Envelope Glycoprotein of Arenaviruses

{kind=link}
{kind=link}
Abstract
:1. Introduction
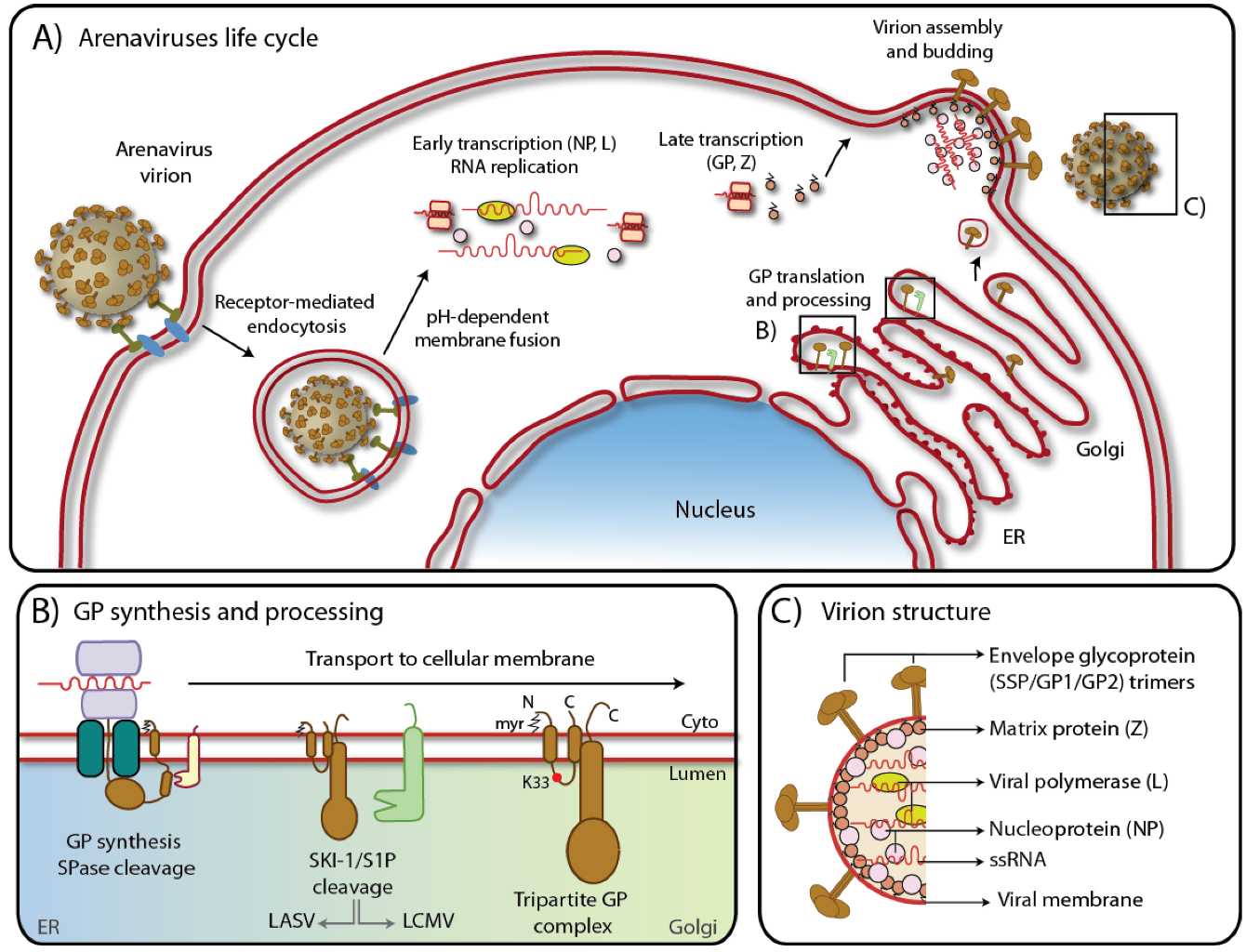
1.2. Arenavirus life-cycle

2. The biosynthesis of the glycoprotein precursor
2.1. The signal peptide is not degraded and forms a stable tripartite complex with GP1/GP2
2.2. Glycosylation of the GP1/GP2 complex
2.3. GPC is cleaved by the cellular protease SKI-1/S1P
3. Role of the envelope glycoprotein on virus assembly and particle formation
Acknowledgments
Conflict of Interest
References and Notes
- Armstrong, C.; Lillie, R. Experimental lymphocytic choriomeningitis of monkeys and mice produced by a virus encountered in studies of the 1933 St. Luis encephalitis epidemic. Publ Health Rep 1934, 49, 1019–1024. [Google Scholar] [CrossRef]
- Rivers, T.M.; McNair Scott, T.F. Meningitis in Man Caused by a Filterable Virus. Science 1935, 81, 439–440. [Google Scholar]
- Traub, E. A Filterable Virus Recovered from White Mice. Science 1935, 81, (2099), 298–299. [Google Scholar]
- Charrel, R.N.; Coutard, B.; Baronti, C.; Canard, B.; Nougairede, A.; Frangeul, A.; Morin, B.; Jamal, S.; Schmidt, C.L.; Hilgenfeld, R.; Klempa, B.; de Lamballerie, X. Arenaviruses and hantaviruses: from epidemiology and genomics to antivirals. Antiviral research 2011, 90, 102–114. [Google Scholar] [CrossRef]
- Geisbert, T.W.; Jahrling, P.B. Exotic emerging viral diseases: progress and challenges. Nature medicine 2004, 10, S110–S121. [Google Scholar] [CrossRef]
- McCormick, J.B.; King, I.J.; Webb, P.A.; Johnson, K.M.; O'Sullivan, R.; Smith, E.S.; Trippel, S.; Tong, T.C. A case-control study of the clinical diagnosis and course of Lassa fever. The Journal of infectious diseases 1987, 155, 445–455. [Google Scholar] [CrossRef]
- Peters, C.J. Human infection with arenaviruses in the Americas. Current topics in microbiology and immunology 2002, 262, 65–74. [Google Scholar] [CrossRef]
- Maiztegui, J.I. Clinical and epidemiological patterns of Argentine haemorrhagic fever. Bulletin of the World Health Organization 1975, 52, 567–575. [Google Scholar]
- Parodi, A.S.; Greenway, D.J.; Rugiero, H.R.; Frigerio, M.; De La Barrera, J.M.; Mettler, N.; Garzon, F.; Boxaca, M.; Guerrero, L.; Nota, N. [Concerning the epidemic outbreak in Junin]. El Dia medico 1958, 30, 2300–2301. [Google Scholar]
- Weissenbacher, M.C.; Laguens, R.P.; Coto, C.E. Argentine hemorrhagic fever. Current topics in microbiology and immunology 1987, 134, 79–116. [Google Scholar]
- Razonable, R.R. Rare, unusual, and less common virus infections after organ transplantation. Current opinion in organ transplantation 2011, 16, 580–587. [Google Scholar] [CrossRef]
- Bonthius, D.J. Lymphocytic choriomeningitis virus: a prenatal and postnatal threat. Advances in pediatrics 2009, 56, 75–86. [Google Scholar] [CrossRef]
- Mahy, B.W.; Dykewicz, C.; Fisher-Hoch, S.; Ostroff, S.; Tipple, M.; Sanchez, A. Virus zoonoses and their potential for contamination of cell cultures. Developments in biological standardization 1991, 75, 183–189. [Google Scholar]
- Delgado, S.; Erickson, B.R.; Agudo, R.; Blair, P.J.; Vallejo, E.; Albarino, C.G.; Vargas, J.; Comer, J.A.; Rollin, P.E.; Ksiazek, T.G.; Olson, J.G.; Nichol, S.T. Chapare virus, a newly discovered arenavirus isolated from a fatal hemorrhagic fever case in Bolivia. PLoS pathogens 2008, 4, e1000047. [Google Scholar] [CrossRef]
- Briese, T.; Paweska, J.T.; McMullan, L.K.; Hutchison, S.K.; Street, C.; Palacios, G.; Khristova, M.L.; Weyer, J.; Swanepoel, R.; Egholm, M.; Nichol, S.T.; Lipkin, W.I. Genetic detection and characterization of Lujo virus, a new hemorrhagic fever-associated arenavirus from southern Africa. PLoS pathogens 2009, 5, e1000455. [Google Scholar] [CrossRef]
