Insights into the Roles of Cyclophilin A During Influenza Virus Infection

{kind=link}
Abstract
:1. Introduction
2. The Localization and Function of CypA
3. CypA as an Important Host Factor in Viral Infections
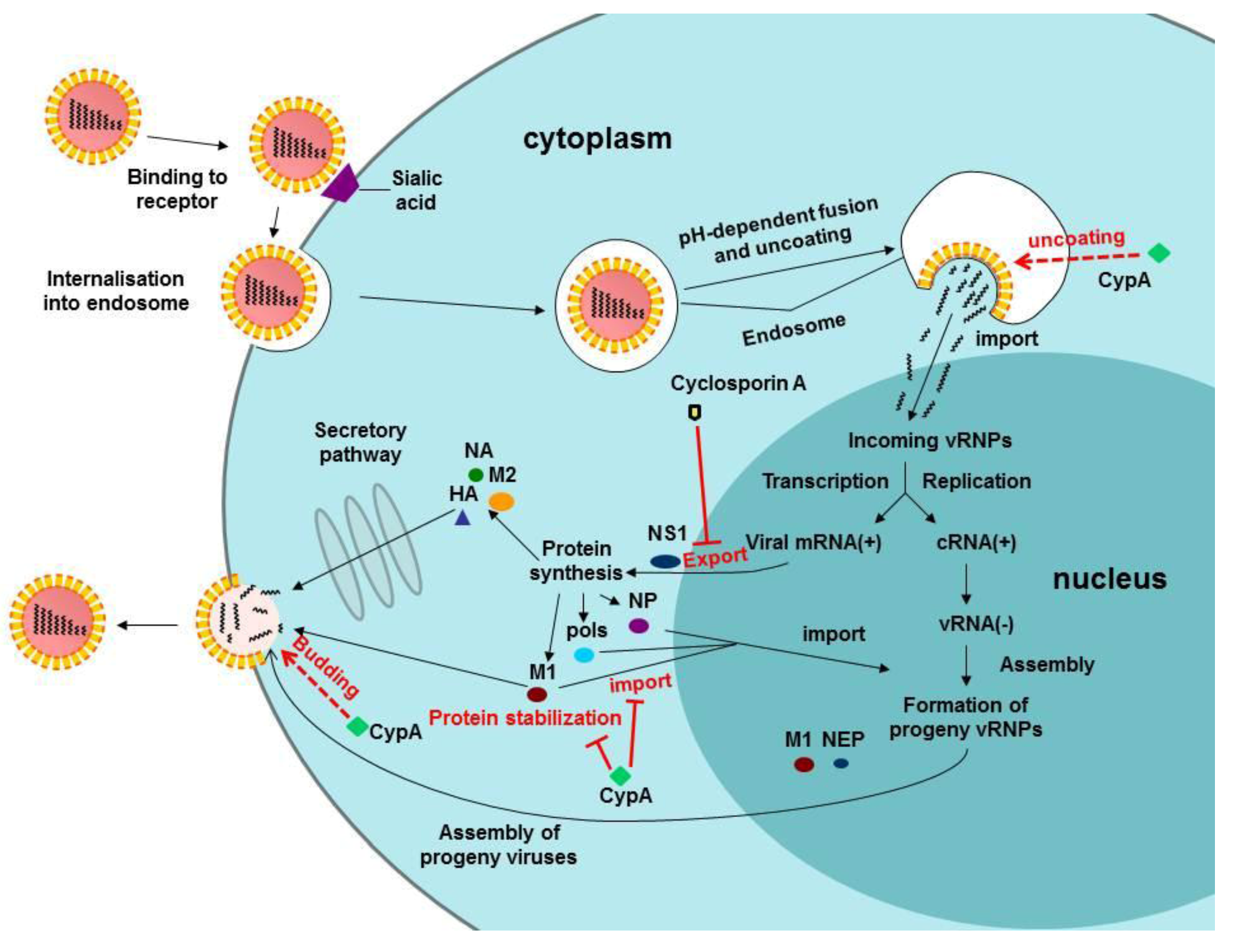
4. CypA and Influenza Virus Infection

Conclusion and Future Directions
Conflict of Interest
Acknowledgements
References and Notes
- Wise, H.M.; Foeglein, A.; Sun, J.; Dalton, R.M.; Patel, S.; Howard, W.; Anderson, E.C.; Barclay, W.S.; Digard, P. A complicated message: Identification of a novel PB1-related protein translated from influenza A virus segment 2 mRNA. J. Virol. 2009, 83, 8021–8031. [Google Scholar]
- Chen, W.; Calvo, P.A.; Malide, D.; Gibbs, J.; Schubert, U.; Bacik, I.; Basta, S.; O'Neill, R.; Schickli, J.; Palese, P.; et al. A novel influenza A virus mitochondrial protein that induces cell death. Nature Medicine 2001, 7, 1306–1312. [Google Scholar] [CrossRef]
- Wise, H.M.; Hutchinson, E.C.; Jagger, B.W.; Stuart, A.D.; Kang, Z.H.; Robb, N.; Schwartzman, L.M.; Kash, J.C.; Fodor, E.; Firth, A.E.; et al. Identification of a novel splice variant form of the influenza a virus m2 ion channel with an antigenically distinct ectodomain. PLoS Pathog. 2012, 8, e1002998. [Google Scholar] [CrossRef]
- Jagger, B.W.; Wise, H.M.; Kash, J.C.; Walters, K.A.; Wills, N.M.; Xiao, Y.L.; Dunfee, R.L.; Schwartzman, L.M.; Ozinsky, A.; Bell, G.L.; et al. An overlapping protein-coding region in influenza A virus segment 3 modulates the host response. Science 2012, 337, 199–204. [Google Scholar] [CrossRef]
- Zhirnov, O.P.; Okhuchi, M.; Avakiants, V.S.; Ovcharenko, A.V.; Klenk Kh, D. Interaction of influenza virus matrix protein M1 with histones. Mol. Biol. (Mosk.) 1997, 31, 137–143. [Google Scholar]
- Reinhardt, J.; Wolff, T. The influenza A virus M1 protein interacts with the cellular receptor of activated C kinase (RACK) 1 and can be phosphorylated by protein kinase C. Vet. Microbiol. 2000, 74, 87–100. [Google Scholar] [CrossRef]
- Garcia-Robles, I.; Akarsu, H.; Muller, C.W.; Ruigrok, R.W.; Baudin, F. Interaction of influenza virus proteins with nucleosomes. Virology 2005, 332, 329–336. [Google Scholar] [CrossRef]
- Watanabe, K.; Fuse, T.; Asano, I.; Tsukahara, F.; Maru, Y.; Nagata, K.; Kitazato, K.; Kobayashi, N. Identification of Hsc70 as an influenza virus matrix protein (M1) binding factor involved in the virus lifecycle. FEBS letters 2006, 580, 5785–5790. [Google Scholar] [CrossRef]
