Recent Evidence of Hantavirus Circulation in the American Tropic

Abstract
:1. Introduction

{kind=link}
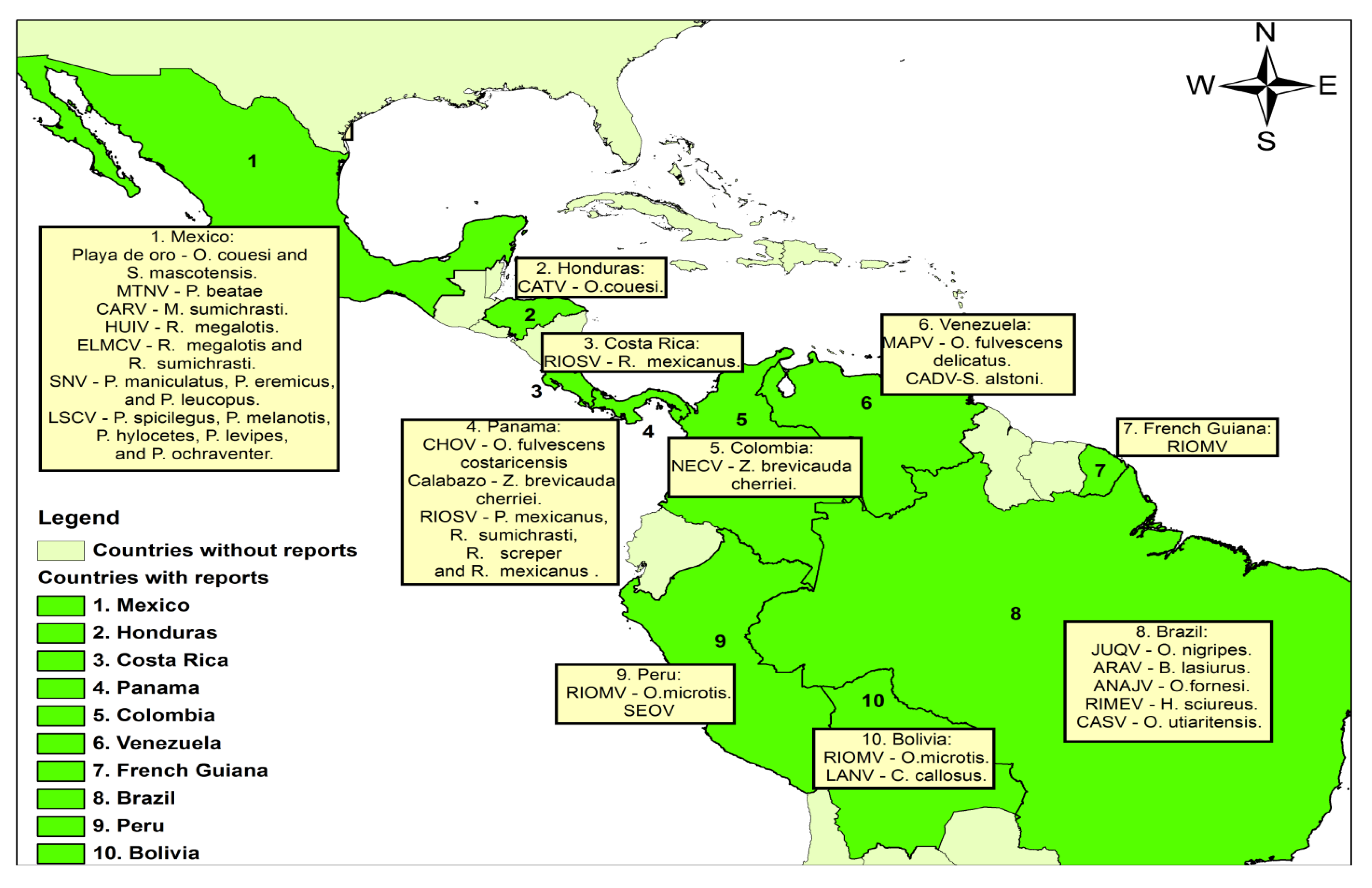
{kind=link}
| Country | Genotype | Acronym | Associated Host | Reference |
|---|---|---|---|---|
| Mexico | Playa de Oro | Oryzomys Couesi and Sigmodon mascotensis | [14] | |
| Montano | MTNV | Peromyscus beatae | [15] | |
| Carrizal | CARV | Megadontomys sumichrasti | [15] | |
| Huitzilac | HUIV | Reithrodontomys megalotis | [15] | |
| Moro Canyon | ELMCV | Reithrodontomys megalotis and R. sumichrasti | [16] | |
| Sin Nombre | SNV | Peromyscus maniculatus, P. eremicus, and P. leucopus | [16] | |
| Limestone Canyon | LSCV | Peromyscus spicilegus, P. melanotis, P. hylocetes, P. levipes and P ochraventer | [16] | |
| Honduras | Catacamas | CATV | Oryzomys couesi | [17] |
| Costa Rica | Rio segundo | RIOSV | Reithrodontomys mexicanus | [18] |
| Panamá | a Choclo | CHOV | Oligoryzomys fulvescens costaricensis | [19,20] |
| Calabazo | Zygodontomys brevicauda cherriei | [20,21] | ||
| Rio Segundo | RIOSV | Peromyscus mexicanus, Reithrodontomys sumichrasti, R. screper and R. mexicanus | [21] | |
| Colombia | Necoclí | NECV | Zygodontomys brevicauda cherriei | [22] |
| Venezuela | Maporal | MAPV | Oligoryzomys fulvescens delicatus | [19] |
| Caño Delgadito | CADV | Sigmodon alstoni | [23] | |
| French Guiana | a Rio Mamoré | RIOM | Human case | [24] |
| Bolivia | b Rio Mamoré | RIOMV | Oligoryzomys microtis | [25,26] |
| a Laguna Negra | LANV | Calomys callosus | [26] | |
| c Bermejo | BMJV | Human case | [27] | |
| Perú | a Rio Mamoré | RIOMV | Oligoryzomys microtis | [28,29] |
| a Seoul | SEOV | Human case | [30] | |
| Brazil | a Juquitiba | JUQV | Oryzomys nigripes | [31,32] |
| a Araraquara | ARAV | Bolomys lasiurus | [31,32] | |
| a Castelo dos Sonhos | CASV | Oligoryzomys utiaritensis | [31,33] | |
| Anajatuba | ANAJV | Oligoryzomys fornesi | [34] | |
| Rio Mearim | RIMEV | Holochilus sciureus | [34] |

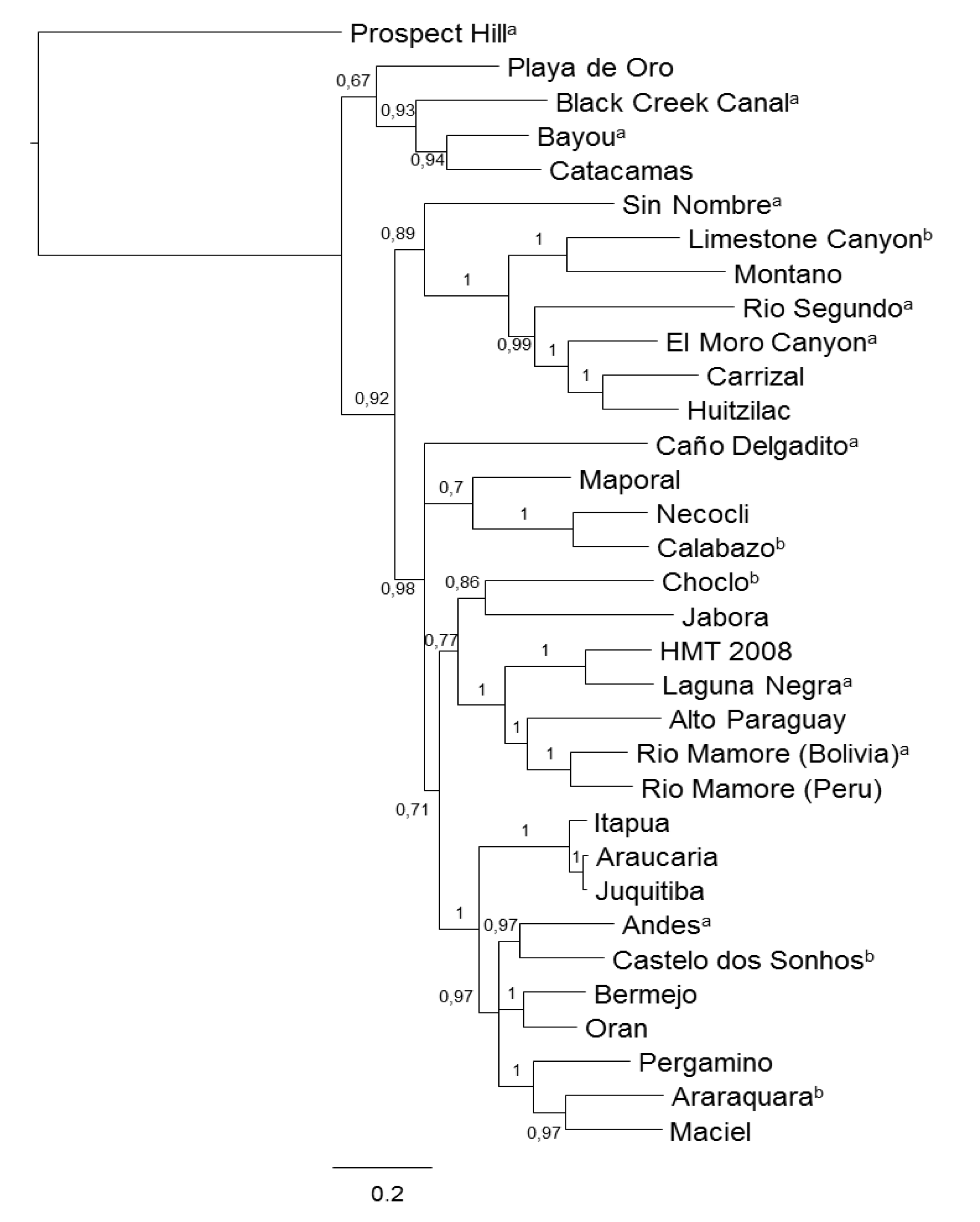
2. Reports of Hantaviruses in the Northern Cone of South America and the Brazilian Amazon
2.1. Venezuela
2.2. Colombia
2.3. Ecuador
2.4. Bolivia
2.5. Peru
2.6. French Guiana
2.7. Brazil
3. Reports of Hantavirus Circulation in Mexico, Central America and the Caribbean Islands
3.1. Mexico
3.2. Panama
3.3. Other Central American Countries
4. Caribbean Islands
5. Conclusions
| Country (State/Providence) | Positive/Tested by Rodent Species a, (Overall Percentage), Antigent b | Ref. |
|---|---|---|
| Mexico (Colima ) | 23/358 Oryzomys Couesi, 6/87 Sigmodon mascotensis and 1/77 Baiomys musculus, (5%) | [14] |
| Mexico (DF and Jalisco) | 1/8 Peromyscus maniculatus and 1/1 Reitrodontomys sumichrasti (6%) | [61] |
| Mexico (Tamaulipas) | 7/31 Peromyscus levipes, (7/31) | [62] |
| Mexico (Morelo and Guerrero) | 17/50 Peromyscus beatae, 1/6 Megadontomys thomasi, 1/6 Neotoma picta, 6/15 Reithrodontomys sumichrasti, and 2/25 R.megalotis, (12.7%) | [15] |
