
Viroids, Satellite RNAs and Prions: Folding of Nucleic Acids and Misfolding of Proteins

1
Institut für Physikalische Biologie, Faculty of Mathematics and Natural Sciences, Heinrich Heine University Düsseldorf, 40204 Düsseldorf, Germany
2
Institute for Neurodegenerative Diseases, Weill Institute for Neurosciences, University of California San Francisco, San Francisco, CA 94158, USA
3
Department of Neurology, Weill Institute for Neurosciences, University of California San Francisco, San Francisco, CA 94158, USA
4
Department of Biochemistry and Biophysics, University of California San Francisco, San Francisco, CA 94158, USA
*
Author to whom correspondence should be addressed.
Viruses 2024, 16(3), 360; https://doi.org/10.3390/v16030360
Submission received: 20 January 2024
/
Revised: 22 February 2024
/
Accepted: 23 February 2024
/
Published: 26 February 2024
(This article belongs to the Special Issue Viroids and Satellites and Their Vector Interactions—This Special Issue Is Dedicated to the Memory of Theodor O. Diener Who Discovered Viroids)
Abstract
:Theodor (“Ted”) Otto Diener (* 28 February 1921 in Zürich, Switzerland; † 28 March 2023 in Beltsville, MD, USA) pioneered research on viroids while working at the Plant Virology Laboratory, Agricultural Research Service, USDA, in Beltsville. He coined the name viroid and defined viroids’ important features like the infectivity of naked single-stranded RNA without protein-coding capacity. During scientific meetings in the 1970s and 1980s, viroids were often discussed at conferences together with other “subviral pathogens”. This term includes what are now called satellite RNAs and prions. Satellite RNAs depend on a helper virus and have linear or, in the case of virusoids, circular RNA genomes. Prions, proteinaceous infectious particles, are the agents of scrapie, kuru and some other diseases. Many satellite RNAs, like viroids, are non-coding and exert their function by thermodynamically or kinetically controlled folding, while prions are solely host-encoded proteins that cause disease by misfolding, aggregation and transmission of their conformations into infectious prion isoforms. In this memorial, we will recall the work of Ted Diener on subviral pathogens.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
‘Dogma’ is generally defined as a principle or set of principles thought to be true. A well-known dogma formulated by Francis Crick in 1958 [1,2] stated that biological information flows from DNA to RNA to proteins. With the discovery of reverse transcription, the first exception became known. Another example of an outdated dogma was that viruses—and phages—were the smallest infectious agents. They possess a nucleic acid genome that encodes a few proteins. It was Theodor (“Ted”) O. Diener who first described a naked RNA, which is 5 to 10-fold smaller than the smallest known viral genome and has no coding capacity [3,4,5,6]. More unexpected was the discovery of proteinaceous particles called prions, which were first discovered in studies of the infectious agent that causes scrapie of sheep [7,8]. Notably, Diener was extremely skeptical about prions initially [9], but he later collaborated with Prusiner on a study that compared the properties of viroids and prions [10]. Of note, satellite RNAs are similar in size, structure, and pathological effects, but they do not autonomously replicate; rather, they need a specific virus, or helper virus, to multiply. Satellite RNAs were discovered at about the same time as viroids [11,12], and discussing their features here helps to clarify the differences among viroids, subviral satellites and prions. Figure 1 shows a timeline of discoveries focused on this article’s topics.
2. Viroids
In the late 1960s, Diener and co-workers described a free RNA that is infectious in plants, which causes potato spindle tuber disease [3,65]. They concluded that the RNA is too small to contain the genetic information necessary for self-replication and that it must rely on host enzymes for its replication. Joseph S. Semancik and Lewis G. Weathers reported similar findings for the causal agent of citrus exocortis disease [66,67,68,69]. For such an unconventional agent, Diener suggested the term viroid [4], which was adopted in 1972. In the 1970s, the purification methods for small RNAs improved substantially, in particular by ultracentrifugation and gel electrophoresis, and later by chromatography [25]. In Germany, Heinz Ludwig Sänger successfully isolated viroids from several viroid diseases in larger quantities [70,71,72]. He convinced the groups of Albrecht K. Kleinschmidt and Günther Klotz, Hans J. Gross, and Detlev Riesner to jointly study viroids. This cooperation of the “German team” [73] resulted in several breakthrough publications demonstrating the following properties: viroid RNA circularity, the molecular weight, secondary structure, and mechanism of structure formation [30,74]. Hans Gross and his group published the complete nucleotide sequence of the potato spindle tuber viroid (PSTVd) and confirmed its rod-shaped secondary structure in 1978 [29,75]. The biophysical studies demonstrated that certain extra-stable structural elements of viroids, named hairpins I and II, are not part of the native, thermodynamically favored structure; rather, they are present as metastable elements in kinetically favored structures during replication and are critical for replication and processing [22,23,76,77,78,79]. Furthermore, the structural studies contributed to easy, fast diagnostic methods [80] and were even used later to find or exclude nucleic acids in prions (see Section 4).
After the structural research, interest turned to the mechanism of viroid replication and pathogenicity. Ted Diener set the pace, and viroid research groups grew all over the world, including those who studied avocado sun blotch viroid (ASBVd) [28], coconut cadang-cadang viroid (CCCVd) [31,32], and others.
Today, viroids are classified into the families Pospiviroidae (named after PSTVd; 39 members) and Avsunviroidae (named after ASBVd; 5 members) “on the basis of their biological, biochemical and structural features. Members of the family Avsunviroidae can form, in the strands of both polarities, hammerhead ribozymes that mediate replication in chloroplasts, in which these viroids accumulate. Members of the family Pospiviroidae lack hammerhead ribozymes but contain a central conserved region (CCR) in their rod-like or quasi rod-like conformation and replicate in the nucleus, wherein these viroids also accumulate” [81].
3. Viroids: The Archetype of Further Subviral Pathogens?
After the sequence of PSTVd and some early results about the replication mechanism were determined [26,27,82,83], viroids were established and accepted as subviral pathogens. The viroid researchers became a worldwide family essentially, meeting in Beltsville [84] and during many symposia (Figure 2). But during those symposia, other unknown pathogens were discussed including the agents of kuru [60,85], called unconventional virus, and scrapie, a sheep disease leading to amyloidosis. Similar to viroids, these particles could not be seen under a light microscope and nobody knew what to look for using an electron microscope since a molecular test did not exist. Could this type of unidentified infectious animal agents be a new class of viroids? Semancik, Hanson and coworkers published two papers in Nature arguing that scrapie had an essential DNA component of small molecular weight [86,87]. Even Ted Diener similarly speculated in a contribution to the 2nd International Congress for Virology, Budapest (1971) [9]. After the groundbreaking publication of Stanley B. Prusiner in Science 1982, in which he coined the term “prion” [88], controversies were plentiful and discussed during joint sessions on subviral pathogens; the last one took place at the 6th International Congress for Virology in Sendai (1984). After that, the scientific directions split, but the earlier often controversial discussions helped clarify the fundamental differences between the plant and animal subviral pathogens.
For young scientists, the research on subviral pathogens remained very attractive, and Ted Diener and Detlev Riesner organized a summer school of the Studienstiftung des Deutschen Volkes (German Academic Scholarship Foundation) in Bled, Slovenia, on “Subviral pathogens: Viroids and Prions” in 1994.
