Low Copper and High Manganese Levels in Prion Protein Plaques

{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
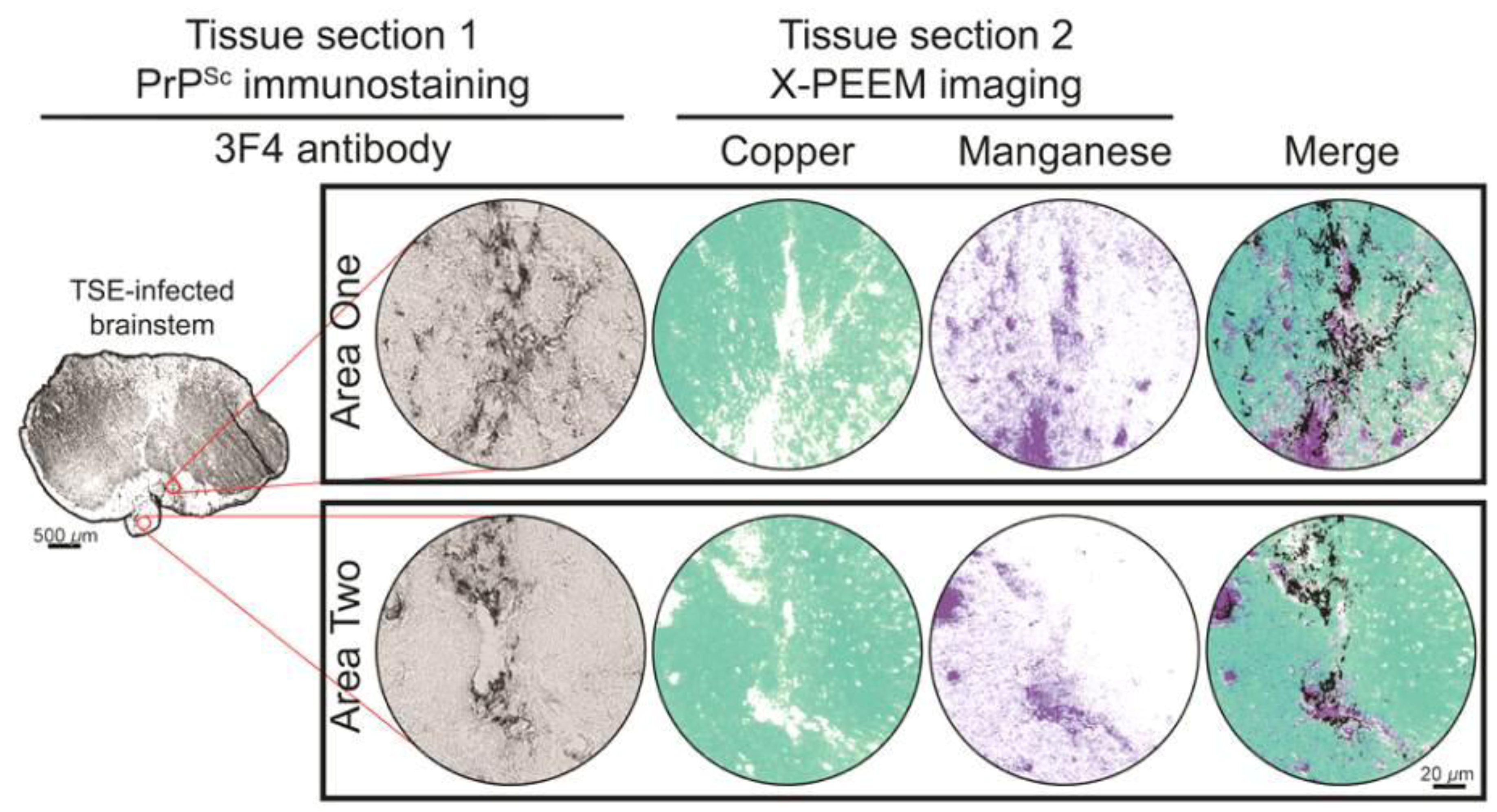
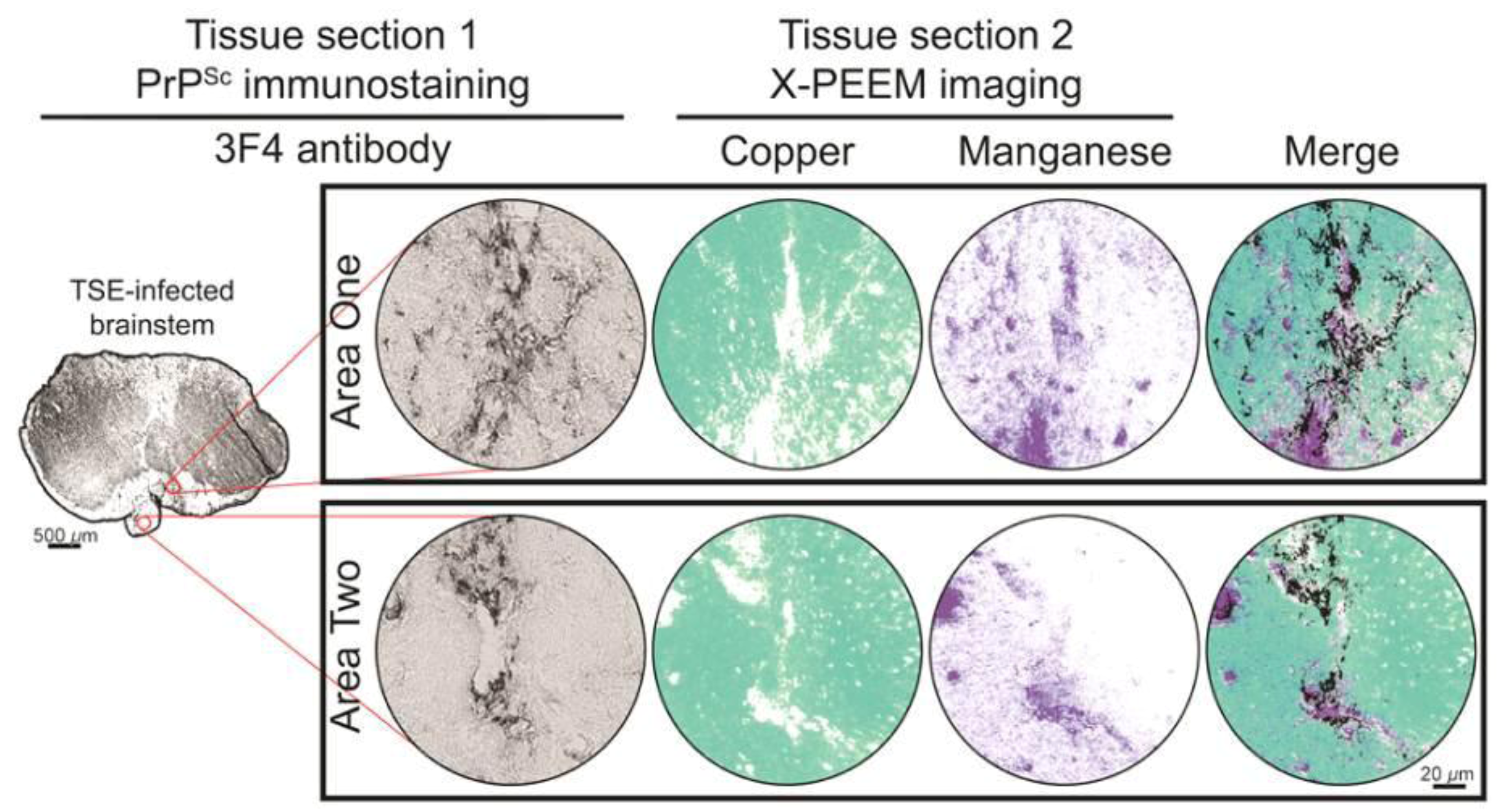
3. Results and Discussion
4. Conclusions
Acknowledgments
Conflict of Interest
References and Notes
- Colby, D.W.; Prusiner, S.B. Prions. Cold Spring Harbor Perspect. Biol. 2011, 3. [Google Scholar] [CrossRef]
- Davies, P.; Brown, D.R. The chemistry of copper binding to PrP: Is there sufficient evidence to elucidate a role for copper in protein function? Biochem. J. 2008, 410, 237–244. [Google Scholar] [CrossRef]
- Brazier, M.W.; Davies, P.; Player, E.; Marken, F.; Viles, J.H.; Brown, D.R. Manganese binding to the prion protein. J. Biol. Chem. 2008, 283, 12831–12839. [Google Scholar]
- Brown, D.R. Prions and manganese: A maddening beast. Metallomics 2011, 3, 229–238. [Google Scholar] [CrossRef]
- Wong, B.S.; Chen, S.G.; Colucci, M.; Xie, Z.; Pan, T.; Liu, T.; Li, R.; Gambetti, P.; Sy, M.S.; Brown, D.R. Aberrant metal binding by prion protein in human prion disease. J. Neurochem. 2001, 78, 1400–1408. [Google Scholar] [CrossRef]
- Thackray, A.M.; Knight, R.; Haswell, S.J.; Bujdoso, R.; Brown, D.R. Metal imbalance and compromised antioxidant function are early changes in prion disease. Biochem. J. 2002, 362, 253–258. [Google Scholar] [CrossRef]
- Mitteregger, G.; Korte, S.; Shakarami, M.; Herms, J.; Kretzschmar, H.A. Role of copper and manganese in prion disease progression. Brain Res. 2009, 1292, 155–164. [Google Scholar] [CrossRef]
- Hortells, P.; Monleon, E.; Acin, C.; Vargas, A.; Vasseur, V.; Salomon, A.; Ryffel, B.; Cesbron, J.Y.; Badiola, J.J.; Monzon, M. The effect of metal imbalances on scrapie neurodegeneration. Zoonoses Public Health 2010, 57, 358–366. [Google Scholar]