- Kohl, K.S.; Arthur, R.R.; O'Connor, R.; Fernandez, J. Assessment of public health events through International Health Regulations, United States, 2007-2011. Emerging infectious diseases 2012, 18, 1047–1053. [Google Scholar] [CrossRef]
- de la Torre, J.C. Molecular and cell biology of the prototypic arenavirus LCMV: implications for understanding and combating hemorrhagic fever arenaviruses. Annals of the New York Academy of Sciences 2009, 1171 Suppl 1, E57–E64. [Google Scholar] [CrossRef]
- Buchmeier, M.J.; de la Torre, J.C.; Peters, C.J. Arenaviridae: the viruses and their replication. In Fields Virology, 4th; Knipe D., L., Howley P., M., Eds.; Lippincott-Raven: Philadelphia, 2007; pp. 1791–1828. [Google Scholar]
- Emonet, S.F.; de la Torre, J.C.; Domingo, E.; Sevilla, N. Arenavirus genetic diversity and its biological implications. In Infection, genetics and evolution : journal of molecular epidemiology and evolutionary genetics in infectious diseases; 2009; Volume 9, pp. 417–429. [Google Scholar]
- Schlie, K.; Maisa, A.; Lennartz, F.; Stroher, U.; Garten, W.; Strecker, T. Characterization of Lassa virus glycoprotein oligomerization and influence of cholesterol on virus replication. Journal of virology 2010, 84, 983–992. [Google Scholar] [CrossRef]
- Fields, B.; Knipe, D.M.; Howley, P.M. Fields Virology, 5th ed; 2007. [Google Scholar]
- Cao, W.; Henry, M.D.; Borrow, P.; Yamada, H.; Elder, J.H.; Ravkov, E.V.; Nichol, S.T.; Compans, R.W.; Campbell, K.P.; Oldstone, M.B. Identification of alpha-dystroglycan as a receptor for lymphocytic choriomeningitis virus and Lassa fever virus. Science 1998, 282, (5396), 2079–2081. [Google Scholar]
- Kunz, S.; Borrow, P.; Oldstone, M.B. Receptor structure, binding, and cell entry of arenavirus. Current topics in microbiology and immunology 2002, 262, 111–137. [Google Scholar] [CrossRef]
- Radoshitzky, S.R.; Abraham, J.; Spiropoulou, C.F.; Kuhn, J.H.; Nguyen, D.; Li, W.; Nagel, J.; Schmidt, P.J.; Nunberg, J.H.; Andrews, N.C.; Farzan, M.; Choe, H. Transferrin receptor 1 is a cellular receptor for New World haemorrhagic fever arenaviruses. Nature 2007, 446, 92–96. [Google Scholar] [CrossRef]
- Radoshitzky, S.R.; Kuhn, J.H.; Spiropoulou, C.F.; Albarino, C.G.; Nguyen, D.P.; Salazar-Bravo, J.; Dorfman, T.; Lee, A.S.; Wang, E.; Ross, S.R.; Choe, H.; Farzan, M. Receptor determinants of zoonotic transmission of New World hemorrhagic fever arenaviruses. Proc Natl Acad Sci U S A 2008, 105, (7), 2664–2669. [Google Scholar]
- Pasqual, G.; Rojek, J.M.; Masin, M.; Chatton, J.Y.; Kunz, S. Old world arenaviruses enter the host cell via the multivesicular body and depend on the endosomal sorting complex required for transport. PLoS pathogens 2011, 7, e1002232. [Google Scholar] [CrossRef]
- Pythoud, C.; Rodrigo, W.W.; Pasqual, G.; Rothenberger, S.; Martinez-Sobrido, L.; de la Torre, J.C.; Kunz, S. Arenavirus nucleoprotein targets interferon regulatory factor-activating kinase IKKepsilon. Journal of virology 2012, 86, 7728–7738. [Google Scholar]
- Aisen, P. Transferrin receptor 1. The international journal of biochemistry & cell biology 2004, 36, 2137–2143. [Google Scholar]
- Huang, C.; Kolokoltsova, O.A.; Yun, N.E.; Seregin, A.V.; Poussard, A.L.; Walker, A.G.; Brasier, A.R.; Zhao, Y.; Tian, B.; de la Torre, J.C.; Paessler, S. Junin virus infection activates the type I interferon pathway in a RIG-I-dependent manner. PLoS neglected tropical diseases 2012, 6, e1659. [Google Scholar] [CrossRef]
