigmodon hispidis | HCPS |
| Andes virus | Oligoryzomys longicaudatus | HCPS |
| Laguna Negra virus | Calomys laucha | HCPS |
| Araraquara virus | Bolomys lasiurus | HCPS |
| Choclo virus | Oligoryzomys fulvenscens | HCPS |
| Juquitiba virus | Oligoryzomys nigripes | HCPS |
| Castelo dos Sonhos | Oligoryzomys utiaritensis | HCPS |
| Bermejo virus | Oligoryzomys chacoensis | HCPS |
| Lechiguanas virus | Oligoryzomys flavescens | HCPS |
2. Infection of Rodent Reservoirs
3. Antibody Responses
4. Signatures of Immunomodulatory Activities of Hantaviruses
| Viral Protein | Putative Effect |
|---|---|
| Gn ITAM | May promote ubiquitination of Gn |
| Interaction with Fyn, Lyk, Syk and ZAP-70 kinases | |
| Inhibition of RIG-I and TBK1 pathways and IRF3 signaling | |
| N | Interference with TNF-mediated NF-κB nuclear translocation |
| Inhibition of STAT1 phosphorylation | |
| Inhibition of CTL-mediated apoptosis (granzyme B and caspase 3) | |
| Inhibition of TBK1 activation | |
| NSs | Inhibition of Ifnβ expression, NF-κB and IRF-3 activities |
5. Immune Responses of Rodent Reservoirs

6. Syrian Hamster Models of Hantavirus Disease
7. Future Directions
7.1. RNA-Seq
7.2. MicroRNA
7.3. Metabolomics
7.4. Cellular Immunology
8. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References and Notes
- Schmaljohn, C.; Hjelle, B. Hantaviruses: A global disease problem. Emerg. Infect. Dis. 1997, 3, 95–104. [Google Scholar] [CrossRef]
- Jaaskelainen, K.M.; Kaukinen, P.; Minskaya, E.S.; Plyusnina, A.; Vapalahti, O.; Elliott, R.M.; Weber, F.; Vaheri, A.; Plyusnin, A. Tula and Puumala hantavirus NSs ORFs are functional and the products inhibit activation of the interferon-beta promoter. J. Med. Virol. 2007, 79, 1527–1536. [Google Scholar] [CrossRef]
- Vera-Otarola, J.; Solis, L.; Soto-Rifo, R.; Ricci, E.P.; Pino, K.; Tischler, N.D.; Ohlmann, T.; Darlix, J.L.; Lopez-Lastra, M. The Andes hantavirus NSs protein is expressed from the viral small mRNA by a leaky scanning mechanism. J. Virol. 2012, 86, 2176–2187. [Google Scholar] [CrossRef]
- Guo, W.P.; Lin, X.D.; Wang, W.; Tian, J.H.; Cong, M.L.; Zhang, H.L.; Wang, M.R.; Zhou, R.H.; Wang, J.B.; Li, M.H.; et al. Phylogeny and origins of hantaviruses harbored by bats, insectivores, and rodents. PLoS Pathog. 2013, 9, e1003159. [Google Scholar] [CrossRef]
- Easterbrook, J.D.; Klein, S.L. Immunological mechanisms mediating hantavirus persistence in rodent reservoirs. PLoS Pathog. 2008, 4, e1000172. [Google Scholar] [CrossRef]
- Lehmer, E.M.; Jones, J.D.; Bego, M.G.; Varner, J.M.; Jeor, S.S.; Clay, C.A.; Dearing, M.D. Long-term patterns of immune investment by wild deer mice infected with Sin Nombre virus. Physiol. Biochem. Zool. 2010, 83, 847–857. [Google Scholar] [CrossRef]
- Martin, L.B., 2nd; Weil, Z.M.; Nelson, R.J. Refining approaches and diversifying directions in ecoimmunology. Integr. Comp. Biol. 2006, 46, 1030–1039. [Google Scholar] [CrossRef]
- Bagamian, K.H.; Towner, J.S.; Kuenzi, A.J.; Douglass, R.J.; Rollin, P.E.; Waller, L.A.; Mills, J.N. Transmission ecology of Sin Nombre hantavirus in naturally infected North American deermouse populations in outdoor enclosures. PLoS One 2012, 7, e47731. [Google Scholar]
- Bagamian, K.H.; Towner, J.S.; Mills, J.N.; Kuenzi, A.J. Increased detection of Sin Nombre hantavirus RNA in antibody-positive deer mice from Montana, USA: Evidence of male bias in RNA viremia. Viruses 2013, 5, 2320–2328. [Google Scholar] [CrossRef]
- Bininda-Emonds, O.R.; Cardillo, M.; Jones, K.E.; MacPhee, R.D.; Beck, R.M.; Grenyer, R.; Price, S.A.; Vos, R.A.; Gittleman, J.L.; Purvis, A. The delayed rise of present-day mammals. Nature 2007, 446, 507–512. [Google Scholar] [CrossRef]