- Liu, X.; Sun, L.; Yu, M.; Wang, Z.; Xu, C.; Xue, Q.; Zhang, K.; Ye, X.; Kitamura, Y.; Liu, W. Cyclophilin A interacts with influenza A virus M1 protein and impairs the early stage of the viral replication. Cell Microbiol. 2009, 11, 730–741. [Google Scholar] [CrossRef]
- Xu, C.; Meng, S.; Liu, X.; Sun, L.; Liu, W. Chicken cyclophilin A is an inhibitory factor to influenza virus replication. Virol. J. 2010, 7, 372. [Google Scholar] [CrossRef]
- Liu, X.; Zhao, Z.; Xu, C.; Sun, L.; Chen, J.; Zhang, L.; Liu, W. Cyclophilin A restricts influenza A virus replication through degradation of the M1 protein. PLoS One 2012, 7, e31063. [Google Scholar]
- Liu, X.; Zhao, Z.; Li, Z.; Xu, C.; Sun, L.; Chen, J.; Liu, W. Cyclosporin A inhibits the influenza virus replication through cyclophilin A-dependent and -independent pathways. PLoS One 2012, 7, e37277. [Google Scholar]
- Braaten, D.; Luban, J. Cyclophilin A regulates HIV-1 infectivity, as demonstrated by gene targeting in human T cells. EMBO J. 2001, 20, 1300–1309. [Google Scholar] [CrossRef]
- Bose, S.; Mathur, M.; Bates, P.; Joshi, N.; Banerjee, A.K. Requirement for cyclophilin A for the replication of vesicular stomatitis virus New Jersey serotype. J. Gen. Virol. 2003, 84, 1687–1699. [Google Scholar] [CrossRef]
- Castro, A.P.; Carvalho, T.M.; Moussatche, N.; Damaso, C.R. Redistribution of cyclophilin A to viral factories during vaccinia virus infection and its incorporation into mature particles. J. Virol. 2003, 77, 9052–9068. [Google Scholar] [CrossRef]
- Towers, G.J.; Hatziioannou, T.; Cowan, S.; Goff, S.P.; Luban, J.; Bieniasz, P.D. Cyclophilin A modulates the sensitivity of HIV-1 to host restriction factors. Nature Medicine 2003, 9, 1138–1143. [Google Scholar] [CrossRef]
- Luban, J. Cyclophilin A, TRIM5 and resistance to human immunodeficiency virus type 1 infection. J. Virol. 2007, 81, 1054–1061. [Google Scholar] [CrossRef]
- Yang, F.; Robotham, J.M.; Nelson, H.B.; Irsigler, A.; Kenworthy, R.; Tang, H. Cyclophilin A is an essential cofactor for hepatitis C virus infection and the principal mediator of cyclosporine resistance in vitro. J. Virol. 2008, 82, 5269–5278. [Google Scholar]
- Tian, X.; Zhao, C.; Zhu, H.; She, W.; Zhang, J.; Liu, J.; Li, L.; Zheng, S.; Wen, Y.M.; Xie, Y. Hepatitis B virus (HBV) surface antigen interacts with and promotes cyclophilin a secretion: Possible link to pathogenesis of HBV infection. J. Virol. 2010, 84, 3373–3381. [Google Scholar] [CrossRef]
