Understanding the Process of Envelope Glycoprotein Incorporation into Virions in Simian and Feline Immunodeficiency Viruses

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Simian Immunodeficiency Viruses
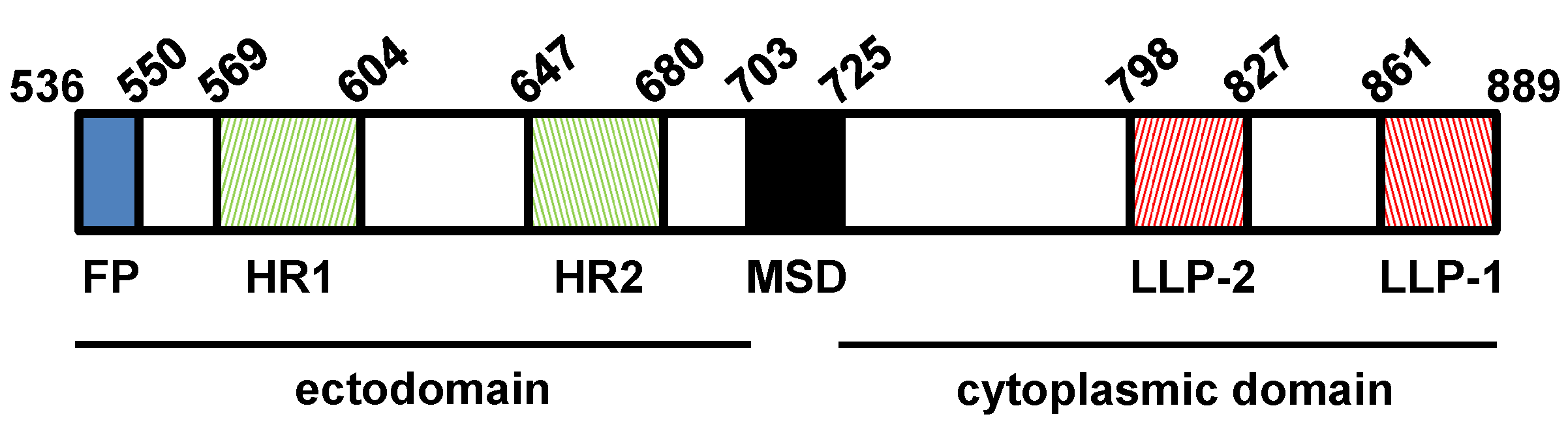
3. The SIV Env Glycoprotein
3.1. SIV gp120
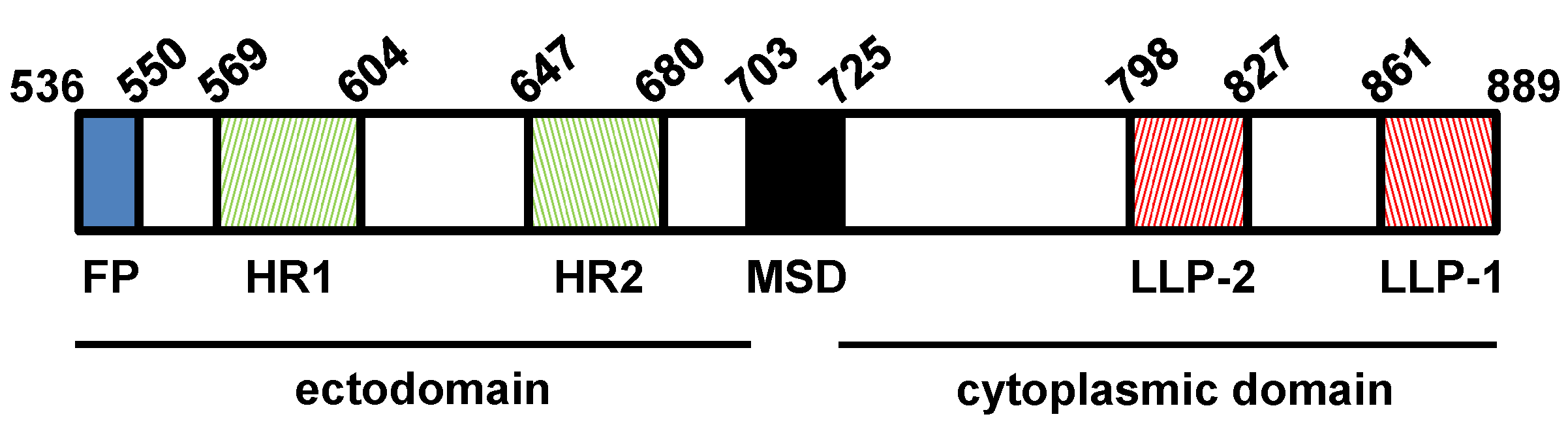
3.2. SIV gp41
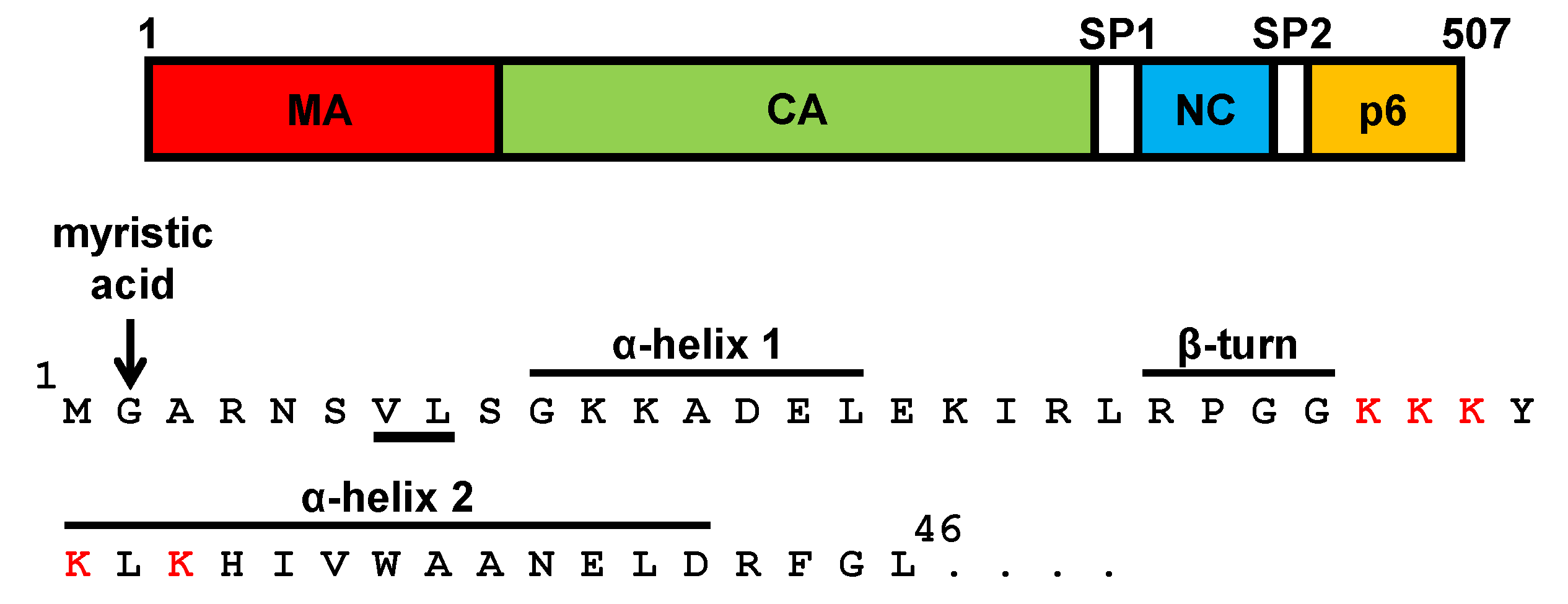
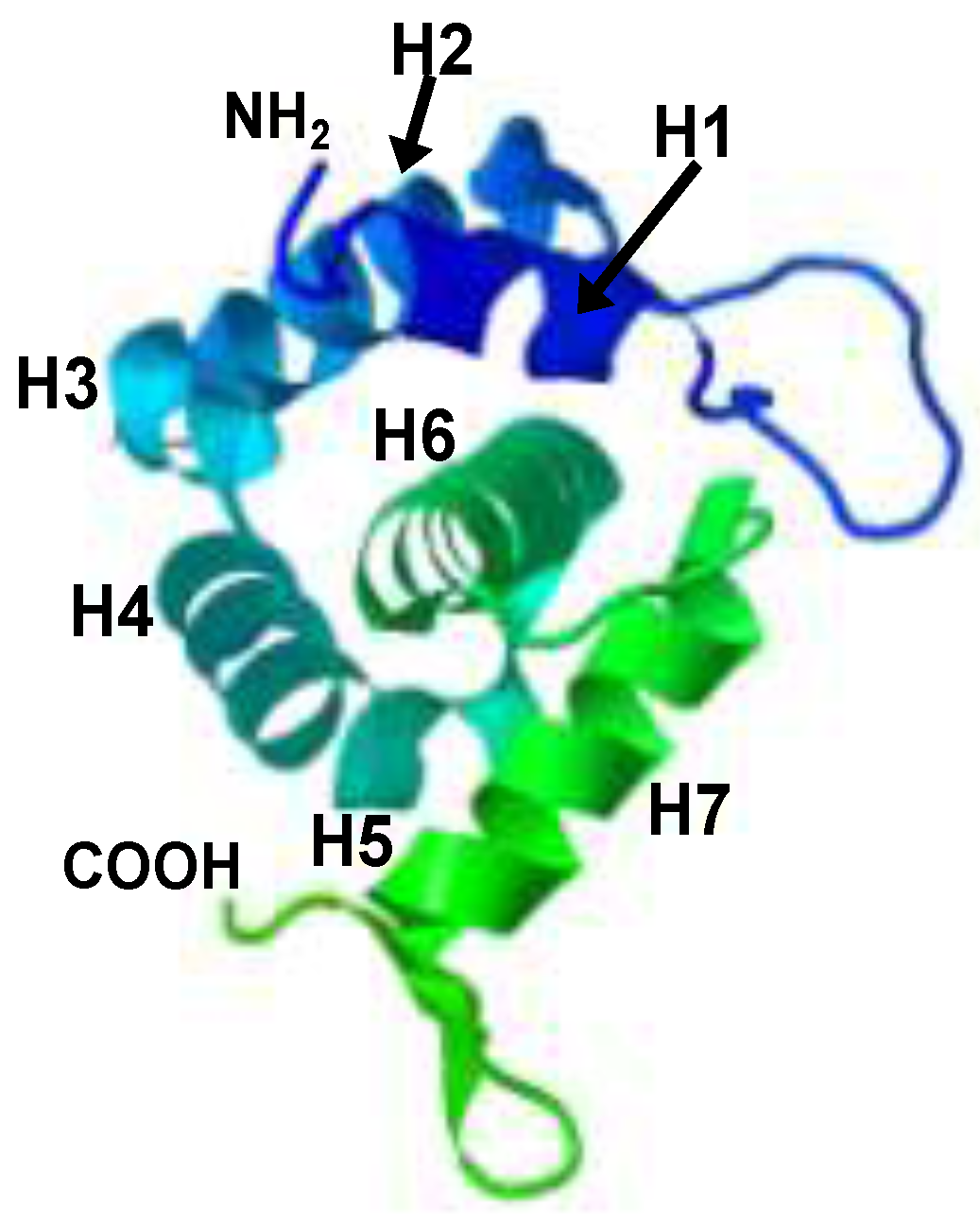
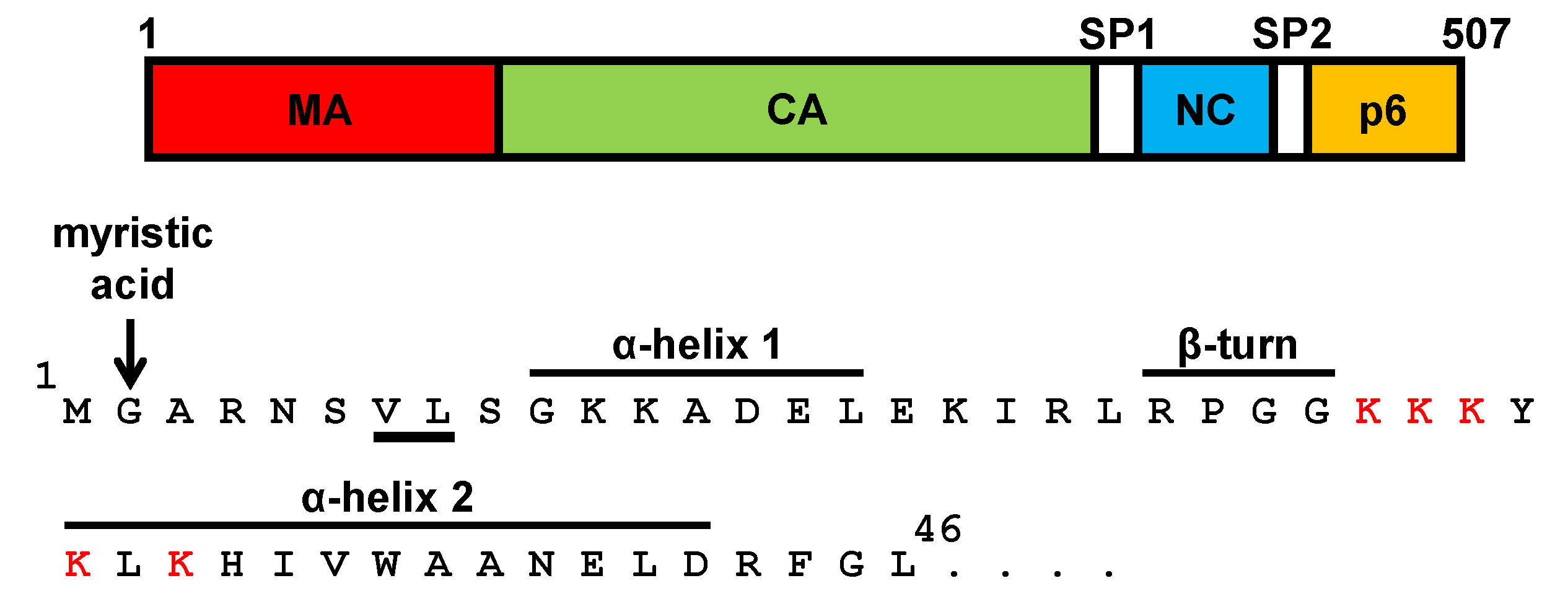
4. The Matrix Domain of SIV Gag
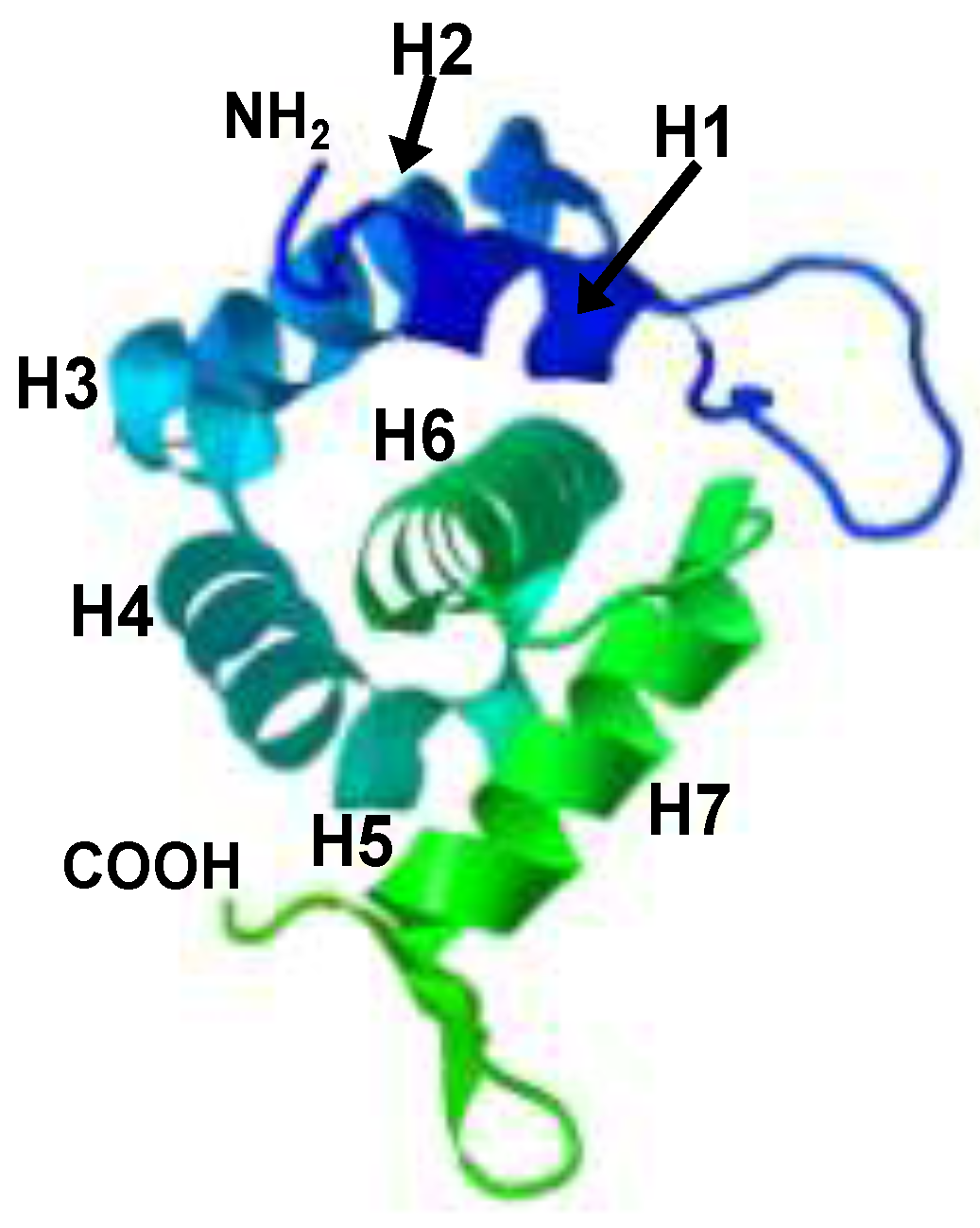
5. Env Incorporation into SIV Virions
The Interplay between the MA Domain of Gag and the Env CD
6. Physical Interaction between the SIV MA and the gp41 CD
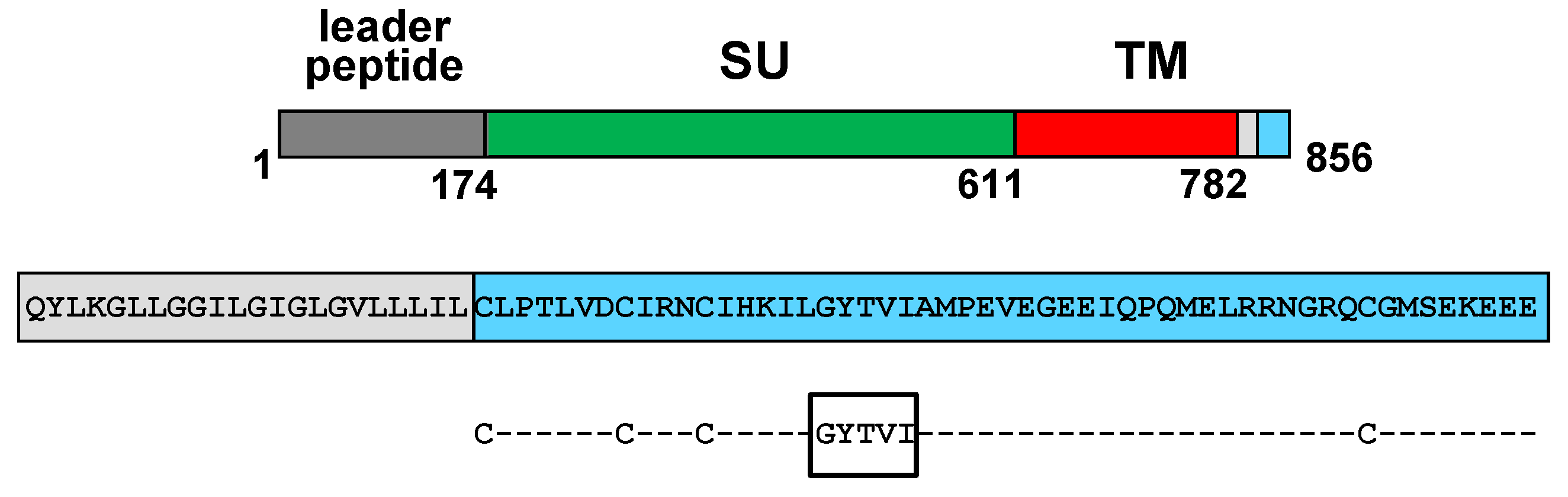
7. Current Knowledge on the Process of FIV Env Incorporation into Virions
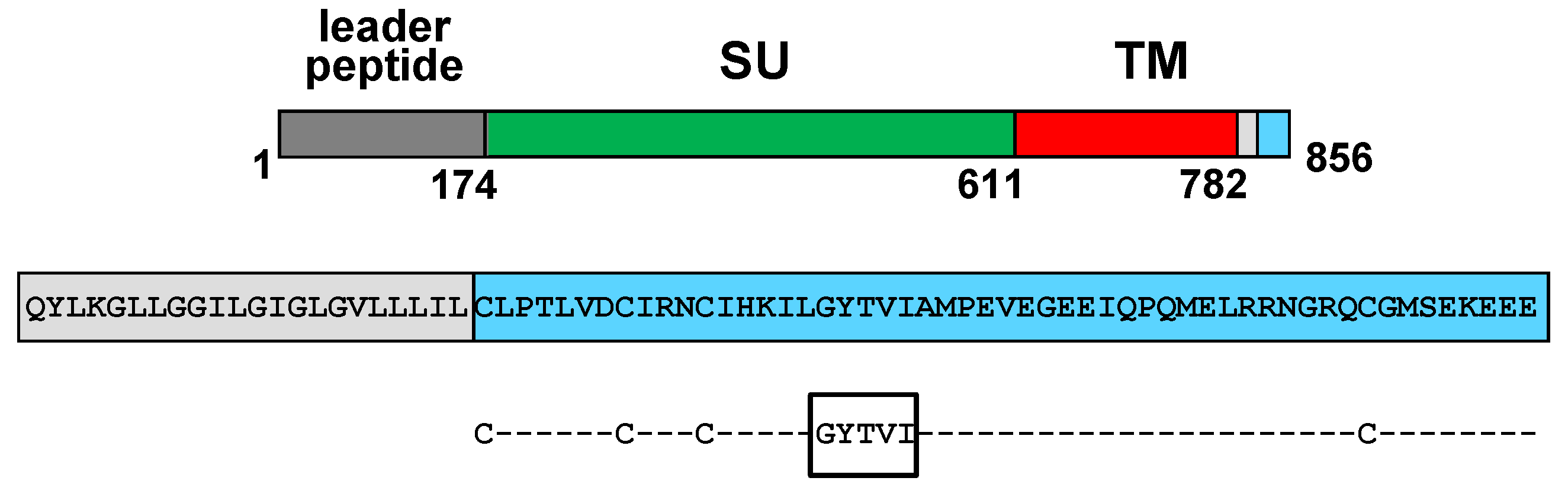
8. Conclusions
Acknowledgments
Conflicts of Interest
References and Notes
- Clements, J.E.; Wong-Staal, F. Molecular biology of lentiviruses. Semin. Virol. 1992, 3, 137–146. [Google Scholar]
- Clements, J.E.; Zink, M.C. Molecular biology and pathogenesis of animal lentivirus infections. Clin. Microbiol. Rev. 1996, 9, 100–117. [Google Scholar]
- Seelamgari, A.; Maddukuri, A.; Berro, R.; de la Fuente, C.; Kehn, K.; Deng, L.; Dadgar, S.; Bottazzi, M.E.; Ghedin, E.; Pumfery, A.; et al. Role of viral regulatory and accessory proteins in HIV-1 replication. Front. Biosci. 2004, 9, 2388–2413. [Google Scholar]
- Gilbert, J.R.; Wong-Staal, F. HIV-2 and SIV vector systems. Somat. Cell Mol. Genet. 2001, 26, 83–98. [Google Scholar] [CrossRef]
- De Parseval, A.; Elder, J.H. Demonstration that ORF2 encodes the feline immunodeficiency virus transactivating (Tat) protein and characterization of a unique gene product with partial rev activity. J. Virol. 1999, 73, 608–617. [Google Scholar]
- Gemeniano, M.C.; Sawai, E.T.; Leutenegger, C.M.; Sparger, E.E. Feline immunodeficiency virus ORF-A is required for virus particle formation and virus infectivity. J. Virol. 2003, 77, 8819–8830. [Google Scholar]
- Gemeniano, M.C.; Sawai, E.T.; Sparger, E.E. Feline immunodeficiency virus ORF-A localizes to the nucleus and induces cell cycle arrest. Virology 2004, 325, 167–174. [Google Scholar] [CrossRef]
- Sundquist, W.I.; Kräusslich, H.-G. HIV-1 Assembly, budding, and maturation. Cold Spring Harb. Perspect. Med. 2012, 2, a006924. [Google Scholar]
- Ganser-Pornillos, B.K.; Yeager, M.; Sundquist, W.I. The structural biology of HIV assembly. Curr. Opin. Struct. Biol. 2008, 18, 203–217. [Google Scholar] [CrossRef]
- Gheysen, D.; Jacobs, E.; de Foresta, F.; Thiriart, C.; Francotte, M.; Thines, D.; de Wilde, M. Assembly and release of HIV-1 precursor Pr55gag virus-like particles from recombinant baculovirus-infected insect cells. Cell 1989, 59, 103–112. [Google Scholar] [CrossRef]
- González, S.A.; Affranchino, J.L.; Gelderblom, H.R.; Burny, A. Assembly of the matrix protein of simian immunodeficiency virus into virus-like particles. Virology 1993, 194, 548–556. [Google Scholar] [CrossRef]
- Manrique, M.L.; Celma, C.C.; González, S.A.; Affranchino, J.L. Mutational analysis of the feline immunodeficiency virus matrix protein. Virus Res. 2001, 76, 103–113. [Google Scholar] [CrossRef]
- Campbell, S.; Rein, A. In vitro assembly properties of human immunodeficiency virus type 1 Gag protein lacking the p6 domain. J. Virol. 1999, 73, 2270–2279. [Google Scholar]
- Huseby, D.; Barklis, R.L.; Alfadhli, A.; Barklis, E. Assembly of human immunodeficiency virus precursor Gag proteins. J. Biol. Chem. 2005, 280, 17664–17670. [Google Scholar] [CrossRef]
- Rauddi, M.L.; Mac Donald, C.L.; Affranchino, J.L.; González, S.A. Mapping of the self-interaction domains in the simian immunodeficiency virus Gag polyprotein. AIDS Res. Hum. Retroviruses 2011, 27, 303–316. [Google Scholar] [CrossRef]
