
Orthoflavivirus Lammi in Russia: Possible Transovarial Transmission and Trans-Stadial Survival in Aedes cinereus (Diptera, Culicidae)

 ,
, 
 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sampling Technique for Mosquitoes
2.2. Preparation of Mosquito Suspensions
2.3. Detection of Orthoflaviviruses by RT-PCR and Sequencing
2.4. Isolation of Lammi Virus on Cell Culture
2.5. Phylogenetic Analysis
3. Results
3.1. Collection of Mosquitoes and Lammi Virus Detection
3.2. Isolation of Lammi Virus in Cell Line
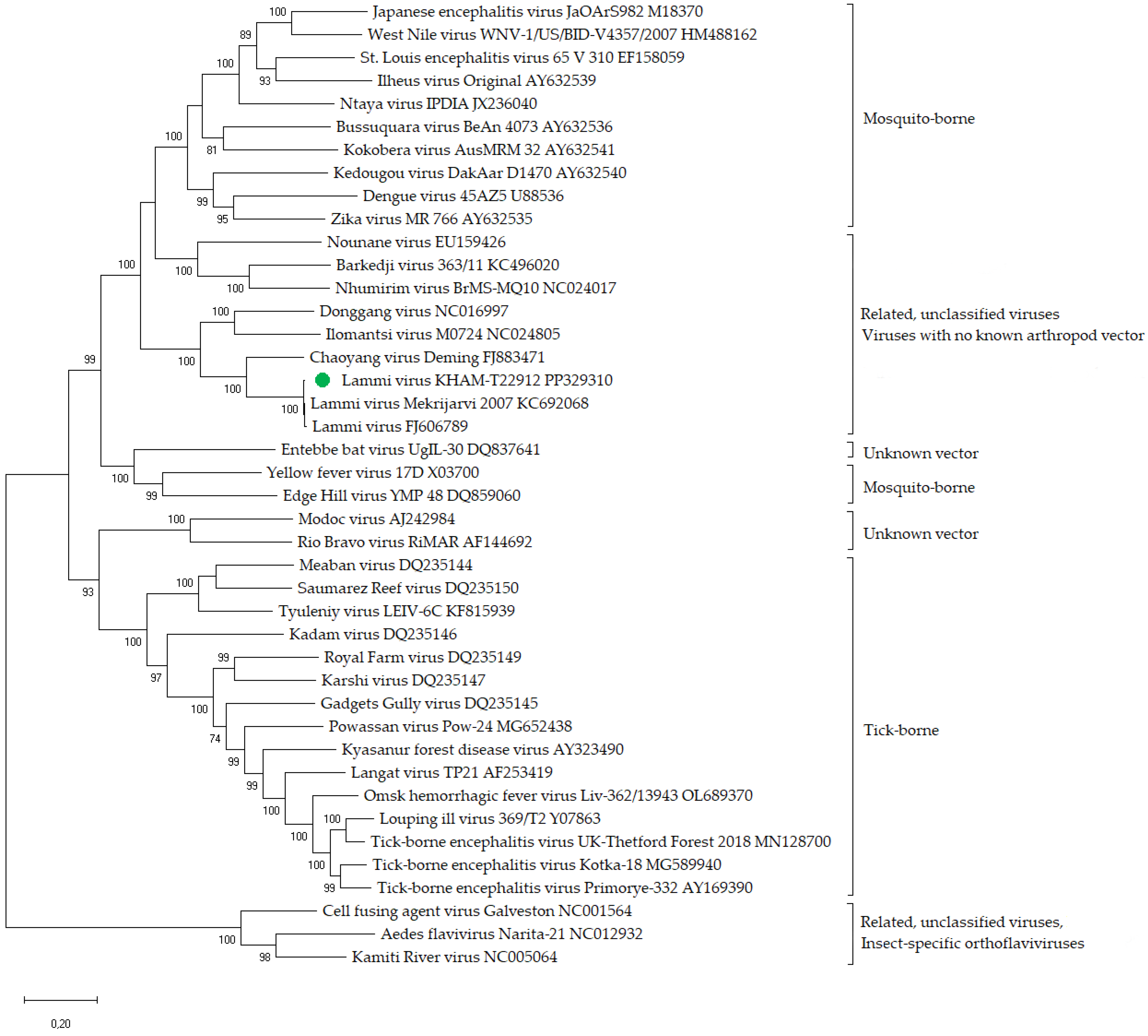
3.3. Phylogenetic Analysis and Genomic Identity Assessment
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Shi, M.; Lin, X.-D.; Vasilakis, N.; Tian, J.-H.; Li, C.-X.; Chen, L.-J.; Eastwood, G.; Diao, X.-N.; Chen, M.-H.; Chen, X.; et al. Divergent Viruses Discovered in Arthropods and Vertebrates Revise the Evolutionary History of the Flaviviridae and Related Viruses. J. Virol. 2016, 90, 659–669. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; He, B.; Huang, S.Y.; Wei, F.; Zhu, X.Q. Severe Fever with Thrombocytopenia Syndrome, an Emerging Tick-Borne Zoonosis. Lancet Infect. Dis. 2014, 14, 763–772. [Google Scholar] [CrossRef] [PubMed]
- López, Y.; Miranda, J.; Mattar, S.; Gonzalez, M.; Rovnak, J. First Report of Lihan Tick Virus (Phlebovirus, Phenuiviridae) in Ticks, Colombia. Virol. J. 2020, 17, 63. [Google Scholar] [CrossRef] [PubMed]
- Litov, A.G.; Belova, O.A.; Kholodilov, I.S.; Gadzhikurbanov, M.N.; Gmyl, L.V.; Oorzhak, N.D.; Saryglar, A.A.; Ishmukhametov, A.A.; Karganova, G.G. Possible Arbovirus Found in Virome of Melophagus ovinus. Viruses 2021, 13, 2375. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.-D.; Wang, W.; Wang, N.N.; Qiu, K.; Zhang, X.; Tana, G.; Liu, Q. Prevalence of the Emerging Novel Alongshan Virus Infection in Sheep and Cattle in Inner Mongolia, Northeastern China. Parasit. Vectors 2019, 12, 450. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.-D.; Wang, B.; Wei, F.; Han, S.-Z.; Zhang, L.; Yang, Z.-T.; Yan, Y.; Lv, X.-L.; Li, L.; Wang, S.-C.; et al. A New Segmented Virus Associated with Human Febrile Illness in China. N. Engl. J. Med. 2019, 380, 2116–2125. [Google Scholar] [CrossRef] [PubMed]
- Xiao, P.; Han, J.; Zhang, Y.; Li, C.; Guo, X.; Wen, S.; Tian, M.; Li, Y.; Wang, M.; Liu, H.; et al. Metagenomic Analysis of Flaviviridae in Mosquito Viromes Isolated from Yunnan Province in China Reveals Genes from Dengue and Zika Viruses. Front. Cell. Infect. Microbiol. 2018, 8, 359. [Google Scholar] [CrossRef] [PubMed]