- Pasqual, G.; Rojek, J.M.; Masin, M.; Chatton, J.Y.; Kunz, S. Old world arenaviruses enter the host cell via the multivesicular body and depend on the endosomal sorting complex required for transport. PLoS Pathog 2011, 7, (9), e1002232. [Google Scholar]
- Iapalucci, S.; Lopez, N.; Franze-Fernandez, M.T. The 3' end termini of the Tacaribe arenavirus subgenomic RNAs. Virology 1991, 182, 269–278. [Google Scholar] [CrossRef]
- Perez, M.; de la Torre, J.C. Characterization of the genomic promoter of the prototypic arenavirus lymphocytic choriomeningitis virus. Journal of virology 2003, 77, 1184–1194. [Google Scholar] [CrossRef]
- Baird, N.L.; York, J.; Nunberg, J.H. Arenavirus infection induces discrete cytosolic structures for RNA replication. Journal of virology 2012. [Google Scholar]
- Perez, M.; Craven, R.C.; de la Torre, J.C. The small RING finger protein Z drives arenavirus budding: implications for antiviral strategies. Proc Natl Acad Sci U S A 2003, 100, (22), 12978–12983. [Google Scholar]
- Strecker, T.; Eichler, R.; Meulen, J.; Weissenhorn, W.; Dieter Klenk, H.; Garten, W.; Lenz, O. Lassa virus Z protein is a matrix protein and sufficient for the release of virus-like particles [corrected]. Journal of virology 2003, 77, 10700–10705. [Google Scholar]
- Urata, S.; Noda, T.; Kawaoka, Y.; Yokosawa, H.; Yasuda, J. Cellular factors required for Lassa virus budding. Journal of virology 2006, 80, 4191–4195. [Google Scholar] [CrossRef]
- Capul, A.A.; Perez, M.; Burke, E.; Kunz, S.; Buchmeier, M.J.; de la Torre, J.C. Arenavirus Z-glycoprotein association requires Z myristoylation but not functional RING or late domains. J Virol 2007, 81, (17), 9451–9460. [Google Scholar]
- Agnihothram, S.S.; York, J.; Trahey, M.; Nunberg, J.H. Bitopic membrane topology of the stable signal peptide in the tripartite Junin virus GP-C envelope glycoprotein complex. Journal of virology 2007, 81, 4331–4337. [Google Scholar] [CrossRef]
- Beyer, W.R.; Popplau, D.; Garten, W.; von Laer, D.; Lenz, O. Endoproteolytic processing of the lymphocytic choriomeningitis virus glycoprotein by the subtilase SKI-1/S1P. Journal of virology 2003, 77, 2866–2872. [Google Scholar] [CrossRef]
- Lenz, O.; ter Meulen, J.; Klenk, H.D.; Seidah, N.G.; Garten, W. The Lassa virus glycoprotein precursor GP-C is proteolytically processed by subtilase SKI-1/S1P. In Proceedings of the National Academy of Sciences of the United States of America; 2001; 98, pp. 12701–12705. [Google Scholar]
- Rojek, J.M.; Lee, A.M.; Nguyen, N.; Spiropoulou, C.F.; Kunz, S. Site 1 protease is required for proteolytic processing of the glycoproteins of the South American hemorrhagic fever viruses Junin, Machupo, and Guanarito. Journal of virology 2008, 82, 6045–6051. [Google Scholar] [CrossRef]
- York, J.; Romanowski, V.; Lu, M.; Nunberg, J.H. The signal peptide of the Junin arenavirus envelope glycoprotein is myristoylated and forms an essential subunit of the mature G1-G2 complex. Journal of virology 2004, 78, 10783–10792. [Google Scholar] [CrossRef]
- Eichler, R.; Lenz, O.; Garten, W.; Strecker, T. The role of single N-glycans in proteolytic processing and cell surface transport of the Lassa virus glycoprotein GP-C. Virology journal 2006, 3, 41. [Google Scholar] [CrossRef]
- Nunberg, J.H.; York, J. The curious case of arenavirus entry, and its inhibition. Viruses 2012, 4, (1), 83–101. [Google Scholar] [CrossRef]
- Froeschke, M.; Basler, M.; Groettrup, M.; Dobberstein, B. Long-lived signal peptide of lymphocytic choriomeningitis virus glycoprotein pGP-C. J Biol Chem 2003, 278, (43), 41914–41920. [Google Scholar]