- Affranchino, J.L.; González, S.A. In vitro assembly of feline immunodeficiency virus Gag polyprotein. Virus Res. 2010, 150, 153–157. [Google Scholar] [CrossRef]
- Checkley, M.A.; Luttge, B.G.; Freed, E.O. HIV-1 envelope glycoprotein biosynthesis, trafficking, and incorporation. J. Mol. Biol. 2011, 410, 582–608. [Google Scholar] [CrossRef]
- Johnson, M.C. Mechanisms for Env glycoprotein acquisition by retroviruses. AIDS Res. Hum. Retroviruses 2011, 27, 239–247. [Google Scholar] [CrossRef]
- Postler, T.S.; Desrosiers, R.C. The tale of the long tail: The cytoplasmic domain of HIV-1 gp41. J. Virol. 2013, 87, 2–15. [Google Scholar] [CrossRef]
- Daniel, M.D.; Letvin, N.L.; King, N.W.; Kannagi, M.; Sehgal, P.K.; Hunt, R.D.; Kanki, P.J.; Essex, M.; Desrosiers, R.C. Isolation of a T-cell tropic HTLVIII-like retrovirus from macaques. Science 1985, 228, 1201–1204. [Google Scholar]
- Desrosiers, R.C. Nonhuman Lentiviruses. In Fields Virology, 5th ed.; Knipe, D.M., Howley, P.M., Eds.; Lippincott Williams & Wilkins: Philadelphia, PA, USA, 2007; Volume 2, Chapter 59; pp. 2215–2243. [Google Scholar]
- Letvin, N.L.; King, N.W. Immunologic and pathologic manifestations of the infection of rhesus monkeys with simian immunodeficiency virus of Macaques. J. Acquir. Immune Defic. Syndr. 1990, 3, 1023–1040. [Google Scholar]
- Hirsch, V.M.; Olmsted, R.A.; Murphey-Corb, M.; Purcell, R.H.; Johnson, P.R. An African primate lentivirus (SIVsm) closely related to HIV-2. Nature 1989, 339, 389–392. [Google Scholar] [CrossRef]
- Lemey, P.; Pybus, O.G.; Wang, B.; Saksena, N.K.; Salemi, M.; Vandamme, A.M. Tracing the origin and history of the HIV-2 epidemic. Proc. Natl. Acad. Sci. USA 2003, 100, 6588–6592. [Google Scholar] [CrossRef]
- Gao, F.; Bailes, E.; Robertson, D.L.; Chen, Y.; Rodenburg, C.M.; Michael, S.F.; Cummins, L.B.; Arthur, L.O.; Peeters, M.; Shaw, G.M.; et al. Origin of HIV-1 in the chimpanzee Pan troglodytes troglodytes. Nature 1999, 397, 436–441. [Google Scholar] [CrossRef]
- Keele, B.F.; van Heuverswyn, F.; Li, Y.; Bailes, E.; Takehisa, J.; Santiago, M.L.; Bibollet-Ruche, F.; Chen, Y.; Wain, L.V.; Liegeois, F.; et al. Chimpanzee reservoirs of pandemic and nonpandemic HIV-1. Science 2006, 313, 523–526. [Google Scholar] [CrossRef]
- Veronese, F.D.; Joseph, B.; Copeland, T.D.; Oroszlan, S.; Gallo, R.C.; Sarngadharan, M.G. Identification of Simian immunodeficiency virus SIVMAC env gene products. J. Virol. 1989, 63, 1416–1419. [Google Scholar]
- Zhu, P.; Chertova, E.; Bess, J., Jr.; Lifson, J.D.; Arthur, L.O.; Liu, J.; Taylor, K.A.; Roux, K.H. Electron tomography analysis of envelope glycoprotein trimers on HIV and Simian immunodeficiency virus virions. Proc. Natl. Acad. Sci. USA 2003, 100, 15812–15817. [Google Scholar]
- Sattentau, Q.J.; Clapham, P.R.; Weiss, R.A.; Beverley, P.C.; Montagnier, L.; Alhalabi, M.F.; Gluckmann, J.C.; Klatzmann, D. The human and simian immunodeficiency viruses HIV-1, HIV-2 and SIV interact with similar epitopes on their cellular receptor, the CD4 molecule. AIDS 1988, 2, 101–105. [Google Scholar] [CrossRef]
- Chen, Z.; Zhou, P.; Ho, D.D.; Landau, N.R.; Marx, P.A. Genetically divergent strains of simian immunodeficiency virus use CCR5 as a coreceptor for entry. J. Virol. 1997, 71, 2705–2714. [Google Scholar]
- Edinger, A.L.; Amedee, A.; Miller, K.; Doranz, B.J.; Endres, M.; Sharron, M.; Samson, M.; Lu, Z.H.; Clements, J.E.; Murphey-Corb, M.; et al. Differential utilization of CCR5 by macrophage and T cell tropic simian immunodeficiency virus strains. Proc. Natl. Acad. Sci. USA 1997, 94, 4005–4010. [Google Scholar]
- Sina, S.T.; Ren, W.; Cheng-Mayer, C. Coreceptor use in nonhuman primate models of HIV infection. J. Transl. Med. 2010, 9. [Google Scholar] [CrossRef]