- Qin, X.-C.; Shi, M.; Tian, J.-H.; Lin, X.-D.; Gao, D.-Y.; He, J.-R.; Wang, J.-B.; Li, C.-X.; Kang, Y.-J.; Yu, B.; et al. A Tick-Borne Segmented RNA Virus Contains Genome Segments Derived from Unsegmented Viral Ancestors. Proc. Natl. Acad. Sci. USA 2014, 111, 6744–6749. [Google Scholar] [CrossRef] [PubMed]
- Kholodilov, I.S.; Belova, O.A.; Morozkin, E.S.; Litov, A.G.; Ivannikova, A.Y.; Makenov, M.T.; Shchetinin, A.M.; Aibulatov, S.V.; Bazarova, G.K.; Bell-Sakyi, L.; et al. Geographical and Tick-Dependent Distribution of Flavi-like Alongshan and Yanggou Tick Viruses in Russia. Viruses 2021, 13, 458. [Google Scholar] [CrossRef]
- Kholodilov, I.S.; Belova, O.A.; Ivannikova, A.Y.; Gadzhikurbanov, M.N.; Makenov, M.T.; Yakovlev, A.S.; Polienko, A.E.; Dereventsova, A.V.; Litov, A.G.; Gmyl, L.V.; et al. Distribution and Characterisation of Tick-Borne Flavi-, Flavi-like, and Phenuiviruses in the Chelyabinsk Region of Russia. Viruses 2022, 14, 2699. [Google Scholar] [CrossRef]
- Postler, T.S.; Beer, M.; Blitvich, B.J.; Bukh, J.; de Lamballerie, X.; Drexler, J.F.; Imrie, A.; Kapoor, A.; Karganova, G.G.; Lemey, P.; et al. Renaming of the Genus Flavivirus to Orthoflavivirus and Extension of Binomial Species Names within the Family Flaviviridae. Arch. Virol. 2023, 168, 224. [Google Scholar] [CrossRef] [PubMed]
- Růžek, D.; Yakimenko, V.V.; Karan, L.S.; Tkachev, S.E. Omsk Haemorrhagic Fever. Lancet 2010, 376, 2104–2113. [Google Scholar] [CrossRef] [PubMed]
- Ruzek, D.; Avšič Županc, T.; Borde, J.; Chrdle, A.; Eyer, L.; Karganova, G.; Kholodilov, I.; Knap, N.; Kozlovskaya, L.; Matveev, A.; et al. Tick-Borne Encephalitis in Europe and Russia: Review of Pathogenesis, Clinical Features, Therapy, and Vaccines. Antivir. Res. 2019, 164, 23–51. [Google Scholar] [CrossRef] [PubMed]
- Chancey, C.; Grinev, A.; Volkova, E.; Rios, M. The Global Ecology and Epidemiology of West Nile Virus. BioMed Res. Int. 2015, 2015, 376230. [Google Scholar] [CrossRef] [PubMed]
- Roberts, A.; Gandhi, S. Japanese Encephalitis Virus: A Review on Emerging Diagnostic Techniques. Front. Biosci. 2020, 25, 1875–1893. [Google Scholar] [CrossRef]
- Guzman, M.G.; Harris, E. Dengue. Lancet 2015, 385, 453–465. [Google Scholar] [CrossRef] [PubMed]
- International Committee on Taxonomy of Viruses Genus: Orthoflavivirus. Available online: https://talk.ictvonline.org/ictv-reports/ictv_online_report/positive-sense-rna-viruses/w/flaviviridae/360/genus-flavivirus (accessed on 27 March 2024).