- Jonsson, C.B.; Figueiredo, L.T.; Vapalahti, O. A global perspective on hantavirus ecology, epidemiology, and disease. Clin. Microbiol. Rev. 2010, 23, 412–441. [Google Scholar] [CrossRef]
- Botten, J.; Mirowsky, K.; Kusewitt, D.; Bharadwaj, M.; Yee, J.; Ricci, R.; Feddersen, R.M.; Hjelle, B. Experimental infection model for Sin Nombre hantavirus in the deer mouse (Peromyscus maniculatus). Proc. Natl. Acad. Sci. USA 2000, 97, 10578–10583. [Google Scholar] [CrossRef]
- Schountz, T.; Acuna-Retamar, M.; Feinstein, S.; Prescott, J.; Torres-Perez, F.; Podell, B.; Peters, S.; Ye, C.; Black, W.C.; Hjelle, B. Kinetics of immune responses in deer mice experimentally infected with Sin Nombre virus. J. Virol. 2012, 86, 10015–10027. [Google Scholar] [CrossRef]
- Yanagihara, R.; Amyx, H.L.; Gajdusek, D.C. Experimental infection with Puumala virus, the etiologic agent of nephropathia epidemica, in bank voles (Clethrionomys glareolus). J. Virol. 1985, 55, 34–38. [Google Scholar]
- Lee, H.W.; Lee, P.W.; Baek, L.J.; Song, C.K.; Seong, I.W. Intraspecific transmission of Hantaan virus, etiologic agent of Korean hemorrhagic fever, in the rodent Apodemus agrarius. Am. J. Trop. Med. Hyg. 1981, 30, 1106–1112. [Google Scholar]
- Kariwa, H.; Kimura, M.; Yoshizumi, S.; Arikawa, J.; Yoshimatsu, K.; Takashima, I.; Hashimoto, N. Modes of Seoul virus infections: Persistency in newborn rats and transiency in adult rats. Arch. Virol. 1996, 141, 2327–2338. [Google Scholar] [CrossRef]
- Mori, M.; Rothman, A.L.; Kurane, I.; Montoya, J.M.; Nolte, K.B.; Norman, J.E.; Waite, D.C.; Koster, F.T.; Ennis, F.A. High levels of cytokine-producing cells in the lung tissues of patients with fatal hantavirus pulmonary syndrome. J. Infect. Dis. 1999, 179, 295–302. [Google Scholar] [CrossRef]
- Mustonen, J.; Makela, S.; Outinen, T.; Laine, O.; Jylhava, J.; Arstila, P.T.; Hurme, M.; Vaheri, A. The pathogenesis of nephropathia epidemica: New knowledge and unanswered questions. Antivir. Res. 2013, 100, 589–604. [Google Scholar] [CrossRef]
- Easterbrook, J.D.; Klein, S.L. Seoul virus enhances regulatory and reduces proinflammatory responses in male Norway rats. J. Med. Virol. 2008, 80, 1308–1318. [Google Scholar] [CrossRef]
- Easterbrook, J.D.; Zink, M.C.; Klein, S.L. Regulatory T cells enhance persistence of the zoonotic pathogen Seoul virus in its reservoir host. Proc. Natl. Acad. Sci. USA 2007, 104, 15502–15507. [Google Scholar] [CrossRef]
- Li, W.; Klein, S.L. Seoul virus-infected rat lung endothelial cells and alveolar macrophages differ in their ability to support virus replication and induce regulatory T cell phenotypes. J. Virol. 2012, 86, 11845–11855. [Google Scholar] [CrossRef]
- Hooper, J.W.; Larsen, T.; Custer, D.M.; Schmaljohn, C.S. A lethal disease model for hantavirus pulmonary syndrome. Virology 2001, 289, 6–14. [Google Scholar] [CrossRef]
- Milazzo, M.L.; Eyzaguirre, E.J.; Molina, C.P.; Fulhorst, C.F. Maporal viral infection in the Syrian golden hamster: A model of hantavirus pulmonary syndrome. J. Infect. Dis. 2002, 186, 1390–1395. [Google Scholar] [CrossRef]
- Hanson, J.D.; Utrera, A.; Fulhorst, C.F. The delicate pygmy rice rat (Oligoryzomys delicatus) is the principal host of Maporal virus (family Bunyaviridae, genus Hantavirus). Vector Borne Zoonotic Dis. 2011, 11, 691–696. [Google Scholar] [CrossRef]
- Toro, J.; Vega, J.D.; Khan, A.S.; Mills, J.N.; Padula, P.; Terry, W.; Yadon, Z.; Valderrama, R.; Ellis, B.A.; Pavletic, C.; et al. An outbreak of hantavirus pulmonary syndrome, Chile, 1997. Emerg. Infect. Dis. 1998, 4, 687–694. [Google Scholar] [CrossRef]