- Liu, H.Y.; Soda, Y.; Shimizu, N.; Haraguchi, Y.; Jinno, A.; Takeuchi, Y.; Hoshino, H. CD4-dependent and CD4-independent utilization of coreceptors by human immunodeficiency viruses type 2 and simian immunodeficiency viruses. Virology 2000, 278, 276–288. [Google Scholar] [CrossRef]
- Lorin, A.; Lins, L.; Stroobant, V.; Brasseur, R.; Charloteaux, B. The minimal fusion peptide of simian immunodeficiency virus corresponds to the 11 first residues of gp32. J. Pept. Sci. 2008, 14, 423–428. [Google Scholar] [CrossRef]
- Malashkevich, V.N.; Chan, D.C.; Chutkowski, C.T.; Kim, P.S. Crystal structure of the simian immunodeficiency virus (SIV) gp41 core: Conserved helical interactions underlie the broad inhibitory activity of gp41 peptides. Proc. Natl. Acad. Sci. USA 1998, 95, 9134–9139. [Google Scholar] [CrossRef]
- Dewhurst, S.; Embretson, J.E.; Anderson, D.C.; Mullins, J.I.; Fultz, P.N. Sequence analysis and acute pathogenicity of molecularly cloned SIVSMM-PBj14. Nature 1990, 345, 636–639. [Google Scholar] [CrossRef]
- Miller, M.A.; Garry, R.F.; Jaynes, J.M.; Montelaro, R.C. A Structural correlation between lentivirus transmembrane proteins and natural cytolytic peptides. AIDS Res. Hum. Retroviruses 1991, 7, 511–519. [Google Scholar] [CrossRef]
- Celma, C.C.P.; Manrique, J.M.; Affranchino, J.L.; Hunter, E.; González, S.A. Domains in the simian immunodeficiency virus gp41 cytoplasmic tail required for envelope incorporation into particles. Virology 2001, 283, 253–261. [Google Scholar] [CrossRef]
- Srinivas, S.K.; Srinivas, R.V.; Anantharamaiah, G.M.; Segrest, J.P.; Compans, R.W. Membrane Interactions of synthetic peptides corresponding to amphipathic helical segments of the human immunodeficiency virus type-1 envelope glycoprotein. J. Biol. Chem. 1992, 267, 7121–7127. [Google Scholar]
- Miller, M.A.; Cloyd, M.D.; Liebmann, J.; Rinaldo, C.R., Jr.; Islam, K.R.; Wang, S.Z.; Mietzner, T.A.; Montelaro, R.C. Alterations in cell membrane permeability by the lentivirus lytic peptide (LLP-1) of HIV-1 transmembrane protein. Virology 1993, 196, 89–100. [Google Scholar] [CrossRef]
- Viard, M.; Ablan, S.D.; Zhou, M.; Veenstra, T.D.; Freed, E.O.; Raviv, Y.; Blumenthal, R. Photoinduced reactivity of the HIV-1 envelope glycoprotein with a membrane-embedded probe reveals insertion of portions of the HIV-1 gp41 cytoplasmic tail into the viral membrane. Biochemistry 2008, 47, 1977–1983. [Google Scholar] [CrossRef]
- Srinivas, S.K.; Srinivas, R.V.; Anantharamaiah, G.M.; Compans, R.W.; Segrest, J.P. Cytosolic domain of the human immunodeficiency virus envelope glycoproteins binds to calmodulin and inhibits calmodulin-regulated proteins. J. Biol. Chem. 1993, 268, 22895–22899. [Google Scholar]
- Yuan, T.; Mietzner, T.A.; Montelaro, R.C.; Vogel, H.J. Characterization of the calmodulin binding domain of SIV transmembrane glycoprotein by NMR and CD spectroscopy. Biochemistry 1995, 34, 10690–10696. [Google Scholar] [CrossRef]
- Ishikawa, H.; Sasaki, M.; Noda, S.; Koga, Y. Apoptosis induction by the binding of the carboxyl terminus of human immunodeficiency virus type 1 gp160 to calmodulin. J. Virol. 1998, 72, 6574–6580. [Google Scholar]
- Micoli, K.J.; Pan, G.; Wu, Y.; Williams, J.P.; Cook, W.J.; McDonald, J.M. Requirement of calmodulin binding by HIV-1 gp160 for enhanced FAS mediated apoptosis. J. Biol. Chem. 2000, 275, 1233–1240. [Google Scholar]
- Beary, T.P.; Tencza, S.B.; Mietzner, T.A.; Montelaro, R.C. Interruption of T-Cell signal transduction by lentivirus lytic peptides from HIV-1 transmembrane protein. J. Pept. Res. 1998, 51, 75–79. [Google Scholar]