4. Prions
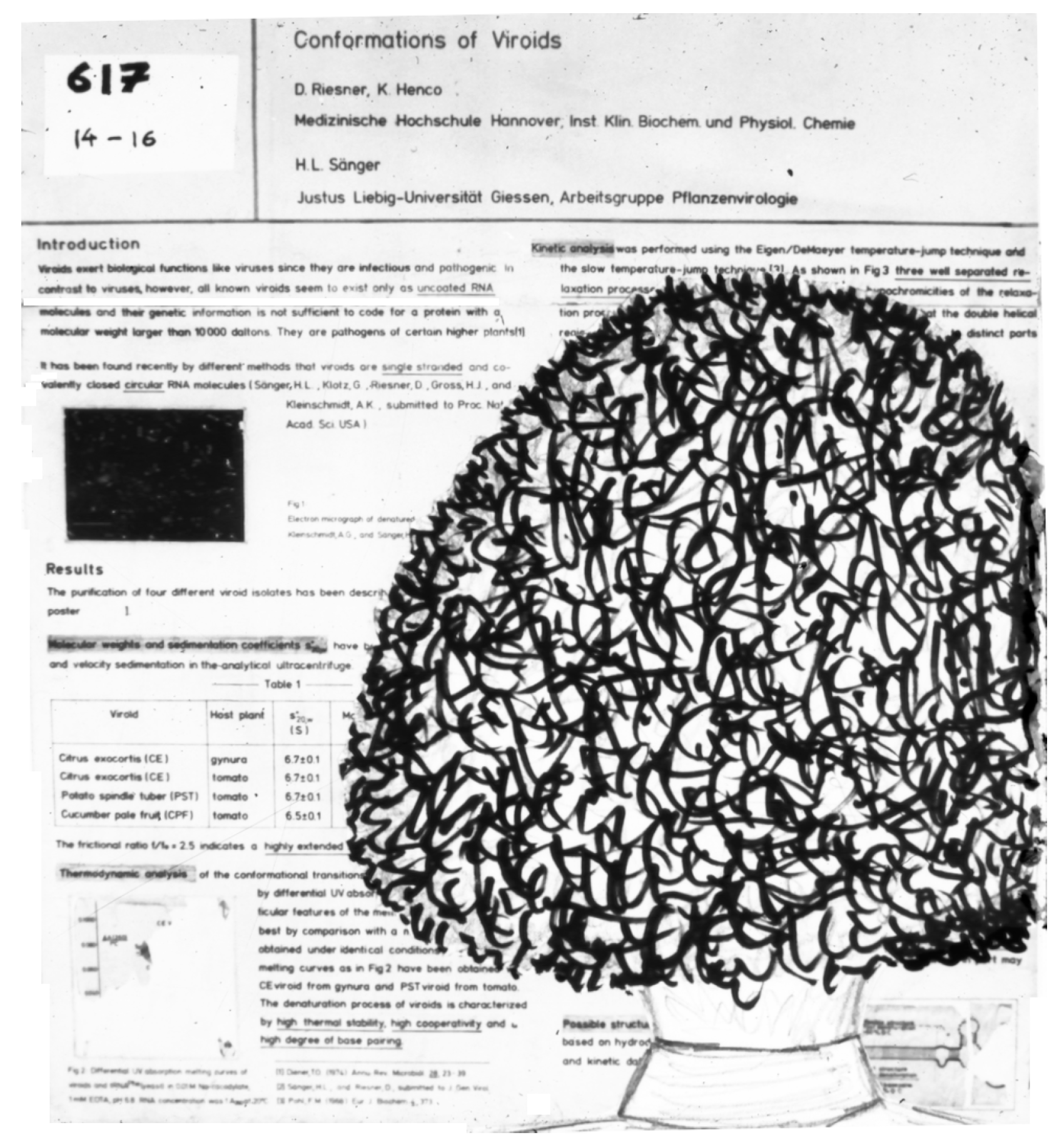
Stanley B. Prusiner systematically continued the earlier studies of Alper et al. [61] into the inactivation properties of scrapie infectivity; Latarjet et al. [89] had shown that the scrapie UV spectrum of inactivation is similar to that of proteins rather than that of nucleic acids. Several years later, Stanley Prusiner and Detlev Riesner had a prolonged discussion about viroids and prions prompted by Riesner’s “Viroid Poster” at the 10th International Congress of Biochemistry, Hamburg (1976). Prusiner came to study the poster and blocked the whole poster with his voluminous hairstyle (Figure 3). While nobody else could look at the poster, Prusiner and Riesner fell into an intensive discussion on viroids and the scrapie agent.
In 1978, during a meeting in Munich, Germany, organized by Sänger, Prusiner reported that the sedimentation behavior of scrapie infectivity was heterogeneous and different from viruses and viroids. Prusiner summarized the results of his subsequent studies: chemical and physical procedures that destroy nucleic acids do not destroy scrapie infectivity, while chemical and physical procedures that destroy proteins do destroy scrapie infectivity. In his groundbreaking publication in 1982 [88], he concluded: “Novel proteinaceous infectious particles cause scrapie”. Whereas viroids were already accepted, prions remained surrounded by considerable skepticism for many more decades. In contrast to most of the skeptics, Ted Diener, who had published a paper entitled “Is the scrapie agent a viroid” [9] in 1972, accepted the new data underpinning the prion hypothesis and published a paper on the fundamental differences between viroids and prions with Prusiner and his colleagues in 1982 [10].
Using some viroid research approaches, Riesner and colleagues applied the quantitative methods of nucleic acid analysis—even counting single molecules—and showed in systematic and extended studies that highly infectious samples from hamster brains contained more infectious units than nucleic acid molecules longer than 25 nucleotides [57,58]. We will not outline the whole research development on prions here; Prusiner was awarded the Nobel Prize “for his discovery of Prions—a new biological principle of infection” [56] in 1997, and the final molecular proof was found in 2004 when a synthetic prion—i. e., a synthetic protein that never has seen an animal—was created [55,90].
Due to the ongoing research of Prusiner’s group and others, the present day mechanism of prion amplification was determined. The prion protein (PrP) can exist in a cellular, non-pathological conformation (PrPc), which is mainly -helical, or in an aggregated, pathological conformation (PrPSc, i. e., scrapie), which has a higher proportion of -sheets. PrPc is expressed from a single-copy gene of the host [59] and presented on the outer surface of the cell. If PrPc encounters invading PrPSc during infection, PrPSc forces PrPc to misfold into the pathological conformation of PrPSc. When PrPSc aggregates, then breaks apart and encounters nascent PrPc, a PrPSc dimer or oligomer is formed. This replication process continues until the accumulation of PrPSc prions kills the host cells.
The infectivity of prions results from a conformational change of PrPc into PrPSc. The first PrPSc particles can be generated spontaneously by the rare aggregation of several PrPc molecules or can be facilitated by specific mutations. The foregoing process produces prion diseases that can originate spontaneously, by infection or as familial diseases. Prion disease was the first example in medicine of three origins of the same disease [91].
5. Satellites and Virusoids
At the conference in Munich in 1978, Jacobus (“Jap”) M. Kaper presented data that CARNA 5 (cucumber mosaic virus |CMV] associated RNA 5), the satellite RNA of CMV (satCMV), was able to modify CMV symptoms depending on CMV strain, CARNA 5 variant, and host plant [49,92]. Virions of CMV contain the three genomic RNAs 1–3, a subgenomic RNA 4, and sometimes a fifth RNA, giving rise to the name CARNA 5. Thus, an RNA was identified that is similar in size to viroids, can also induce pathological effects, and—in contrast to viroids—is linear, depending on a helper CMV to replicate and accumulating in planta in double-stranded form [93].