- Galat, A. Peptidylproline cis-trans-isomerases: immunophilins. Eur. J. Biochem. 1993, 216, 689–707. [Google Scholar] [CrossRef]
- Gothel, S.F.; Marahiel, M.A. Peptidyl-prolyl cis-trans isomerases, a superfamily of ubiquitous folding catalysts. Cell Mol. Life Sci. 1999, 55, 423–436. [Google Scholar] [CrossRef]
- Liu, J.; Albers, M.W.; Wandless, T.J.; Luan, S.; Alberg, D.G.; Belshaw, P.J.; Cohen, P.; MacKintosh, C.; Klee, C.B.; Schreiber, S.L. Inhibition of T cell signaling by immunophilin-ligand complexes correlates with loss of calcineurin phosphatase activity. Biochemistry 1992, 31, 3896–3901. [Google Scholar]
- Sherry, B.; Yarlett, N.; Strupp, A.; Cerami, A. Identification of cyclophilin as a proinflammatory secretory product of lipopolysaccharide-activated macrophages. P. Natl. Acad. Sci. USA 1992, 89, 3511–3515. [Google Scholar] [CrossRef]
- Xu, Q.; Leiva, M.C.; Fischkoff, S.A.; Handschumacher, R.E.; Lyttle, C.R. Leukocyte chemotactic activity of cyclophilin. J. Biol. Chem. 1992, 267, 11968–11971. [Google Scholar]
- Kirsch, A.H.; Diaz, L.A., Jr.; Bonish, B.; Antony, P.A.; Fox, D.A. The pattern of expression of CD147/neurothelin during human T-cell ontogeny as defined by the monoclonal antibody 8D6. Tissue Antigens 1997, 50, 147–152. [Google Scholar] [CrossRef]
- Mutin, M.; Dignat-George, F.; Sampol, J. Immunologic phenotype of cultured endothelial cells: Quantitative analysis of cell surface molecules. Tissue Antigens 1997, 50, 449–458. [Google Scholar] [CrossRef]
- Inoue, H.; Sawada, M.; Ryo, A.; Tanahashi, H.; Wakatsuki, T.; Hada, A.; Kondoh, N.; Nakagaki, K.; Takahashi, K.; Suzumura, A.; et al. Serial analysis of gene expression in a microglial cell line. Glia 1999, 28, 265–271. [Google Scholar] [CrossRef]
- Kasinrerk, W.; Tokrasinwit, N.; Phunpae, P. CD147 monoclonal antibodies induce homotypic cell aggregation of monocytic cell line U937 via LFA-1/ICAM-1 pathway. Immunology 1999, 96, 184–192. [Google Scholar] [CrossRef]
- Wimazal, F.; Ghannadan, M.; Muller, M.R.; End, A.; Willheim, M.; Meidlinger, P.; Schernthaner, G.H.; Jordan, J.H.; Hagen, W.; Agis, H.; et al. Expression of homing receptors and related molecules on human mast cells and basophils: A comparative analysis using multi-color flow cytometry and toluidine blue/immunofluorescence staining techniques. Tissue Antigens 1999, 54, 499–507. [Google Scholar] [CrossRef]