- Grard, G.; Moureau, G.; Charrel, R.N.; Lemasson, J.J.; Gonzalez, J.P.; Gallian, P.; Gritsun, T.S.; Holmes, E.C.; Gould, E.A.; de Lamballerie, X. Genetic Characterization of Tick-Borne Flaviviruses: New Insights into Evolution, Pathogenetic Determinants and Taxonomy. Virology 2007, 361, 80–92. [Google Scholar] [CrossRef] [PubMed]
- Cook, S.; Holmes, E.C. A Multigene Analysis of the Phylogenetic Relationships among the Flaviviruses (Family: Flaviviridae) and the Evolution of Vector Transmission. Arch. Virol. 2006, 151, 309–325. [Google Scholar] [CrossRef]
- Blitvich, B.J.; Firth, A.E. Insect-Specific Flaviviruses: A Systematic Review of Their Discovery, Host Range, Mode of Transmission, Superinfection Exclusion Potential and Genomic Organization. Viruses 2015, 7, 1927–1959. [Google Scholar] [CrossRef] [PubMed]
- Gordon Smith, C.E. A Virus Resembling Russian Spring-Summer Encephalitis Virus from an Ixodid Tick in Malaya. Nature 1956, 178, 581–582. [Google Scholar] [CrossRef]
- Bancroft, W.H.; Scott, R.M.; Snitbhan, R.; Weaver, R.E., Jr.; Gould, D.J. Isolation of Langat Virus from Haemaphysalis papuana Thorell in Thailand. Am. J. Trop. Med. Hyg. 1976, 25, 500–504. [Google Scholar] [CrossRef] [PubMed]
- Deardorff, E.R.; Nofchissey, R.A.; Cook, J.A.; Hope, A.G.; Tsvetkova, A.; Talbot, S.L.; Ebel, G.D. Powassan Virus in Mammals, Alaska and New Mexico, USA, and Russia, 2004–2007. Emerg. Infect. Dis. 2013, 19, 2012–2016. [Google Scholar] [CrossRef] [PubMed]
- Greig, J.R.; Brownlee, A.; Wilson, D.R.; Gordon, W.S. The Nature of Louping Ill. Vet. Rec. 1931, 11, 325–333. [Google Scholar]
- Weinberger, M.; Pitlik, S.D.; Gandacu, D.; Lang, R.; Nassar, F.; Ben David, D.; Rubinstein, E.; Izthaki, A.; Mishal, J.; Kitzes, R.; et al. West Nile Fever Outbreak, Israel, 2000: Epidemiologic Aspects. Emerg. Infect. Dis. 2001, 7, 686–691. [Google Scholar] [CrossRef]
- Wikan, N.; Smith, D.R. Zika Virus: History of a Newly Emerging Arbovirus. Lancet Infect. Dis. 2016, 16, e119–e126. [Google Scholar] [CrossRef] [PubMed]
- Gubler, D.J. Potential Yellow Fever Epidemics in Unexposed Populations. Bull. World Health Organ. 2018, 96, 299. [Google Scholar] [CrossRef] [PubMed]
- Leung, X.Y.; Islam, R.M.; Adhami, M.; Ilic, D.; McDonald, L.; Palawaththa, S.; Diug, B.; Munshi, S.U.; Karim, M.N. A Systematic Review of Dengue Outbreak Prediction Models: Current Scenario and Future Directions. PLoS Negl. Trop. Dis. 2023, 17, e0010631. [Google Scholar] [CrossRef] [PubMed]
- Hoad, V.C.; Kiely, P.; Seed, C.R.; Viennet, E.; Gosbell, I.B. An Outbreak of Japanese Encephalitis Virus in Australia; What Is the Risk to Blood Safety? Viruses 2022, 14, 1935. [Google Scholar] [CrossRef]
- Parida, M.; Dash, P.K.; Tripathi, N.K.; Ambuj; Sannarangaiah, S.; Saxena, P.; Agarwal, S.; Sahni, A.K.; Singh, S.P.; Rathi, A.K.; et al. Japanese Encephalitis Outbreak, India, 2005. Emerg. Infect. Dis. 2006, 12, 1427–1430. [Google Scholar] [CrossRef]
- Curren, E.J.; Lindsey, N.P.; Fischer, M.; Hills, S.L. St. Louis Encephalitis Virus Disease in the United States, 2003–2017. Am. J. Trop. Med. Hyg. 2018, 99, 1074–1079. [Google Scholar] [CrossRef]
- Selvey, L.A.; Dailey, L.; Lindsay, M.; Armstrong, P.; Tobin, S.; Koehler, A.P.; Markey, P.G.; Smith, D.W. The Changing Epidemiology of Murray Valley Encephalitis in Australia: The 2011 Outbreak and a Review of the Literature. PLoS Negl. Trop. Dis. 2014, 8, 18. [Google Scholar] [CrossRef] [PubMed]