Plant infections with CMV and necrogenic CARNA 5 variants lead to systemic necrosis and destruction of plants [46,48], exemplified by several epidemics [45]. In contrast, infections with CMV and non-necrogenic variants show only marginal symptoms and a yield that is similar to that of non-infected plants, even above that of plants infected with CMV without a satellite. This opened the possibility to (cross-)protect crops by pre-infection or “vaccination” with CMV and non-necrogenic satellites [94,95]. It was important, however, to verify that during this biological control process, no necrogenic CARNA 5 variants were emerging; that is, the non-necrogenic variant should differ from any necrogenic variant by more than a single mutation [96]. Following this line, Tien Po (Academia Sinica, Beijing, China) analyzed double-stranded CARNA 5 (dsCARNA 5) variants during his sabbatical stay in Düsseldorf in 1986 and 1987. The major method for analysis was temperature-gradient gel electrophoresis (TGGE) [97,98] that can separate macromolecules of identical length that differ by a few or even single mutations, which influence their denaturation behavior. Indeed, several dsCARNA 5 samples consisted of different molecular species that were only separated at temperatures leading to a partial denaturation but co-migrated as dsRNA at low temperature and as single strands after full denaturation. Common to all samples were three transitions: two low-temperature transitions, due to partial denaturation leading to strong gel retardation, and a high-temperature transition into single strands, which migrated much faster than the partially denatured molecules. For the non-necrogenic variants, the transition at the lowest temperature was below the corresponding transition of the necrogenic variants [99,100].
Virusoids are small, circular, single-stranded satellite RNAs that are encapsidated by respective plant viruses and replicated by the viral polymerase; their native secondary structure is mostly rod-like with a few small bifurcations. John W. Randles, recipient of a Ludwig Leichhardt fellowship from the Alexander von Humboldt Foundation, arrived for his sabbatical in Düsseldorf with viroid and virusoid samples in 1981. Together, Randles, Steger and Riesner showed that CCCVd possesses thermodynamic and kinetic features of a typical viroid including an extra-stable hairpin I but not an extra-stable hairpin II. In contrast, the virusoids of subterranean clover mottle virus (satSNMV) and velvet tobacco mottle virus (satVTMoV) are thermodynamically less stable than viroids, despite similar GC content, and do not possess extra-stable hairpin(s) [24,101]. Thomas C. Goodman, a postdoctoral fellow of the Alexander von Humboldt Foundation, showed that DNA-dependent RNA polymerase II, the enzyme that replicates members of Pospiviroidae, binds PSTVd specifically at one end of its secondary structure. In contrast, satSNMV and satVTMoV are only nonspecifically bound, similar to tRNA, for example [102].
6. Outlook
The discovery of viroids was a strenuous dive into the new world of subviral pathogens and included a prolonged battle against the disbelief of many scientists. Consequently, we cannot forget the pioneering work of Theodor O. Diener and his successors: Joseph S. Semancik [69] and Heinz L. Sänger [70], who described first the citrus exocortis viroid; Rudra P. Singh and his work on PSTVd [103]; John W. Randles and his discovery of CCCVd [31,32]; and Robert H. Symons, who first described the ASBVd and its self-splicing by hammerhead ribozymes [28,104]. The discovery of new viroid(-like) RNAs has increased in recent years mostly due to the use of high-throughput sequencing [105,106,107,108].
Of course, the knowledge on these subviral particles has expanded greatly over the following half century. Viroids are now subdivided into the families Pospiviroidae and Avsunviroidae, which are located mainly in the nucleus and the chloroplast, respectively. Their locations also suggest their usage of different replication strategies, host polymerases, structural elements, and trafficking [13,109,110,111]. The knowledge on viroid-induced symptoms is increasing, but still not solved fully. At least in the case of peach latent mosaic viroid variants that induce an extensive chlorosis of peach, a specific viroid-derived small RNA is involved [15,112]; mechanisms are less clear in other cases [113,114]. Elimination of some viroids during pollen maturation involves a depression of viroid replication and an increase in degradation processes [115,116].
Current knowledge on replication of virusoids and satellite RNAs by the polymerase of their helper virus is discussed in recent reviews [117,118] showing that complex secondary and tertiary structures of satellite RNAs are involved in suppression of the helper virus and symptoms [119,120,121,122,123].
What started with research in plants and animals had later impact in general molecular biology, extending to human disease and therapeutic developments. Here follows a few spot lights:
- Hammerhead ribozymes have been detected in most genomes [16,127]. Retrotransposons with hammerhead ribozymes, called retrozymes, have been found encoded in diverse plant genomes [17] and have stimulated new ideas about the possible origin of viroid and viroid-like RNAs [128,129,130,131]. Viroid-like RNAs, termed mycoviroids, have been detected in fungi [132,133,134].
- In the early 1980s, there was only a minor interest in rare neurodegenerative diseases like kuru, Creutzfeldt–Jakob and Gerstmann–Sträussler–Scheinker in humans and scrapie in sheep, which seems not to be transmissible to humans. This situation changed drastically with Prusiner’s concept of prions and the discovery of the bovine spongiform encephalopathy (BSE) epidemic in the mid-1980s. Later, variant Creutzfeldt–Jakob disease (vCJD) was found to be due to bovine prions. With widespread testing of slaughtered cattle in Europe and elimination of bovine offal as a source of feed for cattle, sheep, and pigs, BSE has been eliminated from the roster of lethal human illnesses.
Much more than a spotlight is our knowledge of prions that is leading to effective therapeutics for Alzheimer’s and Parkinson’s diseases as well as related disorders. Recent breakthroughs in cryo-electron microscopy and solid-state NMR spectroscopy are helping elucidate the fibril structures of proteins that cause prion diseases [50,51,52,54,142]. Hopefully, by understanding the biophysics, molecular biology and protein interactions involved in these diseases, effective therapeutics can be developed.
Author Contributions
Writing—original draft preparation, G.S. and D.R.; writing—review and editing, G.S., D.R. and S.B.P. All authors have read and agreed to the published version of the manuscript.
