The Role of Equine Herpesvirus Type 4 Glycoprotein K in Virus Replication

Abstract
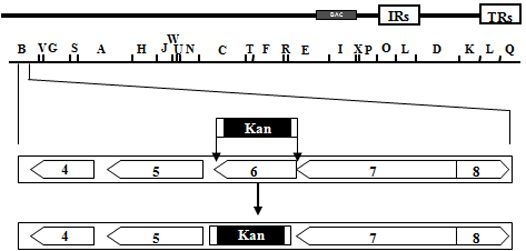
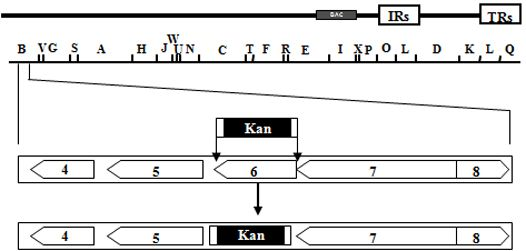
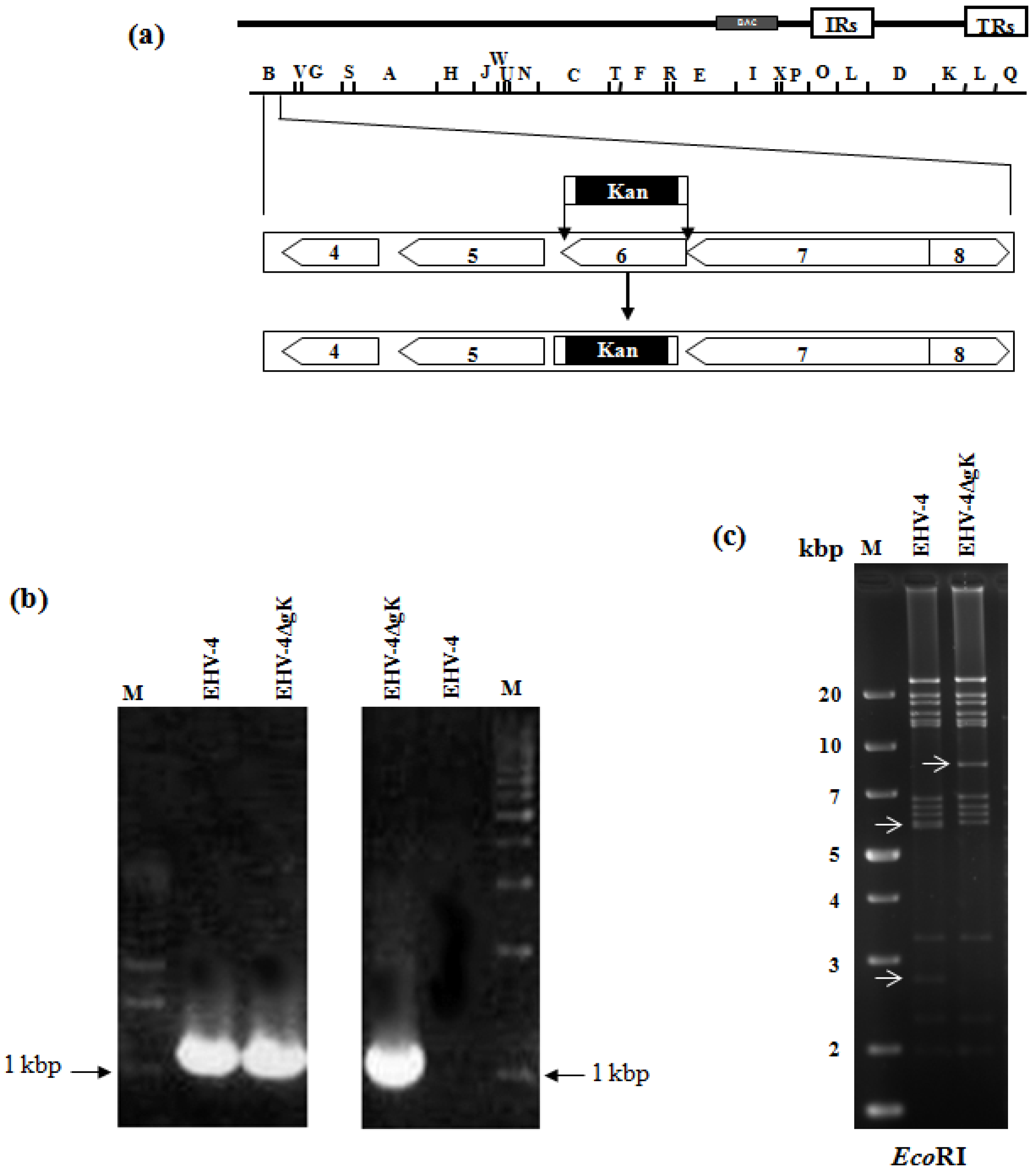
:
{kind=link}
{kind=link}
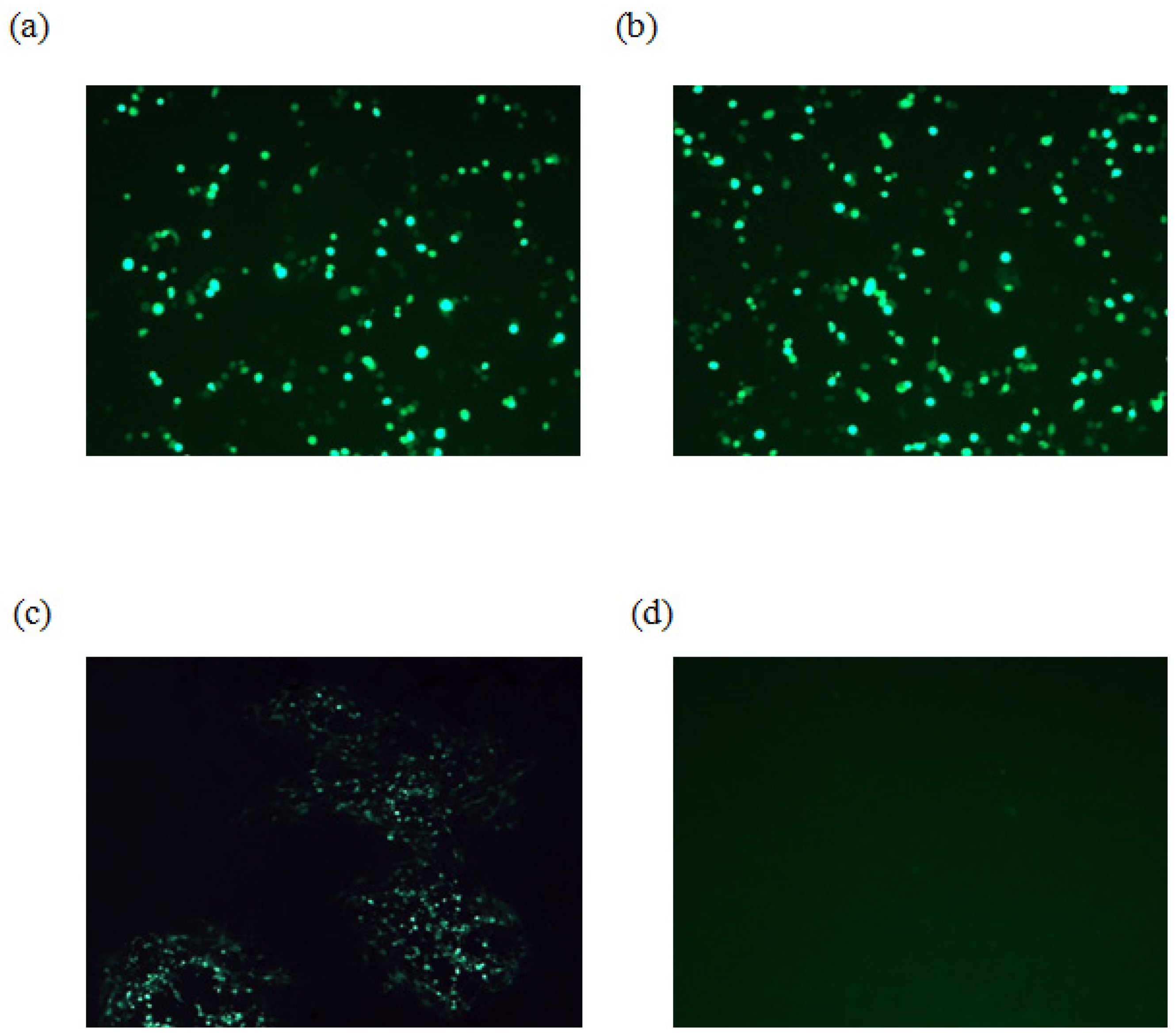{kind=link}
Acknowledgements
Conflict of Interest
References and Notes
- Davison, A.J.; Eberle, R.; Ehlers, B.; Hayward, G.S.; McGeoch, D.J.; Minson, A.C.; Pellett, P.E.; Roizman, B.; Studdert, M.J.; Thiry, E. The order Herpesvirales. Arch. Virol. 2009, 154, 171–177. [Google Scholar] [CrossRef]
- Roizman, B. Herpesviridae. In Virology, 3rd; Field, B.N., Knipe, D.M., Howley, P.M., Channock, R.M., Melnick, J.L., Monath, T.P., Roizman, B., Straus, S.E., Eds.; Lippincott-Raven: Philadelphia, NY, USA, 1996; pp. 2221–2230. [Google Scholar]
- Patel, J.R.; Heldens, J. Equine herpesviruses 1 (EHV-1) and 4 (EHV-4)--epidemiology, disease and immunoprophylaxis: a brief review. Vet. J. 2005, 170, 14–23. [Google Scholar] [CrossRef]
- Foster, T.P.; Kousoulas, K.G. Genetic analysis of the role of herpes simplex virus type 1 glycoprotein K in infectious virus production and egress. J. Virol. 1999, 73, 8457–8468. [Google Scholar]
- Hutchinson, L.; Johnson, D.C. Herpes simplex virus glycoprotein K promotes egress of virus particles. J. Virol. 1995, 69, 5401–5413. [Google Scholar]
