
Dissemination of the Flavivirus Subgenomic Replicon Genome and Viral Proteins by Extracellular Vesicles
1
Department of Microbiology, Dokkyo Medical University School of Medicine, 880 Kita-kobayashi, Mibu 321-0293, Tochigi, Japan
2
Department of Pathology, Dokkyo Medical University Hospital, 880 Kita-kobayashi, Mibu 321-0293, Tochigi, Japan
*
Author to whom correspondence should be addressed.
Viruses 2024, 16(4), 524; https://doi.org/10.3390/v16040524
Submission received: 4 March 2024
/
Revised: 22 March 2024
/
Accepted: 26 March 2024
/
Published: 28 March 2024
(This article belongs to the Special Issue Boosting Flavivirus Research: A Pandengue Net Initiative)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Extracellular vesicles (EVs) such as exosomes have been shown to play physiological roles in cell-to-cell communication by delivering various proteins and nucleic acids. In addition, several studies revealed that the EVs derived from the cells that are infected with certain viruses could transfer the full-length viral genomes, resulting in EVs-mediated virus propagation. However, the possibility cannot be excluded that the prepared EVs were contaminated with infectious viral particles. In this study, the cells that harbor subgenomic replicon derived from the Japanese encephalitis virus and dengue virus without producing any replication-competent viruses were employed as the EV donor. It was demonstrated that the EVs in the culture supernatants of those cells were able to transfer the replicon genome to other cells of various types. It was also shown that the EVs were incorporated by the recipient cells primarily through macropinocytosis after interaction with CD33 and Tim-1/Tim-4 on HeLa and K562 cells, respectively. Since the methods used in this study are free from contamination with infectious viral particles, it is unequivocally indicated that the flavivirus genome can be transferred by EVs from cell to cell, suggesting that this pathway, in addition to the classical receptor-mediated infection, may play some roles in the viral propagation and pathogenesis.
1. Introduction
Genus Flavivirus consists of more than 70 members including important pathogens such as Japanese encephalitis virus (JEV) and dengue virus (DENV) [1]. Flaviviruses are distributed worldwide and billions of people are at risk for the diseases caused by flavivirus infection [2]. Flavivirus infection leads to various symptoms ranging from mild flu-like illness to severe diseases, such as meningitis and encephalitis for JEV and hemorrhagic fever for DENV [3]. However, the pathological mechanisms of the diseases caused by flaviviruses have not been fully elucidated.
JEV and DENV are arthropod-borne enveloped viruses, which approximately contain 11-kb positive-strand RNA genomes. The viral genome encodes three structural proteins, namely capsid, precursor membrane (prM), and envelope (E), as well as seven nonstructural (NS) proteins, namely NS1, NS2A, NS2B, NS3, NS4A, NS4B, and NS5 [4]. The infectious virion is approximately 50–70 nm in diameter and contains mature M and E proteins on its surface. The E protein contains the receptor-binding domain and the fusion peptide. The NS proteins are known to be involved in viral replication and also modulation of host immune responses [5]. For example, the NS5 protein possesses a guanylyl transferase/methyltransferase domain and an RNA-dependent RNA polymerase domain essential for viral genome replication, while it is known to block type I interferon signaling as well [5].
Extracellular vesicles (EVs) have been known to contain structures such as exosomes and microvesicles. Exosomes are extracellular vesicles sized approximately 40–100 nm in diameter and are formed within a multivesicular endosome (MVE) and released when the MVE is fused with the plasma membrane, whereas microvesicles are extracellular vesicles sized 50–1000 nm in diameter and formed by budding from the plasma membrane [6]. Most of the cell types are known to produce and release EVs. When EVs were originally discovered in the 1980s, their functions were unknown. However, recent studies have demonstrated that EVs bear a variety of physiological and pathological functions. For instance, EVs carry membrane proteins, intra-cellular proteins, extra-cellular proteins, DNA, and RNA and play roles in cell-to-cell communications by delivering those cargoes [7]. It has also been reported that EVs are involved in tumor growth and tumor metastasis [8,9]. In addition, recent studies have revealed that EVs are involved in the infection of certain viruses by transferring viral components and host cellular factors [10]. For example, exosomes produced by the cells infected with human immunodeficiency virus (HIV), human T-cell leukemia virus 1, or hepatitis C virus (HCV) were shown to deliver viral proteins [11,12,13]. It has also been reported that EVs can deliver viral genomes of HIV, HCV, human pegi virus, hepatitis A virus, and enterovirus, contributing to virus dissemination [13,14,15,16,17]. Furthermore, it has been shown that EVs, which contain cellular factors, such as miRNA and proteins, could affect immune responses, suggesting that EVs might be involved in viral pathogenesis [18]. These findings imply that virus propagation could be mediated by EVs in addition to the classical receptor-mediated infection. As to flavivirus, several studies have suggested that its genome RNA could be transferred by EVs from cell to cell [19,20,21]. However, those studies utilized the EVs derived from virus-infected cells, raising the possibility that the EV samples may have been contaminated with infectious viral particles.
To exclude this possibility, the cells that harbor subgenomic replicon derived from JEV and DENV without producing any replication-competent virus were employed in this study as the EV donor. Since the replicon carries the luciferase gene, the transfer of the replicon RNA could be quantitatively evaluated by measuring the luciferase activity in the recipient cells. The results indicated that the EVs produced by the replicon-harboring cells contained the replicon RNA, which could be transferred to other cells of various types and replicated in the recipient cells. In addition, the EVs were found to contain viral proteins including the non-structural proteins that are not normally found in the viral particles. Among those, DENV NS1 proteins were shown to exist inside the EVs. This is the first study that unequivocally demonstrated EV-mediated cell-to-cell transfer of flaviviral RNA. The results suggest that EV-mediated transfer of the viral components, in addition to the classical receptor-mediated infection, may play previously unknown roles in the propagation and pathogenesis of flaviviruses.
2. Materials and Methods
2.1. Cells and Viruses
Vero (monkey kidney), HeLa (human cervix adenocarcinoma), PK15 (porcine kidney), and SK-N-SH (human neuroblastoma) cells were grown at 37 °C with 5% CO2 in Eagle’s minimum essential media (MEM), supplemented with 10% fetal bovine serum (FBS), 10 mM nonessential amino acid (NEAA; Thermo Fisher Scientific, Waltham, MA, USA), and penicillin (1000 unit/mL)-streptomycin (100 µg/mL) (PS). BHK (hamster kidney) and HepG2 (human hepatoma) cells were grown at 37 °C with 5% CO2 in Dulbecco’s modified MEM supplemented with 10% FBS, NEAA, and PS. K562 (human leukemia) cells were grown at 37 °C with 5% CO2 in RPMI1640 media supplemented with 10% FBS. C6/36 (Aedes albopictus mosquito) cells were grown at 30 °C with 5% CO2 in MEM supplemented with 10% FBS, NEAA, and PS. JEV of the Muar strain and DENV of the New Guinea C (NGC) strain were propagated in C6/36 as described previously [22]. Following virus infection and EV inoculation, cells were maintained in MEM supplemented with 1% FBS, NEAA, and 10 mM HEPES (MEM-1%FBS).
2.2. Establishment of Subgenomic Replicon Harboring Cell Lines
To construct the subgenomic replicon, JM-PnL (Figure 1), the puromycin N-acetyl-transferase (PAC), and the deep sea shrimp-derived nano-luciferase (nLuc) genes were individually amplified by PCR and the obtained DNA fragments were inserted into the previously reported JEV Muar strain-based replicon clone [23]. A similar strategy was applied to construct the DENV NGC-derived replicon, DN-PnL (Figure 1), bearing the PAC and nLuc genes. The obtained clones of plasmid DNA were linearized by Swa I digestion and then subjected to in vitro RNA transcription using a MEGAscript T7 Kit (Thermo Fisher Scientific). The prepared replicon RNAs were electroporated into BHK cells by using the GENE PULSER II (Bio Rad, Hercules, CA, USA) at 500 V and 25 µF [22]. To select the cells successfully harboring the replicon RNA, puromycin was added at 10 µg/mL from the next day and the surviving cells were designated as BHK/JM-PnL and BHK/DN-PnL, respectively.
2.3. Immunostaining
Expression of the viral proteins was detected by immunostaining as described previously [22]. Briefly, cells fixed with 50% acetone–50% methanol solution were serially incubated with the 6H4 anti-NS1 mouse monoclonal antibody [24], biotinylated anti-mouse IgG (Vector Laboratories, Burlingame, CA, USA), and peroxidase-conjugated avidin-biotin complex (Vector Laboratories). The cells with virus protein expression were visualized by incubation with the peroxidase substrate, 0.02% 3,3′-diaminobenzidine in 0.008% H2O2.
2.4. EV Sample Preparation
The BHK/JM-PnL and BHK/DN-PnL cells in the culture plates were rinsed with PBS and then cultured in 10% VP-SFM (Thermo Fisher Scientific)–90% MEM supplemented with NEAA (VPSFM-MEM). Then, the culture supernatants were collected daily and subjected to centrifugation at 3000 rpm for 5 min to remove cell debris. The cleared culture fluids were concentrated by about 100-fold with the Amicon Ultra 100 K (Merck, Burlington, MA, USA). Approximately 1 mL of the concentrated culture fluids were subjected to EV isolation by using the MagCapture™ exosome isolation kit PS (FUJIFILM Wako Pure Chemical Corporation, Osaka, Japan) according to the manufacturer’s instruction except that the addition of the beads was repeated three times for each culture fluid preparation. All of the EVs eluted from the three batches of beads were combined and used as the EV sample derived from each fluid preparation. After EV isolation with the beads, the remaining concentrated culture fluids were used as EV-depleted samples in some analyses.
In some experiments where EV production needs to be inhibited, GW4869 (7.5 µM in DMSO; Cayman Chemical, Ann Arbor, MI, USA) was added 24 h before changing the media to VPSFM-MEM, which also contained GW4869.
2.5. Electron Microscopy
Five microliters of the isolated EV sample were applied on a formvar-coated grid (Nisshin EM Co., Ltd., Tokyo, Japan) and incubated for 2 min at room temperature. The grid was stained with 5% uranyl acetate for 30 s at room temperature. The images were obtained by the transmission electron microscope HT7800 (Hitachi High-Tech Corporation, Tokyo, Japan). The diameter of the observed EVs was measured (n = 20).
2.6. EV Inoculation and Luciferase Assay
The prepared EVs and the EV-depleted culture supernatants were diluted in VPSFM-MEM 20-fold and then inoculated onto the target cells that had been seeded in a 96-well plate and rinsed with PBS before the inoculation. In some experiments, the EVs incubated in 0.5 mg/mL of Triton X-100 (Tx-100) at room temperature for 5 min were diluted by 30-fold in VPSFM-MEM and then inoculated onto BHK cells. In some other experiments, the EVs were treated before inoculation with 5 µg/mL RNase A (Nippon Gene, Tokyo, Japan) for 15 min at 37 °C and diluted in VPSFM-MEM by 20-fold.
After inoculation with the EVs, the cells were incubated at 37 °C for 4 h. Then, the inoculum was removed and the cells were maintained in MEM-1%FBS for 72 h. In some experiments, the cells were maintained in MEM-1%FBS in the presence of 300 µM ribavirin (FUJIFILM Wako Pure Chemical Corporation). At 72 h, the cells were rinsed with PBS and lysed with the cell lysis buffer supplied in the Nano-Glo Luciferase Assay System (Promega, Madison, WI, USA) and the luciferase activities were measured by the same Assay System according to the manufacturer’s instructions [23]. The luminescence was measured by a Lumat LB9501 (Berthold Technologies, Bad Wilbad, Germany).
2.7. PCR
The total RNA contained in the isolated EVs was purified by an EV RNA purification system (System Biosciences, Palo Alto, CA, USA), according to the manufacturer’s instructions. Approximately 40 ng of RNA was reverse transcribed by ReverTra Ace (TOYOBO, Tokyo, Japan) with the NS5 gene-specific primers, namely, 5′-agctcttgtacgcgttccgat-3′ and 5′-ctccttctccctccatctgtc-3′ for JEV Muar and DENV NGC strains, respectively, according to the manufacturer’s instructions. By using 3 µL of the obtained cDNA solution, PCR amplification was carried out by KOD Ver 2 (TOYOBO) according to the manufacturer’s instructions. For amplifying the nLuc gene, oligonucleotide 5′-ggtgatcctgcactatgg-3′ was used as the sense primer. As the antisense primers, oligonucleotides 5′-tcaaccacactcgagtcgacg-3′ and 5′-gagcggattccacaaatgccc-3′, which correspond to the adjacent viral sequences of JM-PnL and DN-PnL, respectively, were used. For amplifying the viral NS5 gene, oligonucleotides 5′-tggcagcaagtccct-3′ and 5′- agctcttgtacgcgttccgat-3′ were used as sense and antisense primers, respectively, for the JEV Muar strain. Similarly, oligonucleotides 5′-ggtgagaagcaatgcagcctt-3′ and 5′-tccataggtaccaacttgtcc were used for amplifying the NS5 gene of the DENV NGC strain.
2.8. Real-Time PCR
Total RNAs were obtained by CellAmp™ Direct RNA Prep Kit for RT-PCR (TaKaRa Bio Inc., Shiga, Japan) from C6/36 cells in a 96-well plate inoculated with EVs derived from BHK/JM-PnL and BHK/DN-PnL cells. Total RNAs of the EVs treated with RNase A in the presence/absence of Tx-100 were isolated by a microRNA Extractor® Kit for Purified EV (FUJIFILM Wako Pure Chemical Corporation). JM-PnL and DN-PnL replicon genomes were quantified by RNA-direct™ SYBR® Green Realtime PCR Master Mix (TOYOBO). Primers used for JM-PnL were sense, 5′-ccctcagaaccgtctcggaa-3′, and anti-sense, 5′-ctattcccaggtgtcaatatgctgt-3′, and primers used for DN-PnL were sense, 5′-agttgttagtctacgtggaccga-3′, and anti-sense, 5′-cgcgtttcagcatattgaaag-3′. The real-time PCR was carried out by Thermal Cycler Dice Real-Time System III (Takara Bio Inc.). The in vitro-transcribed JM-PnL and DN-PnL were used as standard.