- Eichler, R.; Lenz, O.; Strecker, T.; Garten, W. Signal peptide of Lassa virus glycoprotein GP-C exhibits an unusual length. FEBS Lett 2003, 538, (1-3), 203–206. [Google Scholar]
- York, J.; Romanowski, V.; Lu, M.; Nunberg, J.H. The signal peptide of the Junin arenavirus envelope glycoprotein is myristoylated and forms an essential subunit of the mature G1-G2 complex. J Virol. 2004, 78, (19), 10783–10792. [Google Scholar]
- York, J.; Nunberg, J.H. Role of the stable signal peptide of Junin arenavirus envelope glycoprotein in pH-dependent membrane fusion. J Virol 2006, 80, (15), 7775–7780. [Google Scholar]
- York, J.; Nunberg, J.H. Intersubunit interactions modulate pH-induced activation of membrane fusion by the Junin virus envelope glycoprotein GPC. J Virol 2009, 83, (9), 4121–4126. [Google Scholar]
- Eichler, R.; Lenz, O.; Strecker, T.; Eickmann, M.; Klenk, H.D.; Garten, W. Identification of Lassa virus glycoprotein signal peptide as a trans-acting maturation factor. EMBO reports 2003, 4, (11), 1084–1088. [Google Scholar]
- York, J.; Nunberg, J.H. Distinct requirements for signal peptidase processing and function in the stable signal peptide subunit of the Junin virus envelope glycoprotein. Virology 2007, 359, 72–81. [Google Scholar] [CrossRef]
- Albarino, C.G.; Bird, B.H.; Chakrabarti, A.K.; Dodd, K.A.; White, D.M.; Bergeron, E.; Shrivastava-Ranjan, P.; Nichol, S.T. Reverse genetics generation of chimeric infectious Junin/Lassa virus is dependent on interaction of homologous glycoprotein stable signal peptide and G2 cytoplasmic domains. Journal of virology 2011, 85, 112–122. [Google Scholar] [CrossRef]
- Agnihothram, S.S.; York, J.; Nunberg, J.H. Role of the stable signal peptide and cytoplasmic domain of G2 in regulating intracellular transport of the Junin virus envelope glycoprotein complex. Journal of virology 2006, 80, 5189–5198. [Google Scholar] [CrossRef]
- York, J.; Dai, D.; Amberg, S.M.; Nunberg, J.H. pH-induced activation of arenavirus membrane fusion is antagonized by small-molecule inhibitors. Journal of virology 2008, 82, 10932–10939. [Google Scholar] [CrossRef]
- Messina, E.L.; York, J.; Nunberg, J.H. Dissection of the role of the stable signal peptide of the arenavirus envelope glycoprotein in membrane fusion. Journal of virology 2012, 86, 6138–6145. [Google Scholar]
- Braakman, I.; van Anken, E. Folding of viral envelope glycoproteins in the endoplasmic reticulum. Traffic 2000, 1, 533–539. [Google Scholar] [CrossRef]
- Bonhomme, C.J.; Capul, A.A.; Lauron, E.J.; Bederka, L.H.; Knopp, K.A.; Buchmeier, M.J. Glycosylation modulates arenavirus glycoprotein expression and function. Virology 2011, 409, (2), 223–233. [Google Scholar]
- Kido, H.; Okumura, Y.; Takahashi, E.; Pan, H.Y.; Wang, S.; Chida, J.; Le, T.Q.; Yano, M. Host envelope glycoprotein processing proteases are indispensable for entry into human cells by seasonal and highly pathogenic avian influenza viruses. Journal of molecular and genetic medicine : an international journal of biomedical research 2008, 3, 167–175. [Google Scholar]
- Hallenberger, S.; Bosch, V.; Angliker, H.; Shaw, E.; Klenk, H.D.; Garten, W. Inhibition of furin-mediated cleavage activation of HIV-1 glycoprotein gp160. Nature 1992, 360, 358–361. [Google Scholar] [CrossRef]
- Kido, H.; Okumura, Y.; Yamada, H.; Mizuno, D.; Higashi, Y.; Yano, M. Secretory leukoprotease inhibitor and pulmonary surfactant serve as principal defenses against influenza A virus infection in the airway and chemical agents up-regulating their levels may have therapeutic potential. Biological chemistry 2004, 385, 1029–1034. [Google Scholar]