Funding
The APC was funded by Heinrich Heine University Düsseldorf.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Crick, F. On protein synthesis. Symp. Soc. Exp. Biol. 1958, 12, 138–163. [Google Scholar]
- Crick, F. Central dogma of molecular biology. Nature 1970, 227, 561–563. [Google Scholar] [CrossRef] [PubMed]
- Diener, T.; Raymer, W. Potato spindle tuber virus: A plant virus with properties of a free nucleic acid. Science 1967, 158, 378–381. [Google Scholar] [CrossRef] [PubMed]
- Diener, T. Potato spindle tuber “virus” IV. A replicating, low molecular weight RNA. Virology 1971, 45, 411–428. [Google Scholar] [CrossRef] [PubMed]
- Diener, T. Viroids: The smallest known agents of infectious disease. Annu. Rev. Microbiol. 1974, 28, 23–39. [Google Scholar] [CrossRef] [PubMed]
- Diener, T. Discovering viroids–a personal perspective. Nat. Rev. Microbiol. 2003, 1, 75–80. [Google Scholar] [CrossRef]
- Bolton, D.; McKinley, M.; Prusiner, S. Identification of a protein that purifies with the scrapie prion. Science 1982, 218, 1309–1311. [Google Scholar] [CrossRef]
- Prusiner, S.; Bolton, D.; Groth, D.; Bowman, K.; Cochran, S.; McKinley, M. Further purification and characterization of scrapie prions. Biochemistry 1982, 21, 6942–6950. [Google Scholar] [CrossRef]
- Diener, T. Is the scrapie agent a viroid? Nat. New. Biol. 1972, 235, 218–219. [Google Scholar] [CrossRef] [PubMed]
- Diener, T.; McKinley, M.; Prusiner, S. Viroids and prions. Proc. Natl. Acad. Sci. USA 1982, 79, 5220–5224. [Google Scholar] [CrossRef]
- Schneider, I. Satellite-like particle of tobacco ringspot virus that resembles tobacco ringspot virus. Science 1969, 166, 1627–1629. [Google Scholar] [CrossRef] [PubMed]
- Schneider, I. Characteristics of a satellite-like virus of tobacco ringspot virus. Virology 1971, 45, 108–122. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.; Dissanayaka Mudiyanselage, S.; Park, W.; Wang, M.; Takeda, R.; Liu, B.; Wang, Y. A nuclear import pathway exploited by pathogenic noncoding RNAs. Plant Cell 2022, 34, 3543–3556. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Qu, J.; Ji, S.; Wallace, A.; Wu, J.; Li, Y.; Gopalan, V.; Ding, B. A land plant-specific transcription factor directly enhances transcription of a pathogenic noncoding RNA template by DNA-dependent RNA polymerase II. Plant Cell 2016, 28, 1094–1107. [Google Scholar] [CrossRef] [PubMed]
- Navarro, B.; Gisel, A.; Rodio, M.; Delgado, S.; Flores, R.; Di Serio, F. Small RNAs containing the pathogenic determinant of a chloroplast-replicating viroid guide the degradation of a host mRNA as predicted by RNA silencing. Plant J. 2012, 70, 991–1003. [Google Scholar] [CrossRef] [PubMed]
- Seehafer, C.; Kalweit, A.; Steger, G.; Gräf, S.; Hammann, C. From alpaca to zebrafish: Hammerhead ribozymes wherever you look. RNA 2011, 17, 21–26. [Google Scholar] [CrossRef] [PubMed]
- de la Peña, M.; Cervera, A. Circular RNAs with hammerhead ribozymes encoded in eukaryotic genomes: The enemy at home. RNA Biol. 2017, 14, 985–991. [Google Scholar] [CrossRef]
- Gago, S.; Elena, S.; Flores, R.; Sanjuán, R. Extremely high mutation rate of a hammerhead viroid. Science 2009, 323, 1308. [Google Scholar] [CrossRef]
- López-Carrasco, A.; Ballesteros, C.; Sentandreu, V.; Delgado, S.; Gago-Zachert, S.; Flores, R.; Sanjuán, R. Different rates of spontaneous mutation of chloroplastic and nuclear viroids as determined by high-fidelity ultra-deep sequencing. PLoS Pathog. 2017, 13, e1006547. [Google Scholar] [CrossRef]
- Itaya, A.; Folimonov, A.; Matsuda, Y.; Nelson, R.; Ding, B. Potato spindle tuber viroid as inducer of RNA silencing in infected tomato. Mol. Plant Micr. Int. 2001, 14, 1332–1334. [Google Scholar] [CrossRef]
- Papaefthimiou, I.; Hamilton, A.; Denti, M.; Baulcombe, D.; Tsagris, M.; Tabler, M. Replicating potato spindle tuber viroid RNA is accompanied by short RNA fragments that are characteristic of post-transcriptional gene silencing. Nucleic Acids Res. 2001, 29, 2395–2400. [Google Scholar] [CrossRef] [PubMed]
- Repsilber, D.; Wiese, U.; Rachen, M.; Schröder, A.; Riesner, D.; Steger, G. Formation of metastable RNA structures by sequential folding during transcription: Time-resolved structural analysis of potato spindle tuber viroid (–)-stranded RNA by temperature-gradient gel electrophoresis. RNA 1999, 5, 574–584. [Google Scholar] [CrossRef] [PubMed]
- Loss, P.; Schmitz, M.; Steger, G.; Riesner, D. Formation of a thermodynamically metastable structure containing hairpin II is critical for infectivity of potato spindle tuber viroid RNA. EMBO J. 1991, 10, 719–727. [Google Scholar] [CrossRef]
- Steger, G.; Hofmann, H.; Förtsch, J.; Gross, H.; Randles, J.; Sänger, H.; Riesner, D. Conformational transitions in viroids and virusoids: Comparison of results from energy minimization algorithm and from experimental data. J. Biomol. Struct. Dyn. 1984, 2, 543–571. [Google Scholar] [CrossRef] [PubMed]
- Colpan, M.; Schumacher, J.; Brüggemann, W.; Sänger, H.; Riesner, D. Large-scale purification of viroid RNA using Cs2SO4 gradient centrifugation and high-performance liquid chromatography. Anal. Biochem. 1983, 131, 257–265. [Google Scholar] [CrossRef]
- Mühlbach, H.; Sänger, H. Viroid replication is inhibited by α-amanitin. Nature 1979, 278, 185–188. [Google Scholar] [CrossRef]
- Schindler, I.M.; Mühlbach, H.P. Involvement of nuclear DNA-dependent RNA polymerases in potato spindle tuber viroid replication: A reevaluation. Plant Sci. 1992, 84, 221–229. [Google Scholar] [CrossRef]
- Palukaitis, P.; Hatta, T.; Alexander, D.; Symons, R. Characterization of a viroid associated with avocado sunblotch disease. Virology 1979, 99, 145–151. [Google Scholar] [CrossRef]
- Gross, H.; Domdey, H.; Lossow, C.; Jank, P.; Raba, M.; Alberty, H.; Sänger, H. Nucleotide sequence and secondary structure of potato spindle tuber viroid. Nature 1978, 273, 203–208. [Google Scholar] [CrossRef]
- Sänger, H.; Klotz, G.; Riesner, D.; Gross, H.; Kleinschmidt, A. Viroids are single-stranded covalently closed circular RNA molecules existing as highly base-paired rod-like structures. Proc. Natl. Acad. Sci. USA 1976, 73, 3852–3856. [Google Scholar] [CrossRef] [PubMed]
- Randles, J. Association of two ribonucleic acid species with cadang-cadang disease of coconut palm. Phytopathology 1975, 65, 163–167. [Google Scholar] [CrossRef]