2.9. Treatment of the Cells with Antibodies and Inhibitors
For treatment with antibodies, HeLa or K562 cells were incubated with anti-CD33 (15 µg/mL; Southern Biotech, Birmingham, AL, USA), anti-Tim-1 (20 µg/mL; BioLegend, San Diego, CA, USA), or anti-Tim-4 antibody (20 µg/mL; Novus Biologicals, Centennial, CO, USA) for 1 h at 37 °C. The prepared EV samples were then inoculated onto the cells in the presence of the antibodies. After incubation for 4 h at 37 °C, the inoculum was removed and the cells were maintained in MEM-1%FBS for 72 h before the luciferase assay.
For treatment with various entry inhibitors, HeLa cells prepared in the 96-well plate were incubated with Pitstop 2 (30 µM; Merck), LY294002 (5 µM; FUJIFILM Wako Pure Chemical Corporation), dynasore (80 µM; Tokyo Chemical Industry Co., Tokyo, Japan), genistein (150 µM; FUJIFILM Wako Pure Chemical Corporation), or 5-[N-ethyl-N-isopropyl] amiloride (EIPA: 100 µM; Cayman Chemical) for 30 min at 37 °C. The prepared EV samples were then inoculated onto the cells in the presence of entry inhibitors. Four hours later, the cells were rinsed with 0.5 M NaCl–0.2 M acetic acid solution and then maintained in MEM–1%FBS for 72 h before the luciferase assay.
2.10. Western Blot
The EVs, culture supernatants derived from BHK/JM-PnL and BHK/DN-PnL cells and EV-depleted culture supernatants added with the NuPAGE LDS sample buffer (Thermo Fisher Scientific) containing 100 mM dithiothretiol were fractionated on 4–12% gradient polyacrylamide gels (NuPAGE; Thermo Fisher Scientific) in the NuPAGE MOPS SDS Running Buffer (Thermo Fisher Scientific) for 80 min at 150 V and then were transferred to Immobilon-P membranes (Merck) in the NuPAGE Transfer Buffer (Thermo Fisher Scientific) for 80 min at 30 V. The membranes were treated with the blocking buffer containing 0.05% Tween 20 and 5% skim milk in 50 mM Tris-buffered saline (pH 8.0) and serially incubated with anti-JEV (NS1, NS2B, NS3, NS4B, and NS5) or anti-DENV (NS1, NS2B, NS3, NS4B, and NS5) rabbit monoclonal antibodies (Gene Tex, Irvine, CA, USA), horseradish peroxidase-conjugated anti-rabbit IgG (Cytiva, Tokyo, Japan), and Lumi-Light Plus (Merck). To detect luminescence, the membranes were examined by LuminoGraph I (ATTO, Tokyo, Japan).
2.11. Proteinase K Protection Assay
EVs were incubated with proteinase K (100 µg/mL) in the presence/absence of Tx-100 (0.5%) for 30 min on ice and then phenylmethylsulfonyl fluoride (2 mM) was added. Those samples were subjected to Western blot as described above.
2.12. Statistical Analysis
Statistical analysis of the data was performed with the Student’s t-test. For multiple comparisons, the Holm–Bonferroni method was used for correction. Differences were considered statistically significant when the p-value was less than 0.05.
3. Results
3.1. Establishment of JEV and DENV Subgenomic Replicon Expressing Cells
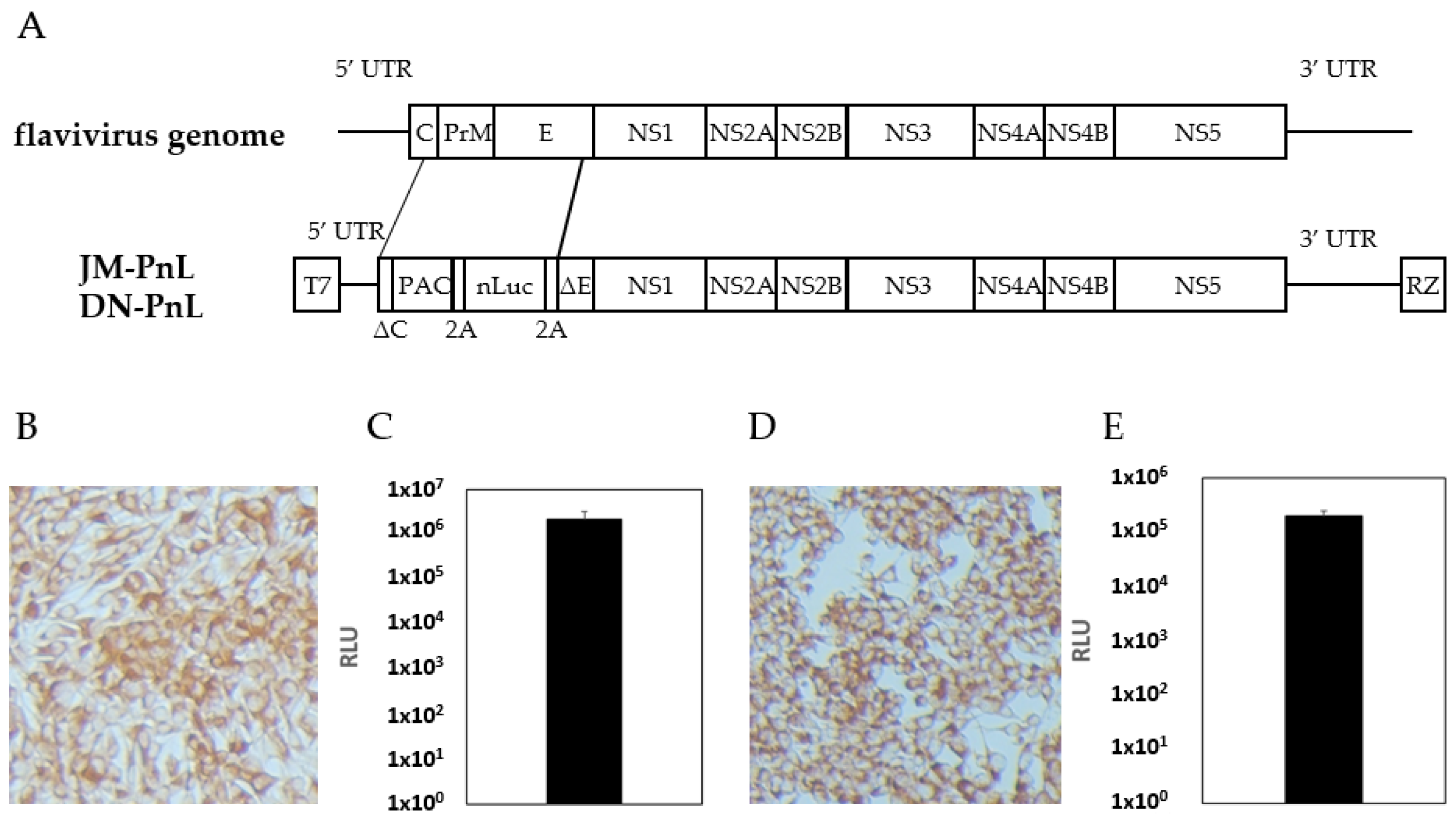
To establish subgenomic replicon harboring cells as EV donor cells, DNA clones of the nLuc gene-bearing replicons JM-PnL and DN-PnL were constructed from the infectious clones of JEV (Muar strain) and DENV (NGC strain), respectively (Figure 1A). The DNA clones were used as templates of in vitro transcription of the replicon RNA, which was then electroporated into BHK cells. Successful introduction of the replicon was confirmed by the detection of the replicon-coded JEV and DENV proteins and of luciferase activity (Figure 1B–E). The obtained BHK cells harboring JM-PnL and DN-PnL were designated as BHK/JM-PnL and BHK/DN-PnL, respectively.
3.2. Incorporation of the Replicon Genome and Viral Proteins in the EVs
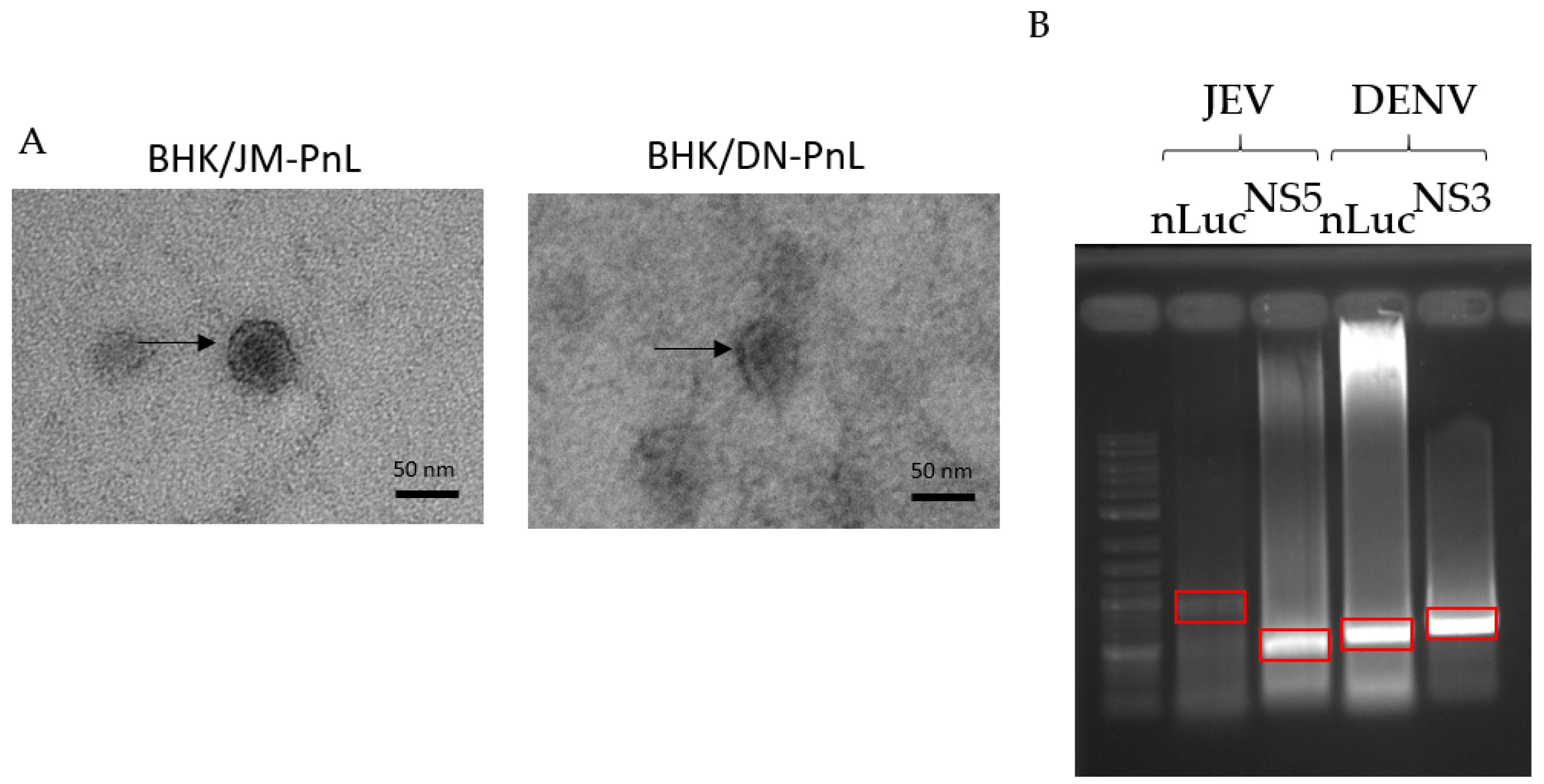
In order to elucidate the functions of EVs derived from BHK/JM-PnL and BHK/DN-PnL cells, EVs were isolated from the concentrated culture supernatants of those cells by using phosphatidyl serine affinity beads. Electron microscopy revealed that the extracellular vesicles in the prepared samples were consistent with EVs in shape (characterized as cup-shape) and size (Averages in diameter of the EVs derived from BHK/JM-PnL and BHK/DN-PnL cells were 39.6 ± 5.01 nm and 40.2 ± 5.45 nm, respectively) (Figure 2A) [25]. To examine whether the replicon genome maintained in BHK cells could be packaged in those EVs, the total RNA extracted from the EV samples was reverse transcribed and by using the obtained cDNA as a template, PCR amplification of nLuc and viral NS5 genes was performed. As shown in Figure 2B, the DNA fragments with expected sizes for nLuc and NS5 genes were amplified for both EV samples derived from BHK/JM-PnL and BHK/DN-PnL. These results suggested that the replicon genome maintained in BHK cells was associated with the EVs.
