Amentoflavone Inhibits HSV-1 and ACV-Resistant Strain Infection by Suppressing Viral Early Infection


 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cells and Viruses
2.2. Compounds, Antibodies, and Reagents
2.3. Cytotoxicity and Antiviral Activity Assay
2.4. Quantitative Real-Time PCR (qRT-PCR)
2.5. Nuclear and Cytoplasmic Protein Extraction
2.6. Western Blot Assay
2.7. Immunofluorescence Assay
2.8. Flow Cytometry Assay
2.9. Luciferase Reporter Gene Assay
2.10. Statistical Analysis
3. Results
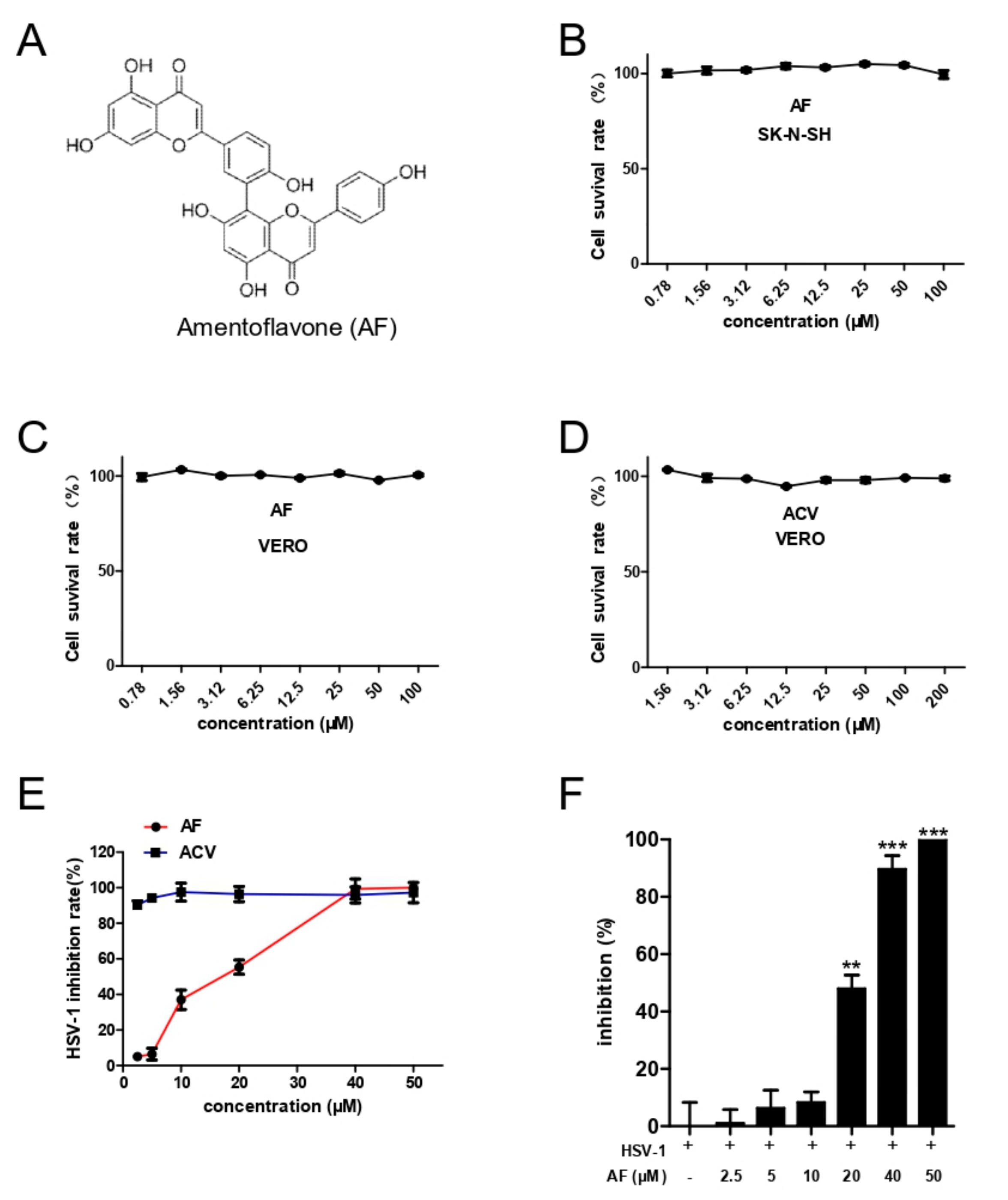
3.1. Cytotoxicity and Comprehensive Antiviral Activity of Amentoflavone
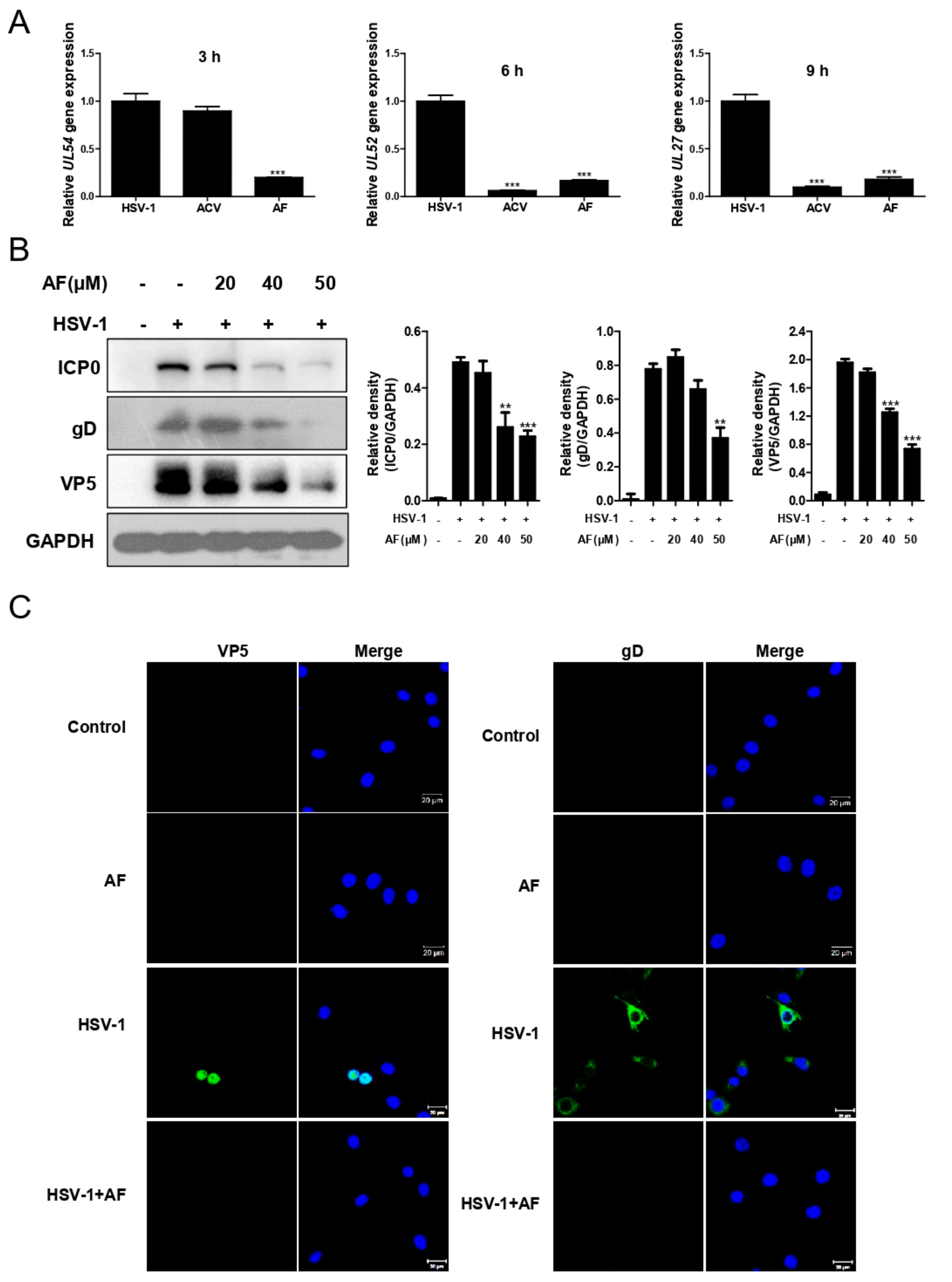
3.2. Amentoflavone Inhibits HSV-1 Gene and Protein Expression
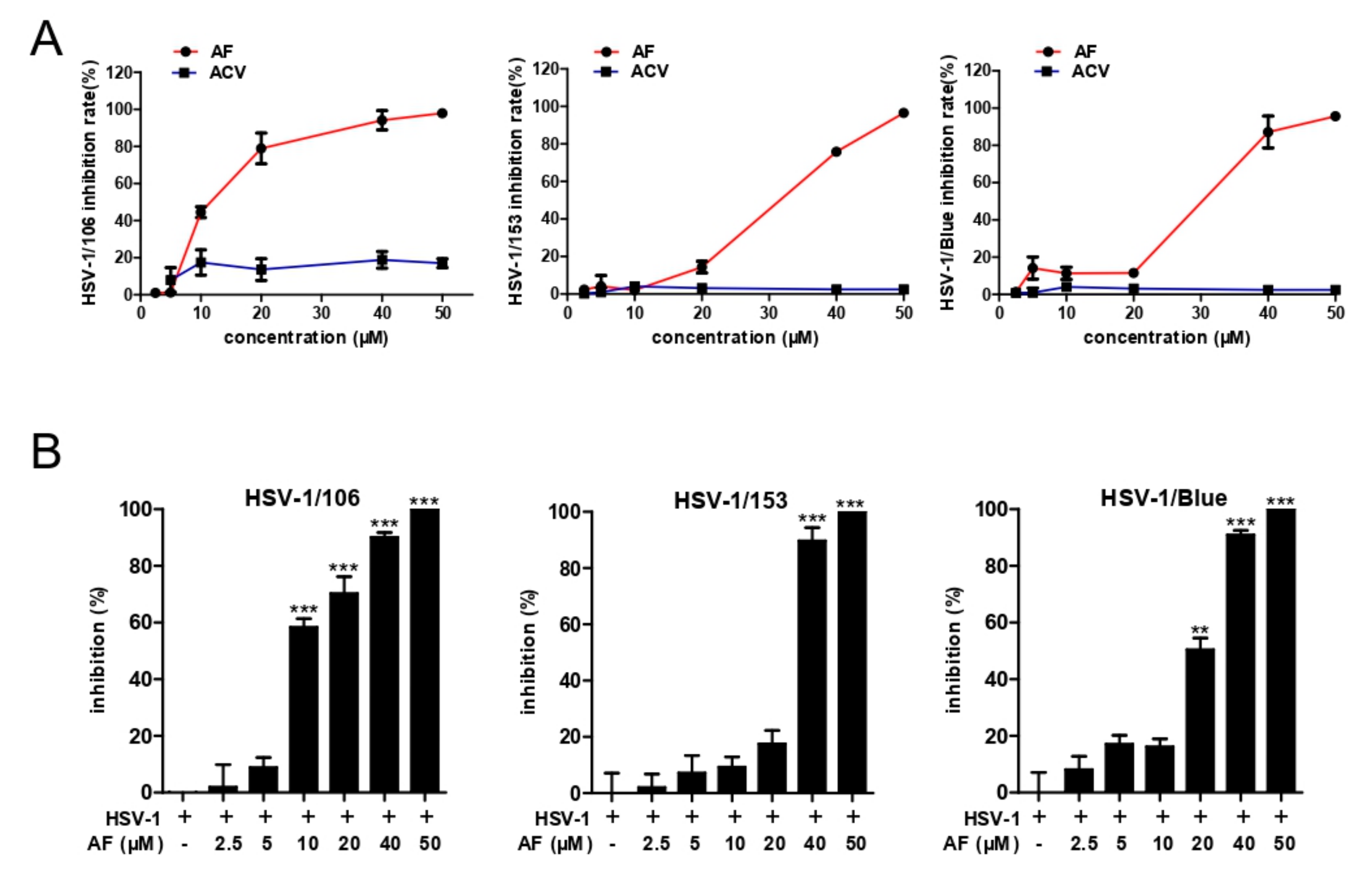
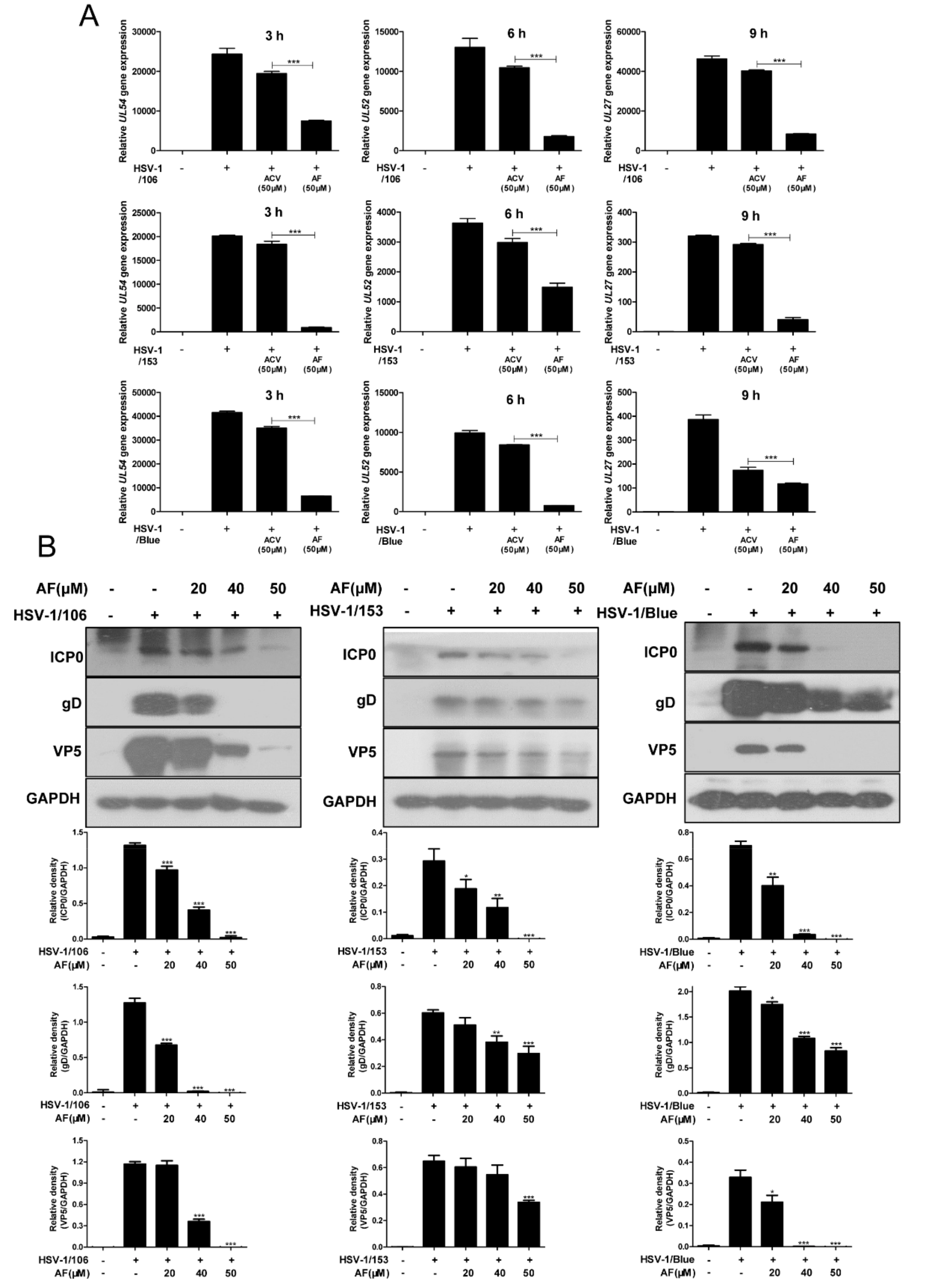
3.3. Amentoflavone Inhibits ACV-Resistant Strains Infection
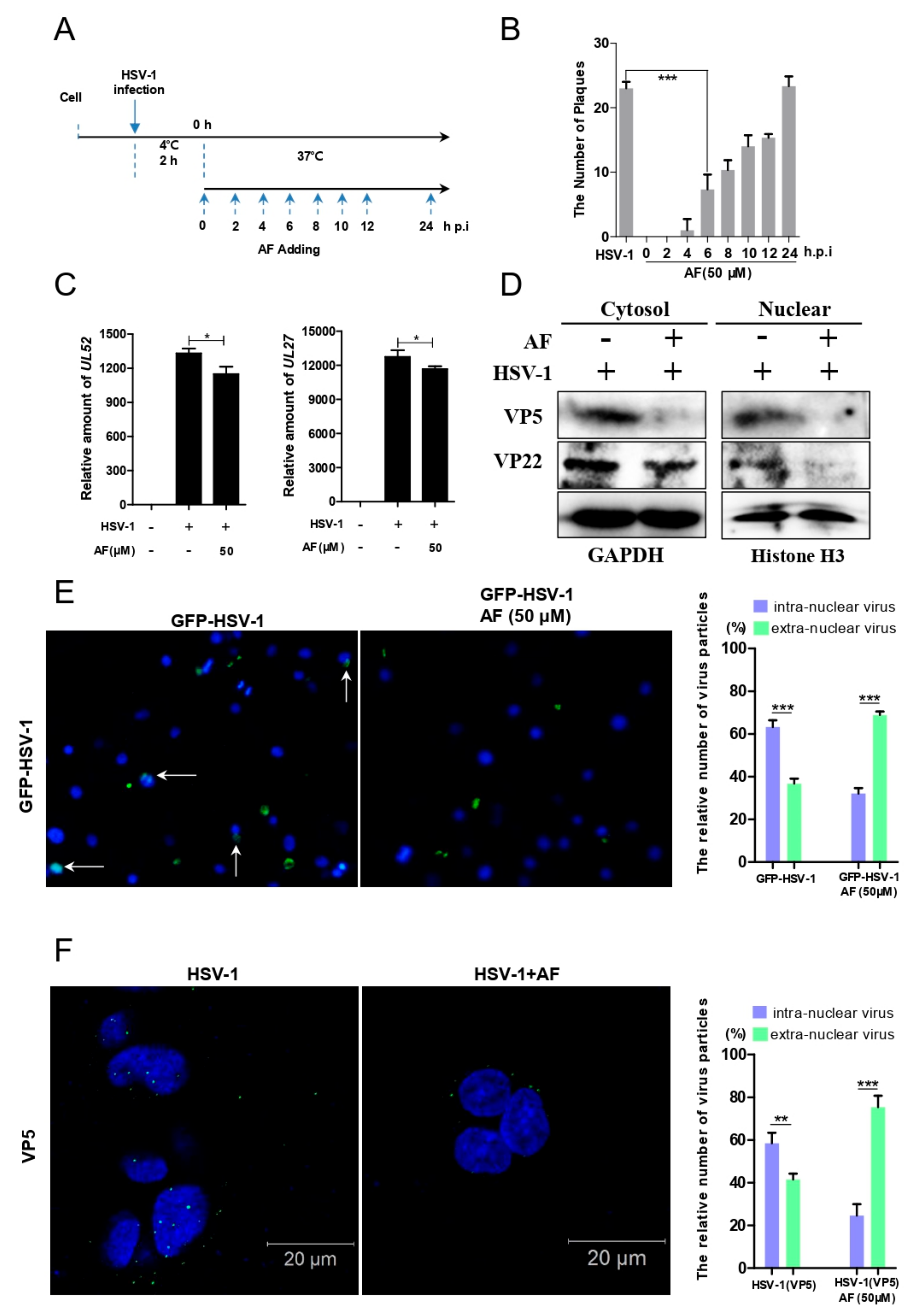
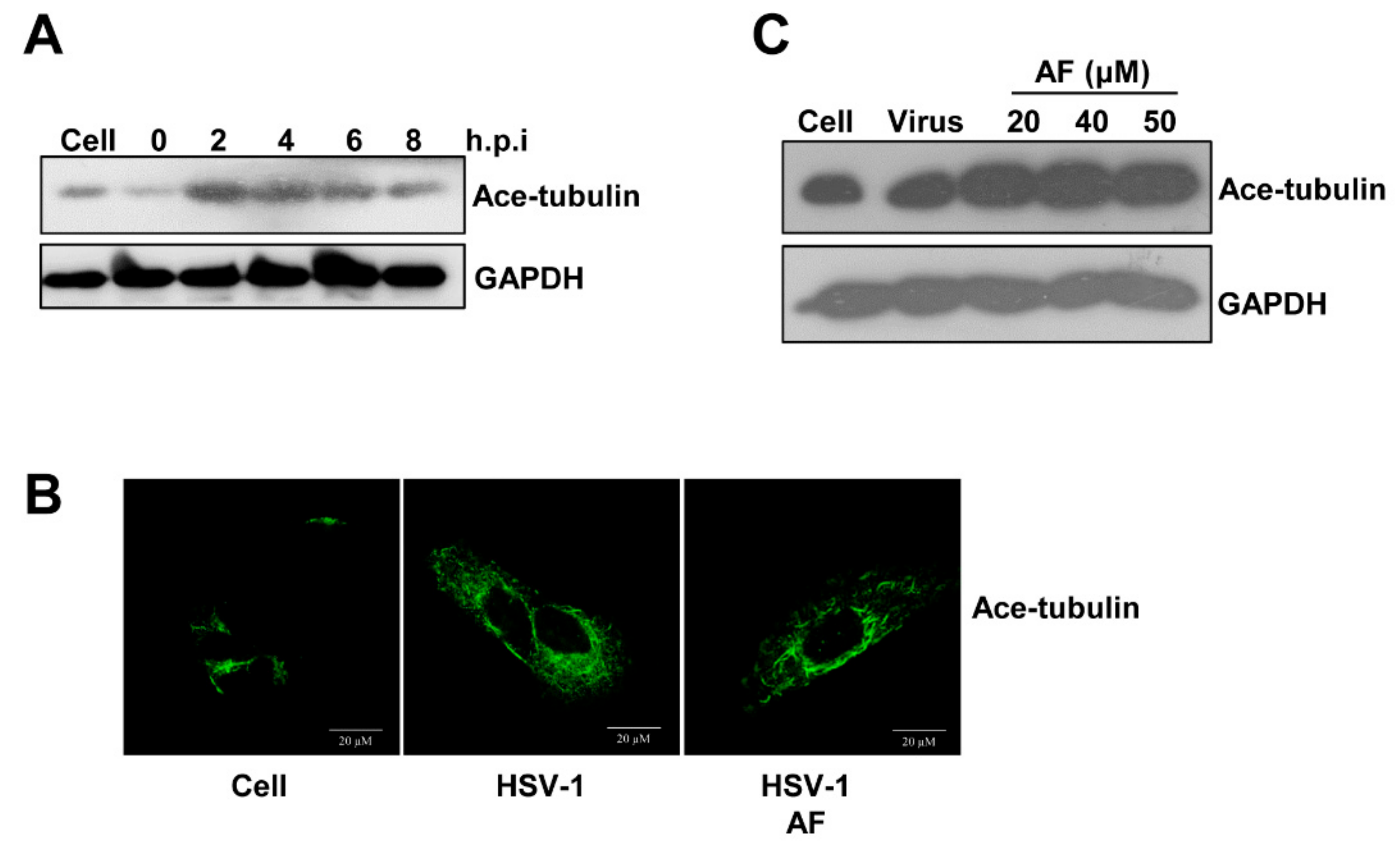
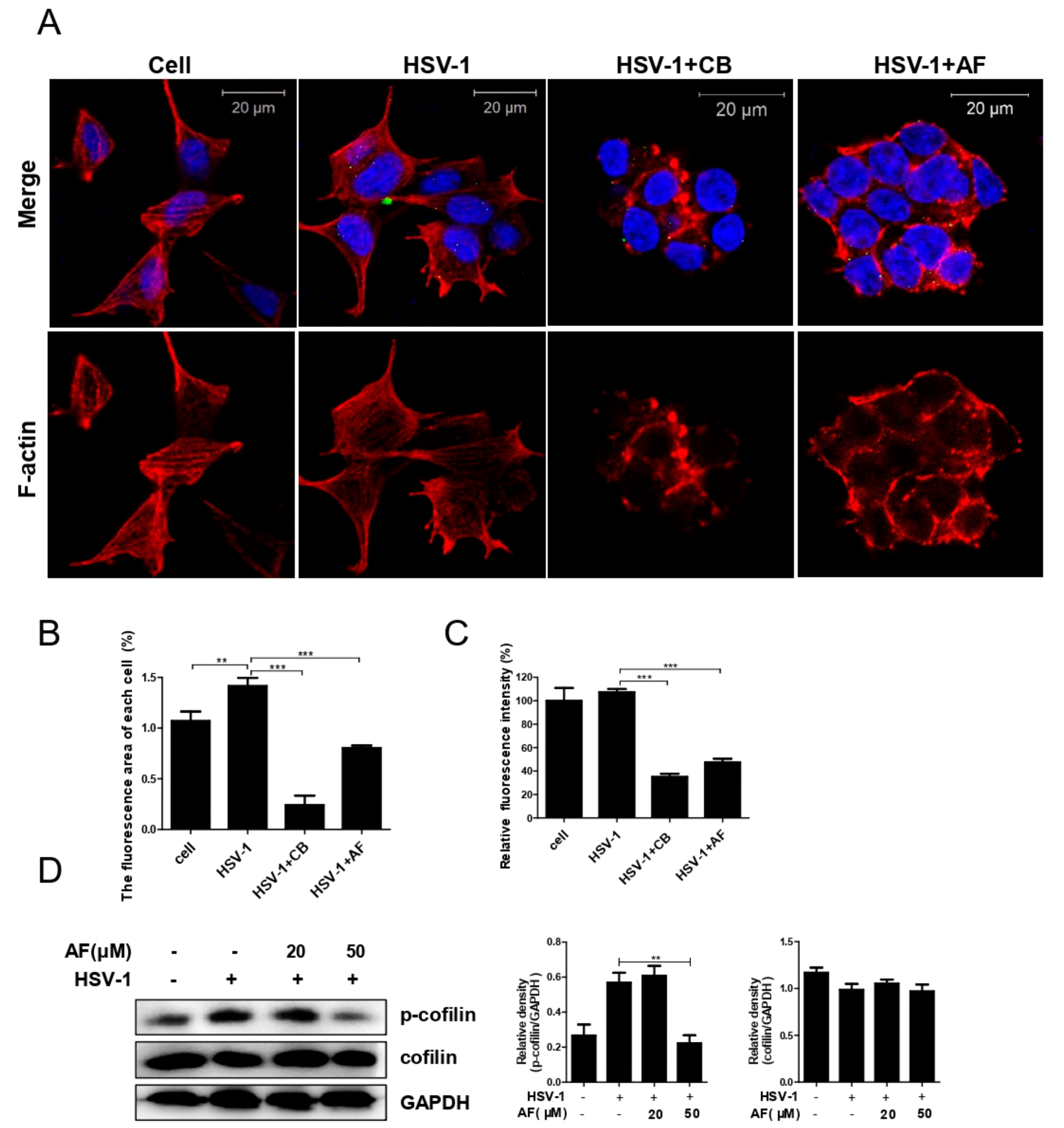
3.4. Amentoflavone Reduces the Nuclear Import of HSV-1
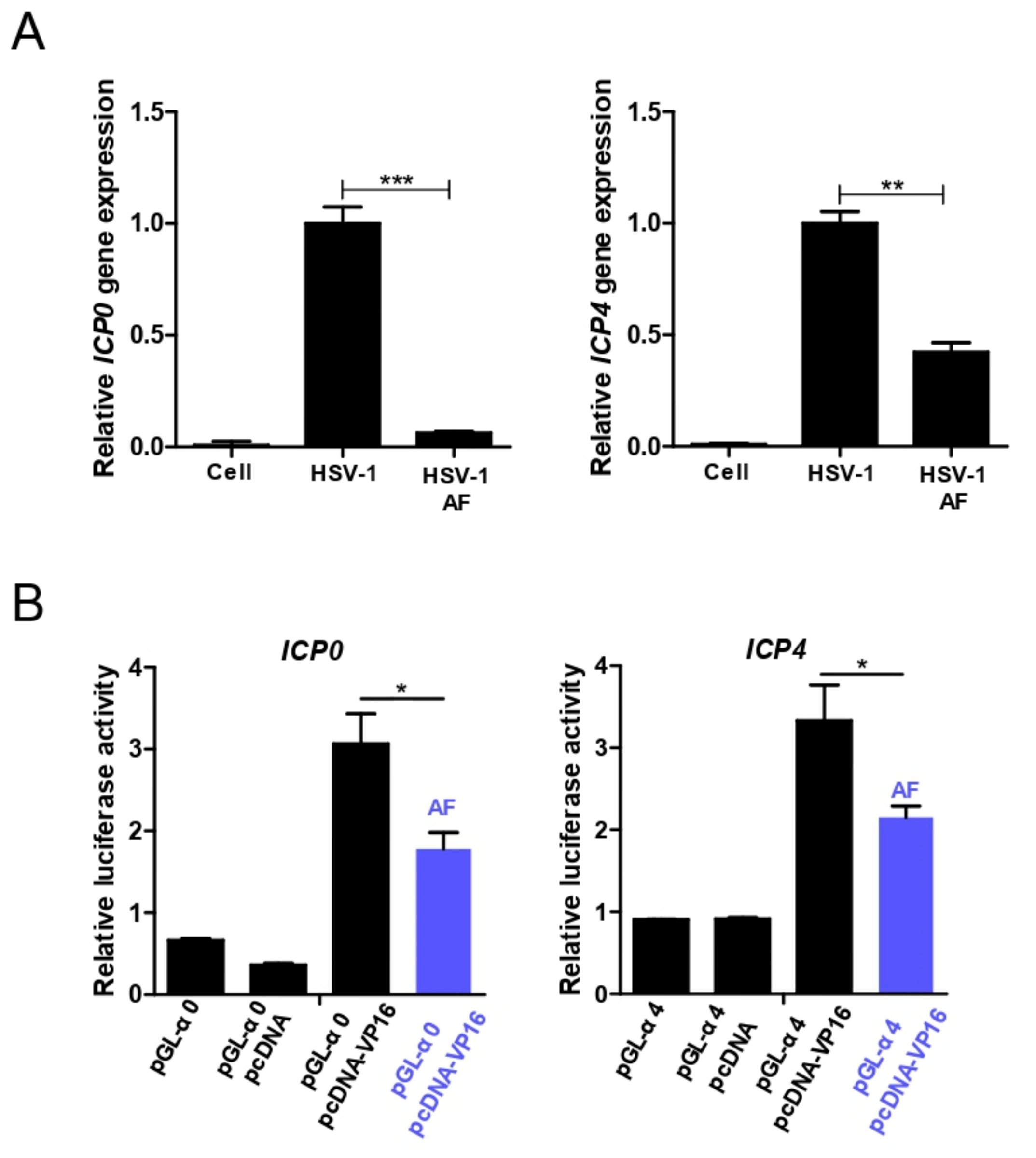
3.5. Amentoflavone Reduces Immediate Early Gene Promoter Activity
4. Discussion
Author Contributions
Funding
Conflicts of Interest
References
- Midaksiewirska, A.; Karabin, K.; Chudzik, E.; Dzieciatkowski, T.; Przybylski, M.; Majewska, A.; Łuczak, M.; Młynarczyk, G. Application of real-time pcr assay for investigating the presence of herpes simplex virus type 1 DNA. Med. Dosw. Mikrobiol. 2010, 62, 85–92. [Google Scholar]