- Grard, G.; Moureau, G.; Charrel, R.N.; Holmes, E.C.; Gould, E.A.; de Lamballerie, X. Genomics and Evolution of Aedes-Borne Flaviviruses. J. Gen. Virol. 2010, 91, 87–94. [Google Scholar] [CrossRef] [PubMed]
- Hobson-Peters, J.; Wei, A.; Yam, Y.; Wei, J.; Lu, F.; Setoh, Y.X.; May, F.J.; Kurucz, N.; Walsh, S.; Prow, N.A.; et al. A New Insect-Specific Flavivirus from Northern Australia Suppresses Replication of West Nile Virus and Murray Valley Encephalitis Virus in Co-Infected Mosquito Cells. PLoS ONE 2013, 8, e56534. [Google Scholar] [CrossRef] [PubMed]
- Fauver, J.R.; Grubaugh, N.D.; Krajacich, B.J.; Weger-Lucarelli, J.; Lakin, S.M.; Fakoli, L.S.; Bolay, F.K.; Diclaro, J.W.; Dabiré, K.R.; Foy, B.D.; et al. West African Anopheles gambiae Mosquitoes Harbor a Taxonomically Diverse Virome Including New Insect-Specific Flaviviruses, Mononegaviruses, and Totiviruses. Virology 2016, 498, 288–299. [Google Scholar] [CrossRef]
- Grard, G.; Lemasson, J.J.; Sylla, M.; Dubot, A.; Cook, S.; Molez, J.F.; Pourrut, X.; Charrel, R.; Gonzalez, J.P.; Munderloh, U.; et al. Ngoye Virus: A Novel Evolutionary Lineage within the Genus Flavivirus. J. Gen. Virol. 2006, 87, 3273–3277. [Google Scholar] [CrossRef] [PubMed]
- Cholleti, H.; Hayer, J.; Abilio, A.P.; Mulandane, F.C.; Verner-Carlsson, J.; Falk, K.I.; Fafetine, J.M.; Berg, M.; Blomström, A.L. Discovery of Novel Viruses in Mosquitoes from the Zambezi Valley of Mozambique. PLoS ONE 2016, 11, e0162751. [Google Scholar] [CrossRef] [PubMed]
- Cook, S.; Moureau, G.; Harbach, R.E.; Mukwaya, L.; Goodger, K.; Ssenfuka, F.; Gould, E.; Holmes, E.C.; de Lamballerie, X. Isolation of a Novel Species of Flavivirus and a New Strain of Culex Flavivirus (Flaviviridae) from a Natural Mosquito Population in Uganda. J. Gen. Virol. 2009, 90, 2669–2678. [Google Scholar] [CrossRef] [PubMed]
- Crabtree, M.B.; Sang, R.C.; Stollar, V.; Dunster, L.M.; Miller, B.R. Genetic and Phenotypic Characterization of the Newly Described Insect Flavivirus, Kamiti River Virus. Arch. Virol. 2003, 148, 1095–1118. [Google Scholar] [CrossRef] [PubMed]
- Junglen, S.; Kopp, A.; Kurth, A.; Pauli, G.; Ellerbrok, H.; Leendertz, F.H. A New Flavivirus and a New Vector: Characterization of a Novel Flavivirus Isolated from Uranotaenia Mosquitoes from a Tropical Rain Forest. J. Virol. 2009, 83, 4462–4468. [Google Scholar] [CrossRef]
- Alkan, C.; Zapata, S.; Bichaud, L.; Moureau, G.; Lemey, P.; Firth, A.E.; Gritsun, T.S.; Gould, E.A.; de Lamballerie, X.; Depaquit, J.; et al. Ecuador Paraiso Escondido Virus, a New Flavivirus Isolated from New World Sand Flies in Ecuador, Is the First Representative of a Novel Clade in the Genus Flavivirus. J. Virol. 2015, 89, 11773–11785. [Google Scholar] [CrossRef]
- Guzman, H.; Contreras-Gutierrez, M.A.; Travassos da Rosa, A.P.A.; Nunes, M.R.T.; Cardoso, J.F.; Popov, V.L.; Young, K.I.; Savit, C.; Wood, T.G.; Widen, S.G.; et al. Characterization of Three New Insect-Specific Flaviviruses: Their Relationship to the Mosquito-Borne Flavivirus Pathogens. Am. J. Trop. Med. Hyg. 2018, 98, 410–419. [Google Scholar] [CrossRef]