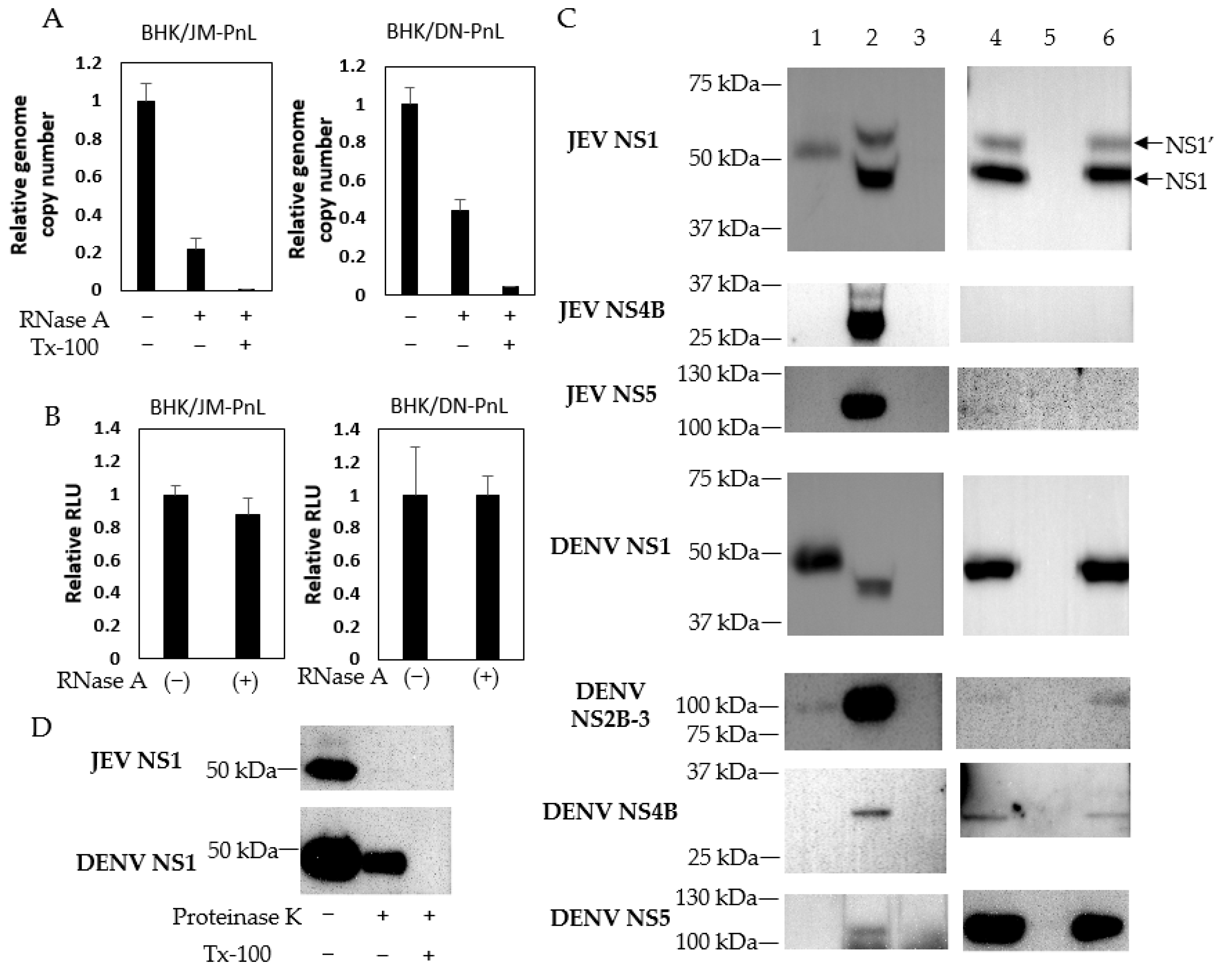
To analyze whether the replicon genome was packaged inside of the EVs or attached to the surface of the EVs, the EV samples were treated with RNase A in the presence/absence of Tx-100. RNase A resistant JM-PnL and DN-PnL replicon genomes were detected, whereas those were diminished when treated with RNase A in the presence of Tx-100 (Figure 3A). In addition, the RNase A treatment did not affect luciferase activities in the EVs-inoculated cells (Figure 3B). These results suggested that the replicon genome could be packaged inside of the EVs.
Whereas Western blot analysis detected viral NS1 and NS1′ proteins of approximately 45 kDa and 55 kDa, respectively, in the JEV-infected BHK cells, EVs produced by BHK/JM-PnL cells were shown to contain the protein of approximately 50 kDa reactive with the anti-NS1 antibody (Figure 3C). Similarly, EVs produced by BHK/DN-PnL cells contained the protein of approximately 50 kDa reactive with the anti-NS1 antibody while viral NS1 protein of approximately 45 kDa was detected in the DENV-infected BHK cells. In addition, the NS2B-NS3 complex of 100 kDa appeared to be incorporated in the EV produced by BHK/DN-PnL cells (Figure 3C) but not BHK/JM-PnL cells. The incorporation of other viral proteins encoded by the replicon, such as NS4B and NS5, into the EV derived from both BHK/JM-PnL and BHK/DN-PnL cells was not detected (Figure 3C).
To see if the detected viral proteins were packaged inside of the EVs or attached to the surface of the EVs, the resistance of those proteins to proteinase K in the presence/absence of Tx-100 was examined. DENV NS1 was reduced but detected after treatment with proteinase K alone, whereas DENV NS1 was diminished by treatment with proteinase K in the presence of Tx-100 (Figure 3D). Those results indicated that DENV NS1 exists both inside of the EVs and on the surface. In contrast, JEV NS1 existing inside of the EVs was not detected, thus most of the JEV NS1 seems to be attached to the surface of the EVs. Those viral proteins remained in the culture fluids after EV isolations, indicating that viral proteins associated with EVs are not the majority among viral proteins detected in the culture supernatants.
3.3. Cell-to-Cell Transfer of the Replicon Genome Mediated by EVs
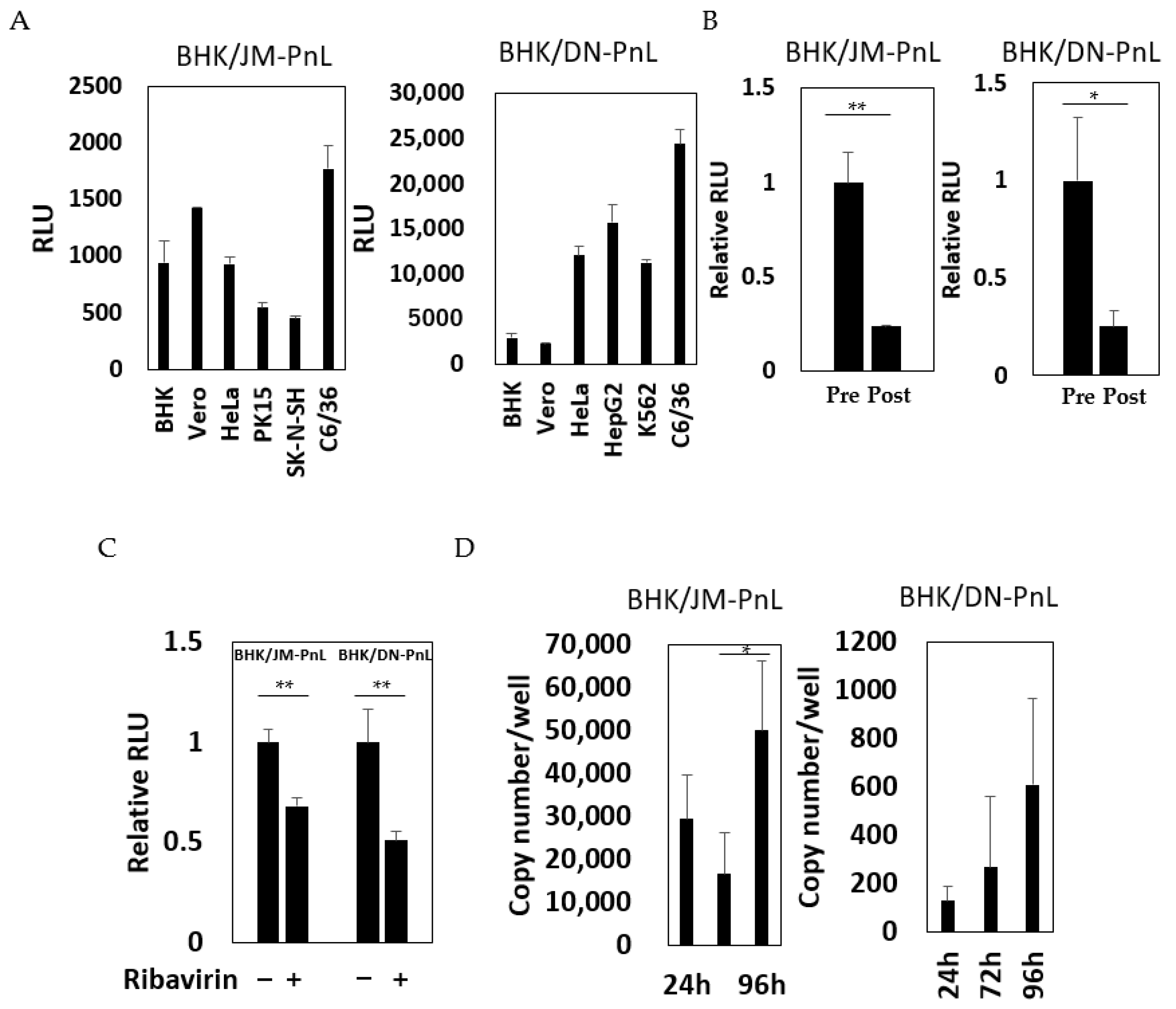
In order to test whether the replicon RNA associated with the EVs could be transferred to other cells, the EV samples prepared from the culture supernatants of BHK/JM-PnL and BHK/DN-PnL cells were inoculated onto various recipient cells. Luciferase activities were detected in all of the recipient cell lines tested (Figure 4A), suggesting that the replicon RNA could be transferred by EVs. To confirm that the detected luciferase activities resulted from the transfer of EV-associated replicon RNA, the EV-depleted culture supernatants of BHK/JM-PnL and BHK/DN-PnL cells were inoculated onto BHK cells. The luciferase activities of the cells with the EV-depleted culture supernatants were decreased, indicating that EVs could mediate cell-to-cell transfer of the replicon RNA genomes (Figure 4B). To further examine if luciferase activities detected in the EVs-inoculated cells resulted from replication of the transferred replicon genomes, the effects of ribavirin on the luciferase activities in the EV-inoculated cells were examined and replicon genomes in the EV-inoculated cells were quantified by real-time PCR. Ribavirin treatment reduced luciferase activities (Figure 4C) and the replicon genomes in the recipient cells were increased (Figure 4D), showing that transferred replicon genomes were replicable. Those results indicated the possibility that replicon RNA could be transferred from BHK/JM-PnL and BHK/DN-PnL cells to the cells of various origins in an EV-mediated manner.
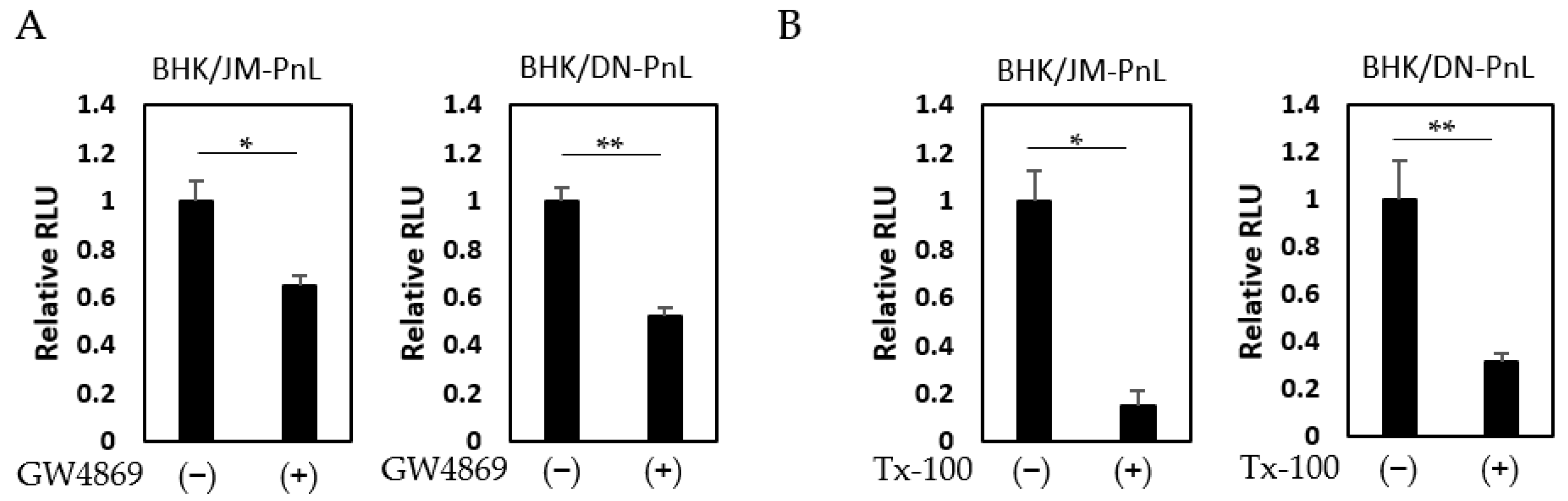
To confirm this possibility, EVs prepared from the culture supernatants of BHK/JM-PnL and BHK/DN-PnL cells that had been treated with the EV production inhibitor, GW4869 (neutral sphingomyelinase) known to inhibit EV production, were inoculated onto BHK cells. The luciferase activities in the recipient BHK cells inoculated with EVs from GW4869-treated donor cells were significantly lower compared with those in BHK cells inoculated with EVs from the untreated donor cells (Figure 5A). Because GW4869 did not affect the replication of the JM-PnL and DN-PnL (Figure S1), the reduction in the luciferase activities (Figure 5A) resulted from the lower production of EVs. Furthermore, treatment of the EV samples with Tx-100, which disrupts the EV membrane, also reduced luciferase activities (Figure 5B). These results clearly demonstrate that EVs are involved in the cell-to-cell transfer of the replicon genome RNA.