- Centers for Disease Control and Prevention. Laboratory management of agents associated with hantavirus pulmonary syndrome: Interim biosafety guidelines. Morb. Mortal. Wkly. Rep. 1994, 43, 1–7. [Google Scholar]
- Calisher, C.H.; Peters, C.J.; Douglass, R.J.; Kuenzi, A.J. Hantaviral infections of rodents: Possible scenarios. Archi. Virol. 2009, 154, 1195–1197. [Google Scholar] [CrossRef]
- Bernshtein, A.D.; Apekina, N.S.; Mikhailova, T.V.; Myasnikov, Y.A.; Khlyap, L.A.; Korotkov, Y.S.; Gavrilovskaya, I.N. Dynamics of Puumala hantavirus infection in naturally infected bank voles (Clethrinomys glareolus). Arch. Virol. 1999, 144, 2415–2428. [Google Scholar] [CrossRef]
- Calisher, C.H.; Root, J.J.; Mills, J.N.; Rowe, J.E.; Reeder, S.A.; Jentes, E.S.; Wagoner, K.; Beaty, B.J. Epizootiology of Sin Nombre and El Moro Canyon hantaviruses, southeastern Colorado, 1995–2000. J. Wildl. Dis. 2005, 41, 1–11. [Google Scholar] [CrossRef]
- Kallio, E.R.; Henttonen, H.; Koskela, E.; Lundkvist, A.; Mappes, T.; Vapalahti, O. Maternal antibodies contribute to sex-based difference in hantavirus transmission dynamics. Biol. Lett. 2013, 9, 20130887. [Google Scholar] [CrossRef]
- Hutchinson, K.L.; Rollin, P.E.; Shieh, W.J.; Zaki, S.; Greer, P.W.; Peters, C.J. Transmission of Black Creek Canal virus between cotton rats. J. Med. Virol. 2000, 60, 70–76. [Google Scholar] [CrossRef]
- Hannah, M.F.; Bajic, V.B.; Klein, S.L. Sex differences in the recognition of and innate antiviral responses to Seoul virus in Norway rats. Brain Behav. Immun. 2008, 22, 503–516. [Google Scholar] [CrossRef]
- Calisher, C.H.; Sweeney, W.; Mills, J.N.; Beaty, B.J. Natural history of Sin Nombre virus in western Colorado. Emerg. Infect. Dis. 1999, 5, 126–134. [Google Scholar] [CrossRef]
- Delfraro, A.; Tome, L.; D'Elia, G.; Clara, M.; Achaval, F.; Russi, J.C.; Rodonz, J.R. Juquitiba-like hantavirus from 2 nonrelated rodent species, Uruguay. Emerg. Infect. Dis. 2008, 14, 1447–1451. [Google Scholar] [CrossRef]
- Klingstrom, J.; Heyman, P.; Escutenaire, S.; Sjolander, K.B.; de Jaegere, F.; Henttonen, H.; Lundkvist, A. Rodent host specificity of European hantaviruses: Evidence of Puumala virus interspecific spillover. J. Med. Virol. 2002, 68, 581–588. [Google Scholar] [CrossRef]
- Schlegel, M.; Klempa, B.; Auste, B.; Bemmann, M.; Schmidt-Chanasit, J.; Buchner, T.; Groschup, M.H.; Meier, M.; Balkema-Buschmann, A.; Zoller, H.; et al. Dobrava-belgrade virus spillover infections, Germany. Emerg. Infect. Dis. 2009, 15, 2017–2020. [Google Scholar] [CrossRef]
- Spengler, J.R.; Haddock, E.; Gardner, D.; Hjelle, B.; Feldmann, H.; Prescott, J. Experimental Andes virus infection in deer mice: Characteristics of infection and clearance in a heterologous rodent host. PLoS One 2013, 8, e55310. [Google Scholar]
- Schountz, T.; Shaw, T.I.; Glenn, T.C.; Feldmann, H.; Prescott, J. Expression profiling of lymph node cells from deer mice infected with Andes virus. BMC Immunol. 2013, 14, 18. [Google Scholar] [CrossRef]
- Schonrich, G.; Rang, A.; Lutteke, N.; Raftery, M.J.; Charbonnel, N.; Ulrich, R.G. Hantavirus-induced immunity in rodent reservoirs and humans. Immunol. Rev. 2008, 225, 163–189. [Google Scholar] [CrossRef]
- Botten, J.; Mirowsky, K.; Kusewitt, D.; Ye, C.; Gottlieb, K.; Prescott, J.; Hjelle, B. Persistent Sin Nombre virus infection in the deer mouse (Peromyscus maniculatus) model: Sites of replication and strand-specific expression. J. Virol. 2003, 77, 1540–1550. [Google Scholar] [CrossRef]