- Pauvolid-Corrêa, A.; Solberg, O.D.; Couto-Lima, D.; Kenney, J.; Nogueira, R.; Komar, N. Nhumirim Virus, a Novel Flavivirus Isolated from Mosquitoes from the Pantanal, Brazil Alex. Physiol. Behav. 2011, 176, 139–148. [Google Scholar] [CrossRef]
- Hoshino, K.; Isawa, H.; Tsuda, Y.; Sawabe, K.; Kobayashi, M. Isolation and Characterization of a New Insect Flavivirus from Aedes albopictus and Aedes flavopictus Mosquitoes in Japan. Virology 2009, 391, 119–129. [Google Scholar] [CrossRef] [PubMed]
- Misencik, M.J.; Grubaugh, N.D.; Andreadis, T.G.; Ebel, G.D.; Armstrong, P.M. Isolation of a Novel Insect-Specific Flavivirus from Culiseta melanura in the Northeastern United States. Vector-Borne Zoonotic Dis. 2016, 16, 181–190. [Google Scholar] [CrossRef] [PubMed]
- Kuwata, R.; Sugiyama, H.; Yonemitsu, K.; Van Dung, N.; Terada, Y.; Taniguchi, M.; Shimoda, H.; Takano, A.; Maeda, K. Isolation of Japanese Encephalitis Virus and a Novel Insect-Specific Flavivirus from Mosquitoes Collected in a Cowshed in Japan. Arch. Virol. 2015, 160, 2151–2159. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.S.; Grubaugh, N.D.; Kondig, J.P.; Turell, M.J.; Kim, H.C.; Klein, T.A.; O’Guinn, M.L. Isolation and Genomic Characterization of Chaoyang Virus Strain ROK144 from Aedes vexans Nipponii from the Republic of Korea. Virology 2013, 435, 220–224. [Google Scholar] [CrossRef] [PubMed]
- Zuo, S.; Zhao, Q.; Guo, X.; Zhou, H.; Cao, W.; Zhang, J. Detection of Quang Binh Virus from Mosquitoes in China. Virus Res. 2014, 180, 31–38. [Google Scholar] [CrossRef] [PubMed]
- Crabtree, M.B.; Nga, P.T.; Miller, B.R. Isolation and Characterization of a New Mosquito Flavivirus, Quang Binh Virus, from Vietnam. Arch. Virol. 2009, 154, 857–860. [Google Scholar] [CrossRef]
- Kolodziejek, J.; Pachler, K.; Bin, H.; Mendelson, E.; Shulman, L.; Orshan, L.; Nowotny, N. Barkedji Virus, a Novel Mosquito-Borne Flavivirus Identified in Culex perexiguus Mosquitoes, Israel, 2011. J. Gen. Virol. 2013, 94, 2449–2457. [Google Scholar] [CrossRef]
- Charles, J.; Tangudu, C.S.; Firth, A.E.; Blitvich, B.J. Complete Genome Sequences of Two Insect-Specific Flaviviruses. Arch. Virol. 2017, 162, 3913–3917. [Google Scholar] [CrossRef]
- Parreira, R.; Cook, S.; Lopes, Â.; Pedro, A.; Matos, D.; Paulo, A.; De Almeida, G.; Piedade, J.; Esteves, A. Genetic Characterization of an Insect-Specific Flavivirus Isolated from Culex theileri Mosquitoes Collected in Southern Portugal. Virus Res. 2012, 167, 152–161. [Google Scholar] [CrossRef]
- Carrera, J.P.; Guzman, H.; Beltrán, D.; Díaz, Y.; López-Vergès, S.; Torres-Cosme, R.; Popov, V.; Widen, S.G.; Wood, T.G.; Weaver, S.C.; et al. Mercadeo Virus: A Novel Mosquito-Specific Flavivirus from Panama. Am. J. Trop. Med. Hyg. 2015, 93, 1014–1019. [Google Scholar] [CrossRef]
- Huhtamo, E.; Cook, S.; Moureau, G.; Uzcátegui, N.Y.; Sironen, T.; Kuivanen, S.; Putkuri, N.; Kurkela, S.; Harbach, R.E.; Firth, A.E.; et al. Novel Flaviviruses from Mosquitoes: Mosquito-Specific Evolutionary Lineages within the Phylogenetic Group of Mosquito-Borne Flaviviruses. Virology 2014, 464–465, 320–329. [Google Scholar] [CrossRef] [PubMed]
- Huhtamo, E.; Putkuri, N.; Kurkela, S.; Manni, T.; Vaheri, A.; Vapalahti, O.; Uzcategui, N.Y. Characterization of a Novel Flavivirus from Mosquitoes in Northern Europe That Is Related to Mosquito-Borne Flaviviruses of the Tropics. J. Virol. 2009, 83, 9532–9540. [Google Scholar] [CrossRef]