- Kennedy, P.G.E.; Chaudhuri, A. Herpes simplex encephalitis. Tidsskr. Den Nor. Lægeforen. Tidsskr. Prakt. Med. Ny Række 1981, 101, 1577. [Google Scholar] [CrossRef] [PubMed]
- Glaser, R.; Pearson, G.R.; Jones, J.F.; Hillhouse, J.; Kennedy, S.; Mao, H.; Kiecolt-Glaser, J.K. Stress-related activation of epstein-barr virus. Brain Behav. Immun. 1991, 5, 219. [Google Scholar] [CrossRef]
- Agostini, S.; Mancuso, R.; Hernis, A.; Costa, A.S.; Nemni, R.; Clerici, M. HSV-1-specific igg subclasses distribution and serum neutralizing activity in alzheimer’s disease and in mild cognitive impairment. J. Alzheimer’s Dis. 2018, 63, 131–138. [Google Scholar] [CrossRef]
- Gudmundsson, K.S.; Johns, B.A. Imidazo[1,2-]pyridines with potent activity against herpesviruses. Bioorg. Med. Chem. Lett. 2007, 17, 2735–2739. [Google Scholar] [CrossRef] [PubMed]
- Christophers, J.; Clayton, J.; Craske, J.; Ward, R.; Collins, P.; Trowbridge, M.; Darby, G. Survey of resistance of herpes simplex virus to acyclovir in northwest england. Antimicrob. Agents Chemother. 1998, 42, 868. [Google Scholar] [CrossRef] [PubMed]
- Brice, A.; Moseley, G.W. Viral interactions with microtubules: Orchestrators of host cell biology? Future Virol. 2013, 8, 229–243. [Google Scholar] [CrossRef]
- Radtke, K.; Dohner, K.; Sodeik, B. Viral interactions with the cytoskeleton: A hitchhiker’s guide to the cell. Cell Microbiol. 2006, 8, 387–400. [Google Scholar] [CrossRef]
- Favoreel, H.W.; Enquist, L.W.; Feierbach, B. Actin and rho gtpases in herpesvirus biology. Trends Microbiol. 2007, 15, 426–433. [Google Scholar] [CrossRef]