| Mexico (18 States) | 1/43 Baiomys musculus, 1/48 B. taylori, 1/10 Oryzomys Couesi, 1/19 Peromyscus eremicus, 2/8 P. hylocetes, 2/15 P. leucopus, 4/51 P. levipes, 3/20 P. maniculatus, 1/29 P. megalops, 3/135 P. melanotis, 2/11 P. ochraventer, 2/31 P. spicilegus, 1/43 Peromyscus spp., 5/45 Reithrodontomys megalotis, 1/4 R. microdon, 5/23 R. sumichrasti, (4%), CADV | [16] |
| Costa Rica (Heredia, Cartago and Puntarenas) | 1/3 Reithrodontomys mexicanus, (33%) c | [18] |
| Honduras (Olancho) | 5/24 Oryzomys couesi, (20.8%) CADV | [17] |
| Barbados (Bridgetown) | 19/68 Rattus Norvegicus, (25.3%), mix of old world hantaviruses | [67] |
| Panama (Los Santos) | 4/50 Zydodontomys brevicauda and 2/15 Oligoryzomys fulvenses (then it was reclassify in O.fulvescens costaricensis), (5%) | [20] |
| Panama (Los Santos) | 5/108 Zydodontomys brevicauda, 4/72 Oligoryzomys fulvescens costaricensis, 1/22 Peromyscus mexicanus, 3/4 Reithrodontomys sumichrastis; 1/7 R. Mexicanus, 1/2 R. creper, (27%) | [21] |
| Venezuela (Cojedes, Portuguesa, Barinas) | 1/13 Oryzomys bicolor, 10/166 Sigmodon alstoni, 1/45 Zygodontomys brevicauda and 1/29 Rattus rattus, (5%), PHV | [20] |
| Colombia (Cordoba) | 1/17 Heteromys sp., 4/47 Oryzomys sp., 1/11 Oligoryzomys sp. and 1/2 (50%) Proechimys sp., (2,1%) | [44] |
| Colombia (Antioquia) | 15/109 Zygodontomys brevicauda cherriei, (4.2%), SNV and MCLV | [22] |
| Peru (Loreto) | 12/50 Oligoryzomys microtis, (24%) | [28] |
| Brazil (Maranhão) | 1/40 Bolomys lasiurus, 5/12 Oligoryzomys fornesi and 15/52 Holochilus sciureus, (20.2%), SNV and ANDV | [34] |
| Country (State/Province) | Frequency of Seropositive/Tested (%), Ig a, antigen b | Studied Population | Ref. |
|---|---|---|---|
| Barbados (Bridgetown) | 11/60 (18%) IgM and 4/60 (6,6%) IgG, using mix of old world hantaviruses | Leptospiroses-like cases | [67] |
| Trinidad and Tobago | 27/236 (11,4) IgG, mix of old and new world hantaviruses | Healthy abattoir and livestock farm workers | [68] |
| Panama (Los Santos and Veraguas) | 371/1129 (32.9%) IgG | Healthy general population | [66] |
| Venezuela (Portuguesa, Capital region, Carabobo, Lara, Tachira, Anzoategui, Zulia, some samples from military recruits) | 80/1380 (5.8%), IgG, New York virus | Healthy general population | [40] |
| Colombia (Cordoba) | 12/88 (13.5%), IgG | Healthy rural volunteers | [43] |
| Bolivia (Chapare) | 61/500 (12.2%) IgG | Healthy general population | [50] |
| Bolivia (Chapare) | 9/372 (2.4%), IgG, ANDV or LANV | Febrile patients | [50] |
| Peru (Loreto) | 15/5175 (0.2%) IgM and all samples were negative for IgG, ANDV | Febrile patients | [53] |
| Peru (Loreto) | 36/2063 (1.7%), IgG, LANV, SNV, RIOMV | Healthy general population | [53] |
| French Guiana | All samples were negative for IgM and 6/420 (1.4%) had IgG | Febrile patients | [54] |
| Brazil (Maranhão) | 65/1389 (4.7%) IgG | Healthy general population | [57] |
| Brazil (Pará) | 3/2737 (0.1%) IgM and 148/2737 (5.4%) IgG, ANDV | Healthy general population | [58] |
| Brazil (Amazon) | All samples were negative for IgM and 10/1731 (0.6%) for IgG, ARAV | Healthy general population | [59] |