- Huhtamo, E.; Moureau, G.; Cook, S.; Julkunen, O.; Putkuri, N.; Kurkela, S.; Uzcátegui, N.Y.; Harbach, R.E.; Gould, E.A.; Vapalahti, O.; et al. Novel Insect-Specific Flavivirus Isolated from Northern Europe. Virology 2012, 433, 471–478. [Google Scholar] [CrossRef] [PubMed]
- Pabbaraju, K.; Ho, K.C.F.; Wong, S.; Fox, J.D.; Kaplen, B.; Tyler, S.; Drebot, M.; Tilley, P.A.G. Surveillance of Mosquito-Borne Viruses in Alberta Using Reverse Transcription Polymerase Chain Reaction with Generic Primers. J. Med. Entomol. 2009, 46, 640–648. [Google Scholar] [CrossRef]
- Marklewitz, M.; Zirkel, F.; Kurth, A.; Drosten, C.; Junglena, S. Evolutionary and Phenotypic Analysis of Live Virus Isolates Suggests Arthropod Origin of a Pathogenic RNA Virus Family. Proc. Natl. Acad. Sci. USA 2015, 112, 7536–7541. [Google Scholar] [CrossRef]
- Hall-Mendelin, S.; McLean, B.J.; Bielefeldt-Ohmann, H.; Hobson-Peters, J.; Hall, R.A.; Van Den Hurk, A.F. The Insect-Specific Palm Creek Virus Modulates West Nile Virus Infection in and Transmission by Australian Mosquitoes. Parasites Vectors 2016, 9, 414. [Google Scholar] [CrossRef] [PubMed]
- Öhlund, P.; Delhomme, N.; Hayer, J.; Hesson, J.C.; Blomström, A.L. Transcriptome Analysis of an Aedes Albopictus Cell Line Single-and Dual-Infected with Lammi Virus and WNV. Int. J. Mol. Sci. 2022, 23, 875. [Google Scholar] [CrossRef]
- Klimova, E.A.; Karetkina, G.N.; Shakaryan, A.K.; Sayfullin, M.A.; Karan, L.S.; Larichev, V.F.; Grigoreva, Y.E.; Morozkin, E.S.; Lyapeikova, E.A.; Abramova, E.N.; et al. West Nile Fever on the Territory of the Moscow Agglomeration. Infect. Dis. News Opin. Train. 2021, 10, 13–21. [Google Scholar] [CrossRef]
- Öhlund, P.; Hayer, J.; Hesson, J.C.; Blomström, A.L. Small RNA Response to Infection of the Insect-Specific Lammi Virus and Hanko Virus in an Aedes albopictus Cell Line. Viruses 2021, 13, 2181. [Google Scholar] [CrossRef]
- DiMenna, M.A.; Bueno, R.J.; Parmenter, R.R.; Norris, D.E.; Sheyka, J.M.; Molina, J.L.; LaBeau, E.M.; Hatton, E.S.; Glass, G.E. Emergence of West Nile Virus in Mosquito (Diptera: Culicidae) Communities of the New Mexico Rio Grande Valley. J. Med. Entomol. 2006, 43, 594–599. [Google Scholar] [CrossRef] [PubMed]
- Khalin, A.V.; Aibulatov, S.V. Northernmost Records of Mosquito Species (Diptera: Culicidae) in Northwestern Russia. Zoosystematica Ross. 2021, 30, 46–63. [Google Scholar] [CrossRef]
- Rasnitsyn, S.P. Sampling Methods for Bloodsucking Dipterans. In Guide to Medical Entomology; Derbeneva-Ukhova, V.P., Ed.; Meditsina: Moscow, Russia, 1974; pp. 163–176. (In Russian) [Google Scholar]
- Khalin, A.V.; Aibulatov, S.V.; Przhiboro, A.A. Sampling Techniques for Bloodsucking Dipterans (Diptera: Culicidae, Simuliidae, Ceratopogonidae, Tabanidae). Entomol. Rev. 2021, 101, 1219–1243. [Google Scholar] [CrossRef]
- Becker, N.; Petrić, D.; Zgomba, M.; Boase, C.; Madon, M.; Dahl, C.; Kaiser, A. Mosquitoes: Identification, Ecology and Control, 3rd ed.; Springer Nature: Cham, Switzerland, 2020. [Google Scholar]
- Gutsevich, A.V.; Monchadsky, A.S.; Stackelberg, A. Mosquitoes of the Family Culicidae. In Fauna of the USSR. Insecta, Diptera; Bychowsky, B.E., Ed.; Nauka: Leningrad, Russia, 1970; Volume 3, p. 384. (In Russian) [Google Scholar]