- Sauter, M.M.; Pelchen-Matthews, A.; Bron, R.; Marsh, M.; LaBranche, C.C.; Vance, P.J.; Romano, J.; Haggarty, B.S.; Hart, T.K.; Lee, W.M.; et al. An internalization signal in the simian immunodeficiency virus transmembrane protein cytoplasmic domain modulates expression of envelope glycoproteins on the cell surface. J. Cell Biol. 1996, 132, 795–811. [Google Scholar] [CrossRef]
- Bowers, K.; Pelchen-Matthews, A.; Hoening, S.; Vance, P.J.; Creary, L.; Haggarty, B.S.; Romano, J.; Ballensiefen, W.; Hoxie, J.A.; Marsh, M. The simian immunodeficiency virus envelope glycoprotein contains multiple signals that regulate its cell surface expression and endocytosis. Traffic 2000, 1, 661–674. [Google Scholar] [CrossRef]
- Boge, M.; Wyss, S.; Bonifacino, J.S.; Thali, M. A membrane-proximal tyrosine-based signal mediates internalization of the HIV-1 envelope glycoprotein via interaction with the AP-2 clathrin adaptor. J. Biol. Chem. 1998, 273, 15773–15778. [Google Scholar] [CrossRef]
- Byland, R.; Vance, P.J.; Hoxie, J.A.; Marsh, M. A conserved dileucine motif mediates clathrin and AP-2-dependent endocytosis of the HIV-1 envelope protein. Mol. Biol. Cell 2007, 18, 414–425. [Google Scholar]
- Kodama, T.; Wooley, D.P.; Naidu, Y.M.; Kestler, H.W.; Daniel, M.D.; Li, Y.; Desrosiers, R.C. Significance of premature stop codons in env of simian immunodeficiency virus. J. Virol. 1989, 63, 4709–4714. [Google Scholar]
- Chakrabarti, L.; Emerman, M.; Tiollais, P.; Sonigo, P. The Cytoplasmic domain of simian immunodeficiency virus transmembrane protein modulates infectivity. J. Virol. 1989, 63, 4395–4403. [Google Scholar]
- Zingler, K.; Littman, D.R. Truncation of the cytoplasmic domain of the simian immunodeficiency virus envelope glycoprotein increases Env incorporation into particles and fusogenicity and infectivity. J. Virol. 1993, 67, 2824–2831. [Google Scholar]
- Johnston, P.B.; Dubay, J.W.; Hunter, E. Truncations of the simian immunodeficiency virus transmembrane protein confer expanded host range by removing a block to virus entry into cells. J. Virol. 1993, 67, 3077–3086. [Google Scholar]
- Affranchino, J.L.; González, S.A. Mutations at the C-terminus of the simian immunodeficiency virus envelope glycoprotein affect gp120-gp41 stability on virions. Virology 2006, 347, 217–225. [Google Scholar] [CrossRef]
- Henderson, L.E.; Benveniste, R.E.; Sowder, R.; Copeland, T.D.; Schultz, A.M.; Oroszlan, S. Molecular characterization of Gag proteins from simian immunodeficiency virus (SIVMne). J. Virol. 1988, 62, 2587–2595. [Google Scholar]
- Delchambre, M.; Gheysen, D.; Thinès, D.; Thiriart, C.; Jacobs, C.; Verdin, E.; Horth, M.; Burny, A.; Bex, F. The Gag Precursor of the simian immunodeficiency virus assembles into virus-like particles. EMBO J. 1989, 8, 2653–2660. [Google Scholar]
- González, S.A.; Affranchino, J.L. Substitution of leucine 8 in the simian immunodeficiency virus matrix protein impairs particle formation without affecting N-myristylation of the Gag precursor. Virology 1998, 240, 27–35. [Google Scholar] [CrossRef]
- Manrique, M.L.; González, S.A.; Affranchino, J.L. Functional relationship between the matrix proteins of feline and simian immunodeficiency viruses. Virology 2004, 329, 157–167. [Google Scholar] [CrossRef]
- Yuan, X.; Yu, X.; Lee, T.-H.; Essex, M. Mutations in the N-terminal region of human immunodeficiency virus type 1 matrix protein block intracellular transport of the Gag precursor. J. Virol. 1993, 67, 6387–6394. [Google Scholar]
- Zhou, W.; Parent, L.J.; Wills, J.W.; Resh, M.D. Identification of a membrane-binding domain within the amino-terminal region of human immunodeficiency virus type 1 Gag protein which interacts with acidic phospholipids. J. Virol. 1994, 68, 2556–2569. [Google Scholar]
- Ono, A.; Freed, E.O. Binding of human immunodeficiency virus type 1 Gag to membrane: Role of the matrix amino terminus. J. Virol. 1999, 73, 4136–4144. [Google Scholar]