3.4. Cellular Factors Involved in the EV-Mediated Transfer of the Replicon Genome
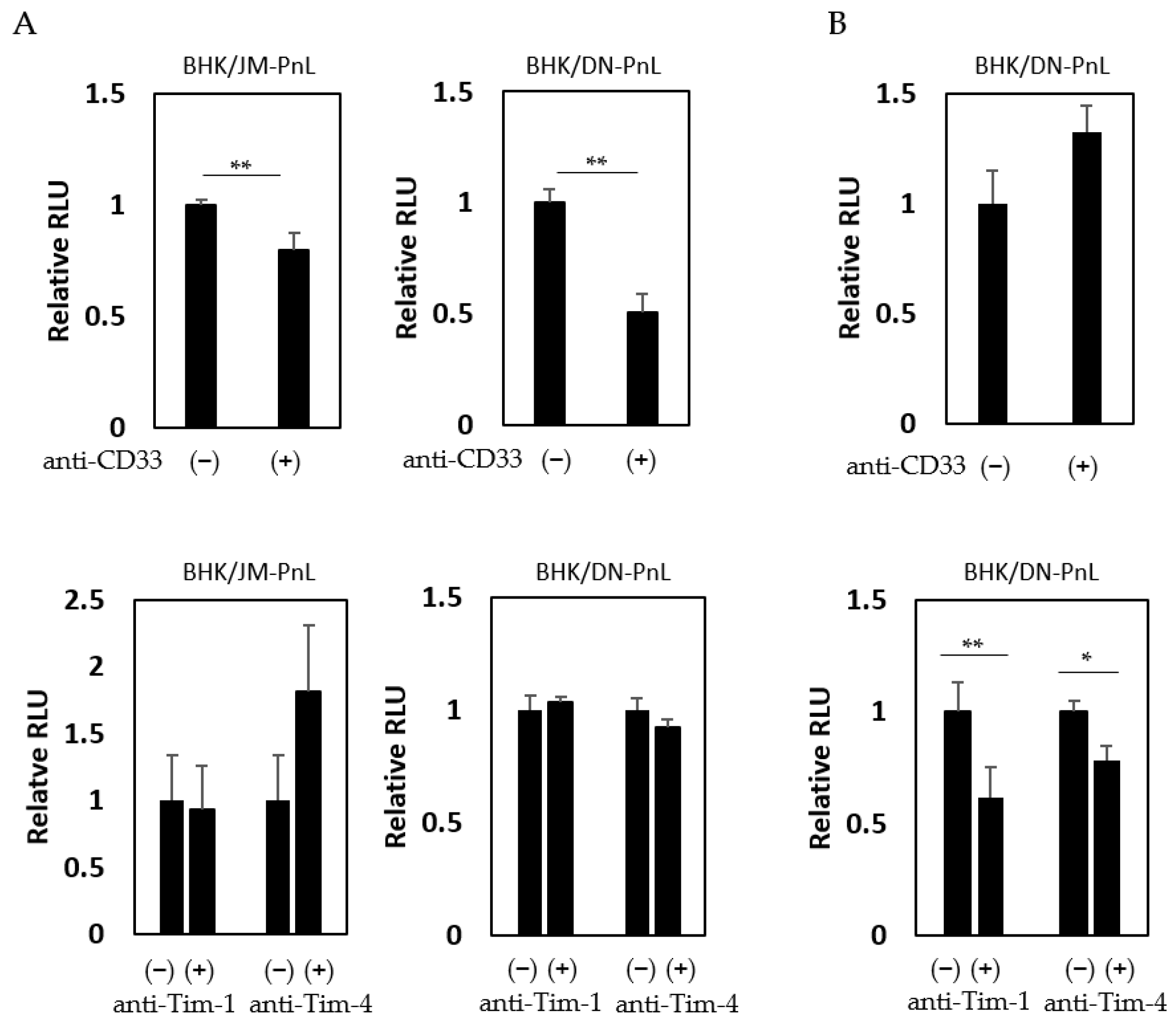
It has previously been shown that cell surface proteins, such as CD33, Tim-1, and Tim-4, are involved in EV-mediated transfer of cargo molecules [26]. To examine if these proteins are involved in the EV-mediated transfer of the replicon genome demonstrated in this study, HeLa cells were inoculated with EVs derived from BHK/JM-PnL and BHK/DN-PnL cells in the presence of the anti-CD33, anti-Tim-1, or anti-Tim-4 antibodies. Luciferase activities in the recipient cells were decreased when they were inoculated with the EVs in the presence of the anti-CD33 antibody but not the other antibodies (Figure 6A), suggesting that CD33 might be involved in the EV-mediated transfer of the replicon genome into HeLa cells. On the other hand, when K562 cells were used as the recipient cells, anti-Tim-1 and anti-Tim-4 antibodies but not anti-CD33 antibodies, reduced luciferase activities (Figure 6B), suggesting that Tim-1 and Tim-4 rather than CD33 might be crucial for the EV-mediated transfer of the replicon genome into K562 cells.
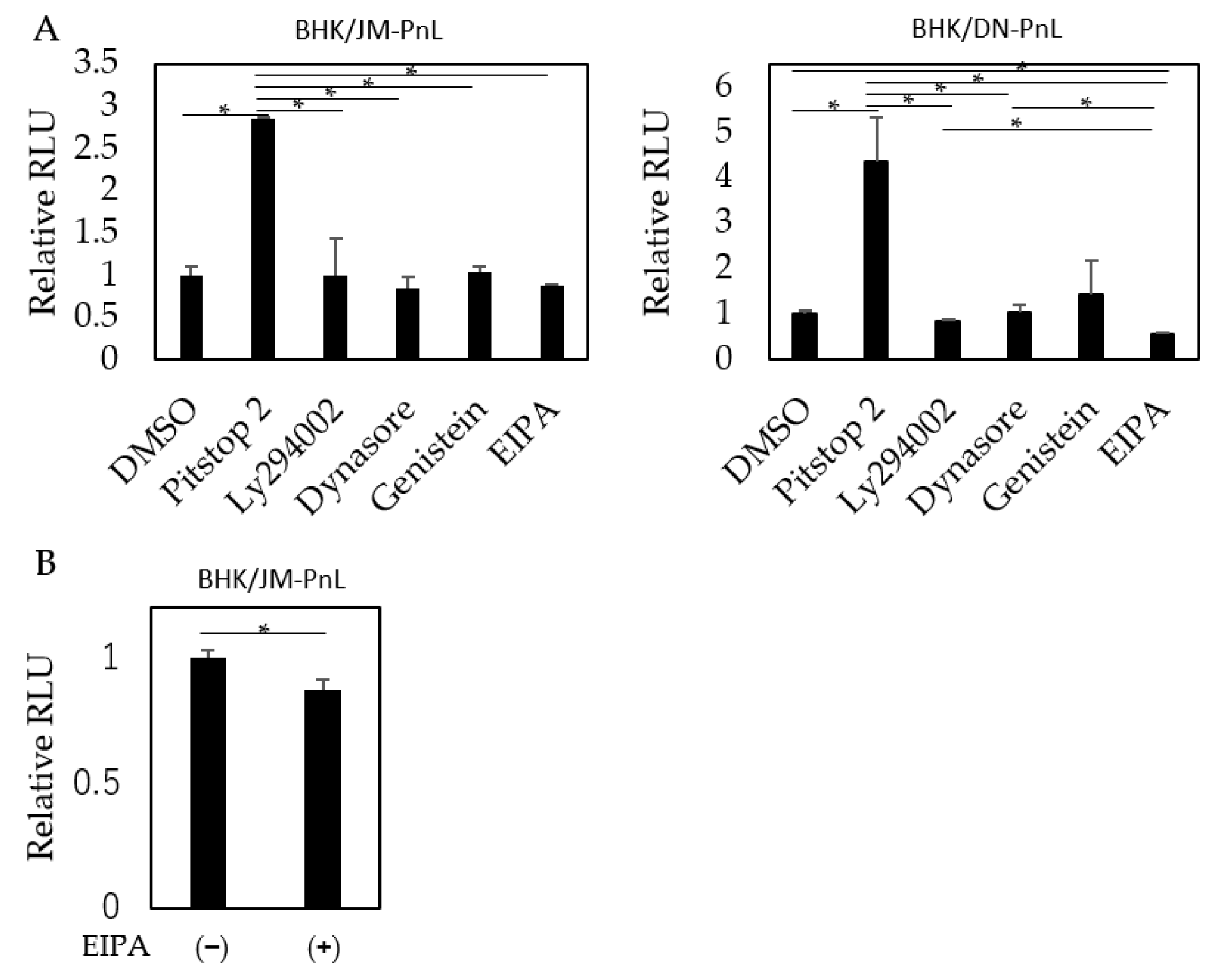
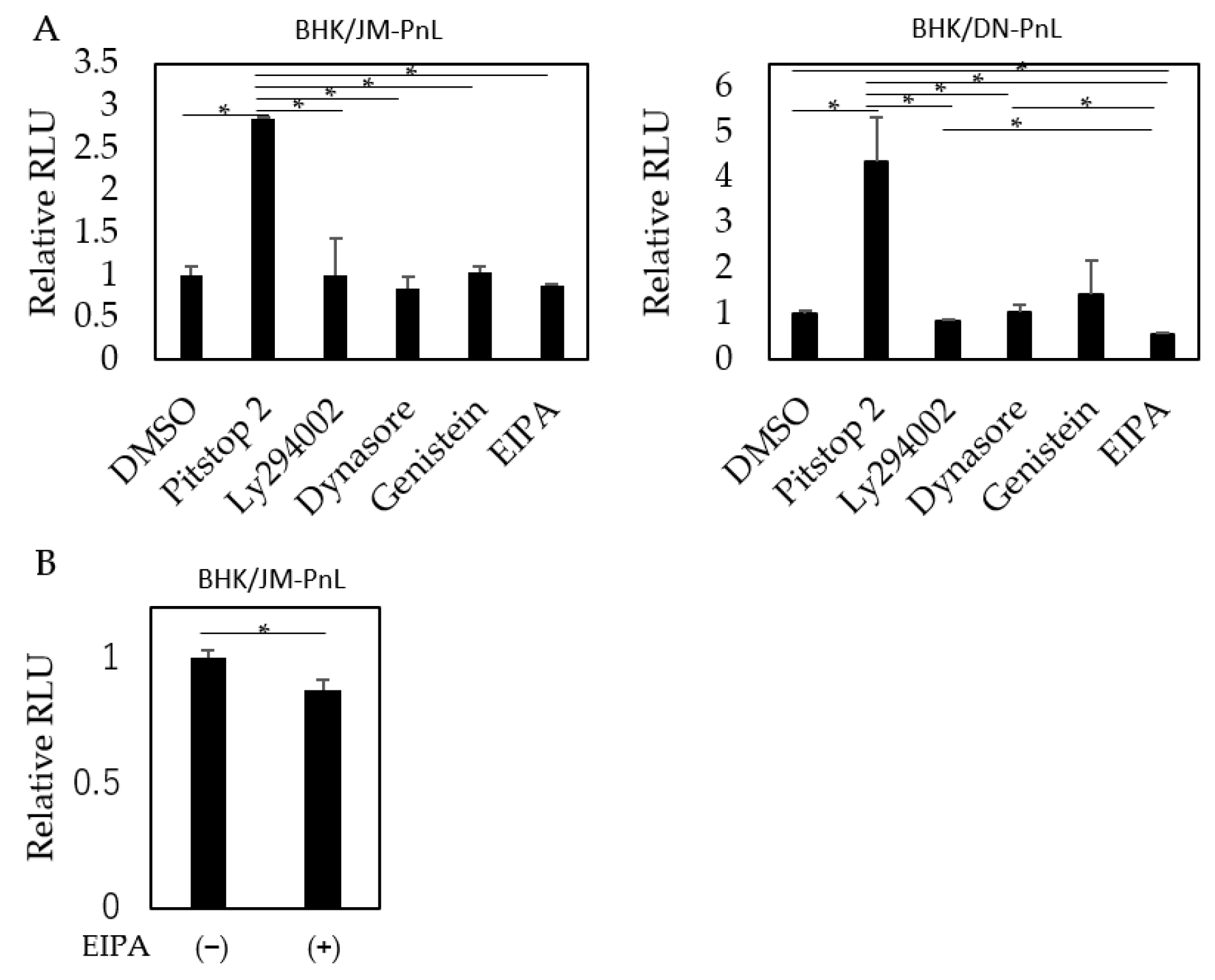
To elucidate the pathway by which the replicon genome is internalized into the recipient cells, HeLa cells were treated before EV inoculation with Pitstop 2, dynasore, and genistein, which inhibit clathrin-, dynamin-, and caveolae-mediated endocytosis, respectively; LY294002, which inhibits clathrin-independent endocytosis; or EIPA, which inhibits macropinocytosis. The results showed that luciferase activities in the recipient cells were significantly decreased when the cells were pretreated with EIPA (Figure 7A,B). Interestingly, pretreatment of the recipient cells with Pitstop 2 markedly increased luciferase activities (Figure 7).
4. Discussion
EVs, which carry biomolecules, have been shown to play a variety of physiological roles. For example, exosomes have been shown to enhance immune responses by delivering antigen-presenting molecules [27] and DNA [28]. Evidence is also accumulating that EVs are involved in the pathogenesis of various diseases. For instance, exosomes appear to promote tumor growth and metastasis by delivering molecules, such as TGF-β [29], EGFR [30], and miRNAs [31,32]. In addition, recent studies using virus-infected cells have demonstrated the incorporation of viral proteins, as well as viral and cellular RNAs, into EVs, suggesting the possibility that EVs could be involved in viral propagation, pathogenesis, and immune evasion [33,34,35,36,37,38,39,40,41,42]. Several studies reported that EVs could transfer the full-length genome of certain viruses including flaviviruses and that virus dissemination might be mediated by EVs in addition to the classical receptor-mediated infection [13,14,15,16,17,19,20,21]. However, EVs derived from virus-infected cells were used in those previous studies. Therefore, the possibility cannot be excluded that the observed phenomena were due to contamination of the infectious viral particles in the EV samples. In contrast, the cells that harbor flavivirus-derived subgenomic replicons do not produce infectious viral particles; the EVs used in this study were prepared from the culture supernatants of those cells. Therefore, this is the first study that unequivocally demonstrates the ability of EVs to mediate cell-to-cell transfer of flavivirus genome-derived RNA.