- Haglund, C.M.; Welch, M.D. Pathogens and polymers: Microbe-host interactions illuminate the cytoskeleton. J. Cell Boil. 2011, 195, 7–17. [Google Scholar] [CrossRef]
- Yoder, A.; Yu, D.; Dong, L.; Iyer, S.R.; Xu, X.; Kelly, J.; Liu, J.; Wang, W.; Vorster, P.J.; Agulto, L.; et al. HIV envelope-cxcr4 signaling activates cofilin to overcome cortical actin restriction in resting cd4 t cells. Cell 2008, 134, 782–792. [Google Scholar] [CrossRef]
- Zheng, K.; Kitazato, K.; Wang, Y.; He, Z. Pathogenic microbes manipulate cofilin activity to subvert actin cytoskeleton. Crit. Rev. Microbiol. 2016, 42, 677–695. [Google Scholar] [CrossRef] [PubMed]
- Zheng, K.; Xiang, Y.; Wang, X.; Wang, Q.; Zhong, M.; Wang, S.; Wang, X.; Fan, J.; Kitazato, K.; Wang, Y. Epidermal growth factor receptor-pi3k signaling controls cofilin activity to facilitate herpes simplex virus 1 entry into neuronal cells. mBio 2014, 5, e00958-13. [Google Scholar] [CrossRef]
- Zheng, K.; Kitazato, K.; Wang, Y. Viruses exploit the function of epidermal growth factor receptor. Rev. Med. Virol. 2014, 24, 274–286. [Google Scholar] [CrossRef] [PubMed]
- Jin, F.; Ma, K.; Chen, M.; Zou, M.; Wu, Y.; Li, F.; Wang, Y. Pentagalloylglucose blocks the nuclear transport and nucleocapsid egress process to inhibit hsv-1 infection. Jpn. J. Infect. Dis. 2015, 69. [Google Scholar] [CrossRef] [PubMed]
- Woo, E.R.; Kim, H.J.; Kwak, J.H.; Lim, Y.K.; Park, S.K.; Kim, H.S.; Lee, C.K.; Park, H. Anti-herpetic activity of various medicinal plant extracts. Arch. Pharmacal. Res. 1997, 20, 58–67. [Google Scholar] [CrossRef]