- He, H.; Zhou, D.; Fan, W.; Fu, X.; Zhang, J.; Shen, Z.; Li, J.; Wu, Y. Cyclophilin A inhibits rotavirus replication by facilitating host IFN-I production. Biochem. Biophys. Res. Commun. 2012, 422, 664–669. [Google Scholar] [CrossRef]
- Zhou, D.; Mei, Q.; Li, J.; He, H. Cyclophilin A and viral infections. Biochem. Biophys. Res. Commun. 2012, 424, 647–650. [Google Scholar] [CrossRef]
- Shaw, M.L.; Stone, K.L.; Colangelo, C.M.; Gulcicek, E.E.; Palese, P. Cellular proteins in influenza virus particles. PLoS Pathog. 2008, 4, e1000085. [Google Scholar] [CrossRef]
- Franke, E.K.; Yuan, H.E.; Luban, J. Specific incorporation of cyclophilin A into HIV-1 virions. Nature 1994, 372, 359–362. [Google Scholar] [CrossRef]
- An, P.; Wang, L.H.; Hutcheson-Dilks, H.; Nelson, G.; Donfield, S.; Goedert, J.J.; Rinaldo, C.R.; Buchbinder, S.; Kirk, G.D.; O'Brien, S.J.; et al. egulatory polymorphisms in the cyclophilin A gene, PPIA, accelerate progression to AIDS. PLoS Pathog. 2007, 3, e88. [Google Scholar] [CrossRef] [Green Version]
- Colgan, J.; Yuan, H.E.; Franke, E.K.; Luban, J. Binding of the human immunodeficiency virus type 1 Gag polyprotein to cyclophilin A is mediated by the central region of capsid and requires Gag dimerization. J. Virol. 1996, 70, 4299–4310. [Google Scholar]
- Manel, N.; Hogstad, B.; Wang, Y.; Levy, D.E.; Unutmaz, D.; Littman, D.R. A cryptic sensor for HIV-1 activates antiviral innate immunity in dendritic cells. Nature 2010, 467, 214–217. [Google Scholar]
- Zander, K.; Sherman, M.P.; Tessmer, U.; Bruns, K.; Wray, V.; Prechtel, A.T.; Schubert, E.; Henklein, P.; Luban, J.; et al. Cyclophilin A interacts with HIV-1 Vpr and is required for its functional expression. J. Biol. Chem. 2003, 278, 43202–43213. [Google Scholar]
- Bruns, K.; Fossen, T.; Wray, V.; Henklein, P.; Tessmer, U.; Schubert, U. Structural characterization of the HIV-1 Vpr N terminus: Evidence of cis/trans-proline isomerism. J. Biol. Chem. 2003, 278, 43188–43201. [Google Scholar]
- Solbak, S.M.; Reksten, T.R.; Roder, R.; Wray, V.; Horvli, O.; Raae, A.J.; Henklein, P.; Fossen, T. HIV-1 p6-Another viral interaction partner to the host cellular protein cyclophilin A. Biochim. Biophys. Acta 2012, 1824, 667–678. [Google Scholar] [CrossRef]
- Solbak, S.M.; Wray, V.; Horvli, O.; Raae, A.J.; Flydal, M.I.; Henklein, P.; Nimtz, M.; Schubert, U.; Fossen, T. The host-pathogen interaction of human cyclophilin A and HIV-1 Vpr requires specific N-terminal and novel C-terminal domains. BMC Struct. Biol. 2011, 11, 49. [Google Scholar] [CrossRef]