Since the ability of EVs to endow the luciferase activity to other cells was resistant to RNase A treatment, the replicon genome was likely packaged inside of the EVs (Figure 3B). The mechanism by which the replicon genome is packaged in the EVs is still unknown. It has been reported that RNA-binding proteins, such as the heterogeneous nuclear ribonucleoprotein family and the Y-box binding protein 1, contribute to the packaging of cellular RNAs into exosomes [43]. Since these cellular proteins have been reported to bind to the genome RNA of JEV and DENV [44,45,46], similar mechanisms might be involved in the incorporation of flavivirus-derived replicon genome into EVs. Figure 2B suggested that the amount of full-length replicon genome of JEV in the EVs seems to be smaller than DENV since the band corresponding to the luciferase-JEV NS1 fragment was faint. Those results implied that the packaging efficiencies of the replicon genome into EVs might be dependent on the virus sequences, which could affect the recruitment of RNA-binding proteins in the host cells to carry the viral gnomes into EVs.
In addition to the replicon genome, proteins reactive with the anti-NS1 protein were detected in the EVs and part of the proteins were shown to be packaged inside of the EVs (Figure 3C,D). The molecular weight of the detected proteins was slightly higher than that of the NS1 protein expressed in the virus-infected cells. Although the actual molecular nature of the detected protein is unknown, it is possible that it corresponds to an incompletely cleaved viral precursor protein, which contains the NS1 epitope. Alternatively, NS1 protein packaged in EVs might be modified because previous studies have demonstrated that proteins found in EVs went through post-translational modification, such as ubiquitination, SUMOylation, or UBL3 modification [47,48,49,50]. The NS2B-NS3 complex was detected in the EVs produced by BHK/DN-PnL cells but not BHK/JM-PnL cells. The reason for the discrepancy is unknown. Flavivirus nonstructural proteins are known to play roles in virus replication and immune evasion [5]. Although the precise abundance of the viral proteins incorporated in the EVs is unknown, it is possible that those viral proteins may contribute to the formation of the EVs containing the replicon genome and to the replication of the replicon genome in the recipient cells.
In this study, treatment of recipient HeLa cells but not K562 cells with the anti-CD33 antibody negatively affected the EV-mediated transfer of the flavivirus-derived replicon genome. This is consistent with the previous study showing that CD33 was involved in the exosome-mediated uptake of fluorescent dye into HeLa cells [51]. In contrast, treatment of recipient K562 cells but not HeLa cells with anti-Tim-1 and Tim-4 antibodies was shown in this study to decrease the exosome uptake. Tim-1 and Tim-4 are members of the T cell immunoglobulin and mucin-domain-containing proteins family and known to bind to phosphatidylserine [52]. While Tim-4 is primarily expressed on antigen-presenting cells, Tim-1 is expressed on various epithelial cells in addition to immune cells [52,53]. It is suggested that receptors for EV uptake are recipient cell-dependent and/or EV producer cell-dependent. Furthermore, it cannot be ruled out that receptors for viral proteins detected in EVs of those on the surface of the EVs play roles in uptake into the recipient cells.
Following the attachment to the surface of recipient cells, EVs are usually internalized into those cells. It has been shown that various mechanisms including endocytosis, macropinocytosis, phagocytosis, and membrane fusion could be involved in the internalization process [54,55,56]. In this study, EIPA treatment of the recipient HeLa cells appeared to negatively affect the internalization of the EVs. Therefore, it is likely that macropinocytosis is involved in the internalization of the EVs carrying the flavivirus-derived replicon into HeLa cells. Apparently, the results contradict the previous studies, which demonstrated clathrin-dependent endocytosis as the major pathway for internalization of the EVs bearing the full-length flaviviral genome [19,21], although partial inhibition of internalization by EIPA treatment observed in this study indicated that other untested internalization pathways might be involved in addition to macropinocytosis. However, EV samples prepared from the virus-infected cells were used in those studies. In addition, neuronal cells were used as the recipient. These differences might be responsible for the apparently incompatible results. Alternatively, the internalization pathway might be dependent on the size of the EVs. Macropinocytosis is known to uptake larger substances (up to 5 µm) than clathrin-dependent endocytosis (~100 nm) [57]. The EVs prepared from flavivirus-infected cells (about 80 to 120 nm on average) [58,59] were larger than those prepared from the replicon-harboring cells in this study (about 40 nm). Therefore, depending on the recipient cell type and the size of the EV, the internalization pathway might be different.
Interestingly, treatment of the recipient cells with Pitstop 2, which inhibits clathrin-dependent endocytosis, markedly enhanced the internalization of the EVs in our system (Figure 7). Although the exact mechanism for this enhancement is unknown, it is possible that inhibition of clathrin-dependent endocytosis by Pitstop 2 led to the activation of other internalization pathways through the mechanism known as cross-regulation [60]. A similar mechanism was speculated in a previous study where treatment of the recipient cells with chlorpromazine, which inhibits clathrin-dependent endocytosis, increased the EV uptake [61].
In conclusion, this study demonstrates that the cells harboring the flavivirus-derived subgenomic replicon, which can avoid contamination of infectious virus particles, serve as a useful EV donor. By using this system, we successfully provided evidence that the RNA molecules derived from JEV and DENV genomes can be transferred from cell to cell in an EV-mediated manner. It was also indicated that the cellular proteins, such as CD33, Tim-1, and Tim-4, are likely involved in the attachment of the EVs on the recipient cell surface. Furthermore, macropinocytosis was shown to play a major role in the EV uptake by the recipient cells in our system. In addition to the classical receptor-mediated infection, EV-mediated cell-to-cell transmission of the flavivirus genome, which could escape the effects of neutralizing antibodies, may play some important roles in viral propagation and pathogenesis. Further studies to elucidate the significance of the EV-mediated transfer of flaviviral components are necessary.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/v16040524/s1, Figure S1: Effects of GW4869 on the replication of JM-PnL and DN-PnL in BHK cells.
Author Contributions
Conceptualization, T.I.; Methodology, T.I.; Investigation, T.I., K.N., and K.M.; Resources, T.I. and M.M.; Writing—Original Draft Preparation, T.I. and K.N.; Writing—Review and Editing, T.I. and M.M.; Supervision, M.M.; Project Administration, T.I.; Funding Acquisition, T.I. All authors have read and agreed to the published version of the manuscript.
Funding
This study was supported by the AMED under grant numbers JP20fk0108123 and JP23fk0108656 as well as JSPS KAKENHI under grant number JP21K08512.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data presented in this study are available on request.
Acknowledgments
We thank Takashi Namatame (Center for Research Collaboration and Support, Dokkyo Medical University) for assistance with DNA sequencing and Yuki Shinozaki and Satomi Yoshida (Department of Microbiology, Dokkyo Medical University School of Medicine) for technical and secretarial assistance.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Pierson, T.C.; Diamond, M.S. Flaviviruses. In Fields Virology, 6th ed.; Knipe, D.M., Howley, P.M., Eds.; Lippincott Williams & Wilkins: Philadelphia, PA, USA, 2013; pp. 747–794. [Google Scholar]
- Pierson, T.C.; Diamond, M.S. The continued threat of emerging flaviviruses. Nat. Microbiol. 2020, 5, 796–812. [Google Scholar] [CrossRef] [PubMed]
- Ishikawa, T.; Yamanaka, A.; Konishi, E. A review of successful flavivirus vaccines and the problems with those flaviviruses for which vaccines are not yet available. Vaccine 2014, 32, 1326–1337. [Google Scholar] [CrossRef] [PubMed]
- Lindenbach, B.D.; Murray, C.; Thiel, H.J.; Rice, C.M. Flaviviridae. In Fields Virology, 6th ed.; Knipe, D.M., Howley, P.M., Eds.; Lippincott Williams & Wilkins: Philadelphia, PA, USA, 2013; pp. 712–746. [Google Scholar]
- Latanova, A.; Starodubova, E.; Karpov, V. Flaviviridae Nonstructural Proteins: The Role in Molecular Mechanisms of Triggering Inflammation. Viruses 2022, 14, 1808. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.; El Andaloussi, S.; Wood, M.J. Exosomes and microvesicles: Extracellular vesicles for genetic information transfer and gene therapy. Hum. Mol. Genet. 2012, 21, R125–R134. [Google Scholar] [CrossRef] [PubMed]
- Kalluri, R.; LeBleu, V.S. The biology, function, and biomedical applications of exosomes. Science 2020, 367, eaau6977. [Google Scholar] [CrossRef] [PubMed]
- Becker, A.; Thakur, B.K.; Weiss, J.M.; Kim, H.S.; Peinado, H.; Lyden, D. Extracellular Vesicles in Cancer: Cell-to-Cell Mediators of Metastasis. Cancer Cell 2016, 30, 836–848. [Google Scholar] [CrossRef] [PubMed]
- Wortzel, I.; Dror, S.; Kenific, C.M.; Lyden, D. Exosome-Mediated Metastasis: Communication from a Distance. Dev. Cell 2019, 49, 347–360. [Google Scholar] [CrossRef] [PubMed]
- Saad, M.H.; Badierah, R.; Redwan, E.M.; El-Fakharany, E.M. A Comprehensive Insight into the Role of Exosomes in Viral Infection: Dual Faces Bearing Different Functions. Pharmaceutics 2021, 13, 1405. [Google Scholar] [CrossRef] [PubMed]
- Fang, Y.; Wu, N.; Gan, X.; Yan, W.; Morrell, J.C.; Gould, S.J. Higher-order oligomerization targets plasma membrane proteins and HIV gag to exosomes. PLoS Biol. 2007, 5, e158. [Google Scholar] [CrossRef]
- Jaworski, E.; Narayanan, A.; Van Duyne, R.; Shabbeer-Meyering, S.; Iordanskiy, S.; Saifuddin, M.; Das, R.; Afonso, P.V.; Sampey, G.C.; Chung, M.; et al. Human T-lymphotropic virus type 1-infected cells secrete exosomes that contain Tax protein. J. Biol. Chem. 2014, 289, 22284–22305. [Google Scholar] [CrossRef]
- Ramakrishnaiah, V.; Thumann, C.; Fofana, I.; Habersetzer, F.; Pan, Q.; de Ruiter, P.E.; Willemsen, R.; Demmers, J.A.; Stalin Raj, V.; Jenster, G.; et al. Exosome-mediated transmission of hepatitis C virus between human hepatoma Huh7.5 cells. Proc. Natl. Acad. Sci. USA 2013, 110, 13109–13113. [Google Scholar] [CrossRef] [PubMed]
- Chivero, E.T.; Bhattarai, N.; Rydze, R.T.; Winters, M.A.; Holodniy, M.; Stapleton, J.T. Human pegivirus RNA is found in multiple blood mononuclear cells in vivo and serum-derived viral RNA-containing particles are infectious in vitro. J. Gen. Virol. 2014, 95, 1307–1319. [Google Scholar] [CrossRef] [PubMed]
- Kadiu, I.; Narayanasamy, P.; Dash, P.K.; Zhang, W.; Gendelman, H.E. Biochemical and biologic characterization of exosomes and microvesicles as facilitators of HIV-1 infection in macrophages. J. Immunol. 2012, 189, 744–754. [Google Scholar] [CrossRef]
- Feng, Z.; Hensley, L.; McKnight, K.L.; Hu, F.; Madden, V.; Ping, L.; Jeong, S.H.; Walker, C.; Lanford, R.E.; Lemon, S.M. A pathogenic picornavirus acquires an envelope by hijacking cellular membranes. Nature 2013, 496, 367–371. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.H.; Du, W.; Hagemeijer, M.C.; Takvorian, P.M.; Pau, C.; Cali, A.; Brantner, C.A.; Stempinski, E.S.; Connelly, P.S.; Ma, H.C.; et al. Phosphatidylserine vesicles enable efficient en bloc transmission of enteroviruses. Cell 2015, 160, 619–630. [Google Scholar] [CrossRef] [PubMed]