- Neubauer, A.; Osterrieder, N. Equine herpesvirus type 1 (EHV-1) glycoprotein K is required for efficient cell-to-cell spread and virus egress. Virology 2004, 329, 18–32. [Google Scholar] [CrossRef]
- Dietz, P.; Klupp, B.G.; Fuchs, W.; Kollner, B.; Weiland, E.; Mettenleiter, T.C. Pseudorabies virus glycoprotein K requires the UL20 gene product for processing. J. Virol. 2000, 74, 5083–5090. [Google Scholar] [CrossRef]
- Spear, P.G. Entry of alphaherpesviruses into cells. seminars in virology 1993, 4, 167–180. [Google Scholar] [CrossRef]
- Zhang, S.; Xiang, J.; Cheng, A.; Wang, M.; Wu, Y.; Yang, X.; Zhu, D.; Jia, R.; Luo, Q.; Chen, Z.; Chen, X. Characterization of duck enteritis virus UL53 gene and glycoprotein K. Virol. J. 2011, 8, 235. [Google Scholar] [CrossRef]
- Azab, W.; El-Sheikh, A.; Abdel-Gawad, A. In vitro characterization of EHV-4 gG-deleted mutant. Virus Genes 2012, 44, 109–111. [Google Scholar] [CrossRef]
- Azab, W.; Kato, K.; Arii, J.; Tsujimura, K.; Yamane, D.; Tohya, Y.; Matsumura, T.; Akashi, H. Cloning of the genome of equine herpesvirus 4 strain TH20p as an infectious bacterial artificial chromosome. Arch. Virol. 2009, 154, 833–842. [Google Scholar] [CrossRef]
- Azab, W.; Osterrieder, N. Glycoproteins D of equine herpesvirus type 1 (EHV-1) and EHV-4 determine cellular tropism independently of integrins. J. Virol. 2012, 86, 2031–2044. [Google Scholar] [CrossRef]