- Paillart, J.-C.; Göttlinger, H.G. Opposing effects of human immunodeficiency virus type 1 matrix mutations support a myristyl switch model of Gag membrane targeting. J. Virol. 1999, 73, 2604–2612. [Google Scholar]
- Saad, J.S.; Miller, J.; Tai, J.; Kim, A.; Ghanam, R.H.; Summers, M.F. Structural basis for targeting HIV-1 Gag proteins to the plasma membrane for virus assembly. Proc. Natl. Acad. Sci. USA 2006, 103, 11364–11369. [Google Scholar] [CrossRef]
- Alfadhli, A.; Still, A.; Barklis, E. Analysis of human immunodeficiency virus type 1 matrix binding to membranes and nucleic acids. J. Virol. 2009, 83, 12196–12203. [Google Scholar] [CrossRef]
- Chukkapalli, V.; Oh, S.J.; Ono, A. Opposing mechanisms involving RNA and lipids regulate HIV-1 Gag membrane binding through the highly basic region of the matrix domain. Proc. Natl. Acad. Sci. USA 2010, 107, 1600–1605. [Google Scholar] [CrossRef]
- Rao, Z.; Belyaev, A.S.; Fry, E.; Roy, P.; Jones, I.M.; Stuart, D.I. Crystal structure of SIV matrix antigen and implications for virus assembly. Nature 1995, 378, 743–747. [Google Scholar] [CrossRef]
- Hill, C.P.; Worthylake, D.; Bancroft, D.P.; Christensen, A.M.; Sundquist, W.I. Crystal structures of the trimeric human immunodeficiency virus type 1 matrix protein: Implications for membrane association and assembly. Proc. Natl. Acad. Sci. USA 1996, 93, 3099–3104. [Google Scholar] [CrossRef]
- González, S.A.; Affranchino, J.L. Mutational analysis of the conserved cysteine residues in the simian immunodeficiency virus matrix protein. Virology 1995, 210, 501–507. [Google Scholar] [CrossRef]
- González, S.A.; Burny, A.; Affranchino, J.L. Identification of domains in the simian immunodeficiency virus matrix protein essential for assembly and envelope glycoprotein incorporation. J. Virol. 1996, 70, 6384–6389. [Google Scholar]
- Manrique, J.M.; Celma, C.C.P.; Hunter, E.; Affranchino, .J.L.; González, S.A. Positive and negative modulation of virus infectivity and envelope glycoprotein incorporation into virions by amino acid substitutions at the N-terminus of the simian immunodeficiency virus matrix protein. J. Virol. 2003, 77, 10881–10888. [Google Scholar]
- Tang, C.; Ndassa, Y.; Summers, M.F. Structure of the N-terminal 283-residue fragment of the immature HIV-1 Gag polyprotein. Nat. Struct. Biol. 2002, 9, 537–543. [Google Scholar]
- Morikawa, Y.; Hockley, D.J.; Nermut, M.V.; Jones, I.M. Roles of matrix, p2, and N-terminal myristoylation in human immunodeficiency virus type 1 Gag assembly. J. Virol. 2000, 74, 16–23. [Google Scholar] [CrossRef]
- Morikawa, Y.; Zhang, W.H.; Hockley, D.J.; Nermut, M.V.; Jones, I.M. Detection of a trimeric human immunodeficiency virus type 1 Gag intermediate is dependent on sequences in the matrix protein, p17. J. Virol. 1998, 72, 7659–7663. [Google Scholar]
- Alfadhli, A.; Barklis, R.L.; Barklis, E. HIV-1 matrix organizes as a hexamer of trimers on membranes containing phosphatidylinositol-(4,5)-bisphosphate. Virology 2009, 387, 466–472. [Google Scholar] [CrossRef]
- Briggs, J.A.; Riches, J.D.; Glass, B.; Bartonova, V.; Zanetti, G.; Kräusslich, H.G. Structure and assembly of immature HIV. Proc. Natl. Acad. Sci. USA 2009, 106, 11090–11095. [Google Scholar]
- Tedbury, P.R.; Ablan, S.D.; Freed, E.O. Global rescue of defects in HIV-1 envelope glycoprotein incorporation: Implications for matrix structure. PLoS Pathog. 2013, 9, e1003739. [Google Scholar] [CrossRef]
- Manrique, J.M.; Celma, C.C.P.; Affranchino, J.L.; Hunter, E.; González, S.A. Small variations in the length of the cytoplasmic domain of the simian immunodeficiency virus transmembrane protein drastically affect envelope incorporation and virus entry. AIDS Res. Hum. Retroviruses 2001, 17, 1615–1624. [Google Scholar] [CrossRef]