- Moulin, C.; Crupi, M.J.F.; Ilkow, C.S.; Bell, J.C.; Boulton, S. Extracellular Vesicles and Viruses: Two Intertwined Entities. Int. J. Mol. Sci. 2023, 24, 1306. [Google Scholar] [CrossRef] [PubMed]
- Zhou, W.; Woodson, M.; Neupane, B.; Bai, F.; Sherman, M.B.; Choi, K.H.; Neelakanta, G.; Sultana, H. Exosomes serve as novel modes of tick-borne flavivirus transmission from arthropod to human cells and facilitates dissemination of viral RNA and proteins to the vertebrate neuronal cells. PLoS Pathog. 2018, 14, e1006764. [Google Scholar] [CrossRef] [PubMed]
- Vora, A.; Zhou, W.; Londono-Renteria, B.; Woodson, M.; Sherman, M.B.; Colpitts, T.M.; Neelakanta, G.; Sultana, H. Arthropod EVs mediate dengue virus transmission through interaction with a tetraspanin domain containing glycoprotein Tsp29Fb. Proc. Natl. Acad. Sci. USA 2018, 115, E6604–E6613. [Google Scholar] [CrossRef] [PubMed]
- Zhou, W.; Woodson, M.; Sherman, M.B.; Neelakanta, G.; Sultana, H. Exosomes mediate Zika virus transmission through SMPD3 neutral Sphingomyelinase in cortical neurons. Emerg. Microbes Infect. 2019, 8, 307–326. [Google Scholar] [CrossRef] [PubMed]
- Ishikawa, T.; Abe, M.; Masuda, M. Construction of an infectious molecular clone of Japanese encephalitis virus genotype V and its derivative subgenomic replicon capable of expressing a foreign gene. Virus Res. 2015, 195, 153–161. [Google Scholar] [CrossRef]
- Honjo, S.; Masuda, M.; Ishikawa, T. Effects of the Japanese Encephalitis Virus Genotype V-Derived Sub-Viral Particles on the Immunogenicity of the Vaccine Characterized by a Novel Virus-Like Particle-Based Assay. Vaccines 2019, 7, 81. [Google Scholar] [CrossRef] [PubMed]
- Kitai, Y.; Shoda, M.; Kondo, T.; Konishi, E. Epitope-blocking enzyme-linked immunosorbent assay to differentiate west nile virus from Japanese encephalitis virus infections in equine sera. Clin. Vaccine Immunol. 2007, 14, 1024–1031. [Google Scholar] [CrossRef] [PubMed]
- Malenica, M.; Vukomanovic, M.; Kurtjak, M.; Masciotti, V.; Dal Zilio, S.; Greco, S.; Lazzarino, M.; Krusic, V.; Percic, M.; Jelovica Badovinac, I.; et al. Perspectives of Microscopy Methods for Morphology Characterisation of Extracellular Vesicles from Human Biofluids. Biomedicines 2021, 9, 603. [Google Scholar] [CrossRef] [PubMed]
- Gonda, A.; Kabagwira, J.; Senthil, G.N.; Wall, N.R. Internalization of Exosomes through Receptor-Mediated Endocytosis. Mol. Cancer Res. 2019, 17, 337–347. [Google Scholar] [CrossRef] [PubMed]
- Zitvogel, L.; Regnault, A.; Lozier, A.; Wolfers, J.; Flament, C.; Tenza, D.; Ricciardi-Castagnoli, P.; Raposo, G.; Amigorena, S. Eradication of established murine tumors using a novel cell-free vaccine: Dendritic cell-derived exosomes. Nat. Med. 1998, 4, 594–600. [Google Scholar] [CrossRef] [PubMed]
- Kitai, Y.; Kawasaki, T.; Sueyoshi, T.; Kobiyama, K.; Ishii, K.J.; Zou, J.; Akira, S.; Matsuda, T.; Kawai, T. DNA-Containing Exosomes Derived from Cancer Cells Treated with Topotecan Activate a STING-Dependent Pathway and Reinforce Antitumor Immunity. J. Immunol. 2017, 198, 1649–1659. [Google Scholar] [CrossRef] [PubMed]
- De Wever, O.; Demetter, P.; Mareel, M.; Bracke, M. Stromal myofibroblasts are drivers of invasive cancer growth. Int. J. Cancer 2008, 123, 2229–2238. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Deng, T.; Liu, R.; Bai, M.; Zhou, L.; Wang, X.; Li, S.; Wang, X.; Yang, H.; Li, J.; et al. Exosome-delivered EGFR regulates liver microenvironment to promote gastric cancer liver metastasis. Nat. Commun. 2017, 8, 15016. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Zhang, S.; Yao, J.; Lowery, F.J.; Zhang, Q.; Huang, W.C.; Li, P.; Li, M.; Wang, X.; Zhang, C.; et al. Microenvironment-induced PTEN loss by exosomal microRNA primes brain metastasis outgrowth. Nature 2015, 527, 100–104. [Google Scholar] [CrossRef]
- Cooks, T.; Pateras, I.S.; Jenkins, L.M.; Patel, K.M.; Robles, A.I.; Morris, J.; Forshew, T.; Appella, E.; Gorgoulis, V.G.; Harris, C.C. Mutant p53 cancers reprogram macrophages to tumor supporting macrophages via exosomal miR-1246. Nat. Commun. 2018, 9, 771. [Google Scholar] [CrossRef]
- Temme, S.; Eis-Hubinger, A.M.; McLellan, A.D.; Koch, N. The herpes simplex virus-1 encoded glycoprotein B diverts HLA-DR into the exosome pathway. J. Immunol. 2010, 184, 236–243. [Google Scholar] [CrossRef]
- Vallhov, H.; Gutzeit, C.; Johansson, S.M.; Nagy, N.; Paul, M.; Li, Q.; Friend, S.; George, T.C.; Klein, E.; Scheynius, A.; et al. Exosomes containing glycoprotein 350 released by EBV-transformed B cells selectively target B cells through CD21 and block EBV infection in vitro. J. Immunol. 2011, 186, 73–82. [Google Scholar] [CrossRef] [PubMed]
- Campbell, T.D.; Khan, M.; Huang, M.B.; Bond, V.C.; Powell, M.D. HIV-1 Nef protein is secreted into vesicles that can fuse with target cells and virions. Ethn. Dis. 2008, 18, S2-14-9. [Google Scholar] [PubMed]
- Pinto, D.O.; DeMarino, C.; Pleet, M.L.; Cowen, M.; Branscome, H.; Al Sharif, S.; Jones, J.; Dutartre, H.; Lepene, B.; Liotta, L.A.; et al. HTLV-1 Extracellular Vesicles Promote Cell-to-Cell Contact. Front. Microbiol. 2019, 10, 2147. [Google Scholar] [CrossRef] [PubMed]
- Kalamvoki, M.; Du, T.; Roizman, B. Cells infected with herpes simplex virus 1 export to uninfected cells exosomes containing STING, viral mRNAs, and microRNAs. Proc. Natl. Acad. Sci. USA 2014, 111, E4991–E4996. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, W.; Philip, P.S.; Tariq, S.; Khan, G. Epstein-Barr virus-encoded small RNAs (EBERs) are present in fractions related to exosomes released by EBV-transformed cells. PLoS ONE 2014, 9, e99163. [Google Scholar] [CrossRef]
- Bukong, T.N.; Momen-Heravi, F.; Kodys, K.; Bala, S.; Szabo, G. Exosomes from hepatitis C infected patients transmit HCV infection and contain replication competent viral RNA in complex with Ago2-miR122-HSP90. PLoS Pathog. 2014, 10, e1004424. [Google Scholar] [CrossRef] [PubMed]
- Honegger, A.; Schilling, D.; Bastian, S.; Sponagel, J.; Kuryshev, V.; Sultmann, H.; Scheffner, M.; Hoppe-Seyler, K.; Hoppe-Seyler, F. Dependence of intracellular and exosomal microRNAs on viral E6/E7 oncogene expression in HPV-positive tumor cells. PLoS Pathog. 2015, 11, e1004712. [Google Scholar] [CrossRef]
- Sampey, G.C.; Saifuddin, M.; Schwab, A.; Barclay, R.; Punya, S.; Chung, M.C.; Hakami, R.M.; Zadeh, M.A.; Lepene, B.; Klase, Z.A.; et al. Exosomes from HIV-1-infected Cells Stimulate Production of Pro-inflammatory Cytokines through Trans-activating Response (TAR) RNA. J. Biol. Chem. 2016, 291, 1251–1266. [Google Scholar] [CrossRef]
- Hoshina, S.; Sekizuka, T.; Kataoka, M.; Hasegawa, H.; Hamada, H.; Kuroda, M.; Katano, H. Profile of Exosomal and Intracellular microRNA in Gamma-Herpesvirus-Infected Lymphoma Cell Lines. PLoS ONE 2016, 11, e0162574. [Google Scholar] [CrossRef]
- Fabbiano, F.; Corsi, J.; Gurrieri, E.; Trevisan, C.; Notarangelo, M.; D’Agostino, V.G. RNA packaging into extracellular vesicles: An orchestra of RNA-binding proteins? J. Extracell. Vesicles 2020, 10, e12043. [Google Scholar] [CrossRef]
- Katoh, H.; Mori, Y.; Kambara, H.; Abe, T.; Fukuhara, T.; Morita, E.; Moriishi, K.; Kamitani, W.; Matsuura, Y. Heterogeneous nuclear ribonucleoprotein A2 participates in the replication of Japanese encephalitis virus through an interaction with viral proteins and RNA. J. Virol. 2011, 85, 10976–10988. [Google Scholar] [CrossRef] [PubMed]
- Phillips, S.L.; Soderblom, E.J.; Bradrick, S.S.; Garcia-Blanco, M.A. Identification of Proteins Bound to Dengue Viral RNA In Vivo Reveals New Host Proteins Important for Virus Replication. mBio 2016, 7, e01865-15. [Google Scholar] [CrossRef] [PubMed]
- Viktorovskaya, O.V.; Greco, T.M.; Cristea, I.M.; Thompson, S.R. Identification of RNA Binding Proteins Associated with Dengue Virus RNA in Infected Cells Reveals Temporally Distinct Host Factor Requirements. PLoS Negl. Trop. Dis. 2016, 10, e0004921. [Google Scholar] [CrossRef] [PubMed]
- Ageta, H.; Ageta-Ishihara, N.; Hitachi, K.; Karayel, O.; Onouchi, T.; Yamaguchi, H.; Kahyo, T.; Hatanaka, K.; Ikegami, K.; Yoshioka, Y.; et al. UBL3 modification influences protein sorting to small extracellular vesicles. Nat. Commun. 2018, 9, 3936. [Google Scholar] [CrossRef] [PubMed]
- Richard, P.; Feng, S.; Manley, J.L. A SUMO-dependent interaction between Senataxin and the exosome, disrupted in the neurodegenerative disease AOA2, targets the exosome to sites of transcription-induced DNA damage. Genes Dev. 2013, 27, 2227–2232. [Google Scholar] [CrossRef] [PubMed]
- Villarroya-Beltri, C.; Gutierrez-Vazquez, C.; Sanchez-Cabo, F.; Perez-Hernandez, D.; Vazquez, J.; Martin-Cofreces, N.; Martinez-Herrera, D.J.; Pascual-Montano, A.; Mittelbrunn, M.; Sanchez-Madrid, F. Sumoylated hnRNPA2B1 controls the sorting of miRNAs into exosomes through binding to specific motifs. Nat. Commun. 2013, 4, 2980. [Google Scholar] [CrossRef] [PubMed]
- Buschow, S.I.; Liefhebber, J.M.; Wubbolts, R.; Stoorvogel, W. Exosomes contain ubiquitinated proteins. Blood Cells Mol. Dis. 2005, 35, 398–403. [Google Scholar] [CrossRef]
- Shimoda, A.; Tahara, Y.; Sawada, S.I.; Sasaki, Y.; Akiyoshi, K. Glycan profiling analysis using evanescent-field fluorescence-assisted lectin array: Importance of sugar recognition for cellular uptake of exosomes from mesenchymal stem cells. Biochem. Biophys. Res. Commun. 2017, 491, 701–707. [Google Scholar] [CrossRef]
- Kobayashi, N.; Karisola, P.; Pena-Cruz, V.; Dorfman, D.M.; Jinushi, M.; Umetsu, S.E.; Butte, M.J.; Nagumo, H.; Chernova, I.; Zhu, B.; et al. TIM-1 and TIM-4 glycoproteins bind phosphatidylserine and mediate uptake of apoptotic cells. Immunity 2007, 27, 927–940. [Google Scholar] [CrossRef]
- Kondratowicz, A.S.; Lennemann, N.J.; Sinn, P.L.; Davey, R.A.; Hunt, C.L.; Moller-Tank, S.; Meyerholz, D.K.; Rennert, P.; Mullins, R.F.; Brindley, M.; et al. T-cell immunoglobulin and mucin domain 1 (TIM-1) is a receptor for Zaire Ebolavirus and Lake Victoria Marburgvirus. Proc. Natl. Acad. Sci. USA 2011, 108, 8426–8431. [Google Scholar] [CrossRef]
- McKelvey, K.J.; Powell, K.L.; Ashton, A.W.; Morris, J.M.; McCracken, S.A. Exosomes: Mechanisms of Uptake. J. Circ. Biomark. 2015, 4, 7. [Google Scholar] [CrossRef] [PubMed]
- Gurung, S.; Perocheau, D.; Touramanidou, L.; Baruteau, J. The exosome journey: From biogenesis to uptake and intracellular signalling. Cell Commun. Signal 2021, 19, 47. [Google Scholar] [CrossRef]
- Mulcahy, L.A.; Pink, R.C.; Carter, D.R. Routes and mechanisms of extracellular vesicle uptake. J. Extracell. Vesicles 2014, 3, 24641. [Google Scholar] [CrossRef] [PubMed]
- Lim, J.P.; Gleeson, P.A. Macropinocytosis: An endocytic pathway for internalising large gulps. Immunol. Cell Biol. 2011, 89, 836–843. [Google Scholar] [CrossRef] [PubMed]
- Reyes-Ruiz, J.M.; Osuna-Ramos, J.F.; De Jesus-Gonzalez, L.A.; Hurtado-Monzon, A.M.; Farfan-Morales, C.N.; Cervantes-Salazar, M.; Bolanos, J.; Cigarroa-Mayorga, O.E.; Martin-Martinez, E.S.; Medina, F.; et al. Isolation and characterization of exosomes released from mosquito cells infected with dengue virus. Virus Res. 2019, 266, 1–14. [Google Scholar] [CrossRef]
- Martinez-Rojas, P.P.; Quiroz-Garcia, E.; Monroy-Martinez, V.; Agredano-Moreno, L.T.; Jimenez-Garcia, L.F.; Ruiz-Ordaz, B.H. Participation of Extracellular Vesicles from Zika-Virus-Infected Mosquito Cells in the Modification of Naive Cells’ Behavior by Mediating Cell-to-Cell Transmission of Viral Elements. Cells 2020, 9, 123. [Google Scholar] [CrossRef]
- Mayor, S.; Pagano, R.E. Pathways of clathrin-independent endocytosis. Nat. Rev. Mol. Cell Biol. 2007, 8, 603–612. [Google Scholar] [CrossRef]
- Verdera, C.H.; Gitz-Francois, J.J.; Schiffelers, R.M.; Vader, P. Cellular uptake of extracellular vesicles is mediated by clathrin-independent endocytosis and macropinocytosis. J. Control. Release 2017, 266, 100–108. [Google Scholar] [CrossRef]
Figure 1.