Acknowledgments
Author Contributions
Conflicts of Interest
References and Notes
- Nuzum, E.O.; Rossi, C.A.; Stephenson, E.H.; LeDuc, J.W. Aerosol transmission of Hantaan and related viruses to laboratory rats. Am. J. Trop. Med. Hyg. 1988, 38, 636–640. [Google Scholar]
- Lee, H.W.; Lee, P.W.; Johnson, K.M. Isolation of the etiologic agent of Korean hemorrhagic fever. J. Infect. Dis. 1978, 137, 298–308. [Google Scholar] [CrossRef]
- Nichol, S.T.; Spiropoulou, C.F.; Morzunov, S.; Rollin, P.E.; Ksiazek, T.G.; Feldmann, H.; Sanchez, A.; Childs, J.; Zaki, S.; Peters, C.J. Genetic identification of a hantavirus associated with an outbreak of acute respiratory illness. Science 1993, 262, 914–917. [Google Scholar]
- Xu, F.; Yang, Z.; Wang, L.; Lee, Y.L.; Yang, C.C.; Xiao, S.Y.; Xiao, H.; Wen, L. Morphological characterization of hantavirus HV114 by electron microscopy. Intervirology 2007, 50, 166–172. [Google Scholar] [CrossRef]
- Jaaskelainen, K.M.; Kaukinen, P.; Minskaya, E.S.; Plyusnina, A.; Vapalahti, O.; Elliott, R.M.; Weber, F.; Vaheri, A.; Plyusnin, A. Tula and Puumala hantavirus NSs ORFs are functional and the products inhibit activation of the interferon-beta promoter. J. Med. Virol. 2007, 79, 1527–1536. [Google Scholar] [CrossRef]
- Vera-Otarola, J.; Solis, L.; Soto-Rifo, R.; Ricci, E.P.; Pino, K.; Tischler, N.D.; Ohlmann, T.; Darlix, J.L.; Lopez-Lastra, M. The Andes hantavirus NSs protein is expressed from the viral small mRNA by a leaky scanning mechanism. J. Virol. 2011, 86, 2176–2187. [Google Scholar]
- Mir, M.A. Hantaviruses. Clin. Lab. Med. 2010, 30, 67–91. [Google Scholar] [CrossRef]
- Lee, P.W.; Amyx, H.L.; Yanagihara, R.; Gajdusek, D.C.; Goldgaber, D.; Gibbs, C.J. Partial characterization of prospect hill virus isolated from meadow voles in the United States. J. Infect. Dis. 1985, 152, 826–829. [Google Scholar] [CrossRef]
- Schmaljohn, C.S.; Dalrymple, J.M. Analysis of hantaan virus RNA: Evidence for a new genus of bunyaviridae. Virology 1983, 131, 482–491. [Google Scholar] [CrossRef]
- Lee, P.W.; Amyx, H.L.; Gajdusek, D.C.; Yanagihara, R.T.; Goldgaber, D.; Gibbs, C.J. New Hemorrhagic fever with renal syndrome-related virus in rodents in the United States. Lancet 1982, 320, 1405. [Google Scholar]
- Centers for Disease Control and Prevention. Outbreak of acute illness southwestern United States, 1993. In MMWR Morb. Mortal. Wkly. Rep.; 1993; 42, pp. 421–424. [Google Scholar]
- Hjelle, B.; Torres-Perez, F. Hantaviruses in the Americas and their role as emerging pathogens. Viruses 2010, 2, 2559–2586. [Google Scholar] [CrossRef]
- Matheus, S.; Djossou, F.; Moua, D.; Bourbigot, A.M.; Hommel, D.; Lacoste, V.; Dussart, P.; Lavergne, A. Hantavirus pulmonary syndrome, French Guiana. Emerg. Infect. Dis. 2010, 16, 739–741. [Google Scholar] [CrossRef]
- Chu, Y.K.; Owen, R.D.; Sanchez-Hernandez, C.; Mde, L.R.-A.; Jonsson, C.B. Genetic characterization and phylogeny of a hantavirus from western Mexico. Virus Res. 2008, 131, 180–188. [Google Scholar]