- Mammano, F.; Kondo, E.; Sodroski, J.; Bukovsky, A.; Göttlinger, H.G. Rescue of human immunodeficiency virus type 1 matrix protein mutants by envelope glycoproteins with short cytoplasmic domains. J. Virol. 1995, 69, 3824–3830. [Google Scholar]
- Freed, E.O.; Martin, M.A. Virion incorporation of envelope glycoproteins with long but not short cytoplasmic tails is blocked by specific, single amino acid substitutions in the human immunodeficiency virus type 1 matrix. J. Virol. 1995, 69, 1984–1989. [Google Scholar]
- Celma, C.C.P.; Manrique, J.M.; Hunter, E.; Affranchino, J.L.; González, S.A. Second-Site revertants of a simian immunodeficiency virus gp41 mutant defective in envelope glycoprotein incorporation. AIDS Res. Hum. Retroviruses 2004, 20, 731–739. [Google Scholar]
- Yu, X.; Yuan, X.; Matsuda, Z.; Lee, T.-H.; Essex, M. The matrix protein of human immunodeficiency virus type 1 is required for incorporation of viral envelope protein into mature virions. J. Virol. 1992, 66, 4966–4971. [Google Scholar]
- Dorfman, T.; Mammano, F.; Haseltine, W.A.; Göttlinger, H.G. Role of the matrix protein in the virion association of the human immunodeficiency virus type 1 envelope glycoprotein. J. Virol. 1994, 68, 1689–1696. [Google Scholar]
- Freed, E.O.; Martin, M.A. Domains of the human immunodeficiency virus type 1 matrix and gp41 cytoplasmic tail required for envelope incorporation into virions. J. Virol. 1996, 70, 341–351. [Google Scholar]
- Murakami, T.; Freed, E.O. Genetic evidence for an interaction between human immunodeficiency virus type 1 matrix and α-helix 2 of the gp41 cytoplasmic tail. J. Virol. 2000, 74, 3548–3554. [Google Scholar] [CrossRef]
- Wyma, D.J.; Kotov, A.; Aiken, C. Evidence for a stable interaction of gp41 with Pr55(Gag) in immature human immunodeficiency virus type 1 particles. J. Virol. 2000, 74, 9381–9387. [Google Scholar] [CrossRef]
- Kalia, V.; Sarkar, S.; Gupta, P.; Montelaro, R.C. Rational site-directed mutations of the LLP-1 and LLP-2 lentivirus lytic peptide domains in the intracytoplasmic tail of human immunodeficiency virus type 1 gp41 indicate common functions in cell-cell fusion but distinct roles in virion envelope incorporation. J. Virol. 2003, 77, 3634–3646. [Google Scholar] [CrossRef]
- Manrique, J.M.; Affranchino, J.L.; González, S.A. In vitro binding of the simian immunodeficiency virus matrix protein to the cytoplasmic domain of the envelope glycoprotein. Virology 2008, 373, 273–279. [Google Scholar]
- Li, J.; Lord, C.I.; Haseltine, W.; Letvin, N.L.; Sodroski, J. Infection of cynomologus monkeys with a chimeric HIV-1/SIVmac virus that expresses the HIV-1 envelope glycoproteins. J. Acquir. Immune. Defic. Syndr. 1992, 5, 639–646. [Google Scholar]
- Lopez-Vergès, S.; Camus, G.; Blot, G.; Beauvoir, R.; Benarous, R.; Berlioz-Torrent, C. Tail-interacting protein TIP47 is a connector between Gag and Env and is required for Env incorporation into HIV-1 virions. Proc. Natl. Acad. Sci. USA 2006, 103, 14947–14952. [Google Scholar]
- Checkley, M.A.; Luttge, B.G.; Mercredi, P.Y.; Kyere, S.K.; Donlan, J.; Murakami, T.; Summers, M.F.; Cocklin, S.; Freed, E.O. Reevaluation of the requirement for TIP47 in human immunodeficiency virus type 1 envelope glycoprotein incorporation. J. Virol. 2013, 87, 3561–3570. [Google Scholar] [CrossRef]
- Pedersen, N.C.; Ho, E.W.; Brown, M.L.; Yamamoto, J.K. Isolation of a T-lymphotropic virus from domestic cats with an immunodeficiency-like syndrome. Science 1987, 235, 790–793. [Google Scholar]
- Brunner, D.; Pedersen, N.C. Infection of peritoneal macrophages in vitro and in vivo with feline immunodeficiency virus. J. Virol. 1989, 63, 5483–5488. [Google Scholar]