- Li, P.; Yue, G.L.; Kwok, H.F.; Long, C.; Lau, B.S.; Kennelly, E.J. Using ultra-performance liquid chromatography quadrupole time of flight mass spectrometry-based chemometrics for the identification of anti-angiogenic biflavonoids from edible garcinia species. J. Agric. Food Chem. 2017, 65, 8348–8355. [Google Scholar] [CrossRef]
- Marzia, I.; Marco, M.; Catia, G.; Francesca, I.; Franco Francesco, V.; Nadia, M. Flavonoids and biflavonoids in tuscan berries of juniperus communis l.: Detection and quantitation by hplc/dad/esi/ms. J. Agric. Food Chem. 2007, 55, 6596–6602. [Google Scholar]
- Sha, L.; Qiuxia, R.; Cuiping, Y.; Tianhong, Z.; Jinglai, L.; Xiaoying, W.; Xinyan, Q.; Xiaojuan, Z.; Zhe, Z.; Zhenqing, Z. Liquid chromatography-tandem mass spectrometry determination and pharmacokinetic analysis of amentoflavone and its conjugated metabolites in rats. J. Agric. Food Chem. 2015, 63, 1957–1966. [Google Scholar]
- Zhang, Z.; Sun, T.; Niu, J.G.; He, Z.Q.; Liu, Y.; Wang, F. Amentoflavone protects hippocampal neurons: Anti-inflammatory, antioxidative, and antiapoptotic effects. Neural Regen. Res. 2015, 10, 1125–1133. [Google Scholar]
- An, J.; Li, Z.; Dong, Y.; Ren, J.; Huo, J. Amentoflavone protects against psoriasis-like skin lesion through suppression of nf-κb-mediated inflammation and keratinocyte proliferation. Mol. Cell. Biochem. 2016, 413, 87–95. [Google Scholar] [CrossRef]