- Randles, J.; Rillo, E.; Diener, T. The viroidlike structure and cellular location of anomalous RNA associated with the cadang-cadang disease. Virology 1976, 74, 128–139. [Google Scholar] [CrossRef] [PubMed]
- Davies, J.; Kaesberg, P.; Diener, T. Potato spindle tuber viroid. XII. An investigation of viroid RNA as a messenger for protein synthesis. Virology 1974, 61, 281–286. [Google Scholar] [CrossRef] [PubMed]
- Hall, T.; Wepprich, R.; Davies, J.; Weathers, L.; Semancik, J. Functional distinctions between the ribonucleic acids from citrus exocortis viroid and plant viruses: Cell-free translation and aminoacylation reactions. Virology 1974, 61, 486–492. [Google Scholar] [CrossRef] [PubMed]
- Katsarou, K.; Adkar-Purushothama, C.; Tassios, E.; Samiotaki, M.; Andronis, C.; Lisón, P.; Nikolaou, C.; Perreault, J.; Kalantidis, K. Revisiting the non-coding nature of pospiviroids. Cells 2022, 11, 265. [Google Scholar] [CrossRef]
- Diener, T. Potato spindle tuber virus: A plant virus with properties of a free nucleic acid. III. Subcellular location of PSTV-RNA and the question of whether virions exist in extracts or in situ. Virology 1971, 43, 75–89. [Google Scholar] [CrossRef]
- Schumacher, J.; Sänger, H.; Riesner, D. Subcellular localization of viroids in highly purified nuclei from tomato leaf tissue. EMBO J. 1983, 2, 1549–1555. [Google Scholar] [CrossRef]
- Harders, J.; Lukács, N.; Robert-Nicoud, M.; Jovin, T.; Riesner, D. Imaging of viroids in nuclei from tomato leaf tissue by in situ hybridization and confocal laser scanning microscopy. EMBO J. 1989, 8, 3941–3949. [Google Scholar] [CrossRef] [PubMed]
- Qi, Y.; Ding, B. Differential subnuclear localization of RNA strands of opposite polarity derived from an autonomously replicating viroid. Plant Cell 2003, 15, 2566–2577. [Google Scholar] [CrossRef]
- Martin, W. “Spindle tuber,” a new potato trouble. Hints to potato growers. N.Y. State Potato Assoc. 1922, 3, 8. [Google Scholar]
- Schultz, E.; Folsom, D. Transmission, variation, and control of certain degeneration diseases of Irish potatoes. J. Agric. Res. 1923, 25, 43–118. [Google Scholar]
- Bar-Joseph, M. A contribution to the natural history of viroids. Int. Organ. Citrus Virol. Conf. 1996, 13, 226–229. [Google Scholar] [CrossRef]
- Keese, P.; Symons, R. Domains in viroids: Evidence of intermolecular RNA rearrangement and their contribution to viroid evolution. Proc. Natl. Acad. Sci. USA 1985, 82, 4582–4586. [Google Scholar] [CrossRef]
- Wüsthoff, K.; Steger, G. Conserved motifs and domains in members of Pospiviroidae. Cells 2022, 11, 230. [Google Scholar] [CrossRef]
- García-Arenal, F.; Palukaitis, P. Structure and functional relationships of satellite RNAs of cucumber mosaic virus. Curr. Top. Microbiol. Immunol. 1999, 239, 37–63. [Google Scholar] [CrossRef]
- Xu, P.; Roossinck, M. Cucumber mosaic virus D satellite RNA-induced programmed cell death in tomato. Plant Cell 2000, 12, 1079–1092. [Google Scholar] [CrossRef]
- Prody, G.; Bakos, J.; Buzayan, J.; Schneider, I.; Bruening, G. Autolytic processing of dimeric plant virus satellite RNA. Science 1986, 231, 1577–1580. [Google Scholar] [CrossRef]
- Kaper, J.; Waterworth, H. Cucumber mosaic virus associated RNA 5: Causal agent for tomato necrosis. Science 1977, 196, 429–431. [Google Scholar] [CrossRef]
- Waterworth, H.; Kaper, J.; Tousignant, M. CARNA 5. the small cucumber mosaic virus-dependent replicating RNA, regulates disease expression. Science 1979, 204, 845–847. [Google Scholar] [CrossRef]
- Gremer, L.; Schölzel, D.; Schenk, C.; Reinartz, E.; Labahn, J.; Ravelli, R.; Tusche, M.; Lopez-Iglesias, C.; Hoyer, W.; Heise, H.; et al. Fibril structure of amyloid-β(1-42) by cryo-electron microscopy. Science 2017, 358, 116–119. [Google Scholar] [CrossRef]
- Ayers, J.; Lee, J.; Monteiro, O.; Woerman, A.; Lazar, A.; Condello, C.; Paras, N.; Prusiner, S. Different α-synuclein prion strains cause dementia with Lewy bodies and multiple system atrophy. Proc. Natl. Acad. Sci. USA 2022, 119, e2113489119. [Google Scholar] [CrossRef]
- Condello, C.; Maxwell, A.; Castillo, E.; Aoyagi, A.; Graff, C.; Ingelsson, M.; Lannfelt, L.; Bird, T.; Keene, C.; Seeley, W.; et al. Aβ and tau prions feature in the neuropathogenesis of Down syndrome. Proc. Natl. Acad. Sci. USA 2022, 119, e2212954119. [Google Scholar] [CrossRef]
- Artikis, E.; Kraus, A.; Caughey, B. Structural biology of ex vivo mammalian prions. J. Biol. Chem. 2022, 298, 102181. [Google Scholar] [CrossRef]
- Merz, G.; Chalkley, M.; Tan, S.; Tse, E.; Lee, J.; Prusiner, S.; Paras, N.; DeGrado, W.; Southworth, D. Stacked binding of a PET ligand to Alzheimer’s tau paired helical filaments. Nat. Commun. 2023, 14, 3048. [Google Scholar] [CrossRef]
- Legname, G.; Baskakov, I.; Nguyen, H.; Riesner, D.; Cohen, F.; DeArmond, S.; Prusiner, S. Synthetic mammalian prions. Science 2004, 305, 673–676. [Google Scholar] [CrossRef]
- Prusiner, S. Prions. Available online: https://www.nobelprize.org/prizes/medicine/1997/prusiner/lecture/ (accessed on 11 January 2024).
- Kellings, K.; Meyer, N.; Mirenda, C.; Prusiner, S.; Riesner, D. Further analysis of nucleic acids in purified scrapie prion preparations by improved return refocusing gel electrophoresis. J. Gen. Virol. 1992, 73, 1025–1029. [Google Scholar] [CrossRef]
- Safar, J.; Kellings, K.; Serban, A.; Groth, D.; Cleaver, J.; Prusiner, S.; Riesner, D. Search for a prion-specific nucleic acid. J. Virol. 2005, 79, 10796–10806. [Google Scholar] [CrossRef]
- Basler, K.; Oesch, B.; Scott, M.; Westaway, D.; Wälchli, M.; Groth, D.; McKinley, M.; Prusiner, S.; Weissmann, C. Scrapie and cellular PrP isoforms are encoded by the same chromosomal gene. Cell 1986, 46, 417–428. [Google Scholar] [CrossRef]
- Gajdusek, D. Unconventional Viruses and the Origin and Disappearance of Kuru. Available online: https://www.nobelprize.org/prizes/medicine/1976/gajdusek/lecture/ (accessed on 11 January 2024).
- Alper, T.; Cramp, W.; Haig, D.; Clarke, M. Does the agent of scrapie replicate without nucleic acid? Nature 1967, 214, 764–766. [Google Scholar] [CrossRef]
- Leopoldt, J. Nützliche und auf Die Erfahrung Gegründete Einleitung zu der Landwirthschaft: Fünf Theile. Mit Kupfer und Baurissen; Christian Friedrich Günther: Berlin/Glogau, Prussia, Germany, 1759; Available online: https://www.digitale-sammlungen.de/en/view/bsb10228944?page=374.375 (accessed on 11 February 2024).