- Brazier, M.W.; Volitakis, I.; Kvasnicka, M.; White, A.R.; Underwood, J.R.; Green, J.E.; Han, S.; Hill, A.F.; Masters, C.L.; Collins, S.J. Manganese chelation therapy extends survival in a mouse model of M1000 prion disease. J. Neurochem. 2010, 114, 440–451. [Google Scholar] [CrossRef]
- Bolea, R.; Hortells, P.; Martin-Burriel, I.; Vargas, A.; Ryffel, B.; Monzon, M.; Badiola, J.J. Consequences of dietary manganese and copper imbalance on neuronal apoptosis in a murine model of scrapie. Neuropathol. Appl. Neurobiol. 2010, 36, 300–311. [Google Scholar] [CrossRef]
- Sigurdsson, E.M.; Brown, D.R.; Alim, M.A.; Scholtzova, H.; Carp, R.; Meeker, H.C.; Prelli, F.; Frangione, B.; Wisniewski, T. Copper chelation delays the onset of prion disease. J. Biol. Chem. 2003, 278, 46199–46202. [Google Scholar]
- Wong, B.S.; Brown, D.R.; Pan, T.; Whiteman, M.; Liu, T.; Bu, X.; Li, R.; Gambetti, P.; Olesik, J.; Rubenstein, R.; et al. Oxidative impairment in scrapie-infected mice is associated with brain metals perturbations and altered antioxidant activities. J. Neurochem. 2001, 79, 689–698. [Google Scholar]
- Hesketh, S.; Sassoon, J.; Knight, R.; Brown, D.R. Elevated manganese levels in blood and CNS in human prion disease. Mol. Cell. Neurosci. 2008, 37, 590–598. [Google Scholar] [CrossRef]
- Hesketh, S.; Sassoon, J.; Knight, R.; Hopkins, J.; Brown, D.R. Elevated manganese levels in blood and central nervous system occur before onset of clinical signs in scrapie and bovine spongiform encephalopathy. J. Animal Sci. 2007, 85, 1596–1609. [Google Scholar] [CrossRef]