- Liu, H.; Yue, Q.; He, S. Amentoflavone suppresses tumor growth in ovarian cancer by modulating skp2. Life Sci. 2017, 189, 96. [Google Scholar] [CrossRef] [PubMed]
- Ma, S.C.; But, P.P.; Ooi, V.E.; He, Y.H.; Lee, S.H.; Lee, S.F.; Lin, R.C. Antiviral amentoflavone from selaginella sinensis. Boil. Pharm. Bull. 2001, 24, 311. [Google Scholar] [CrossRef]
- Lin, Y.M.; Flavin, M.T.; Schure, R.; Chen, F.C.; Sidwell, R.; Barnard, D.L.; Huffman, J.H.; Kern, E.R. Antiviral activities of biflavonoids. Planta Med. 1999, 65, 120–125. [Google Scholar] [CrossRef]
- Wilsky, S.; Sobotta, K.; Wiesener, N.; Pilas, J.; Althof, N.; Munder, T.; Wutzler, P.; Henke, A. Inhibition of fatty acid synthase by amentoflavone reduces coxsackievirus b3 replication. Arch Virol. 2012, 157, 259–269. [Google Scholar] [CrossRef] [PubMed]
- Xiang, Y.; Zheng, K.; Ju, H.; Wang, S.; Pei, Y.; Ding, W.; Chen, Z.; Wang, Q.; Qiu, X.; Zhong, M.; et al. Cofilin 1-mediated biphasic f-actin dynamics of neuronal cells affect herpes simplex virus 1 infection and replication. J. Virol. 2012, 86, 8440–8451. [Google Scholar] [CrossRef]
- Wang, Y.; Wang, R.; Li, F.; Wang, Y.; Zhang, Z.; Wang, Q.; Ren, Z.; Jin, F.; Kitazato, K.; Wang, Y. Heat-shock protein 90alpha is involved in maintaining the stability of vp16 and vp16-mediated transactivation of alpha genes from herpes simplex virus-1. Mol. Med. 2018, 24, 65. [Google Scholar] [CrossRef]
- Liu, X.; Main, D.; Ma, Y.; He, B. Herpes simplex virus 1 inhibits tank-binding kinase 1 through formation of the us11-hsp90 complex. J. Virol. 2018. [Google Scholar] [CrossRef]