- Kariwa, H.; Yoshida, H.; Sanchez-Hernandez, C.; Mde, L.R.-A.; Almazan-Catalan, J.A.; Ramos, C.; Miyashita, D.; Seto, T.; Takano, A.; Totani, M.; et al. Genetic diversity of hantaviruses in Mexico: Identification of three novel hantaviruses from Neotominae rodents. Virus Res. 2012, 163, 486–494. [Google Scholar] [CrossRef]
- Milazzo, M.L.; Cajimat, M.N.B.; Romo, H.E.; Estrada-Franco, J.G.; Iñiguez-Dávalos, L.I.; Bradley, R.D.; Fulhorst, C.F. Geographic distribution of hantaviruses associated with neotomine and sigmodontine rodents, Mexico. Emerg. Infect. Dis. 2012, 18, 571–576. [Google Scholar]
- Milazzo, M.L.; Cajimat, M.N.; Hanson, J.D.; Bradley, R.D.; Quintana, M.; Sherman, C.; Velasquez, R.T.; Fulhorst, C.F. Catacamas virus, a hantaviral species naturally associated with Oryzomys couesi (Coues’ oryzomys) in Honduras. Am. J. Trop. Med. Hyg. 2006, 75, 1003–1010. [Google Scholar]
- Hjelle, B.; Anderson, B.; Torrez-Martinez, N.; Song, W.; Gannon, W.L.; Yates, T.L. Prevalence and geographic genetic variation of hantaviruses of new world harvest mice (Reithrodontomys): Identification of a divergent genotype from a Costa Rican Reithrodontomys mexicanus. Virology 1995, 207, 452–459. [Google Scholar] [CrossRef]
- Hanson, J.D.; Utrera, A.; Fulhorst, C.F. The delicate pygmy rice rat (Oligoryzomys delicatus) is the principal host of Maporal virus (family Bunyaviridae, genus Hantavirus). Vector Borne Zoonotic Dis. 2011, 11, 691–696. [Google Scholar] [CrossRef]
- Vincent, M.J.; Quiroz, E.; Gracia, F.; Sanchez, A.J.; Ksiazek, T.G.; Kitsutani, P.T.; Ruedas, L.A.; Tinnin, D.S.; Caceres, L.; Garcia, A.; et al. Hantavirus pulmonary syndrome in Panama: Identification of novel hantaviruses and their likely reservoirs. Virology 2000, 277, 14–19. [Google Scholar] [CrossRef]
- Salazar-Bravo, J.; Armien, B.; Suzan, G.; Armien, A.; Ruedas, L.A.; Avila, M.; Zaldivar, Y.; Pascale, J.M.; Gracia, F.; Yates, T.L. Serosurvey of wild rodents for hantaviruses in Panama, 2000–2002. J. Wildl. Dis. 2004, 40, 103–109. [Google Scholar] [CrossRef]
- Londono, A.F.; Diaz, F.J; Agudelo-Florez, P.; Levis, S.; Rodas, J.D. Genetic evidence of hantavirus infections in wild rodents from northwestern Colombia. Vector Borne Zoonotic Dis. 2011, 6, 701–708. [Google Scholar]
- Fulhorst, C.F.; Monroe, M.C.; Salas, R.A.; Duno, G.; Utrera, A.; Ksiazek, T.G.; Nichol, S.T.; de Manzione, N.M.; Tovar, D.; Tesh, R.B. Isolation, characterization and geographic distribution of Cano Delgadito virus, a newly discovered South American hantavirus (family Bunyaviridae). Virus Res. 1997, 51, 159–171. [Google Scholar] [CrossRef]
- Matheus, S.; Lavergne, A.; de Thoisy, B.; Dussart, P.; Lacoste, V. Complete genome sequence of a novel hantavirus variant of Rio Mamore Virus, Maripa Virus, from French Guiana. J. Virol. 2012, 86. [Google Scholar] [CrossRef]
- Bharadwaj, M.; Botten, J.; Torrez-Martinez, N.; Hjelle, B. Rio mamore virus: Genetic characterization of a newly recognized hantavirus of the pygmy rice rat, Oligoryzomys microtis, from Bolivia. Am. J. Trop. Med. Hyg. 1997, 57, 368–374. [Google Scholar]