- Frazer, B.H.; Girasole, M.; Wiese, L.M.; Franz, T.; De Stasio, G. Spectromicroscope for the photoelectron imaging of nanostructures with x-rays (SPHINX): Performance in biology, medicine and geology. Ultramicroscopy 2004, 99, 87–94. [Google Scholar] [CrossRef]
- Frazer, B.H.; Sonderegger, B.R.; Gilbert, B.; Richter, K.L.; Salt, C.; Wiese, L.M.; Rajesh, D.; Howard, S.P.; Fowler, J.F.; Mehta, M.P.; et al. Mapping of physiological and trace elements with X-PEEM. J. Phys. IV 2003, 104, 349–352. [Google Scholar]
- Gilbert, B.; Andres, R.; Perfetti, P.; Margaritondo, G.; Rempfer, G.; De Stasio, G. Charging phenomena in PEEM imaging and spectroscopy. Ultramicroscopy 2000, 83, 129–139. [Google Scholar]
- Bessen, R.A.; Marsh, R.F. Identification of two biologically distinct strains of transmissible mink encephalopathy in hamsters. J. Gen. Virol. 1992, 73, 329–334. [Google Scholar] [CrossRef]
- Hoefert, V.B.; Aiken, J.M.; McKenzie, D.; Johnson, C.J. Labeling of the scrapie-associated prion protein in vitro and in vivo. Neurosci. Lett. 2004, 371, 176–180. [Google Scholar]
- Gilbert, B.; Perfetti, L.; Hansen, R.; Mercanti, D.; Casalbore, P.; Andres, R.; Perfetti, P.; Margaritondo, G.; De Stasio, G. UV-ozone ashing of cells and tissues for spatially resolved trace element analysis. Front. Biosci. 2000, 5, 10–17. [Google Scholar]
- Johnson, C.J.; Gilbert, P.U.; McKenzie, D.; Pedersen, J.A.; Aiken, J.M. Ultraviolet-ozone treatment reduces levels of disease-associated prion protein and prion infectivity. BMC Res. Notes 2009, 2, 121. [Google Scholar] [CrossRef]
- R Development Core Team, R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2012.
- Kort, E. rtiff: A tiff reader for R. version 1.4.1; 2012. [Google Scholar]