- Cheng, H.Y.; Lin, C.C.; Lin, T.C. Antiherpes simplex virus type 2 activity of casuarinin from the bark of terminalia arjuna linn. Antivir. Res. 2002, 55, 447–455. [Google Scholar] [CrossRef]
- Reed, L.J.; Muench, H. A simple method of estimating fifty per cent endpoints. Am J Epidemiol. 1938, 27, 493–497. [Google Scholar] [CrossRef]
- Mena, J.A.; Ramírez, O.T.; Palomares, L.A. Titration of non-occluded baculovirus using a cell viability assay. Biotechniques 2003, 34, 260–262. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Fujun, J.; Yiliang, W.; Danlin, Z.; Junwei, L.; Zhen, Z.; Rongze, W.; Dong, D.; Kai, Z.; Yifei, W. Hsp90 inhibitor at-533 blocks hsv-1 nuclear egress and assembly. J. Biochem. 2018, 164, 397–406. [Google Scholar] [CrossRef] [PubMed]
- Pei, Y.; Chen, Z.P.; Ju, H.Q.; Komatsu, M.; Ji, Y.H.; Liu, G.; Guo, C.W.; Zhang, Y.J.; Yang, C.R.; Wang, Y.F. Autophagy is involved in anti-viral activity of pentagalloylglucose (pgg) against herpes simplex virus type 1 infection in vitro. Biochem. Biophys. Res. Commun. 2011, 405, 186–191. [Google Scholar] [CrossRef] [PubMed]
- Burns, W.H.; Saral, R.; Santos, G.W.; Laskin, O.L.; Lietman, P.S.; Mclaren, C.; Barry, D.W. Isolation and characterisation of resistant herpes simplex virus after acyclovir therapy. Lancet 1982, 319, 421–423. [Google Scholar] [CrossRef]
- Jin, F.; Li, S.; Zheng, K.; Zhuo, C.; Ma, K.; Chen, M.; Wang, Q.; Zhang, P.; Fan, J.; Ren, Z.; et al. Silencing herpes simplex virus type 1 capsid protein encoding genes by sirna: A promising antiviral therapeutic approach. PLoS ONE 2014, 9, e96623. [Google Scholar] [CrossRef] [PubMed]