- Blair, W.S.; Cao, J.; Jackson, L.; Jimenez, J.; Peng, Q.; Wu, H.; Isaacson, J.; Butler, S.L.; Chu, A.; Graham, J.; Malfait, A.M.; Tortorella, M.; Patick, A.K. Identification and characterization of UK-201844, a novel inhibitor that interferes with human immunodeficiency virus type 1 gp160 processing. Antimicrobial agents and chemotherapy 2007, 51, 3554–3561. [Google Scholar] [CrossRef]
- Watanabe, M.; Hirano, A.; Stenglein, S.; Nelson, J.; Thomas, G.; Wong, T.C. Engineered serine protease inhibitor prevents furin-catalyzed activation of the fusion glycoprotein and production of infectious measles virus. Journal of virology 1995, 69, 3206–3210. [Google Scholar]
- Ozden, S.; Lucas-Hourani, M.; Ceccaldi, P.E.; Basak, A.; Valentine, M.; Benjannet, S.; Hamelin, J.; Jacob, Y.; Mamchaoui, K.; Mouly, V.; Despres, P.; Gessain, A.; Butler-Browne, G.; Chretien, M.; Tangy, F.; Vidalain, P.O.; Seidah, N.G. Inhibition of Chikungunya virus infection in cultured human muscle cells by furin inhibitors: impairment of the maturation of the E2 surface glycoprotein. The Journal of biological chemistry 2008, 283, 21899–21908. [Google Scholar]
- White, J.M.; Delos, S.E.; Brecher, M.; Schornberg, K. Structures and mechanisms of viral membrane fusion proteins: multiple variations on a common theme. Critical reviews in biochemistry and molecular biology 2008, 43, 189–219. [Google Scholar] [CrossRef]
- Kunz, S.; Edelmann, K.H.; de la Torre, J.C.; Gorney, R.; Oldstone, M.B. Mechanisms for lymphocytic choriomeningitis virus glycoprotein cleavage, transport, and incorporation into virions. Virology 2003, 314, 168–178. [Google Scholar] [CrossRef]
- Damonte, E.B.; Mersich, S.E.; Candurra, N.A. Intracellular processing and transport of Junin virus glycoproteins influences virion infectivity. Virus research 1994, 34, 317–326. [Google Scholar] [CrossRef]
- Pasquato, A.; Burri, D.J.; Traba, E.G.; Hanna-El-Daher, L.; Seidah, N.G.; Kunz, S. Arenavirus envelope glycoproteins mimic autoprocessing sites of the cellular proprotein convertase subtilisin kexin isozyme-1/site-1 protease. Virology 2011, 417, 18–26. [Google Scholar] [CrossRef]
- Eschli, B.; Quirin, K.; Wepf, A.; Weber, J.; Zinkernagel, R.; Hengartner, H. Identification of an N-terminal trimeric coiled-coil core within arenavirus glycoprotein 2 permits assignment to class I viral fusion proteins. J Virol. 2006, 80, (12), 5897–5907. [Google Scholar]
- Igonet, S.; Vaney, M.C.; Vonhrein, C.; Bricogne, G.; Stura, E.A.; Hengartner, H.; Eschli, B.; Rey, F.A. X-ray structure of the arenavirus glycoprotein GP2 in its postfusion hairpin conformation. In Proceedings of the National Academy of Sciences of the United States of America; 2011; 108, pp. 19967–19972. [Google Scholar]
- Pasquato, A.; Pullikotil, P.; Asselin, M.C.; Vacatello, M.; Paolillo, L.; Ghezzo, F.; Basso, F.; Di Bello, C.; Dettin, M.; Seidah, N.G. The proprotein convertase SKI-1/S1P. In vitro analysis of Lassa virus glycoprotein-derived substrates and ex vivo validation of irreversible peptide inhibitors. The Journal of biological chemistry 2006, 281, 23471–23481. [Google Scholar]
- Elagoz, A.; Benjannet, S.; Mammarbassi, A.; Wickham, L.; Seidah, N.G. Biosynthesis and cellular trafficking of the convertase SKI-1/S1P: ectodomain shedding requires SKI-1 activity. The Journal of biological chemistry 2002, 277, 11265–11275. [Google Scholar]