Establishment of JEV- and DENV-subgenomic replicon harboring cells. (A) Structures of the flavivirus genome, JM-PnL, and DN-PnL were shown. UTR: untranslated region, RZ: hepatitis delta virus derived ribozyme. (B) Immunostaining of BHK/JM-PnL cells. Cells were stained with anti-NS1 antibody. (C) Luciferase activities of BHK/JM-PnL cells (n = 2). (D) Immunostaining of BHK/DN-PnL cells. Cells were stained with anti-NS1 antibody (n = 2). (E) Luciferase activities of BHK/DN-PnL cells.
Figure 1.
Establishment of JEV- and DENV-subgenomic replicon harboring cells. (A) Structures of the flavivirus genome, JM-PnL, and DN-PnL were shown. UTR: untranslated region, RZ: hepatitis delta virus derived ribozyme. (B) Immunostaining of BHK/JM-PnL cells. Cells were stained with anti-NS1 antibody. (C) Luciferase activities of BHK/JM-PnL cells (n = 2). (D) Immunostaining of BHK/DN-PnL cells. Cells were stained with anti-NS1 antibody (n = 2). (E) Luciferase activities of BHK/DN-PnL cells.
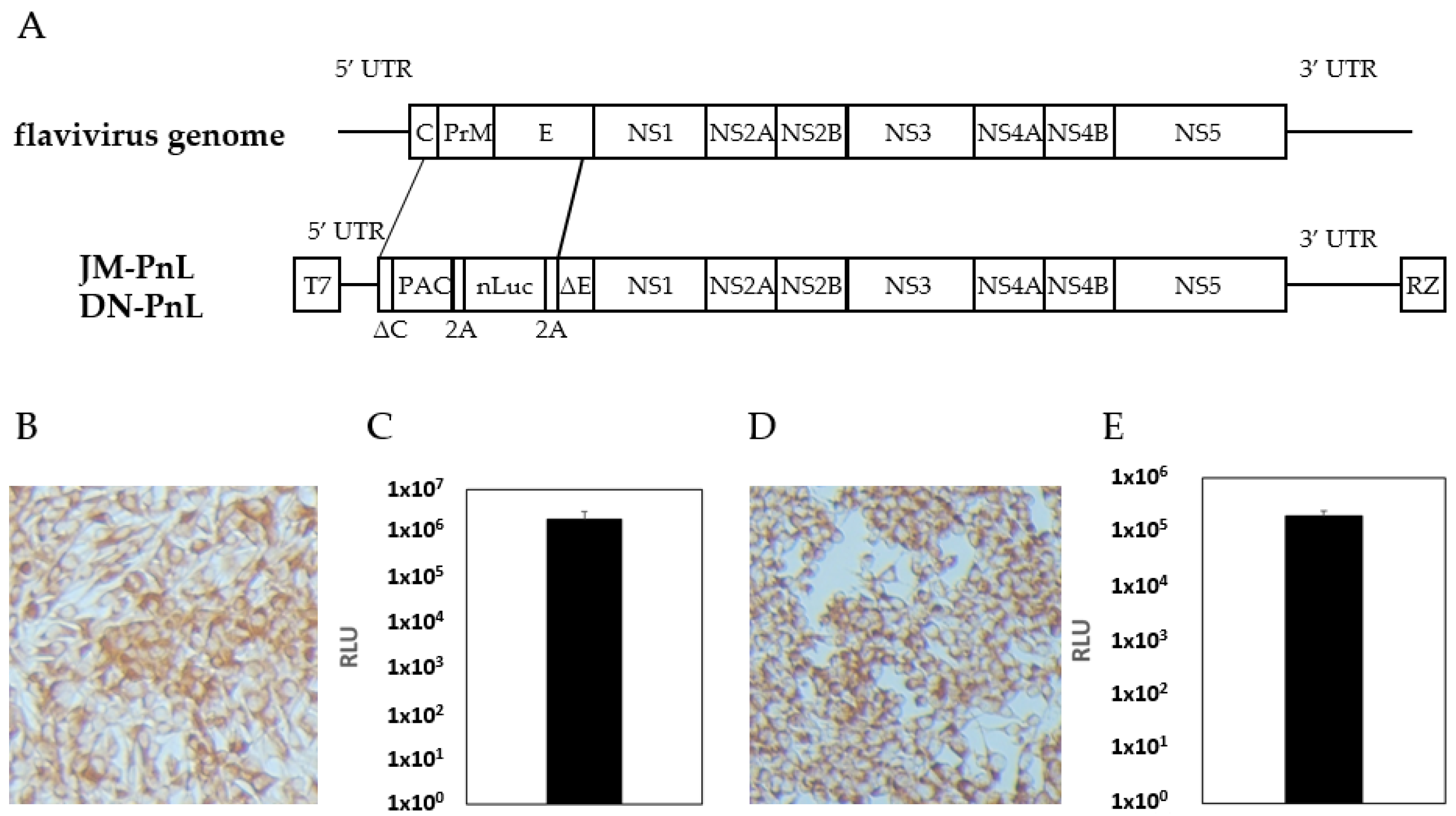
Figure 2.
Detection of subgenomic replicon RNA within EVs. (A) Morphological analysis of EVs derived from BHK/JM-PnL and BHK/DN-PnL cells. Arrow indicates the typical image of EVs. Bar represents 50 nm. (B) Total RNAs were isolated from EVs derived from BHK/JM-PnL and BHK/DN-PnL cells. Luciferase-JEV NS1 (889 bp), JEV NS5 (464 bp), Luciferase-DENV NS1 (557 bp), and DENV NS5 (718 bp) fragments were amplified. The bands surrounded by red boxes corresponded to the expected size.
Figure 2.
Detection of subgenomic replicon RNA within EVs. (A) Morphological analysis of EVs derived from BHK/JM-PnL and BHK/DN-PnL cells. Arrow indicates the typical image of EVs. Bar represents 50 nm. (B) Total RNAs were isolated from EVs derived from BHK/JM-PnL and BHK/DN-PnL cells. Luciferase-JEV NS1 (889 bp), JEV NS5 (464 bp), Luciferase-DENV NS1 (557 bp), and DENV NS5 (718 bp) fragments were amplified. The bands surrounded by red boxes corresponded to the expected size.
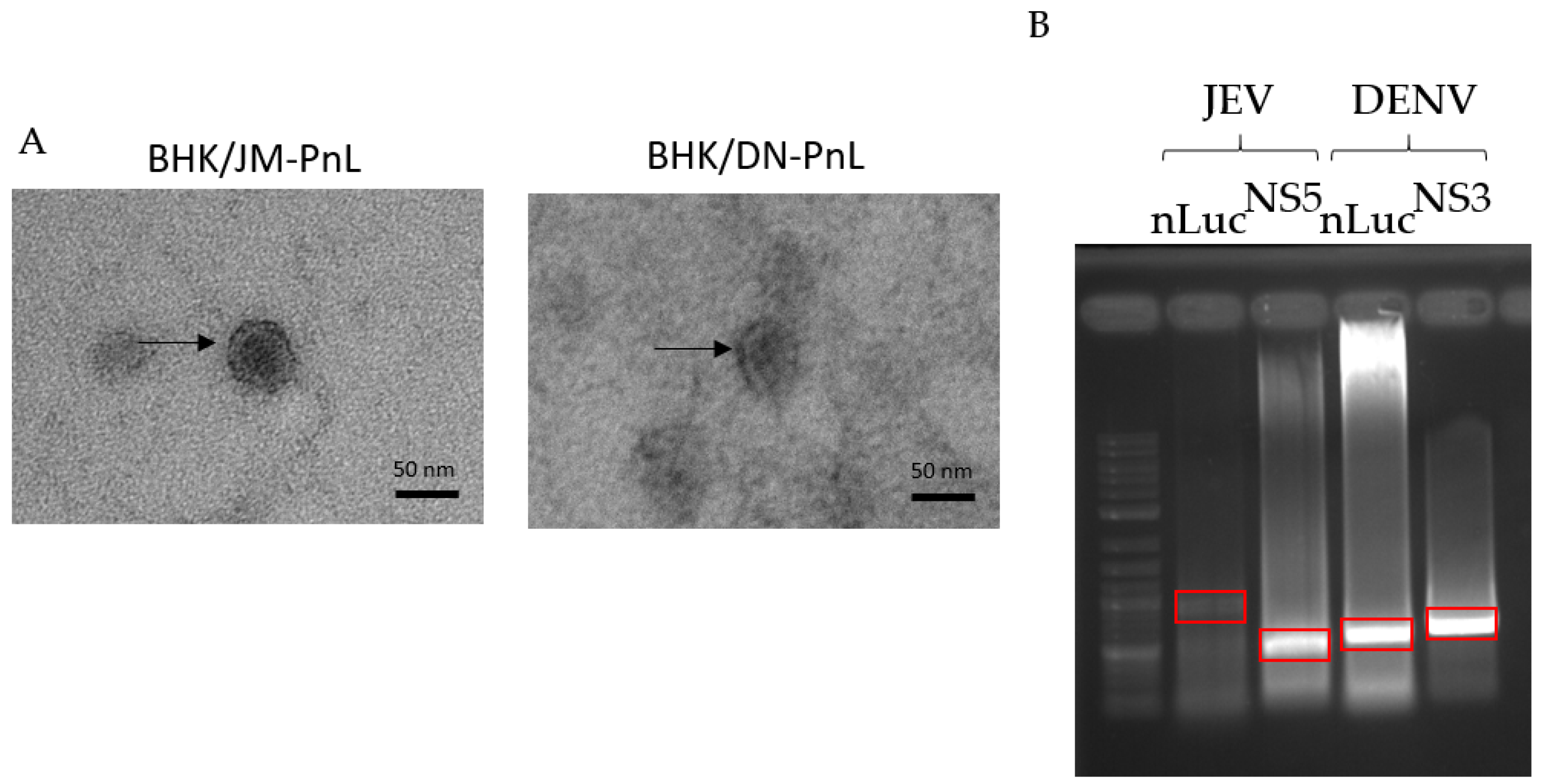
Figure 3.
Existence of replicon RNA and viral proteins in EVs. (A) JM-PnL and DN-PnL replicon genomes contained in EVs treated with RNase A and/or Tx-100 were quantified by real-time PCR (n = 2). (B) BHK/JM-PnL cell- and BHK/DN-PnL cell-derived EVs treated with RNase A were inoculated onto BHK cells. Data were expressed as relative luminescence units to the negative control (n = 3). (C) Western blot analysis of isolated EVs derived from BHK/JM-PnL cells and BHK/DN-PnL cells (lane 1). JEV Muar strain and DENV2 NGC strain-infected BHK cells were used as a positive control (lane 2) and naïve BHK cells were used as a negative control (lane 3). The presence of the JEV NS1, DENV NS1, and DENV NS2B-3 in the culture supernatants derived from BHK/JM-PnL or BHK/DN-PnL cells (lane 4), BHK cells (lane 5), and EV-depleted culture supernatants (lane 6) were examined. The brightness of each picture was adjusted upon manuscript preparation. (D) Western blot analysis of isolated EVs treated with proteinase K in the presence/absence of Tx-100.
Figure 3.
Existence of replicon RNA and viral proteins in EVs. (A) JM-PnL and DN-PnL replicon genomes contained in EVs treated with RNase A and/or Tx-100 were quantified by real-time PCR (n = 2). (B) BHK/JM-PnL cell- and BHK/DN-PnL cell-derived EVs treated with RNase A were inoculated onto BHK cells. Data were expressed as relative luminescence units to the negative control (n = 3). (C) Western blot analysis of isolated EVs derived from BHK/JM-PnL cells and BHK/DN-PnL cells (lane 1). JEV Muar strain and DENV2 NGC strain-infected BHK cells were used as a positive control (lane 2) and naïve BHK cells were used as a negative control (lane 3). The presence of the JEV NS1, DENV NS1, and DENV NS2B-3 in the culture supernatants derived from BHK/JM-PnL or BHK/DN-PnL cells (lane 4), BHK cells (lane 5), and EV-depleted culture supernatants (lane 6) were examined. The brightness of each picture was adjusted upon manuscript preparation. (D) Western blot analysis of isolated EVs treated with proteinase K in the presence/absence of Tx-100.