- Ahlm, C.; Alexeyev, O.A.; Elgh, F.; Aava, B.; Wadell, G.; Tarnvik, A.; Juto, P.; Palo, T. High prevalence of hantavirus antibodies in bank voles (Clethrionomys glareolus) captured in the vicinity of households afflicted with nephropathia epidemica. Am. J. Trop. Med. Hyg. 1997, 56, 674–678. [Google Scholar]
- Mills, J.N.; Johnson, J.M.; Ksiazek, T.G.; Ellis, B.A.; Rollin, P.E.; Yates, T.L.; Mann, M.O.; Johnson, M.R.; Campbell, M.L.; Miyashiro, J.; et al. A survey of hantavirus antibody in small-mammal populations in selected United States National Parks. Am. J. Trop. Med. Hyg. 1998, 58, 525–532. [Google Scholar]
- Schountz, T.; Calisher, C.H.; Richens, T.R.; Rich, A.A.; Doty, J.B.; Hughes, M.T.; Beaty, B.J. Rapid field immunoassay for detecting antibody to Sin Nombre virus in deer mice. Emerg. Infect. Dis. 2007, 13, 1604–1607. [Google Scholar] [CrossRef]
- Bego, M.G.; Bawiec, D.; Dandge, D.; Martino, B.; Dearing, D.; Wilson, E.; St Jeor, S. Development of an ELISA to detect Sin Nombre virus-specific IgM from deer mice (Peromyscus maniculatus). J. Virol. Methods 2008, 151, 204–210. [Google Scholar] [CrossRef]
- Childs, J.E.; Ksiazek, T.G.; Spiropoulou, C.F.; Krebs, J.W.; Morzunov, S.; Maupin, G.O.; Gage, K.L.; Rollin, P.E.; Sarisky, J.; Enscore, R.E.; et al. Serologic and genetic identification of Peromyscus maniculatus as the primary rodent reservoir for a new hantavirus in the southwestern United States. J. Infect. Dis. 1994, 169, 1271–1280. [Google Scholar] [CrossRef]
- Delfraro, A.; Clara, M.; Tome, L.; Achaval, F.; Levis, S.; Calderon, G.; Enria, D.; Lozano, M.; Russi, J.; Arbiza, J. Yellow pigmy rice rat (Oligoryzomys flavescens) and hantavirus pulmonary syndrome in Uruguay. Emerg. Infect. Dis. 2003, 9, 846–852. [Google Scholar] [CrossRef]
- Glass, G.E.; Livingstone, W.; Mills, J.N.; Hlady, W.G.; Fine, J.B.; Biggler, W.; Coke, T.; Frazier, D.; Atherley, S.; Rollin, P.E.; et al. Black Creek Canal Virus infection in Sigmodon hispidus in southern Florida. Am. J. Trop. Med. Hyg. 1998, 59, 699–703. [Google Scholar]
- Lee, B.H.; Yoshimatsu, K.; Araki, K.; Ogino, M.; Okumura, M.; Tsuchiya, K.; Kariwa, H.; Arikawa, J. Detection of antibody for the serodiagnosis of hantavirus infection in different rodent species. Arch. Vrol. 2003, 148, 1885–1897. [Google Scholar] [CrossRef]
- Ravkov, E.V.; Rollin, P.E.; Ksiazek, T.G.; Peters, C.J.; Nichol, S.T. Genetic and serologic analysis of Black Creek Canal virus and its association with human disease and Sigmodon hispidus infection. Virology 1995, 210, 482–489. [Google Scholar] [CrossRef]
- Rawlings, J.A.; Torrez-Martinez, N.; Neill, S.U.; Moore, G.M.; Hicks, B.N.; Pichuantes, S.; Nguyen, A.; Bharadwaj, M.; Hjelle, B. Cocirculation of multiple hantaviruses in Texas, with characterization of the small (S) genome of a previously undescribed virus of cotton rats (Sigmodon hispidus). Am. J. Trop. Med. Hyg. 1996, 55, 672–679. [Google Scholar]
- Tersago, K.; Schreurs, A.; Linard, C.; Verhagen, R.; van Dongen, S.; Leirs, H. Population, environmental, and community effects on local bank vole (Myodes glareolus) Puumala virus infection in an area with low human incidence. Vector Borne Zoonotic Dis. 2008, 8, 235–244. [Google Scholar] [CrossRef]
- Hinson, E.R.; Hannah, M.F.; Norris, D.E.; Glass, G.E.; Klein, S.L. Social status does not predict responses to Seoul virus infection or reproductive success among male Norway rats. Brain Behav. Immun. 2006, 20, 182–190. [Google Scholar] [CrossRef]