- Carroll, D.S.; Mills, J.N.; Montgomery, J.M.; Bausch, D.G.; Blair, P.J.; Burans, J.P.; Felices, V.; Gianella, A.; Iihoshi, N.; Nichol, S.T.; et al. Hantavirus Pulmonary syndrome in central Bolivia: Relationships between reservoir hosts, habitats, and viral genotypes. Am. J. Trop Med. Hyg. 2005, 72, 42–46. [Google Scholar]
- Padula, P.; Valle, M.G.D.; Alai, M.G.; Cortada, P.; Villagra, M.; Gianella, A. Andes virus and first case report of bermejo virus causing fatal pulmonary syndrome. Emerg. Infect. Dis. 2002, 8, 437–439. [Google Scholar] [CrossRef]
- Powers, A.M.; Mercer, D.R.; Watts, D.M.; Guzman, H.; Fulhorst, C.F.; Popov, V.L.; Tesh, R.B. Isolation and genetic characterization of a hantavirus (Bunyaviridae: Hantavirus) from a rodent, Oligoryzomys microtis (Muridae), collected in northeastern Peru. Am. J. Trop Med. Hyg. 1999, 61, 92–98. [Google Scholar]
- Casapia, M.; Mamani, E.; Garcia, M.P.; Miraval, M.L.; Valencia, P.; Quino, A.H.; Alvarez, C.; Donaires, L.F. Hantavirus pulmonary syndrome (Rio Mamore virus) in the Peruvian Amazon region. Rev. Peru. Med. Exp. Salud Pública. 2012, 29, 390–395. [Google Scholar] [CrossRef]
- Garcia, P.P.; Herrera, S.; Donaires, A.L.; Álvarez, F.; Arrasco, C.; Cabezas, J.; Rodríguez, C.; Enrique, H. Confirmación etiológica de los dos primeros casos de hantavirosis humana en el Perú. Rev. Peru. Med. Salud Publica 2011, 28, 564–570. (in Spanish). [Google Scholar] [CrossRef]
- Johnson, A.M.; de Souza, L.T.; Ferreira, I.B.; Pereira, L.E.; Ksiazek, T.G.; Rollin, P.E.; Peters, C.J.; Nichol, S.T. Genetic investigation of novel hantaviruses causing fatal HPS in Brazil. J. Med. Virol. 1999, 59, 527–535. [Google Scholar] [CrossRef]
- Suzuki, A.; Bisordi, I.; Levis, S.; Garcia, J.; Pereira, L.E.; Souza, R.P.; Sugahara, T.K.; Pini, N.; Enria, D.; Souza, L.T. Identifying rodent hantavirus reservoirs, Brazil. Emerg. Infect. Dis. 2004, 10, 2127–2134. [Google Scholar] [CrossRef]
- Firth, C.; Tokarz, R.; Simith, D.B.; Nunes, M.R.; Bhat, M.; Rosa, E.S.; Medeiros, D.B.; Palacios, G.; Vasconcelos, P.F.; Lipkin, W.I. Diversity and distribution of hantaviruses in South America. J. Virol. 2012, 86, 13756–13766. [Google Scholar] [CrossRef]
- Rosa, E.S.; Mills, J.N.; Padula, P.J.; Elkhoury, M.R.; Ksiazek, T.G.; Mendes, W.S.; Santos, E.D.; Araújo, G.C.; Martinez, V.P.; Rosa, J.F.; et al. Newly Recognized hantaviruses associated with hantavirus pulmonary syndrome in Northern Brazil: Partial genetic characterization of viruses and serologic implication of likely reservoirs. Vector Borne Zoonotic Dis. 2005, 5, 11–19. [Google Scholar] [CrossRef]
- ArcGis, version 10.0. Environmental System Research Institute: Redlands, CA, USA, 2010.
- Macneil, A.; Nichol, S.T.; Spiropoulou, C.F. Hantavirus pulmonary syndrome. Virus Res. 2011, 162, 138–147. [Google Scholar] [CrossRef]
- MrBayes: Bayesian Inference of Phylogeny, version 3.2.2; Available online: http://www.mybiosoftware.com/phylogenetic-analysis/2463/ (accessed on 1 September 2013).