- Khalin, A.V. Refinements of a Key to Species of the Genus Aedes Meigen (Diptera, Culicidae) from Northwestern Russia Based on the Male Genitalia Characters. Entomol. Rev. 2009, 89, 1137–1153. [Google Scholar] [CrossRef]
- Scaramozzino, N.; Crance, J.-M.; Jouan, A.; DeBriel, D.A.; Stoll, F.; Garin, D. Comparison of Flavivirus Universal Primer Pairs and Development of a Rapid, Highly Sensitive Heminested Reverse Transcription-PCR Assay for Detection of Flaviviruses Targeted to a Conserved Region of the NS5 Gene Sequences. J. Clin. Microbiol. 2001, 39, 1922–1927. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Nei, M. Estimation of the Number of Nucleotide Substitutions in the Control Region of Mitochondrial DNA in Humans and Chimpanzees. Mol. Biol. Evol. 1993, 10, 512–526. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Bolling, B.G.; Weaver, S.C.; Tesh, R.B.; Vasilakis, N. Insect-Specific Virus Discovery: Significance for the Arbovirus Community. Viruses 2015, 7, 4911–4928. [Google Scholar] [CrossRef]
- Litov, A.G.; Belova, O.A.; Kholodilov, I.S.; Kalyanova, A.S.; Gadzhikurbanov, M.N.; Rogova, A.A.; Gmyl, L.V.; Karganova, G.G. Viromes of Tabanids from Russia. Viruses 2023, 15, 2368. [Google Scholar] [CrossRef]
- Shi, H.; Yu, X.; Cheng, G. Impact of the Microbiome on Mosquito-Borne Diseases. Protein Cell 2023, 14, 743–761. [Google Scholar] [CrossRef]
- Carvalho, V.L.; Long, M.T. Insect-Specific Viruses: An Overview and Their Relationship to Arboviruses of Concern to Humans and Animals. Virology 2021, 557, 34–43. [Google Scholar] [CrossRef]
- Olmo, R.P.; Todjro, Y.M.H.; Aguiar, E.R.G.R.; de Almeida, J.P.P.; Ferreira, F.V.; Armache, J.N.; de Faria, I.J.S.; Ferreira, A.G.A.; Amadou, S.C.G.; Silva, A.T.S.; et al. Mosquito Vector Competence for Dengue Is Modulated by Insect-Specific Viruses. Nat. Microbiol. 2023, 8, 135–149. [Google Scholar] [CrossRef]
- McLean, B.J.; Hall-Mendelin, S.; Webb, C.E.; Bielefeldt-Ohmann, H.; Ritchie, S.A.; Hobson-Peters, J.; Hall, R.A.; Van Den Hurk, A.F. The Insect-Specific Parramatta River Virus Is Vertically Transmitted by Aedes vigilax Mosquitoes and Suppresses Replication of Pathogenic Flaviviruses In Vitro. Vector-Borne Zoonotic Dis. 2021, 21, 208–215. [Google Scholar] [CrossRef]
- Koh, C.; Henrion-Lacritick, A.; Frangeul, L.; Saleh, M.C. Interactions of the Insect-Specific Palm Creek Virus with Zika and Chikungunya Viruses in Aedes Mosquitoes. Microorganisms 2021, 9, 1652. [Google Scholar] [CrossRef]
- Khalin, A.V.; Aibulatov, S.V.; Filonenko, I.V. Mosquito Distribution in Northwestern Russia: Species of the Genus Aedes Meigen (Diptera, Culicidae). Entomol. Rev. 2021, 101, 1060–1095. [Google Scholar] [CrossRef]
- Robert, V.; Günay, F.; Le Goff, G.; Boussès, P.; Sulesco, T.; Khalin, A.; Medlock, J.M.; Kampen, H.; Petrić, D.; Schaffner, F. Distribution Chart for Euro-Mediterranean Mosquitoes (Western Palaearctic Region). J. Eur. Mosq. Control Assoc. 2019, 37, 1–28. [Google Scholar]
- Mohrig, W. Die Culiciden Deutschlands. Untersuchungen zur Taxonomie, Biologie und Ökologie der Einheimischen Stechmücken. Parasitologische Schriftenreihe; Heft 18; VEB Gustav Fischer Verlag: Jena, Germany, 1969. [Google Scholar]
- Suslo, D.S.; Khalin, A.V. Seasonal Activity of Mosquito Larvae (Diptera, Culicidae) in the Berezinsky Biosphere Reserve (the Republic of Belarus). Entomol. Rev. 2023, 103, 131–143. [Google Scholar] [CrossRef]