- Vinuesa, C.G.; Linterman, M.A.; Goodnow, C.C.; Randall, K.L. T cells and follicular dendritic cells in germinal center B-cell formation and selection. Immunol. Rev. 2010, 237, 72–89. [Google Scholar] [CrossRef]
- Schountz, T.; Prescott, J.; Cogswell, A.C.; Oko, L.; Mirowsky-Garcia, K.; Galvez, A.P.; Hjelle, B. Regulatory T cell-like responses in deer mice persistently infected with Sin Nombre virus. Proc. Natl. Acad. Sci. USA 2007, 104, 15496–15501. [Google Scholar]
- Safronetz, D.; Zivcec, M.; Lacasse, R.; Feldmann, F.; Rosenke, R.; Long, D.; Haddock, E.; Brining, D.; Gardner, D.; Feldmann, H.; et al. Pathogenesis and host response in Syrian hamsters following intranasal infection with Andes virus. PLoS Pathog. 2011, 7, e1002426. [Google Scholar] [CrossRef]
- Bharadwaj, M.; Mirowsky, K.; Ye, C.; Botten, J.; Masten, B.; Yee, J.; Lyons, C.R.; Hjelle, B. Genetic vaccines protect against Sin Nombre hantavirus challenge in the deer mouse (Peromyscus maniculatus). J. Gen. Virol. 2002, 83, 1745–1751. [Google Scholar]
- Dargeviciute, A.; Brus Sjolander, K.; Sasnauskas, K.; Kruger, D.H.; Meisel, H.; Ulrich, R.; Lundkvist, A. Yeast-expressed Puumala hantavirus nucleocapsid protein induces protection in a bank vole model. Vaccine 2002, 20, 3523–3531. [Google Scholar] [CrossRef]
- Geimonen, E.; LaMonica, R.; Springer, K.; Farooqui, Y.; Gavrilovskaya, I.N.; Mackow, E.R. Hantavirus pulmonary syndrome-associated hantaviruses contain conserved and functional ITAM signaling elements. J. Virol. 2003, 77, 1638–1643. [Google Scholar] [CrossRef]
- Mou, D.L.; Wang, Y.P.; Jiang, H.; Xiao, S.Y.; Yu, X.; Li, G.Y.; Wang, P.Z.; Sun, Y.T.; Nie, Q.H.; Huang, C.X.; et al. Identification of a functional ITAM-like sequence within G1 cytoplasmic tail of Hantaan virus. Bing Du Xue Bao 2007, 23, 424–428. [Google Scholar]
- Geimonen, E.; Fernandez, I.; Gavrilovskaya, I.N.; Mackow, E.R. Tyrosine residues direct the ubiquitination and degradation of the NY-1 hantavirus G1 cytoplasmic tail. J. Virol. 2003, 77, 10760–10868. [Google Scholar] [CrossRef]
- Matthys, V.S.; Cimica, V.; Dalrymple, N.; Glennon, N.B.; Bianco, C.; Mackow, E.R. Hantavirus GnT Elements Mediate TRAF3 Binding and Inhibit RIG-I/TBK1 directed IFNbeta Transcription by Blocking IRF3 Phosphorylation. J. Virol. 2014. [Google Scholar] [CrossRef]
- Ontiveros, S.J.; Li, Q.; Jonsson, C.B. Modulation of apoptosis and immune signaling pathways by the Hantaan virus nucleocapsid protein. Virology 2010, 401, 165–178. [Google Scholar] [CrossRef]
- Taylor, S.L.; Frias-Staheli, N.; Garcia-Sastre, A.; Schmaljohn, C.S. Hantaan virus nucleocapsid protein binds to importin alpha proteins and inhibits tumor necrosis factor alpha-induced activation of nuclear factor kappa B. J. Virol. 2009, 83, 1271–1279. [Google Scholar] [CrossRef]
- Taylor, S.L.; Krempel, R.L.; Schmaljohn, C.S. Inhibition of TNF-alpha-induced activation of NF-kappaB by hantavirus nucleocapsid proteins. Ann. N. Y. Acad. Sci. 2009, 1171, E86–E93. [Google Scholar]
- Gupta, S.; Braun, M.; Tischler, N.D.; Stoltz, M.; Sundstrom, K.B.; Bjorkstrom, N.K.; Ljunggren, H.G.; Klingstrom, J. Hantavirus-infection Confers Resistance to Cytotoxic Lymphocyte-Mediated Apoptosis. PLoS Pathog. 2013, 9, e1003272. [Google Scholar] [CrossRef]
- Cimica, V.; Dalrymple, N.A.; Roth, E.; Nasonov, A.; Mackow, E.R. An innate immunity-regulating virulence determinant is uniquely encoded by the andes virus nucleocapsid protein gene. mBio 2014, 5. [Google Scholar] [CrossRef]
- Virtanen, J.O.; Jaaskelainen, K.M.; Djupsjobacka, J.; Vaheri, A.; Plyusnin, A. Tula hantavirus NSs protein accumulates in the perinuclear area in infected and transfected cells. Arch. Virol. 2010, 155, 117–121. [Google Scholar] [CrossRef]