- Espenshade, P.J.; Cheng, D.; Goldstein, J.L.; Brown, M.S. Autocatalytic processing of site-1 protease removes propeptide and permits cleavage of sterol regulatory element-binding proteins. The Journal of biological chemistry 1999, 274, 22795–22804. [Google Scholar]
- Sakai, J.; Rawson, R.B.; Espenshade, P.J.; Cheng, D.; Seegmiller, A.C.; Goldstein, J.L.; Brown, M.S. Molecular identification of the sterol-regulated luminal protease that cleaves SREBPs and controls lipid composition of animal cells. Molecular cells 1998, 2, 505–514. [Google Scholar] [CrossRef]
- Hawkins, J.L.; Robbins, M.D.; Warren, L.C.; Xia, D.; Petras, S.F.; Valentine, J.J.; Varghese, A.H.; Wang, I.K.; Subashi, T.A.; Shelly, L.D.; Hay, B.A.; Landschulz, K.T.; Geoghegan, K.F.; Harwood, H.J., Jr. Pharmacologic inhibition of site 1 protease activity inhibits sterol regulatory element-binding protein processing and reduces lipogenic enzyme gene expression and lipid synthesis in cultured cells and experimental animals. The Journal of pharmacology and experimental therapeutics 2008, 326, 801–808. [Google Scholar] [CrossRef]
- Ye, J.; Rawson, R.B.; Komuro, R.; Chen, X.; Dave, U.P.; Prywes, R.; Brown, M.S.; Goldstein, J.L. ER stress induces cleavage of membrane-bound ATF6 by the same proteases that process SREBPs. Molecular cell 2000, 6, 1355–1364. [Google Scholar] [CrossRef]
- Schroder, M. Endoplasmic reticulum stress responses. Cellular and molecular life sciences : CMLS 2008, 65, 862–894. [Google Scholar] [CrossRef]
- Marschner, K.; Kollmann, K.; Schweizer, M.; Braulke, T.; Pohl, S. A key enzyme in the biogenesis of lysosomes is a protease that regulates cholesterol metabolism. Science 2011, 333, 87–90. [Google Scholar]
- Seidah, N.G.; Mowla, S.J.; Hamelin, J.; Mamarbachi, A.M.; Benjannet, S.; Toure, B.B.; Basak, A.; Munzer, J.S.; Marcinkiewicz, J.; Zhong, M.; Barale, J.C.; Lazure, C.; Murphy, R.A.; Chretien, M.; Marcinkiewicz, M. Mammalian subtilisin/kexin isozyme SKI-1: A widely expressed proprotein convertase with a unique cleavage specificity and cellular localization. In Proceedings of the National Academy of Sciences of the United States of America; 1999; 96, pp. 1321–1326. [Google Scholar]
- Tassew, N.G.; Charish, J.; Seidah, N.G.; Monnier, P.P. SKI-1 and Furin generate multiple RGMa fragments that regulate axonal growth. Developmental cell 2012, 22, 391–402. [Google Scholar] [CrossRef]
- Gorski, J.P.; Huffman, N.T.; Cui, C.; Henderson, E.P.; Midura, R.J.; Seidah, N.G. Potential role of proprotein convertase SKI-1 in the mineralization of primary bone. Cells, tissues, organs 2009, 189, 25–32. [Google Scholar] [CrossRef]
- Schlombs, K.; Wagner, T.; Scheel, J. Site-1 protease is required for cartilage development in zebrafish. In Proceedings of the National Academy of Sciences of the United States of America; 2003; 100, pp. 14024–14029. [Google Scholar]
- Rutschmann, S.; Crozat, K.; Li, X.; Du, X.; Hanselman, J.C.; Shigeoka, A.A.; Brandl, K.; Popkin, D.L.; McKay, D.B.; Xia, Y.; Moresco, E.M.; Beutler, B. Hypopigmentation and maternal-zygotic embryonic lethality caused by a hypomorphic mbtps1 mutation in mice. G3 2012, 2, 499–504. [Google Scholar] [CrossRef]
- Mitchell, K.J.; Pinson, K.I.; Kelly, O.G.; Brennan, J.; Zupicich, J.; Scherz, P.; Leighton, P.A.; Goodrich, L.V.; Lu, X.; Avery, B.J.; Tate, P.; Dill, K.; Pangilinan, E.; Wakenight, P.; Tessier-Lavigne, M.; Skarnes, W.C. Functional analysis of secreted and transmembrane proteins critical to mouse development. Nature genetics 2001, 28, 241–249. [Google Scholar] [CrossRef]