- Kim, N.H.; Choi, J.K.; Jeong, B.H.; Kim, J.I.; Kwon, M.S.; Carp, R.I.; Kim, Y.S. Effect of transition metals (mn, cu, fe) and deoxycholic acid (da) on the conversion of PrPC to PrPres. FASEB J. 2005, 19, 783–785. [Google Scholar]
- Singh, A.; Isaac, A.O.; Luo, X.; Mohan, M.L.; Cohen, M.L.; Chen, F.; Kong, Q.; Bartz, J.; Singh, N. Abnormal brain iron homeostasis in human and animal prion disorders. PLoS Pathog. 2009, 5, e1000336. [Google Scholar] [CrossRef]
- Watt, N.T.; Hooper, N.M. The prion protein and neuronal zinc homeostasis. TIBS 2003, 28, 406–410. [Google Scholar]
- Watt, N.T.; Taylor, D.R.; Kerrigan, T.L.; Griffiths, H.H.; Rushworth, J.V.; Whitehouse, I.J.; Hooper, N.M. Prion protein facilitates uptake of zinc into neuronal cells. Nat. Commun. 2012, 3, 1134. [Google Scholar]
- Pushie, M.J.; Pickering, I.J.; Martin, G.R.; Tsutsui, S.; Jirik, F.R.; George, G.N. Prion protein expression level alters regional copper, iron and zinc content in the mouse brain. Metallomics 2011, 3, 206–214. [Google Scholar] [CrossRef]
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Johnson, C.J.; Gilbert, P.U.P.A.; Abrecht, M.; Baldwin, K.L.; Russell, R.E.; Pedersen, J.A.; Aiken, J.M.; McKenzie, D. Low Copper and High Manganese Levels in Prion Protein Plaques. Viruses 2013, 5, 654-662. https://doi.org/10.3390/v5020654
Johnson CJ, Gilbert PUPA, Abrecht M, Baldwin KL, Russell RE, Pedersen JA, Aiken JM, McKenzie D. Low Copper and High Manganese Levels in Prion Protein Plaques. Viruses. 2013; 5(2):654-662. https://doi.org/10.3390/v5020654
Chicago/Turabian StyleJohnson, Christopher J., P.U.P.A. Gilbert, Mike Abrecht, Katherine L. Baldwin, Robin E. Russell, Joel A. Pedersen, Judd M. Aiken, and Debbie McKenzie. 2013. "Low Copper and High Manganese Levels in Prion Protein Plaques" Viruses 5, no. 2: 654-662. https://doi.org/10.3390/v5020654