- Blakqori, G.; Delhaye, S.; Habjan, M.; Blair, C.D.; Sanchez-Vargas, I.; Olson, K.E.; Attarzadeh-Yazdi, G.; Fragkoudis, R.; Kohl, A.; Kalinke, U.; et al. La Crosse bunyavirus nonstructural protein NSs serves to suppress the type I interferon system of mammalian hosts. J. Virol. 2007, 81, 4991–4999. [Google Scholar] [CrossRef]
- Habjan, M.; Pichlmair, A.; Elliott, R.M.; Overby, A.K.; Glatter, T.; Gstaiger, M.; Superti-Furga, G.; Unger, H.; Weber, F. NSs protein of rift valley fever virus induces the specific degradation of the double-stranded RNA-dependent protein kinase. J. Virol. 2009, 83, 4365–4375. [Google Scholar] [CrossRef]
- Weber, F.; Bridgen, A.; Fazakerley, J.K.; Streitenfeld, H.; Kessler, N.; Randall, R.E.; Elliott, R.M. Bunyamwera bunyavirus nonstructural protein NSs counteracts the induction of alpha/beta interferon. J. Virol. 2002, 76, 7949–7955. [Google Scholar] [CrossRef]
- Belkaid, Y.; Rouse, B.T. Natural regulatory T cells in infectious disease. Nat. Immunol. 2005, 6, 353–360. [Google Scholar] [CrossRef]
- Rouse, B.T.; Sarangi, P.P.; Suvas, S. Regulatory T cells in virus infections. Immunol. Rev. 2006, 212, 272–286. [Google Scholar] [CrossRef]
- Zaki, S.R.; Greer, P.W.; Coffield, L.M.; Goldsmith, C.S.; Nolte, K.B.; Foucar, K.; Feddersen, R.M.; Zumwalt, R.E.; Miller, G.L.; Khan, A.S.; et al. Hantavirus pulmonary syndrome. Pathogenesis of an emerging infectious disease. Am. J. Pathol. 1995, 146, 552–579. [Google Scholar]
- Hammerbeck, C.D.; Hooper, J.W. T cells are not required for pathogenesis in the Syrian hamster model of hantavirus pulmonary syndrome. J. Virol. 2011, 85, 9929–9944. [Google Scholar] [CrossRef]
- Robertson, S.J.; Hasenkrug, K.J. The role of virus-induced regulatory T cells in immunopathology. Springer Semin. Immunopathol. 2006, 28, 51–62. [Google Scholar] [CrossRef]
- Rouse, B.T.; Suvas, S. Regulatory cells and infectious agents: Detentes cordiale and contraire. J. Immunol. 2004, 173, 2211–2215. [Google Scholar]
- Klein, S.L.; Cernetich, A.; Hilmer, S.; Hoffman, E.P.; Scott, A.L.; Glass, G.E. Differential expression of immunoregulatory genes in male and female Norway rats following infection with Seoul virus. J. Med. Virol. 2004, 74, 180–190. [Google Scholar] [CrossRef]
- Stoltz, M.; Sundstrom, K.B.; Hidmark, A.; Tolf, C.; Vene, S.; Ahlm, C.; Lindberg, A.M.; Lundkvist, A.; Klingstrom, J. A model system for in vitro studies of bank vole borne viruses. PLoS One 2011, 6, e28992. [Google Scholar] [CrossRef]
- Totura, A.L.; Baric, R.S. SARS coronavirus pathogenesis: Host innate immune responses and viral antagonism of interferon. Curr. Opin. Virol. 2012, 2, 264–275. [Google Scholar] [CrossRef]
- Basler, C.F. Nipah and hendra virus interactions with the innate immune system. Curr. Top. Microbiol. Immunol. 2012, 359, 123–152. [Google Scholar]
- Le May, N.; Bouloy, M. Antiviral escape strategies developed by bunyaviruses pathogenic for humans. Front. Biosci. 2012, 1, 1065–1077. [Google Scholar]
- Fukuyama, S.; Kawaoka, Y. The pathogenesis of influenza virus infections: The contributions of virus and host factors. Curr. Opin. Immunol. 2011, 23, 481–486. [Google Scholar] [CrossRef]
- Geimonen, E.; Neff, S.; Raymond, T.; Kocer, S.S.; Gavrilovskaya, I.N.; Mackow, E.R. Pathogenic and nonpathogenic hantaviruses differentially regulate endothelial cell responses. Proc. Natl. Acad. Sci. USA 2002, 99, 13837–13842. [Google Scholar]