- Zhong, M.; Zheng, K.; Chen, M.; Xiang, Y.; Jin, F.; Ma, K.; Qiu, X.; Wang, Q.; Peng, T.; Kitazato, K.; et al. Heat-shock protein 90 promotes nuclear transport of herpes simplex virus 1 capsid protein by interacting with acetylated tubulin. PLoS ONE 2014, 9, e99425. [Google Scholar] [CrossRef]
- Wysocka, J.; Herr, W. The herpes simplex virus vp16-induced complex: The makings of a regulatory switch. Trends Biochem. Sci. 2003, 28, 294–304. [Google Scholar] [CrossRef]
- Yin, D.; Chen, K.L. Study on the inhibitory action on cvb3 replication in vitro by the extracts from selaginella moelledorfii hieron. Chin. J. Hosp. Pharm. 2009, 29, 349–352. [Google Scholar]
- Sodeik, B.; Ebersold, M.W.; Helenius, A. Microtubule-mediated transport of incoming herpes simplex virus 1 capsids to the nucleus. J. Cell Boil. 1997, 136, 1007–1021. [Google Scholar] [CrossRef] [PubMed]
- Youn, K.J.; Angelo, M.; Chao, M.V.; Ian, M.; Wilson, A.C. Transient reversal of episome silencing precedes vp16-dependent transcription during reactivation of latent hsv-1 in neurons. PLoS Pathog. 2012, 8, e1002540. [Google Scholar]
- Orvedahl, A.; Alexander, D.; Talloczy, Z.; Sun, Q.; Wei, Y.; Zhang, W.; Burns, D.; Leib, D.A.; Levine, B. Hsv-1 icp34.5 confers neurovirulence by targeting the beclin 1 autophagy protein. Cell Host Microbe 2007, 1, 23–35. [Google Scholar] [CrossRef] [PubMed]