- Azab, W.; Tsujimura, K.; Kato, K.; Arii, J.; Morimoto, T.; Kawaguchi, Y.; Tohya, Y.; Matsumura, T.; Akashi, H. Characterization of a thymidine kinase-deficient mutant of equine herpesvirus 4 and in vitro susceptibility of the virus to antiviral agents. Antiviral Res. 2010a, 85, 389–395. [Google Scholar]
- Azab, W.; Tsujimura, K.; Maeda, K.; Kobayashi, K.; Mohamed, Y.M.; Kato, K.; Matsumura, T.; Akashi, H. Glycoprotein C of equine herpesvirus 4 plays a role in viral binding to cell surface heparan sulfate. Virus Res. 2010b, 151, 1–9. [Google Scholar]
- Tischer, B.K.; von Einem, J.; Kaufer, B.; Osterrieder, N. Two-step red-mediated recombination for versatile high-efficiency markerless DNA manipulation in Escherichia coli. Biotechniques 2006, 40, 191–197. [Google Scholar]
- Telford, E.A.; Watson, M.S.; Perry, J.; Cullinane, A.A.; Davison, A.J. The DNA sequence of equine herpesvirus-4. J. Gen. Virol. 1998, 79 , 1197–1203. [Google Scholar]
- Stroh, T.; Erben, U.; Kuhl, A.A.; Zeitz, M.; Siegmund, B. Combined pulse electroporation--a novel strategy for highly efficient transfection of human and mouse cells. PLoS One 2010, 5, e9488. [Google Scholar]
- Foster, T.P.; Rybachuk, G.V.; Alvarez, X.; Borkhsenious, O.; Kousoulas, K.G. Overexpression of gK in gK-transformed cells collapses the Golgi apparatus into the endoplasmic reticulum inhibiting virion egress, glycoprotein transport, and virus-induced cell fusion. Virology 2003, 317, 237–252. [Google Scholar] [CrossRef]