- Brown, W.C.; Bissey, L.; Logan, K.S.; Pedersen, N.C.; Elder, J.H.; Collisson, E.W. Feline immunodeficiency virus infects both CD4+ and CD8+ T lymphocytes. J. Virol. 1991, 65, 3359–3364. [Google Scholar]
- English, R.V.; Johnson, C.M.; Gebhard, D.H.; Tompkins, M.B. In vivo lymphocyte tropism of feline immunodeficiency virus. J. Virol. 1993, 67, 5175–5186. [Google Scholar]
- De Parseval, A.; Chatterji, U.; Sun, P.; Elder, J.H. Feline immunodeficiency virus targets activated CD4+ T cells by using CD134 as a binding receptor. Proc. Natl. Acad. Sci. USA 2004, 101, 13044–13049. [Google Scholar]
- Shimojima, M.; Miyasawa, T.; Ikeda, Y.; McMonagle, E.L.; Haining, H.; Akashi, H.; Takeuchi, Y.; Hosie, M.J.; Willett, B.J. Use of CD134 as a primary receptor by the feline immunodeficiency virus. Science 2004, 303, 1192–1195. [Google Scholar] [CrossRef]
- Poeschla, E.M.; Looney, D.J. CXCR4 is required by a nonprimate lentivirus: Heterologous expression of feline immunodeficiency virus in human, rodent, and feline cells. J. Virol. 1998, 72, 6858–6866. [Google Scholar]
- Verschoor, E.J.; Hulskotte, E.G.J.; Ederveen, J.; Koolen, M.J.M.; Horzinek, M.C.; Rottier, P.J.M. Post-translational processing of the feline immunodeficiency virus envelope precursor protein. Virology 1993, 193, 433–438. [Google Scholar] [CrossRef]
- De Parseval, A.; Elder, J.H. Binding of recombinant feline immunodeficiency virus surface glycoprotein to feline cells: Role of CXCR4, cell-surface heparans, and an unidentified non-CXCR4 receptor. J. Virol. 2001, 75, 4528–4539. [Google Scholar] [CrossRef]
- Garg, H.; Fuller, F.J.; Tompkins, W.A.F. Mechanism of feline immunodeficiency virus envelope glycoprotein-mediated fusion. Virology 2004, 321, 274–286. [Google Scholar] [CrossRef]
- Giannecchini, S.; Bonci, F.; Pistello, M.; Matteucci, D.; Sichi, O.; Rovero, P.; Bendinelli, M. The membrane-proximal tryptophan-rich region in the transmembrane glycoprotein ectodomain of feline immunodeficiency virus is important for cell entry. Virology 2004, 320, 156–166. [Google Scholar] [CrossRef]
- Sundstrom, M.; White, R.L.; de Parseval, A.; Sastry, K.J.; Morris, G.; Grant, C.K.; Elder, J.H. Mapping of the CXCR4 Binding site within variable region 3 of the feline immunodeficiency virus surface glycoprotein. J. Virol. 2008, 82, 9134–9142. [Google Scholar] [CrossRef]
- Hu, Q.Y.; Fink, E.; Hong, Y.; Wang, C.; Grant, C.K.; Elder, J.H. Fine definition of the CXCR4-binding region on the V3 loop of feline immunodeficiency virus surface glycoprotein. PLoS One 2010, 5, e10689. [Google Scholar]
- González, S.A.; Falcón, J.I.; Affranchino, J.L. Replacement of the V3 domain in the surface subunit of the feline immunodeficiency virus envelope glycoprotein with the equivalent region of a T cell-tropic human immunodeficiency virus type 1 results in a chimeric surface protein that efficiently binds to CXCR4. AIDS Res. Hum. Retroviruses 2013. [Google Scholar] [CrossRef]
- Celma, C.C.P.; Paladino, M.G.; González, S.A.; Affranchino, J.L. Importance of the short cytoplasmic domain of feline immunodeficiency virus transmembrane glycoprotein for fusion activity and envelope glycoprotein incorporation into virions. Virology 2007, 366, 405–414. [Google Scholar] [CrossRef]
- Wyss, S.; Dimitrov, A.S.; Baribaud, F.; Edwards, T.G.; Blumenthal, R.; Hoxie, J.A. regulation of human immunodeficiency virus type 1 envelope glycoprotein fusion by a membrane-interactive domain in the gp41 cytoplasmic tail. J. Virol. 2005, 79, 12231–12241. [Google Scholar] [CrossRef]
- Edwards, T.G.; Wyss, S.; Reeves, J.D.; Zolla-Pazner, S.; Hoxie, J.A.; Doms, R.W.; Baribaud, F. Truncation of the cytoplasmic domain induces exposure of conserved regions in the ectodomain of human immunodeficiency virus type 1 envelope glycoprotein. J. Virol. 2002, 76, 2683–2691. [Google Scholar] [CrossRef]