- Brandl, K.; Rutschmann, S.; Li, X.; Du, X.; Xiao, N.; Schnabl, B.; Brenner, D.A.; Beutler, B. Enhanced sensitivity to DSS colitis caused by a hypomorphic Mbtps1 mutation disrupting the ATF6-driven unfolded protein response. In Proceedings of the National Academy of Sciences of the United States of America; 2009; 106, pp. 3300–3305. [Google Scholar]
- Popkin, D.L.; Teijaro, J.R.; Sullivan, B.M.; Urata, S.; Rutschmann, S.; de la Torre, J.C.; Kunz, S.; Beutler, B.; Oldstone, M. Hypomorphic mutation in the site-1 protease Mbtps1 endows resistance to persistent viral infection in a cell-specific manner. Cell host & microbe 2011, 9, (3), 212–222. [Google Scholar]
- Burri, D.J.; Pasqual, G.; Rochat, C.; Seidah, N.G.; Pasquato, A.; Kunz, S. Molecular characterization of the processing of arenavirus envelope glycoprotein precursors by subtilisin kexin isozyme-1/site-1 protease. Journal of virology 2012, 86, 4935–4946. [Google Scholar] [CrossRef]
- Candurra, N.A.; Damonte, E.B. Effect of inhibitors of the intracellular exocytic pathway on glycoprotein processing and maturation of Junin virus. Archives of virology 1997, 142, 2179–2193. [Google Scholar] [CrossRef]
- Oldstone, M.B.; Buchmeier, M.J. Restricted expression of viral glycoprotein in cells of persistently infected mice. Nature. 1982, 300, (5890), 360–362. [Google Scholar]
- Pasqual, G.; Burri, D.J.; Pasquato, A.; de la Torre, J.C.; Kunz, S. Role of the host cell's unfolded protein response in arenavirus infection. Journal of virology 2011, 85, 1662–1670. [Google Scholar] [CrossRef]
- Basak, A.; Chretien, M.; Seidah, N.G. A rapid fluorometric assay for the proteolytic activity of SKI-1/S1P based on the surface glycoprotein of the hemorrhagic fever Lassa virus. FEBS letters 2002, 514, 333–339. [Google Scholar] [CrossRef]
- Klewitz, C.; Klenk, H.D.; ter Meulen, J. Amino acids from both N-terminal hydrophobic regions of the Lassa virus envelope glycoprotein GP-2 are critical for pH-dependent membrane fusion and infectivity. The Journal of general virology 2007, 88, 2320–2328. [Google Scholar] [CrossRef]
- Sergel, T.; Morrison, T.G. Mutations in the cytoplasmic domain of the fusion glycoprotein of Newcastle disease virus depress syncytia formation. Virology 1995, 210, 264–272. [Google Scholar] [CrossRef]
- Cathomen, T.; Naim, H.Y.; Cattaneo, R. Measles viruses with altered envelope protein cytoplasmic tails gain cell fusion competence. Journal of virology 1998, 72, 1224–1234. [Google Scholar]
- Gabuzda, D.H.; Lever, A.; Terwilliger, E.; Sodroski, J. Effects of deletions in the cytoplasmic domain on biological functions of human immunodeficiency virus type 1 envelope glycoproteins. Journal of virology 1992, 66, 3306–3315. [Google Scholar]
- Schlie, K.; Strecker, T.; Garten, W. Maturation cleavage within the ectodomain of Lassa virus glycoprotein relies on stabilization by the cytoplasmic tail. FEBS letters 2010, 584, 4379–4382. [Google Scholar] [CrossRef]
- Mohottalage, D.; Goto, N.; Basak, A. Subtilisin kexin isozyme-1 (SKI-1): production, purification, inhibitor design and biochemical applications. Advances in experimental medicine and biology 2009, 611, 83–84. [Google Scholar]
- Rojek, J.M.; Pasqual, G.; Sanchez, A.B.; Nguyen, N.T.; de la Torre, J.C.; Kunz, S. Targeting the proteolytic processing of the viral glycoprotein precursor is a promising novel antiviral strategy against arenaviruses. Journal of virology 2010, 84, 573–584. [Google Scholar] [CrossRef]
- Maisa, A.; Stroher, U.; Klenk, H.D.; Garten, W.; Strecker, T. Inhibition of Lassa virus glycoprotein cleavage and multicycle replication by site 1 protease-adapted alpha(1)-antitrypsin variants. PLoS neglected tropical diseases 2009, 3, e446. [Google Scholar] [CrossRef]