- Hutchinson, L.; Graham, F.L.; Cai, W.; Debroy, C.; Person, S.; Johnson, D.C. Herpes simplex virus (HSV) glycoproteins B and K inhibit cell fusion induced by HSV syncytial mutants. Virology 1993, 196, 514–531. [Google Scholar] [CrossRef]
- Jayachandra, S.; Baghian, A.; Kousoulas, K.G. Herpes simplex virus type 1 glycoprotein K is not essential for infectious virus production in actively replicating cells but is required for efficient envelopment and translocation of infectious virions from the cytoplasm to the extracellular space. J. Virol. 1997, 71, 5012–5024. [Google Scholar]
- Klupp, B.G.; Baumeister, J.; Dietz, P.; Granzow, H.; Mettenleiter, T.C. Pseudorabies virus glycoprotein gK is a virion structural component involved in virus release but is not required for entry. J. Virol. 1998, 72, 1949–1958. [Google Scholar]
© 2012 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Azab, W.; El-Sheikh, A. The Role of Equine Herpesvirus Type 4 Glycoprotein K in Virus Replication. Viruses 2012, 4, 1258-1263. https://doi.org/10.3390/v4081258
Azab W, El-Sheikh A. The Role of Equine Herpesvirus Type 4 Glycoprotein K in Virus Replication. Viruses. 2012; 4(8):1258-1263. https://doi.org/10.3390/v4081258
Chicago/Turabian StyleAzab, Walid, and Abuelyazeed El-Sheikh. 2012. "The Role of Equine Herpesvirus Type 4 Glycoprotein K in Virus Replication" Viruses 4, no. 8: 1258-1263. https://doi.org/10.3390/v4081258