- FigTree, version 1.1.2; Available online: http://tree.bio.ed.ac.uk/software/figtree/ (accessed on 1 September 2013).
- Milazzo, M.L.; Eyzaguirre, E.J.; Molina, C.P.; Fulhorst, C.F. Maporal viral infection in the Syrian Golden Hamster: A model of hantavirus pulmonary syndrome. J. Infect. Dis. 2002, 186, 1390–1395. [Google Scholar] [CrossRef]
- Rivas, Y.J.; Moros, Z.; Moron, D.; Uzcategui, M.G.; Duran, Z.; Pujol, F.H.; Liprandi, F.; Ludert, J.E. The seroprevalences of anti-hantavirus IgG antibodies among selected Venezuelan populations. Ann. Trop Med. Parasitol. 2003, 97, 61–67. [Google Scholar] [CrossRef]
- Fulhorst, C.F.; Milazzo, M.L.; Duno, G.; Salas, R.A. Experimental infection of the Sigmodon alstoni cotton rat with Cano Delgadito virus, a South American hantavirus. Am. J. Trop. Med. Hyg. 2002, 67, 107–111. [Google Scholar]
- Milazzo, M.L.; Duno, G.; Utrera, A.; Richter, M.H.; Duno, F.; de Manzione, N.; Fulhorst, C.F. Natural host relationships of hantaviruses native to western Venezuela. Vector Borne Zoonotic Dis. 2010, 10, 605–611. [Google Scholar] [CrossRef]
- Mattar, S.; Parra, M. Serologic evidence of hantavirus infection in humans, Colombia. Emerg. Infect. Dis. 2004, 10, 2263–2264. [Google Scholar] [CrossRef]
- Alemán, A.; Iguarán, H.; Puerta, H.; Cantillo, C.; Mills, J.; Ariz, W.; Mattar, S. Primera evidencia serológica de infección por hantavirus en roedores, en Colombia. Rev. Salud Pública 2006, 8, 1–12. (in Spanish). [Google Scholar]
- Glass, G.E.; Watson, A.J.; LeDuc, J.W.; Childs, J.E. Domestic cases of hemorrhagic fever with renal syndrome in the United States. Nephron 1994, 68, 48–51. [Google Scholar] [CrossRef]
- Demeester, R.; Bottieau, E.; van Esbroeck, M.; Pourkarim, M.R.; Maes, P.; Clement, J. Hantavirus nephropathy as a pseudo-import pathology from ecuador. Eur. J. Clin. Microbiol. Infect. Dis. 2009, 29, 59–62. [Google Scholar]
- Espinoza, R.; Vial, P.; Noriega, L.M.; Johnson, A.; Nichol, S.T.; Rollin, P.E.; Wells, R.; Zaki, S.; Reynolds, E.; Ksiazek, T.G. Hantavirus pulmonary syndrome in a chilean patient with recent travel in Bolivia. Emerg. Infect. Dis. 1998, 4, 93–95. [Google Scholar] [CrossRef]
- Johnson, A.M.; Bowen, M.D.; Ksiazek, T.G.; Williams, R.J.; Bryan, R.T.; Mills, J.N.; Peters, C.J.; Nichol, S.T. Laguna Negra virus associated with HPS in western Paraguay and Bolivia. Virology 1997, 238, 115–127. [Google Scholar] [CrossRef]
- Levis, S.; Morzunov, S.P.; Rowe, J.E.; Enria, D.; Pini, N.; Calderon, G.; Sabattini, M.; St Jeor, S.C. Genetic diversity and epidemiology of hantaviruses in Argentina. J. Infect. Dis. 1998, 177, 529–538. [Google Scholar]
- Cruz, C.D.; Forshey, B.M.; Vallejo, E.; Agudo, R.; Vargas, J.; Blazes, D.L.; Guevara, C.; Laguna-Torres, V.A.; Halsey, E.S.; Kochel, T.J. Novel strain of Andes Virus associated with fatal human infection, central Bolivia. Emerg. Infect. Dis. 2012, 18, 750–757. [Google Scholar]