- Brown, P.; Bradley, R. 1755 and all that: A historical primer of transmissible spongiform encephalopathy. BMJ 1998, 317, 1688–1692. [Google Scholar] [CrossRef]
- Watson, J.; Crick, F. Molecular structure of nucleic acids: A structure for deoxyribose nucleic acid. Nature 1953, 171, 737–738. [Google Scholar] [CrossRef]
- Diener, T.; Raymer, W. Potato spindle tuber virus: A plant virus with properties of a free nucleic acid. II. Characterization and partial purification. Virology 1969, 37, 351–366. [Google Scholar] [CrossRef]
- Semancik, J.; Weathers, L. Exocortis virus of citrus: Association of infectivity with nucleic acid preparations. Virology 1968, 36, 326–328. [Google Scholar] [CrossRef]
- Semancik, J.; Weathers, L. Properties of the infectious forms of exocortis virus of citrus. Phythopathology 1970, 60, 732–736. [Google Scholar] [CrossRef]
- Semancik, J.; Weathers, L. Exocortis virus: An infectious free-nucleic acid plant virus with unusual properties. Virology 1972, 47, 456–466. [Google Scholar] [CrossRef]
- Semancik, J.; Weathers, L. Exocortis disease: Evidence for a new species of “infectious” low molecular weight RNA in plants. Nat. New Biol. 1972, 237, 242–244. [Google Scholar] [CrossRef]
- Sänger, H. An infectious and replicating RNA of low molecular weight: The agent of the exocortis disease of citrus. Adv. Biosc. 1972, 8, 103–116. [Google Scholar] [CrossRef]
- Gross, H.; Domdey, H.; Sänger, H. Comparative oligonucleotide fingerprints of three plant viroids. Nucleic Acids Res. 1977, 4, 2021–2028. [Google Scholar] [CrossRef]
- Singh, A.; Sänger, H. A simple and rapid method for partial purification of viroid and other cellular ribonucleic acids of higher secondary structure. Cell. Mol. Biol. Incl. Cyto. Enzymol. 1979, 25, 291–303. [Google Scholar]
- Singh, R. The discovery and eradication of potato spindle tuber viroid in Canada. VirusDisease 2014, 25, 415–424. [Google Scholar] [CrossRef]
- Riesner, D.; Henco, K.; Rokohl, U.; Klotz, G.; Kleinschmidt, A.; Domdey, H.; Jank, P.; Gross, H.; Sänger, H. Structure and structure formation of viroids. J. Mol. Biol. 1979, 133, 85–115. [Google Scholar] [CrossRef] [PubMed]
- Steger, G.; Perreault, J.P. Structure and associated biological functions of viroids. Adv. Virus Res. 2016, 94, 141–172. [Google Scholar] [CrossRef]
- Henco, K.; Sänger, H.; Riesner, D. Fine structure melting of viroids as studied by kinetic methods. Nucleic Acids Res. 1979, 6, 3041–3059. [Google Scholar] [CrossRef]
- Qu, F.; Heinrich, C.; Loss, P.; Steger, G.; Tien, P.; Riesner, D. Multiple pathways of reversion in viroids for conservation of structural elements. EMBO J. 1993, 12, 2129–2139. [Google Scholar] [CrossRef]
- Schröder, A.; Riesner, D. Detection and analysis of hairpin II, an essential metastable structural element in viroid replication intermediates. Nucleic Acids Res. 2002, 30, 3349–3359. [Google Scholar] [CrossRef]
- Schrader, O.; Baumstark, T.; Riesner, D. A mini-RNA containing the tetraloop, wobble-pair and loop E motifs of the central conserved region of potato spindle tuber viroid is processed into a minicircle. Nucleic Acids Res. 2003, 31, 988–998. [Google Scholar] [CrossRef]
- Schumacher, J.; Randles, J.; Riesner, D. A two-dimensional electrophoretic technique for the detection of circular viroids and virusoids. Anal. Biochem. 1983, 135, 288–295. [Google Scholar] [CrossRef] [PubMed]
- Di Serio, F.; Owens, R.; Li, S.F.; Matoušek, J.; Pallás, V.; Sano, T.; Verhoeven, J.; Vidalakis, G.; Flores, R. Viroids. Available online: https://ictv.global/report/chapter/viroids/viroids (accessed on 11 January 2024).
- Takahashi, T.; Diener, T. Potato spindle tuber viroid. XIV. Replication in nuclei isolated from infected leaves. Virology 1975, 64, 106–114. [Google Scholar] [CrossRef]
- Branch, A.; Robertson, H. A replication cycle for viroids and other small infectious RNAs. Science 1984, 223, 450–455. [Google Scholar] [CrossRef]
- Owens, R.; Hadidi, A. The remarkable legacy of Theodor O. Diener (1921–2023): Preeminent plant pathologist and the discoverer of viroids. Viruses 2023, 15, 1895. [Google Scholar] [CrossRef]
- Alpers, M. The epidemiology of kuru: Monitoring the epidemic from its peak to its end. Philos. Trans. R. Soc. Lond. B. Biol. Sci. 2008, 363, 3707–3713. [Google Scholar] [CrossRef]
- Marsh, R.; Malone, T.; Semancik, J.; Lancaster, W.; Hanson, R. Evidence for an essential DNA component in the Scrapie agent. Nature 1978, 275, 146–147. [Google Scholar] [CrossRef]
- Malone, T.; Marsh, R.; Hanson, R.; Semancik, J. Evidence for the low molecular weight nature of scrapie agent. Nature 1979, 278, 575–576. [Google Scholar] [CrossRef] [PubMed]
- Prusiner, S. Novel proteinaceous infectious particles cause scrapie. Science 1982, 216, 136–144. [Google Scholar] [CrossRef] [PubMed]
- Latarjet, R.; Muel, B.; Haig, D.; Clarke, M.; Alper, T. Inactivation of the scrapie agent by near monochromatic ultraviolet light. Nature 1970, 227, 1341–1343. [Google Scholar] [CrossRef] [PubMed]
- Colby, D.; Prusiner, S. De novo generation of prion strains. Nat. Rev. Microbiol. 2011, 9, 771–777. [Google Scholar] [CrossRef]
- Prusiner, S. Prions. Proc. Natl. Acad. Sci. USA 1998, 95, 13363–13383. [Google Scholar] [CrossRef]
- Kaper, J.; Tousignant, M. Cucumber mosaic virus-associating RNA 5. I. Role of host plant and helper strain in determining amount of associated RNA 5 with virions. Virology 1977, 80, 186–195. [Google Scholar] [CrossRef]
- Habili, N.; Kaper, J. Cucumber mosaic virus-associated RNA 5. VII. Double-stranded form accumulation and disease attenuation in tobacco. Virology 1981, 112, 250–261. [Google Scholar] [CrossRef]
- Tien, P.; Zhang, X.; Qiu, B.; Qin, B.; Wu, G. Satellite RNA for the control of plant diseases caused by cucumber mosaic virus. Ann. Appl. Biol. 1987, 111, 143–152. [Google Scholar] [CrossRef]
- Gallitelli, D.; Vovlas, C.; Martelli, G.; Montasser, M.; Tousignant, M.; Kaper, J. Satellite-mediated protection of tomato against cucumber mosaic virus: II. Field test under natural epidemic conditions in southern Italy. Plant Dis. 1991, 75, 93–95. [Google Scholar] [CrossRef]
- Palukaitis, P.; Roossinck, M. Spontaneous change of a benign satellite RNA of cucumber mosaic virus to a pathogenic variant. Nat. Biotechnol. 1996, 14, 1264–1268. [Google Scholar] [CrossRef]