- Pasquato, A.; Rochat, C.; Burri, D.J.; Pasqual, G.; de la Torre, J.C.; Kunz, S. Evaluation of the anti-arenaviral activity of the subtilisin kexin isozyme-1/site-1 protease inhibitor PF-429242. Virology 2012, 423, 14–22. [Google Scholar] [CrossRef]
- Hay, B.A.; Abrams, B.; Zumbrunn, A.Y.; Valentine, J.J.; Warren, L.C.; Petras, S.F.; Shelly, L.D.; Xia, A.; Varghese, A.H.; Hawkins, J.L.; Van Camp, J.A.; Robbins, M.D.; Landschulz, K.; Harwood, H.J., Jr. Aminopyrrolidineamide inhibitors of site-1 protease. Bioorganic & medicinal chemistry letters 2007, 17, 4411–4414. [Google Scholar]
- Urata, S.; Yun, N.; Pasquato, A.; Paessler, S.; Kunz, S.; de la Torre, J.C. Antiviral activity of a small-molecule inhibitor of arenavirus glycoprotein processing by the cellular site 1 protease. Journal of virology 2011, 85, 795–803. [Google Scholar] [CrossRef]
- Oldstone, M.B.; Buchmeier, M.J. Restricted expression of viral glycoprotein in cells of persistently infected mice. Nature 1982, 300, 360–362. [Google Scholar] [CrossRef]
- Pullikotil, P.; Benjannet, S.; Mayne, J.; Seidah, N.G. The proprotein convertase SKI-1/S1P: alternate translation and subcellular localization. The Journal of biological chemistry 2007, 282, 27402–27413. [Google Scholar]
- Olmstead, A.D.; Knecht, W.; Lazarov, I.; Dixit, S.B.; Jean, F. Human subtilase SKI-1/S1P is a master regulator of the HCV Lifecycle and a potential host cell target for developing indirect-acting antiviral agents. PLoS Pathog 2012, 8, (1), e1002468. [Google Scholar]
- Hsu, N.Y.; Ilnytska, O.; Belov, G.; Santiana, M.; Chen, Y.H.; Takvorian, P.M.; Pau, C.; van der Schaar, H.; Kaushik-Basu, N.; Balla, T.; Cameron, C.E.; Ehrenfeld, E.; van Kuppeveld, F.J.; Altan-Bonnet, N. Viral reorganization of the secretory pathway generates distinct organelles for RNA replication. Cell 2010, 141, 799–811. [Google Scholar] [CrossRef]
- Perera, R.; Riley, C.; Isaac, G.; Hopf-Jannasch, A.S.; Moore, R.J.; Weitz, K.W.; Pasa-Tolic, L.; Metz, T.O.; Adamec, J.; Kuhn, R.J. Dengue virus infection perturbs lipid homeostasis in infected mosquito cells. PLoS pathogens 2012, 8, e1002584. [Google Scholar]
- Perez, M.; Greenwald, D.L.; de la Torre, J.C. Myristoylation of the RING finger Z protein is essential for arenavirus budding. J Virol 2004, 78, (20), 11443–11448. [Google Scholar]
- Schlie, K.; Maisa, A.; Freiberg, F.; Groseth, A.; Strecker, T.; Garten, W. Viral protein determinants of Lassa virus entry and release from polarized epithelial cells. Journal of virology 2010, 84, 3178–3188. [Google Scholar] [CrossRef]
- Casabona, J.C.; Levingston Macleod, J.M.; Loureiro, M.E.; Gomez, G.A.; Lopez, N. The RING domain and the L79 residue of Z protein are involved in both the rescue of nucleocapsids and the incorporation of glycoproteins into infectious chimeric arenavirus-like particles. Journal of virology 2009, 83, 7029–7039. [Google Scholar] [CrossRef]
© 2012 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Burri, D.J.; Palma, J.R.D.; Kunz, S.; Pasquato, A. Envelope Glycoprotein of Arenaviruses. Viruses 2012, 4, 2162-2181. https://doi.org/10.3390/v4102162
Burri DJ, Palma JRD, Kunz S, Pasquato A. Envelope Glycoprotein of Arenaviruses. Viruses. 2012; 4(10):2162-2181. https://doi.org/10.3390/v4102162
Chicago/Turabian StyleBurri, Dominique J., Joel Ramos Da Palma, Stefan Kunz, and Antonella Pasquato. 2012. "Envelope Glycoprotein of Arenaviruses" Viruses 4, no. 10: 2162-2181. https://doi.org/10.3390/v4102162