- Richter, M.H.; Hanson, J.D.; Cajimat, M.N.; Milazzo, M.L.; Fulhorst, C.F. Geographical range of Rio Mamore virus (family Bunyaviridae, genus Hantavirus) in association with the small-eared pygmy rice rat (Oligoryzomys microtis). Vector Borne Zoonotic Dis. 2010, 10, 613–620. [Google Scholar] [CrossRef]
- Seijo, A.; Pini, N.; Levis, S.; Coto, H.; Deodato, B.; Cernigoi, B.; de Bassadoni, D.; Enria, D. Study of hantavirus Seoul in a human and rodent population from a marginal area in Buenos Aires City. Medicina 2003, 63, 193–196. [Google Scholar]
- Ore, R.M.C.; Forshey, B.M.; Huaman, A.; Villaran, M.V.; Long, K.C.; Kochel, T.J.; Guevara, C.; Montgomery, J.M.; Alvarez, C.A.; Vilcarromero, S. Serologic evidence for human hantavirus infection in Peru. Vector Borne Zoonotic Dis. 2012, 12, 683–689. [Google Scholar] [CrossRef]
- Matheus, S.; Meynard, J.B.; Rollin, P.; Maubert, B.; Morvan, J. New world hantavirus in humans, French Guiana. Emerg. Infect. Dis. 2006, 12, 1294–1295. [Google Scholar] [CrossRef]
- Monroe, M.C.; Morzunov, S.P.; Johnson, A.M.; Bowen, M.D.; Artsob, H.; Yates, T.; Peters, C.J.; Rollin, P.E.; Ksiazek, T.G.; Nichol, S.T. Genetic diversity and distribution of peromyscus-borne hantaviruses in North America. Emerg. Infect. Dis. 1999, 5, 75–86. [Google Scholar] [CrossRef]
- Dos Santos, M.C.; de Lacerda, M.V.G.; Benedetti, S.M.; Albuquerque, B.C.; de Aguiar Filho, A.A.; da Rosa Elkhoury, M.; da Rosa, E.S.T.; da Costa Vasconcelos, P.F.; de Almeida Medeiros, D.B.; Mourão, M.P.G. Human hantavirus infection, Brazilian Amazon. Emerg. Infect. Dis. 2006, 12, 1165–1167. [Google Scholar] [CrossRef]
- Mendes, W.S.; da Silva, A.A.M.; Neiva, R.F.; Costa, N.M.; de Assis, M.S.; Vidigal, P.M.O.; da Leite, M.; da Rosa, E.S.T.; de A. Medeiros, D.B.; de B. Simith, D.; et al. Serologic survey of hantavirus infection, Brazilian Amazon. Emerg. Infect. Dis. 2010, 16, 889–891. [Google Scholar] [CrossRef]
- Medeiros, D.B.; da Rosa, E.S.; Marques, A.A.; Simith, D.B.; Carneiro, A.R.; Chiang, J.O.; Prazeres, I.T.; Vasconcelos, P.F.; Nunes, M.R. Circulation of hantaviruses in the influence area of the cuiaba-santarem highway. Mem. Inst. Oswaldo Cruz. 2010, 105, 665–671. [Google Scholar] [CrossRef]
- Gimaque, J.B.; Bastos Mde, S.; Braga, W.S.; Oliveira, C.M.; Castilho Mda, C.; Figueiredo, R.M.; Galusso, E.S.; Itapirema, E.F.; Fiqueiredo, L.T.; Mourão, M.P. Serological Evidence of hantavirus infection in rural and urban regions in the state of Amazonas, Brazil. Mem. Inst. Oswaldo Cruz. 2012, 107, 135–137. [Google Scholar] [CrossRef]
- Rowe, J.E.; St. Jeor, S.C.; Riolo, J.; Otteson, E.W.; Monroe, M.C.; Henderson, W.W.; Ksiazek, T.G.; Rollin, P.E.; Nichol, S.T. Coexistence of several novel hantaviruses in rodents indigenous to North America. Virology 1995, 213, 122–130. [Google Scholar] [CrossRef]
- Suzan, G.; Ceballos, G.; Mills, J.; Ksiazek, T.G.; Yates, T. Serologic Evidence of hantavirus infection in sigmodontine rodents in Mexico. J. Wildl. Dis. 2001, 37, 391–393. [Google Scholar] [CrossRef]
- Castro-Arellano, I.; Suzan, G.; Leon, R.F.; Jimenez, R.M.; Lacher, T.E. Survey for antibody to hantaviruses in Tamaulipas, Mexico. J. Wildl. Dis. 2009, 45, 207–212. [Google Scholar] [CrossRef]