- Rosenbaum, V.; Riesner, D. Temperature-gradient gel electrophoresis. Thermodynamic analysis of nucleic acids and proteins in purified form and in cellular extracts. Biophys. Chem. 1987, 26, 235–246. [Google Scholar] [CrossRef]
- Riesner, D.; Steger, G. Temperature-gradient gel-electrophoresis. In Handbook of RNA Biochemistry; Hartmann, R., Bindereif, A., Schön, A., Westhof, E., Eds.; Wiley-VCH: Weinheim, Germany, 2014; pp. 427–444. [Google Scholar] [CrossRef]
- Tien, P.; Steger, G.; Rosenbaum, V.; Kaper, J.; Riesner, D. Double-stranded cucumovirus associated RNA 5: Experimental analysis of necrogenic and non-necrogenic variants by temperature-gradient gel electrophoresis. Nucleic Acids Res. 1987, 15, 5069–5083. [Google Scholar] [CrossRef]
- Steger, G.; Tien, P.; Kaper, J.; Riesner, D. Double-stranded cucumovirus associated RNA 5: Which sequence variations may be detected by optical melting and temperature-gradient gel electrophoresis? Nucleic Acids Res. 1987, 15, 5085–5103. [Google Scholar] [CrossRef]
- Randles, J.; Steger, G.; Riesner, D. Structural transitions in viroid-like RNAs associated with cadang-cadang disease, velvet tobacco mottle virus, and Solanum nodiflorum mottle virus. Nucleic Acids Res. 1982, 10, 5569–5586. [Google Scholar] [CrossRef]
- Goodman, T.; Nagel, L.; Rappold, W.; Klotz, G.; Riesner, D. Viroid replication: Equilibrium association constant and comparative activity measurements for the viroid-polymerase interactions. Nucleic Acids Res. 1984, 12, 6231–6246. [Google Scholar] [CrossRef] [PubMed]
- Singh, R.; Clark, M. Infectious low-molecular weight ribonucleic acid from tomato. Biochem. Biophys. Res. Commun. 1971, 44, 1077–1083. [Google Scholar] [CrossRef] [PubMed]
- Hutchins, C.; Rathjen, P.; Forster, A.; Symons, R. Self-cleavage of plus and minus RNA transcripts of avocado sunblotch viroid. Nucleic Acids Res. 1986, 14, 3627–3640. [Google Scholar] [CrossRef]
- Hadidi, A. Next-Generation Sequencing and CRISPR/Cas13 Editing in Viroid Research and Molecular Diagnostics. Viruses 2019, 11, 120. [Google Scholar] [CrossRef]
- Adkar-Purushothama, C.; Iyer, P.; Sano, T.; Perreault, J. sRNA Profiler: A user-focused interface for small RNA mapping and profiling. Cells 2021, 10, 1771. [Google Scholar] [CrossRef]
- Raza, A.; Ding, S.; Wu, Q. Culture-independent discovery of viroids by deep sequencing and computational algorithms. Methods Mol. Biol. 2022, 2316, 251–274. [Google Scholar] [CrossRef]
- Nabi, S.; Baranwal, V.; Rao, G.; Mansoor, S.; Vladulescu, C.; Raja, W.; Jan, B.; Alansi, S. High-throughput RNA sequencing of mosaic infected and non-infected apple (Malus × domestica Borkh.) cultivars: From detection to the reconstruction of whole genome of viruses and viroid. Plants 2022, 11, 675. [Google Scholar] [CrossRef] [PubMed]
- Ortolá, B.; Daròs, J. Viroids: Non-coding circular RNAs able to autonomously replicate and infect higher plants. Biology 2023, 12, 172. [Google Scholar] [CrossRef]
- Steger, G.; Wüsthoff, K.; Matoušek, J.; Riesner, D. Viroids: Non-coding circular RNAs are tiny pathogens provoking a broad response in host plants. In RNA Structure and Function, RNA Technolgies 14; Barciszewski, J., Ed.; Springer: Cham, Germany, 2023; pp. 295–309. [Google Scholar] [CrossRef]
- Ma, J.; Dissanayaka Mudiyanselage, S.; Hao, J.; Wang, Y. Cellular roadmaps of viroid infection. Trends Microbiol. 2023, 31, 1179–1191. [Google Scholar] [CrossRef] [PubMed]
- Delgado, S.; Navarro, B.; Serra, P.; Gentit, P.; Cambra, M.; Chiumenti, M.; De Stradis, A.; Di Serio, F.; Flores, R. How sequence variants of a plastid-replicating viroid with one single nucleotide change initiate disease in its natural host. RNA Biol. 2019, 16, 906–917. [Google Scholar] [CrossRef] [PubMed]
- Flores, R.; Navarro, B.; Delgado, S.; Serra, P.; Di Serio, F. Viroid pathogenesis: A critical appraisal of the role of RNA silencing in triggering the initial molecular lesion. FEMS Microbiol. Rev. 2020, 44, 386–398. [Google Scholar] [CrossRef]
- Di Serio, F.; Owens, R.; Navarro, B.; Serra, P.; Martínez de Alba, Á.; Delgado, S.; Carbonell, A.; Gago-Zachert, S. Role of RNA silencing in plant-viroid interactions and in viroid pathogenesis. Virus Res. 2023, 323, 198964. [Google Scholar] [CrossRef]
- Matoušek, J.; Steinbachová, L.; Drábková, L.; Kocábek, T.; Potěšil, D.; Mishra, A.; Honys, D.; Steger, G. Elimination of viroids from tobacco pollen involves a decrease in propagation rate and an increase of the degradation processes. Int. J. Mol. Sci. 2020, 21, 3029. [Google Scholar] [CrossRef]
- Matoušek, J.; Wüsthoff, K.; Steger, G. “Pathomorphogenic” changes caused by citrus bark cracking viroid and transcription factor TFIIIA-7ZF variants support viroid propagation in tobacco. Int. J. Mol. Sci. 2023, 24, 7790. [Google Scholar] [CrossRef]
- Huang, Y.W.; Chung-Chi Hu, C.C.; Hsu, Y.H.; Na-Sheng Lin, N.S. Replication of satellites. In Viroids and Satellites; Hadidi, A., Randles, J., Flores, R., Palukaitis, P., Eds.; Academic Press, Elsevier: Amsterdam, The Netherlands, 2017; pp. 577–586. [Google Scholar] [CrossRef]
- Badar, U.; Venkataraman, S.; AbouHaidar, M.; Hefferon, K. Molecular interactions of plant viral satellites. Virus Genes 2021, 57, 1–22. [Google Scholar] [CrossRef]
- Masuta, C.; Shimura, H. Satellite RNAs: Their involvement in pathogenesis and RNA silencing. In Viroids and Satellites; Hadidi, A., Randles, J., Flores, R., Palukaitis, P., Eds.; Academic Press, Elsevier: Amsterdam, The Netherlands, 2017; pp. 587–596. [Google Scholar] [CrossRef]
- Du, Q.; Duan, C.; Zhang, Z.; Fang, Y.; Fang, R.; Xie, Q.; Guo, H. DCL4 targets cucumber mosaic virus satellite RNA at novel secondary structures. J. Virol. 2007, 81, 9142–9151. [Google Scholar] [CrossRef] [PubMed]
- Cao, X.; Liu, S.; Yu, C.; Li, X.; Yuan, X. A new strategy of using satellite RNA to control viral plant diseases: Post-inoculation with satellite RNA attenuates symptoms derived from pre-infection with its helper virus. Plant. Biotechnol. J. 2019, 17, 1856–1858. [Google Scholar] [CrossRef] [PubMed]
- He, L.; Wang, Q.; Gu, Z.; Liao, Q.; Palukaitis, P.; Du, Z. A conserved RNA structure is essential for a satellite RNA-mediated inhibition of helper virus accumulation. Nucleic Acids Res. 2019, 47, 8255–8271. [Google Scholar] [CrossRef]