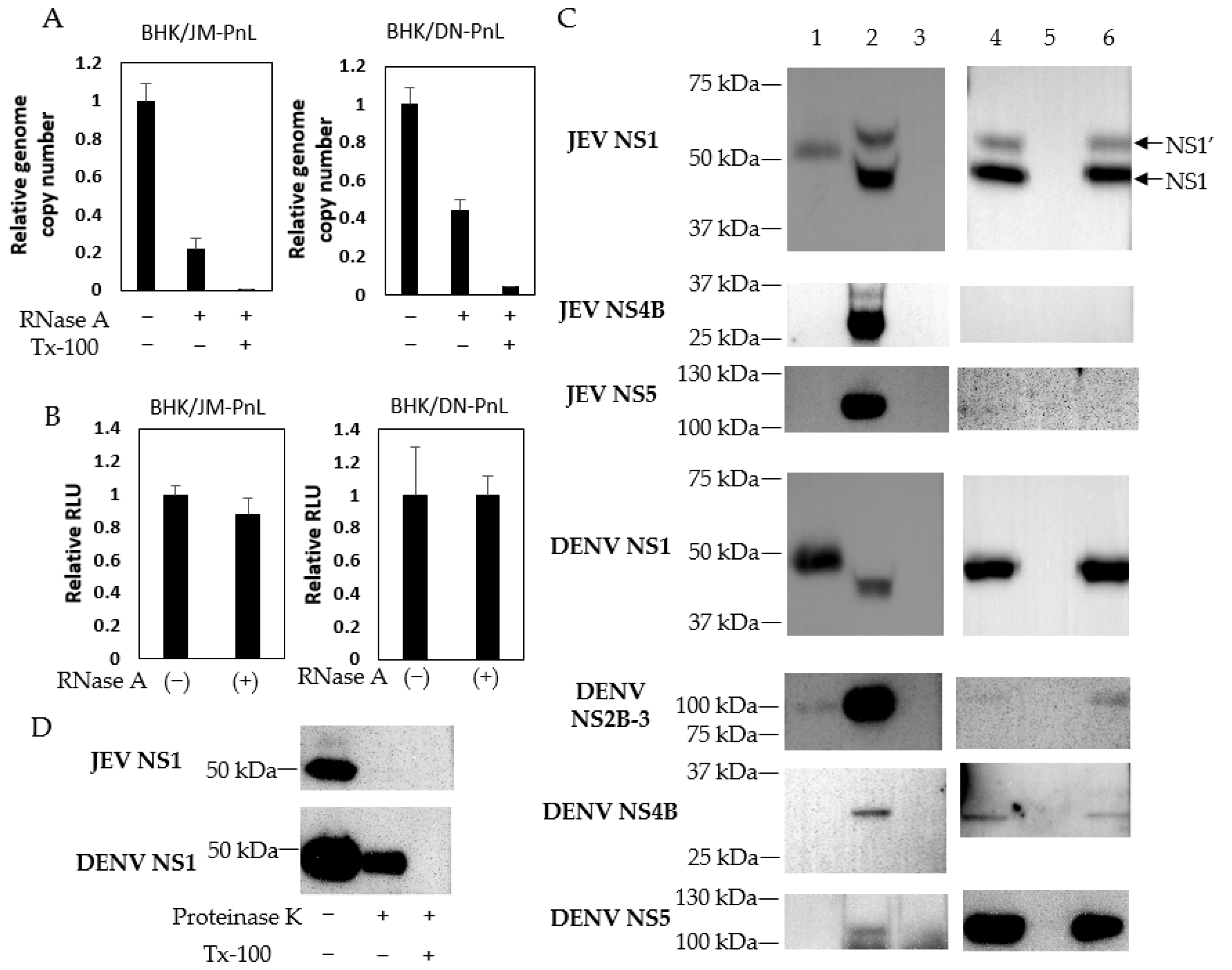
Figure 4.
Transfer of subgenomic replicon genomes to various cell lines by EVs. (A) Luciferase activities of various cells inoculated with EVs derived from BHK/JM-PnL and BHK/DN-PnL cells were measured at 72 h. Luciferase activities obtained from cells inoculated with naïve BHK cell-derived EVs were used as a negative control and subtracted from each luciferase activity (n = 3). (B) Culture supernatants before EV isolation (Pre) and EV-depleted culture supernatants (Post) from BHK/JM-PnL and BHK/DNPnL cells were inoculated on BHK cells (n = 3). (C)Luciferase activities of the BHK cells inoculated with EVs in the presence/absence of ribavirin were measured (n = 3). (D) The JM-PnL and DN-PnL replicon genomes in the EV-inoculated C6/36 cells were quantified by real-time PCR (n = 3). * p < 0.05, ** p < 0.01.
Figure 4.
Transfer of subgenomic replicon genomes to various cell lines by EVs. (A) Luciferase activities of various cells inoculated with EVs derived from BHK/JM-PnL and BHK/DN-PnL cells were measured at 72 h. Luciferase activities obtained from cells inoculated with naïve BHK cell-derived EVs were used as a negative control and subtracted from each luciferase activity (n = 3). (B) Culture supernatants before EV isolation (Pre) and EV-depleted culture supernatants (Post) from BHK/JM-PnL and BHK/DNPnL cells were inoculated on BHK cells (n = 3). (C)Luciferase activities of the BHK cells inoculated with EVs in the presence/absence of ribavirin were measured (n = 3). (D) The JM-PnL and DN-PnL replicon genomes in the EV-inoculated C6/36 cells were quantified by real-time PCR (n = 3). * p < 0.05, ** p < 0.01.

Figure 5.
Cell-to-cell transfer of subgenomic replicon RNA was mediated by EVs. (A) EVs obtained from BHK/JM-PnL and BHK/DN-PnL cells treated with GW4869 were inoculated to BHK cells (n = 3). (B) BHK/JM-PnL cell- and BHK/DN-PnL cell-derived EVs treated with Tx-100 were inoculated onto BHK cells (n = 3). Data were expressed as relative luminescence units to negative control. * p < 0.05, ** p < 0.01.
Figure 5.
Cell-to-cell transfer of subgenomic replicon RNA was mediated by EVs. (A) EVs obtained from BHK/JM-PnL and BHK/DN-PnL cells treated with GW4869 were inoculated to BHK cells (n = 3). (B) BHK/JM-PnL cell- and BHK/DN-PnL cell-derived EVs treated with Tx-100 were inoculated onto BHK cells (n = 3). Data were expressed as relative luminescence units to negative control. * p < 0.05, ** p < 0.01.
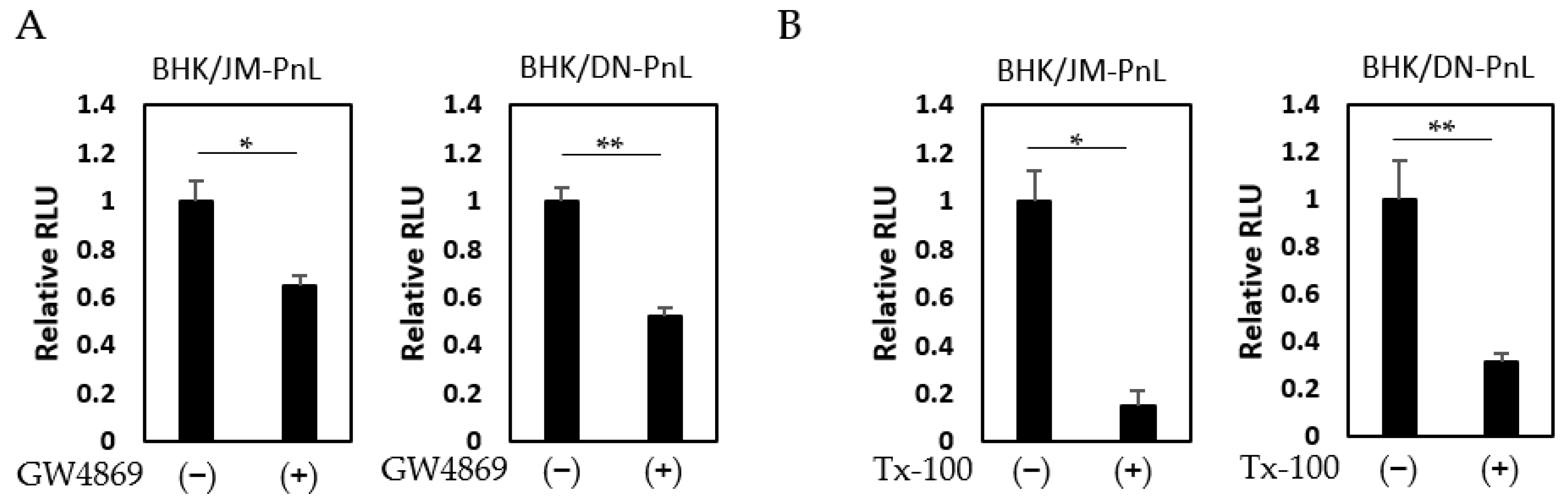
Figure 6.
Interaction of EV with cell surface molecules. (A) HeLa cells were treated with anti-CD33, anti-Tim-1, and anti-Tim-4 antibodies and then inoculated with EVs derived from BHK/JM-PnL or BHK/DN-PnL cells. (B) K562 cells were treated with anti-CD33, anti-Tim-1, and anti-Tim-4 antibodies and then inoculated with EVs derived from BHK/DN-PnL cells. Data were expressed as relative luminescence units to negative control treated with H2O (n = 3). * p < 0.05, ** p < 0.01.
Figure 6.
Interaction of EV with cell surface molecules. (A) HeLa cells were treated with anti-CD33, anti-Tim-1, and anti-Tim-4 antibodies and then inoculated with EVs derived from BHK/JM-PnL or BHK/DN-PnL cells. (B) K562 cells were treated with anti-CD33, anti-Tim-1, and anti-Tim-4 antibodies and then inoculated with EVs derived from BHK/DN-PnL cells. Data were expressed as relative luminescence units to negative control treated with H2O (n = 3). * p < 0.05, ** p < 0.01.
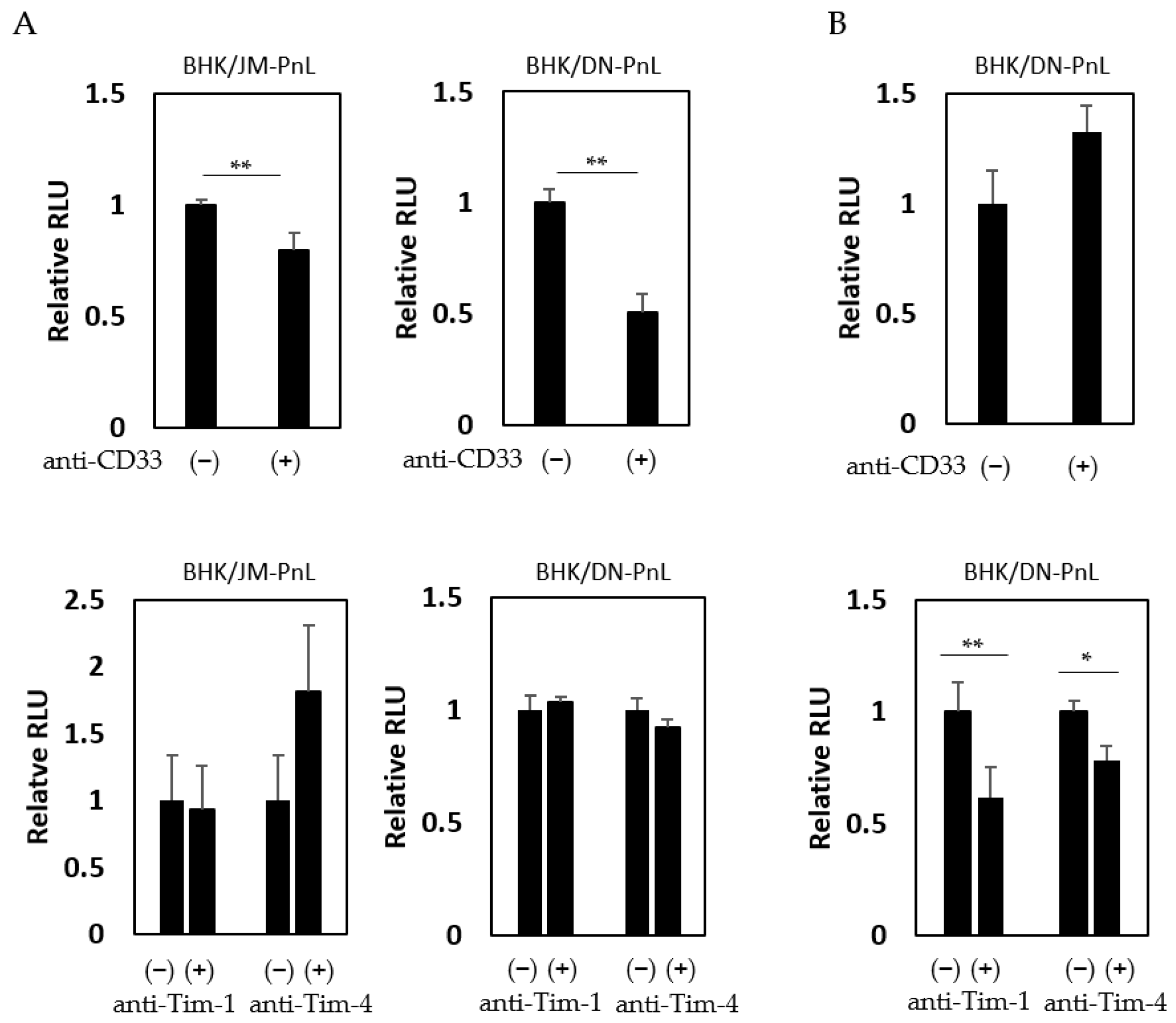
Figure 7.
Effects of various entry inhibitors on EV uptake. (A) HeLa cells were treated with Pitstop 2, LY294002, dynasore, genistein, or EIPA and then inoculated with EVs derived from BHK/JM-PnL and BHK/DN-PnL cells (n = 3). (B) HeLa cells were treated with EIPA and then inoculated with EVs derived from BHK/JM-PnL cells for direct comparison (n = 5). Data were expressed as relative luminescence units to the negative control. * p < 0.05.
Figure 7.
Effects of various entry inhibitors on EV uptake. (A) HeLa cells were treated with Pitstop 2, LY294002, dynasore, genistein, or EIPA and then inoculated with EVs derived from BHK/JM-PnL and BHK/DN-PnL cells (n = 3). (B) HeLa cells were treated with EIPA and then inoculated with EVs derived from BHK/JM-PnL cells for direct comparison (n = 5). Data were expressed as relative luminescence units to the negative control. * p < 0.05.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Ishikawa, T.; Narita, K.; Matsuyama, K.; Masuda, M. Dissemination of the Flavivirus Subgenomic Replicon Genome and Viral Proteins by Extracellular Vesicles. Viruses 2024, 16, 524. https://doi.org/10.3390/v16040524
AMA Style
Ishikawa T, Narita K, Matsuyama K, Masuda M. Dissemination of the Flavivirus Subgenomic Replicon Genome and Viral Proteins by Extracellular Vesicles. Viruses. 2024; 16(4):524. https://doi.org/10.3390/v16040524
Chicago/Turabian StyleIshikawa, Tomohiro, Kentaro Narita, Kinichi Matsuyama, and Michiaki Masuda. 2024. "Dissemination of the Flavivirus Subgenomic Replicon Genome and Viral Proteins by Extracellular Vesicles" Viruses 16, no. 4: 524. https://doi.org/10.3390/v16040524
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.