- Solbak, S.M.; Reksten, T.R.; Wray, V.; Bruns, K.; Horvli, O.; Raae, A.J.; Henklein, P.; Roder, R.; Mitzner, D.; Schubert, U.; et al. The intriguing cyclophilin A-HIV-1 Vpr interaction: Prolyl cis/trans isomerisation catalysis and specific binding. BMC Struct. Biol. 2010, 10, 31. [Google Scholar] [CrossRef]
- Damsker, J.M.; Bukrinsky, M.I.; Constant, S.L. Preferential chemotaxis of activated human CD4+ T cells by extracellular cyclophilin A. J. Leukoc. Biol. 2007, 82, 613–618. [Google Scholar] [CrossRef]
- Yurchenko, V.; Constant, S.; Bukrinsky, M. Dealing with the family: CD147 interactions with cyclophilins. Immunology 2006, 117, 301–309. [Google Scholar] [CrossRef]
- Pushkarsky, T.; Zybarth, G.; Dubrovsky, L.; Yurchenko, V.; Tang, H.; Guo, H.; Toole, B.; Sherry, B.; Bukrinsky, M. CD147 facilitates HIV-1 infection by interacting with virus-associated cyclophilin A. P. Natl. Acad. Sci. USA 2001, 98, 6360–6365. [Google Scholar]
- Saphire, A.C.; Bobardt, M.D.; Gallay, P.A. Host cyclophilin A mediates HIV-1 attachment to target cells via heparans. EMBO J. 1999, 18, 6771–6785. [Google Scholar] [CrossRef]
- Pushkarsky, T.; Yurchenko, V.; Laborico, A.; Bukrinsky, M. CD147 stimulates HIV-1 infection in a signal-independent fashion. Biochem. Biophys. Res. Commun. 2007, 363, 495–499. [Google Scholar] [CrossRef]
- Chatterji, U.; Bobardt, M.; Selvarajah, S.; Yang, F.; Tang, H.; Sakamoto, N.; Vuagniaux, G.; Parkinson, T.; Gallay, P. The isomerase active site of cyclophilin A is critical for hepatitis C virus replication. J. Biol. Chem. 2009, 284, 16998–17005. [Google Scholar]
- Fischer, G.; Gallay, P.; Hopkins, S. Cyclophilin inhibitors for the treatment of HCV infection. Curr. Opin. Investig. Drugs 2010, 11, 911–918. [Google Scholar]
- Liu, Z.; Yang, F.; Robotham, J.M.; Tang, H. Critical role of cyclophilin A and its prolyl-peptidyl isomerase activity in the structure and function of the hepatitis C virus replication complex. J. Virol. 2009, 83, 6554–6565. [Google Scholar]
- Tellinghuisen, T.L.; Foss, K.L.; Treadaway, J.C.; Rice, C.M. Identification of residues required for RNA replication in domains II and III of the hepatitis C virus NS5A protein. J. Virol. 2008, 82, 1073–1083. [Google Scholar]
- Patient, R.; Hourioux, C.; Sizaret, P.Y.; Trassard, S.; Sureau, C.; Roingeard, P. Hepatitis B virus subviral envelope particle morphogenesis and intracellular trafficking. J. Virol. 2007, 81, 3842–3851. [Google Scholar] [CrossRef]