- Suslo, D.S.; Khalin, A.V. Seasonal Activity of Female Mosquitoes (Diptera, Culicidae) in the Berezinsky Biosphere Reserve (Republic of Belarus). Entomol. Rev. 2022, 102, 928–938. [Google Scholar] [CrossRef]


{kind=link}
| Mosquito Species | Mosquito Stage at Collection | Mosquito Stage in Study (Gender) | Number of Individuals |
|---|---|---|---|
| Saint Petersburg, Park Sosnovaya Polyana (59.831347 N, 30.138416 E) | |||
| Aedes cinereus | larvae/pupae | adult (m+f) | 19 * |
| larvae | adult (m+f) | 21 | |
| larvae | 51 | ||
| Aedes cantans | adult | adult (f) | 4 |
| larvae/pupae | adult (m+f) | 3 | |
| pupae | adult (m/f) | 171 | |
| pupae | 44 | ||
| larvae | adult (m/f) | 68 | |
| larvae | 5 | ||
| Aedes communis | adult | adult (f) | 7 |
| pupae | adult (m+f) | 21 | |
| larvae | adult (m+f, f) | 87 | |
| Aedes punctor | pupae | adult (f) | 3 |
| larvae | adult (f) | 6 | |
| Aedes dianteus | larvae | adult (f) | 1 |
| Aedes spp. | larvae | pupae | 4 |
| larvae | 70 | ||
| Saint Petersburg, Polezhaevsky park (59.841955 N, 30.190558 E) | |||
| Aedes cinereus | adult | adult (f) | 4 |
| larvae/pupae | adult (f) | 4 | |
| pupae | adult (m+f) | 12 | |
| larvae | adult (m/f) | 62 | |
| Aedes cantans | adult | adult (f) | 90 |
| Aedes communis | adult | adult (f) | 5 |
| pupae | adult (m+f) | 10 | |
| larvae | adult (m+f) | 11 | |
| Aedes punctor | adult | adult (f) | 4 |
| Aedes dianteus | larvae | adult (f) | 6 |
| Aedes spp. | larvae | larvae | 60 |
| Saint Petersburg, Shungerovskiy Lesopark (59.836207 N, 30.046358 E) | |||
| Aedes communis | adult | adult (f) | 12 |
| pupae | adult (m) | 38 | |
| larvae | adult (m/f) | 287 | |
| larvae | 22 | ||
| Aedes punctor | adult | adult (f) | 2 |
| larvae | adult (m/f) | 13 | |
| larvae | 30 | ||
| Aedes spp. | larvae | pupae | 40 |
| Leningrad Region, Lomonosov District (59.722681 N, 30.179718 E) | |||
| Aedes cinereus | larvae | adult (m+f) | 28 |
| Culex territans | larvae | adult (m+f) | 24 |
| larvae | 25 | ||
| Anopheles claviger | pupae | adult (f) | 2 |
| Culiseta morsitans | larvae | adult (m+f) | 20 |
| Total: | 1396 | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kholodilov, I.S.; Aibulatov, S.V.; Khalin, A.V.; Polienko, A.E.; Klimentov, A.S.; Belova, O.A.; Rogova, A.A.; Medvedev, S.G.; Karganova, G.G. Orthoflavivirus Lammi in Russia: Possible Transovarial Transmission and Trans-Stadial Survival in Aedes cinereus (Diptera, Culicidae). Viruses 2024, 16, 527. https://doi.org/10.3390/v16040527
Kholodilov IS, Aibulatov SV, Khalin AV, Polienko AE, Klimentov AS, Belova OA, Rogova AA, Medvedev SG, Karganova GG. Orthoflavivirus Lammi in Russia: Possible Transovarial Transmission and Trans-Stadial Survival in Aedes cinereus (Diptera, Culicidae). Viruses. 2024; 16(4):527. https://doi.org/10.3390/v16040527
Chicago/Turabian StyleKholodilov, Ivan S., Sergey V. Aibulatov, Alexei V. Khalin, Alexandra E. Polienko, Alexander S. Klimentov, Oxana A. Belova, Anastasiya A. Rogova, Sergey G. Medvedev, and Galina G. Karganova. 2024. "Orthoflavivirus Lammi in Russia: Possible Transovarial Transmission and Trans-Stadial Survival in Aedes cinereus (Diptera, Culicidae)" Viruses 16, no. 4: 527. https://doi.org/10.3390/v16040527