- Wang, C.; Jiang, F.; Zhu, S. Complex small RNA-mediated regulatory networks between viruses/viroids/satellites and host plants. Virus Res. 2022, 311, 198704. [Google Scholar] [CrossRef]
- Hecker, R.; Colpan, M.; Riesner, D. High-performance liquid chromatography of DNA restriction fragments. J. Chromatogr. 1985, 326, 251–261. [Google Scholar] [CrossRef]
- Colpan, M.; Riesner, D. High performance liquid chromatography of nucleic acids. In Modern Physical Methods in Biochemistry; New Comprehensive Biochemistry; Neuberger, A., van Deenen, L., Eds.; Elsevier: Amsterdam, The Netherlands, 1985; Volume 11, pp. 85–105. [Google Scholar] [CrossRef]
- Hecker, R.; Riesner, D. Chromatographic separation of DNA restriction fragments. J. Chromatogr. B Biomed. Sci. Appl. 1987, 418, 97–114. [Google Scholar] [CrossRef] [PubMed]
- Lee, B.; Neri, U.; Roux, S.; Wolf, Y.; Camargo, A.; Krupovic, M.; Simmonds, P.; Kyrpides, N.; Gophna, U.; Dolja, V.; et al. Mining metatranscriptomes reveals a vast world of viroid-like circular RNAs. Cell 2023, 186, 646–661.e4. [Google Scholar] [CrossRef]
- Diener, T. Viroids: “living fossils” of primordial RNAs? Biol. Direct 2016, 11, 15. [Google Scholar] [CrossRef]
- Moelling, K.; Broecker, F. Viroids and the Origin of Life. Int. J. Mol. Sci. 2021, 22, 3476. [Google Scholar] [CrossRef]
- Flores, R.; Navarro, B.; Serra, P.; Di Serio, F. A scenario for the emergence of protoviroids in the RNA world and for their further evolution into viroids and viroid-like RNAs by modular recombinations and mutations. Virus Evol. 2022, 8, veab107. [Google Scholar] [CrossRef] [PubMed]
- Lee, B.; Koonin, E. Viroids and viroid-like circular RNAs: Do they descend from primordial replicators? Life 2022, 12, 103. [Google Scholar] [CrossRef]
- Sun, L.; Hadidi, A. Mycoviroids: Fungi as hosts and vectors of viroids. Cells 2021, 11, 1335. [Google Scholar] [CrossRef] [PubMed]
- Tian, M.; Wei, S.; Bian, R.; Luo, J.; Khan, H.; Tai, H.; Kondo, H.; Hadidi, A.; Andika, I.; Sun, L. Natural cross-kingdom spread of apple scar skin viroid from apple trees to fungi. Cells 2022, 11, 3686. [Google Scholar] [CrossRef] [PubMed]
- Forgia, M.; Navarro, B.; Daghino, S.; Cervera, A.; Gisel, A.; Perotto, S.; Aghayeva, D.; Akinyuwa, M.; Gobbi, E.; Zheludev, I.; et al. Hybrids of RNA viruses and viroid-like elements replicate in fungi. Nat. Commun. 2023, 14, 2591. [Google Scholar] [CrossRef]
- Pisignano, G.; Michael, D.; Visal, T.; Pirlog, R.; Ladomery, M.; Calin, G. Going circular: History, present, and future of circRNAs in cancer. Oncogene 2023, 42, 2783–2800. [Google Scholar] [CrossRef]
- Lindner, G.; Takenaka, K.; Santucci, K.; Gao, Y.; Janitz, M. Protein-coding circular RNAs—Mechanism, detection, and their role in cancer and neurodegenerative diseases. Biochem. Biophys. Res. Commun. 2023, 678, 68–77. [Google Scholar] [CrossRef]
- Singh, S.; Sinha, T.; Panda, A. Regulation of microRNA by circular RNA. Wiley Interdiscip. Rev. RNA 2023, 15, e1820. [Google Scholar] [CrossRef]
- Wang, K.; Choo, Q.; Weiner, A.; Ou, J.; Najarian, R.; Thayer, R.; Mullenbach, G.; Denniston, K.; Gerin, J.; Houghton, M. Structure, sequence and expression of the hepatitis delta (δ) viral genome. Nature 1986, 323, 508–514. [Google Scholar] [CrossRef]
- Chen, P.; Kalpana, G.; Goldberg, J.; Mason, W.; Werner, B.; Gerin, J.; Taylor, J. Structure and replication of the genome of the hepatitis delta virus. Proc. Natl. Acad. Sci. USA 1986, 83, 8774–8778. [Google Scholar] [CrossRef]
- Negro, F.; Lok, A. Hepatitis D: A review. JAMA 2023, 330, 2376–2387. [Google Scholar] [CrossRef] [PubMed]
- Vanwolleghem, T.; Armstrong, P.; Buti, M.; FitzSimons, D.; Valckx, S.; Hendrickx, G.; Van, D. The elimination of hepatitis D as a public health problem: Needs and challenges. J. Viral. Hepat. 2024, 31, 47–50. [Google Scholar] [CrossRef] [PubMed]
- Mishra, S. Emerging trends in cryo-EM-based structural studies of neuropathological amyloids. J. Mol. Biol. 2023, 435, 168361. [Google Scholar] [CrossRef] [PubMed]
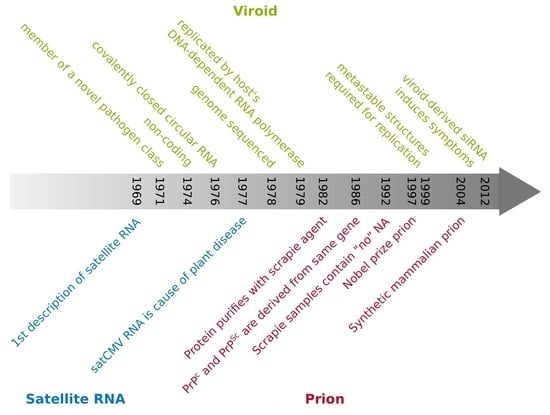
Figure 1.
Timeline of research development of viroids (green text) [4,13,14,15,16,17,18,19,20,21,22,23,24,25,26,27,28,29,30,31,32,33,34,35,36,37,38,39,40,41,42,43,44], satellite RNAs (orange text) [11,12,45,46,47,48,49], prions (red text) [7,50,51,52,53,54,55,56,57,58,59,60,61,62,63], and a few general hallmarks (black text) [64].
Figure 1.
Timeline of research development of viroids (green text) [4,13,14,15,16,17,18,19,20,21,22,23,24,25,26,27,28,29,30,31,32,33,34,35,36,37,38,39,40,41,42,43,44], satellite RNAs (orange text) [11,12,45,46,47,48,49], prions (red text) [7,50,51,52,53,54,55,56,57,58,59,60,61,62,63], and a few general hallmarks (black text) [64].

Figure 2.
At the 7th International Conference of Virology in Edmonton, Canada, 1987. From left to right: Robert A. Owens, Detlev Riesner, Theodor O. Diener, Heinz Ludwig Sänger.
Figure 2.
At the 7th International Conference of Virology in Edmonton, Canada, 1987. From left to right: Robert A. Owens, Detlev Riesner, Theodor O. Diener, Heinz Ludwig Sänger.

Figure 3.
A poster and its visitor at the 10th International Congress of Biochemistry, Hamburg, Germany (1976). Prusiner in front of the poster sketched from memory by Riesner.
Figure 3.
A poster and its visitor at the 10th International Congress of Biochemistry, Hamburg, Germany (1976). Prusiner in front of the poster sketched from memory by Riesner.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Steger, G.; Riesner, D.; Prusiner, S.B. Viroids, Satellite RNAs and Prions: Folding of Nucleic Acids and Misfolding of Proteins. Viruses 2024, 16, 360. https://doi.org/10.3390/v16030360
AMA Style
Steger G, Riesner D, Prusiner SB. Viroids, Satellite RNAs and Prions: Folding of Nucleic Acids and Misfolding of Proteins. Viruses. 2024; 16(3):360. https://doi.org/10.3390/v16030360
Chicago/Turabian StyleSteger, Gerhard, Detlev Riesner, and Stanley B. Prusiner. 2024. "Viroids, Satellite RNAs and Prions: Folding of Nucleic Acids and Misfolding of Proteins" Viruses 16, no. 3: 360. https://doi.org/10.3390/v16030360
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.