- Gracia, F.; Armien, B.; Simpson, S.Q.; Munoz, C.; Broce, C.; Pascale, J.M.; Koster, F. Convalescent pulmonary dysfunction following hantavirus pulmonary syndrome in Panama and the United States. Lung 2010, 188, 387–391. [Google Scholar] [CrossRef]
- Suzan, G.; Giermakowski, J.T.; Marce, E.; Suzan-Azpiri, H.; Armien, B.; Yates, T.L. Modeling hantavirus reservoir species dominance in high seroprevalence areas on the Azuero Peninsula of Panama. Am. J. Trop. Med. Hyg. 2006, 74, 1103–1110. [Google Scholar]
- Suzan, G.; Marce, E.; Giermakowski, J.T.; Mills, J.N.; Ceballos, G.; Ostfeld, R.S.; Armien, B.; Pascale, J.M.; Yates, T.L. Experimental evidence for reduced rodent diversity causing increased hantavirus prevalence. PLoS One. 2009, 4, e5461. [Google Scholar] [CrossRef]
- Armien, B.; Pascale, J.M.; Munoz, C.; Lee, S.J.; Choi, K.L.; Avila, M.; Broce, C.; Armien, A.G.; Gracia, F.; Hjelle, B.; et al. Incidence rate for hantavirus infections without pulmonary syndrome, Panama. Emerg. Infect. Dis. 2011, 17, 1936–1939. [Google Scholar] [CrossRef]
- Groen, J.; Koraka, P.; Edwards, C.N.; Branch, S.L.; Douglas, K.O.; Osterhaus, A.D.; Levett, P.N. Serological evidence of hantavirus in humans and rodents in Barbados. J. Infect. 2002, 45, 109–110. [Google Scholar] [CrossRef]
- Adesiyun, A.; Dookeran, S.; Stewart-Johnson, A.; Rahaman, S.; Bissessar, S.; Thompson, N. Serological evidence of hantavirus infection in farm and abattoir workers in Trinidad a preliminary study. J. Agromedicine 2011, 16, 194–199. [Google Scholar] [CrossRef]
- Lopez, N.; Padula, P.; Rossi, C.; Lazaro, M.E.; Franze-Fernandez, M.T. Genetic identification of a new hantavirus causing severe pulmonary syndrome in Argentina. Virology 1996, 220, 223–226. [Google Scholar] [CrossRef]
- Hjelle, B.; Torrez-Martinez, N.; Koster, F.T. Hantavirus pulmonary syndrome related virus from Bolivia. Lancet 1996, 347, 57. [Google Scholar]
- International Committee on Taxonomy of Viruses. 2012. Available online: http://talk.ictvonline.org/files/ictv_official_taxonomy_updates_since_the_8th_report/m/vertebrate-official/224.aspx/ (accessed on 10 October 2012).
- Coke, R.J.; Hunter, B.M.; Rudge, J.W.; Liverani, M.; Hanvoravongchai, P. Emerging infectious diseases in southeast Asia: Regional challenges to control. Lancet 2011, 377, 599–609. [Google Scholar] [CrossRef]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Montoya-Ruiz, C.; Diaz, F.J.; Rodas, J.D. Recent Evidence of Hantavirus Circulation in the American Tropic. Viruses 2014, 6, 1274-1293. https://doi.org/10.3390/v6031274
Montoya-Ruiz C, Diaz FJ, Rodas JD. Recent Evidence of Hantavirus Circulation in the American Tropic. Viruses. 2014; 6(3):1274-1293. https://doi.org/10.3390/v6031274
Chicago/Turabian StyleMontoya-Ruiz, Carolina, Francisco J. Diaz, and Juan D. Rodas. 2014. "Recent Evidence of Hantavirus Circulation in the American Tropic" Viruses 6, no. 3: 1274-1293. https://doi.org/10.3390/v6031274