- Park, H.J.; Kim, M.M. Amentoflavone induces autophagy and modulates p53. Cell J. 2019, 21, 27–34. [Google Scholar] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cell/Compounds | CC50 (μM) |
|---|---|
| Vero/ACV | >100 |
| Vero/AF | >100 |
| SK-N-SH/AF | >100 |
| Virus (Vero) | EC50 (μM) |
|---|---|
| HSV-1 | 22.13 ± 0.38 |
| HSV-1/106 | 11.11 ± 0.71 |
| HSV-1/153 | 28.22 ± 2.51 |
| HSV-1/Blue | 25.71 ± 3.97 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, F.; Song, X.; Su, G.; Wang, Y.; Wang, Z.; Jia, J.; Qing, S.; Huang, L.; Wang, Y.; Zheng, K.; et al. Amentoflavone Inhibits HSV-1 and ACV-Resistant Strain Infection by Suppressing Viral Early Infection. Viruses 2019, 11, 466. https://doi.org/10.3390/v11050466
Li F, Song X, Su G, Wang Y, Wang Z, Jia J, Qing S, Huang L, Wang Y, Zheng K, et al. Amentoflavone Inhibits HSV-1 and ACV-Resistant Strain Infection by Suppressing Viral Early Infection. Viruses. 2019; 11(5):466. https://doi.org/10.3390/v11050466
Chicago/Turabian StyleLi, Feng, Xiaowei Song, Guifeng Su, Yiliang Wang, Zhaoyang Wang, Jiaoyan Jia, Shurong Qing, Lianzhou Huang, Yuan Wang, Kai Zheng, and et al. 2019. "Amentoflavone Inhibits HSV-1 and ACV-Resistant Strain Infection by Suppressing Viral Early Infection" Viruses 11, no. 5: 466. https://doi.org/10.3390/v11050466
APA StyleLi, F., Song, X., Su, G., Wang, Y., Wang, Z., Jia, J., Qing, S., Huang, L., Wang, Y., Zheng, K., & Wang, Y. (2019). Amentoflavone Inhibits HSV-1 and ACV-Resistant Strain Infection by Suppressing Viral Early Infection. Viruses, 11(5), 466. https://doi.org/10.3390/v11050466