- Luo, C.; Luo, H.; Zheng, S.; Gui, C.; Yue, L.; Yu, C.; Sun, T.; He, P.; Chen, J.; Shen, J.; et al. Nucleocapsid protein of SARS coronavirus tightly binds to human cyclophilin A. Biochem. Biophys. Res. Commun. 2004, 321, 557–565. [Google Scholar] [CrossRef]
- Zhirnov, O.P.; Klenk, H.D. Histones as a target for influenza virus matrix protein M1. Virology 1997, 235, 302–310. [Google Scholar] [CrossRef]
- Pleschka, S.; Wolff, T.; Ehrhardt, C.; Hobom, G.; Planz, O.; Rapp, U.R.; Ludwig, S. Influenza virus propagation is impaired by inhibition of the Raf/MEK/ERK signalling cascade. Nat. Cell Biol. 2001, 3, 301–305. [Google Scholar] [CrossRef]
- Reinhardt, J.; Wolff, T. The influenza A virus M1 protein interacts with the cellular receptor of activated C kinase (RACK) 1 and can be phosphorylated by protein kinase C. Veterinary Microbiol. 2000, 74, 87–100. [Google Scholar] [CrossRef]
- Watanabe, K.; Fuse, T.; Asanoa, I.; Tsukahara, F.; Maru, Y.; Nagata, K.; Kitazato, K.; Kobayashi, N. Identification of Hsc70 as an influenza virus matrix protein (M1) binding factor involved in the virus lifecycle. FEBS Letters 2006, 580, 5785–5790. [Google Scholar] [CrossRef]
- Zhirnov, O.P.; Ksenofontov, A.L.; Kuzmina, S.G.; Klenk, H.D. Interaction of influenza A virus M1 matrix protein with caspases. Biochemistry-Moscow 2002, 67, 534–539. [Google Scholar] [CrossRef]
- Liu, N.; Song, W.; Wang, P.; Lee, K.; Chan, W.; Chen, H.; Cai, Z. Proteomics analysis of differential expression of cellular proteins in response to avian H9N2 virus infection in human cells. Proteomics 2008, 8, 1851–1858. [Google Scholar] [CrossRef]
- Jeong, S.J.; Ryo, A.; Yamamoto, N. The prolyl isomerase Pin1 stabilizes the human T-cell leukemia virus type 1 (HTLV-1) Tax oncoprotein and promotes malignant transformation. Biochem. Biophys. Res. Commun. 2009, 381, 294–299. [Google Scholar] [CrossRef]
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Liu, X.; Zhao, Z.; Liu, W. Insights into the Roles of Cyclophilin A During Influenza Virus Infection. Viruses 2013, 5, 182-191. https://doi.org/10.3390/v5010182
Liu X, Zhao Z, Liu W. Insights into the Roles of Cyclophilin A During Influenza Virus Infection. Viruses. 2013; 5(1):182-191. https://doi.org/10.3390/v5010182
Chicago/Turabian StyleLiu, Xiaoling, Zhendong Zhao, and Wenjun Liu. 2013. "Insights into the Roles of Cyclophilin A During Influenza Virus Infection" Viruses 5, no. 1: 182-191. https://doi.org/10.3390/v5010182