- Kalia, V.; Sarkar, S.; Gupta, P.; Montelaro, R.C. Antibody neutralization escape mediated by point mutations in the intracytoplasmic tail of human immunodeficiency virus type 1 gp41. J. Virol. 2005, 79, 2097–2107. [Google Scholar]
- Poignard, P.; Moulard, M.; Golez, E.; Vivona, V.; Franti, M.; Venturini, S.; Wang, M.; Parren, P.W.H.I.; Burton, D.R. Heterogeneity of envelope molecules expressed on primary human immunodeficiency virus type 1 particles as probed by the binding of neutralizing and nonneutralizing antibodies. J. Virol. 2003, 77, 353–365. [Google Scholar] [CrossRef]
- Moore, P.N.; Crooks, E.T.; Porter, L.; Zhu, P.; Cayanan, C.S.; Grise, H.; Corcoran, P.; Zwick, M.B.; Franti, M.; Morris, L.; et al. Nature of nonfunctional envelope proteins on the surface of human immunodeficiency virus type 1. J. Virol. 2006, 80, 2515–2528. [Google Scholar] [CrossRef]
- González, S.A.; Paladino, M.G.; Affranchino, J.L. Palmitoylation of the feline immunodeficiency virus envelope glycoprotein and its effect on fusion activity and envelope incorporation into virions. Virology 2012, 428, 1–10. [Google Scholar] [CrossRef]
- Salaun, C.; Greaves, J.; Chamberlain, L.H. The intracellular dynamic of protein palmitoylation. J. Cell Biol. 2010, 191, 1229–1238. [Google Scholar] [CrossRef]
- Lindwasser, O.W.; Resh, M.D. Multimerization of human immunodeficiency virus type 1 Gag promotes its localization to barges, raft-like membrane microdomains. J. Virol. 2001, 75, 7913–7924. [Google Scholar] [CrossRef]
- Nguyen, D.H.; Hildreth, J.E. Evidence for budding of human immunodeficiency virus type 1 selectively from glycolipid-enriched membrane lipid rafts. J. Virol. 2000, 74, 3264–3272. [Google Scholar] [CrossRef]
- Ono, A.; Freed, E.O. Plasma membrane rafts play a critical role in HIV-1 assembly and release. Proc. Natl. Acad. Sci. USA 2001, 98, 13925–13930. [Google Scholar]
- Brown, D.A.; Rose, J.K. Sorting of GPI-anchored proteins to glycolipid-enriched membrane subdomains during transport to the apical cell surface. Cell 1992, 68, 533–544. [Google Scholar] [CrossRef]
- Yang, C.; Spies, C.P.; Compans, R.W. The human and simian immunodeficiency virus envelope glycoprotein transmembrane subunits are palmitoylated. Proc. Natl. Acad. Sci. USA 1995, 92, 9871–9875. [Google Scholar] [CrossRef]
- Rousso, I.; Mixon, M.B.; Chen, B.K.; Kim, P.S. Palmitoylation of the HIV-1 envelope glycoprotein is critical for viral infectivity. Proc. Natl. Acad. Sci. USA 2000, 97, 13523–13525. [Google Scholar] [CrossRef]
- Bhattacharya, J.; Peters, P.J.; Clapham, P.R. Human immunodeficiency virus type 1 envelope glycoproteins that lack cytoplasmic domain cysteines: Impact on association with membrane lipid rafts and incorporation onto budding virus particles. J. Virol. 2004, 78, 5500–5506. [Google Scholar] [CrossRef]
- Chan, W.E.; Lin, H.H.; Chen, S.S. Wild-Type viral replication potential of human immunodeficiency virus type 1 envelope mutants lacking palmitoylation signals. J. Virol. 2005, 79, 8374–8387. [Google Scholar] [CrossRef]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Affranchino, J.L.; González, S.A. Understanding the Process of Envelope Glycoprotein Incorporation into Virions in Simian and Feline Immunodeficiency Viruses. Viruses 2014, 6, 264-283. https://doi.org/10.3390/v6010264
Affranchino JL, González SA. Understanding the Process of Envelope Glycoprotein Incorporation into Virions in Simian and Feline Immunodeficiency Viruses. Viruses. 2014; 6(1):264-283. https://doi.org/10.3390/v6010264
Chicago/Turabian StyleAffranchino, José L., and Silvia A. González. 2014. "Understanding the Process of Envelope Glycoprotein Incorporation into Virions in Simian and Feline Immunodeficiency Viruses" Viruses 6, no. 1: 264-283. https://doi.org/10.3390/v6010264