- Kraus, A.A.; Raftery, M.J.; Giese, T.; Ulrich, R.; Zawatzky, R.; Hippenstiel, S.; Suttorp, N.; Kruger, D.H.; Schonrich, G. Differential antiviral response of endothelial cells after infection with pathogenic and nonpathogenic hantaviruses. J. Virol. 2004, 78, 6143–6150. [Google Scholar] [CrossRef]
- Au, R.Y.; Jedlicka, A.E.; Li, W.; Pekosz, A.; Klein, S.L. Seoul virus suppresses NF-kappaB-mediated inflammatory responses of antigen presenting cells from Norway rats. Virology 2010, 400, 115–127. [Google Scholar] [CrossRef]
- Prescott, J.; Safronetz, D.; Haddock, E.; Robertson, S.; Scott, D.; Feldmann, H. The adaptive immune response does not influence hantavirus disease or persistence in the Syrian hamster. Immunology 2013, 140, 168–178. [Google Scholar]
- Wahl-Jensen, V.; Chapman, J.; Asher, L.; Fisher, R.; Zimmerman, M.; Larsen, T.; Hooper, J.W. Temporal Analysis of Andes Virus and Sin Nombre Virus Infection of Syrian Hamsters. J. Virol. 2007, 81, 7449–7462. [Google Scholar] [CrossRef]
- Safronetz, D.; Prescott, J.; Haddock, E.; Scott, D.P.; Feldmann, H.; Ebihara, H. Hamster-Adapted Sin Nombre Virus Causes Disseminated Infection and Efficiently Replicates in Pulmonary Endothelial Cells without Signs of Disease. J. Virol. 2013, 87, 4778–4782. [Google Scholar] [CrossRef]
- Glenn, T.C. Field guide to next-generation DNA sequencers. Mol. Ecol. Resour. 2011, 11, 759–769. [Google Scholar]
- Haas, B.J.; Papanicolaou, A.; Yassour, M.; Grabherr, M.; Blood, P.D.; Bowden, J.; Couger, M.B.; Eccles, D.; Li, B.; Lieber, M.; et al. De novo transcript sequence reconstruction from RNA-seq using the Trinity platform for reference generation and analysis. Nat. Protoc. 2013, 8, 1494–1512. [Google Scholar]
- Schulz, M.H.; Zerbino, D.R.; Vingron, M.; Birney, E. Oases: Robust de novo RNA-seq assembly across the dynamic range of expression levels. Bioinformatics 2012, 28, 1086–1092. [Google Scholar] [CrossRef]
- Li, B.; Dewey, C.N. RSEM: Accurate transcript quantification from RNA-Seq data with or without a reference genome. BMC Bioinform. 2011, 12, 323. [Google Scholar] [CrossRef]
- Anders, S.; Huber, W. Differential expression analysis for sequence count data. Genome Biol. 2010, 11, R106. [Google Scholar] [CrossRef]
- Robinson, M.D.; McCarthy, D.J.; Smyth, G.K. edgeR: A Bioconductor package for differential expression analysis of digital gene expression data. Bioinformatics 2010, 26, 139–140. [Google Scholar]
- Croft, D.; Mundo, A.F.; Haw, R.; Milacic, M.; Weiser, J.; Wu, G.; Caudy, M.; Garapati, P.; Gillespie, M.; Kamdar, M.R.; et al. The Reactome pathway knowledgebase. Nucleic Acids Res. 2014, 42, D472–D477. [Google Scholar] [CrossRef]
- Ogata, H.; Goto, S.; Sato, K.; Fujibuchi, W.; Bono, H.; Kanehisa, M. KEGG: Kyoto Encyclopedia of Genes and Genomes. Nucleic Acids Res. 1999, 27, 29–34. [Google Scholar]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A software environment for integrated models of biomolecular interaction networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef]
- Li, Y.; Shi, X. MicroRNAs in the regulation of TLR and RIG-I pathways. Cell. Mol. Immunol. 2013, 10, 65–71. [Google Scholar] [CrossRef]
- Chowdhury, D.; Novina, C.D. Potential roles for short RNAs in lymphocytes. Immunol. Cell Biol. 2005, 83, 201–210. [Google Scholar] [CrossRef]
- Pepini, T.; Gorbunova, E.E.; Gavrilovskaya, I.N.; Mackow, J.E.; Mackow, E.R. Andes virus regulation of cellular microRNAs contributes to hantavirus-induced endothelial cell permeability. J. Virol. 2010, 84, 11929–11936. [Google Scholar] [CrossRef]
- Shin, O.S.; Kumar, M.; Yanagihara, R.; Song, J.W. Hantaviruses induce cell type- and viral species-specific host microRNA expression signatures. Virology 2013, 446, 217–224. [Google Scholar] [CrossRef]
- Washietl, S.; Will, S.; Hendrix, D.A.; Goff, L.A.; Rinn, J.L.; Berger, B.; Kellis, M. Computational analysis of noncoding RNAs. Wiley Interdiscip. Rev. RNA 2012, 3, 759–778. [Google Scholar] [CrossRef]
- Chukkapalli, V.; Heaton, N.S.; Randall, G. Lipids at the interface of virus-host interactions. Curr. Opin. Microbiol. 2012, 15, 512–518. [Google Scholar] [CrossRef]
- Heaton, N.S.; Randall, G. Multifaceted roles for lipids in viral infection. Trends Microbiol. 2011, 19, 368–375. [Google Scholar] [CrossRef]
- Moser, T.S.; Schieffer, D.; Cherry, S. AMP-activated kinase restricts Rift Valley fever virus infection by inhibiting fatty acid synthesis. PLoS Pathog. 2012, 8, e1002661. [Google Scholar] [CrossRef]
- Perera, R.; Riley, C.; Isaac, G.; Hopf-Jannasch, A.S.; Moore, R.J.; Weitz, K.W.; Pasa-Tolic, L.; Metz, T.O.; Adamec, J.; Kuhn, R.J. Dengue virus infection perturbs lipid homeostasis in infected mosquito cells. PLoS Pathog. 2012, 8, e1002584. [Google Scholar]
- Ritter, J.B.; Wahl, A.S.; Freund, S.; Genzel, Y.; Reichl, U. Metabolic effects of influenza virus infection in cultured animal cells: Intra- and extracellular metabolite profiling. BMC Syst. Biol. 2010, 4, 61. [Google Scholar] [CrossRef]
- Yu, Y.; Clippinger, A.J.; Alwine, J.C. Viral effects on metabolism: Changes in glucose and glutamine utilization during human cytomegalovirus infection. Trends Microbiol. 2011, 19, 360–367. [Google Scholar] [CrossRef]
- Species Search Tool; Aviva Systems Biology: San Diego, CA, USA, 2013. Available online: http://www.avivasysbio.com/aviva/page/sst/ (accessed on 7 March 2014).
- Utermohlen, O.; Dangel, A.; Tarnok, A.; Lehmann-Grube, F. Modulation by gamma interferon of antiviral cell-mediated immune responses in vivo. J. Virol. 1996, 70, 1521–1526. [Google Scholar]
- Wille, A.; Gessner, A.; Lother, H.; Lehmann-Grube, F. Mechanism of recovery from acute virus infection. VIII. Treatment of lymphocytic choriomeningitis virus-infected mice with anti-interferon-gamma monoclonal antibody blocks generation of virus-specific cytotoxic T lymphocytes and virus elimination. Eur. J. Immunol. 1989, 19, 1283–1288. [Google Scholar] [CrossRef]
- Moskophidis, D.; Cobbold, S.P.; Waldmann, H.; Lehmann-Grube, F. Mechanism of recovery from acute virus infection: Treatment of lymphocytic choriomeningitis virus-infected mice with monoclonal antibodies reveals that Lyt-2+ T lymphocytes mediate clearance of virus and regulate the antiviral antibody response. J. Virol. 1987, 61, 1867–1874. [Google Scholar]
- Davenport, B.J.; Willis, D.G.; Prescott, J.; Farrell, R.M.; Coons, T.A.; Schountz, T. Generation of competent bone marrow-derived antigen presenting cells from the deer mouse (Peromyscus maniculatus). BMC Immunol 2004, 5, 23. [Google Scholar] [CrossRef] [Green Version]
- Vaughn, J.; Schountz, T. Discrimination of Peromyscus maniculatus leukocytes by flow cytometry. BIOS 2003, 74, 79–86. [Google Scholar]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Schountz, T.; Prescott, J. Hantavirus Immunology of Rodent Reservoirs: Current Status and Future Directions. Viruses 2014, 6, 1317-1335. https://doi.org/10.3390/v6031317
Schountz T, Prescott J. Hantavirus Immunology of Rodent Reservoirs: Current Status and Future Directions. Viruses. 2014; 6(3):1317-1335. https://doi.org/10.3390/v6031317
Chicago/Turabian StyleSchountz, Tony, and Joseph Prescott. 2014. "Hantavirus Immunology of Rodent Reservoirs: Current Status and Future Directions" Viruses 6, no. 3: 1317-1335. https://doi.org/10.3390/v6031317