Evolution of the Piscine orthoreovirus Genome Linked to Emergence of Heart and Skeletal Muscle Inflammation in Farmed Atlantic Salmon (Salmo salar)

 ,
,  ,
,  , and
, and
Abstract
1. Introduction
2. Materials and Methods
2.1. Virus Isolates
2.2. RNA Isolation
2.3. RT-qPCR and Sanger Sequencing
2.4. NGS and Genome Assembly
2.5. Phylogenetic Analyses
2.6. Protein Structure Analyses
3. Results
3.1. Coding Sequences Obtained by Illumina and Sanger Sequencing
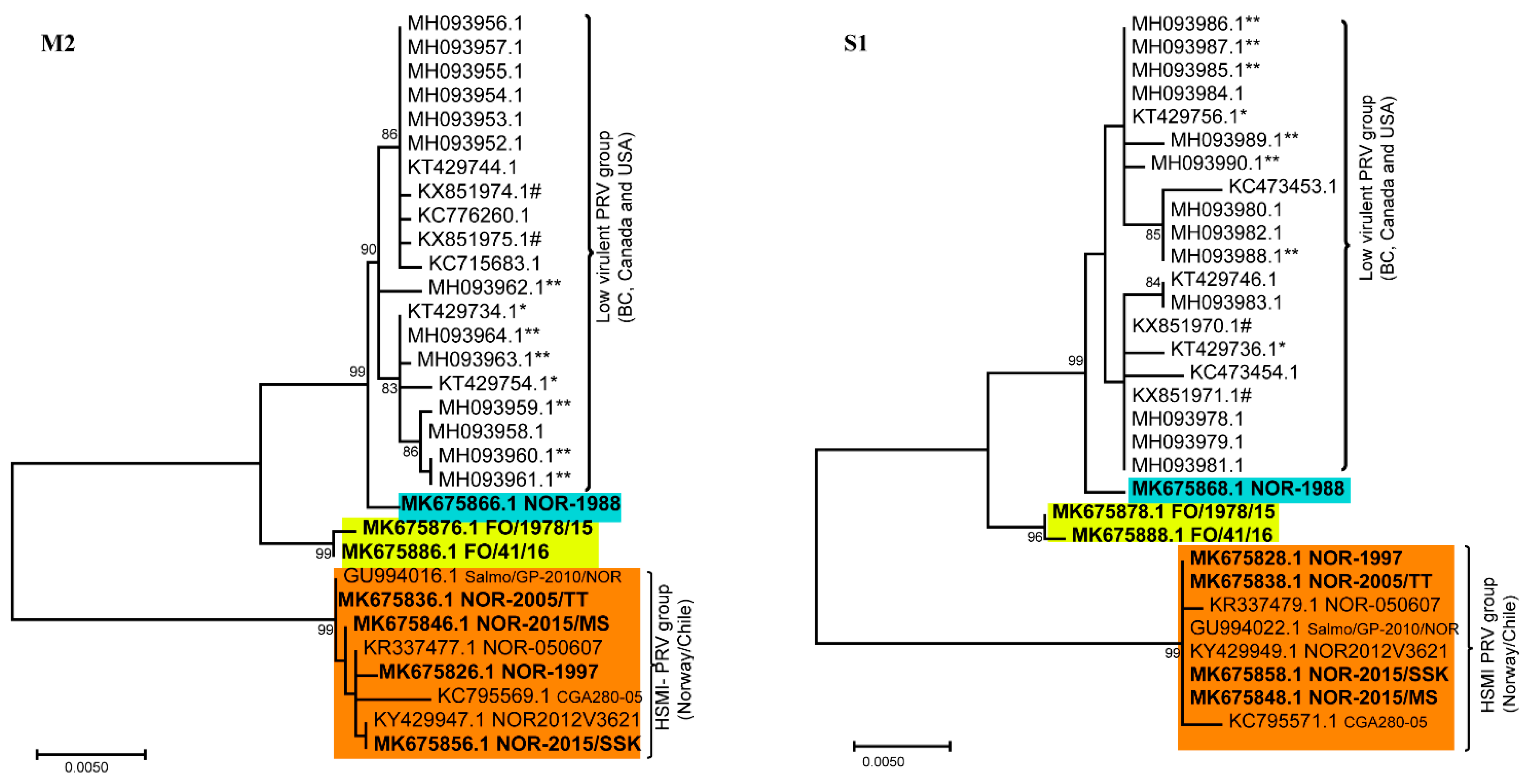
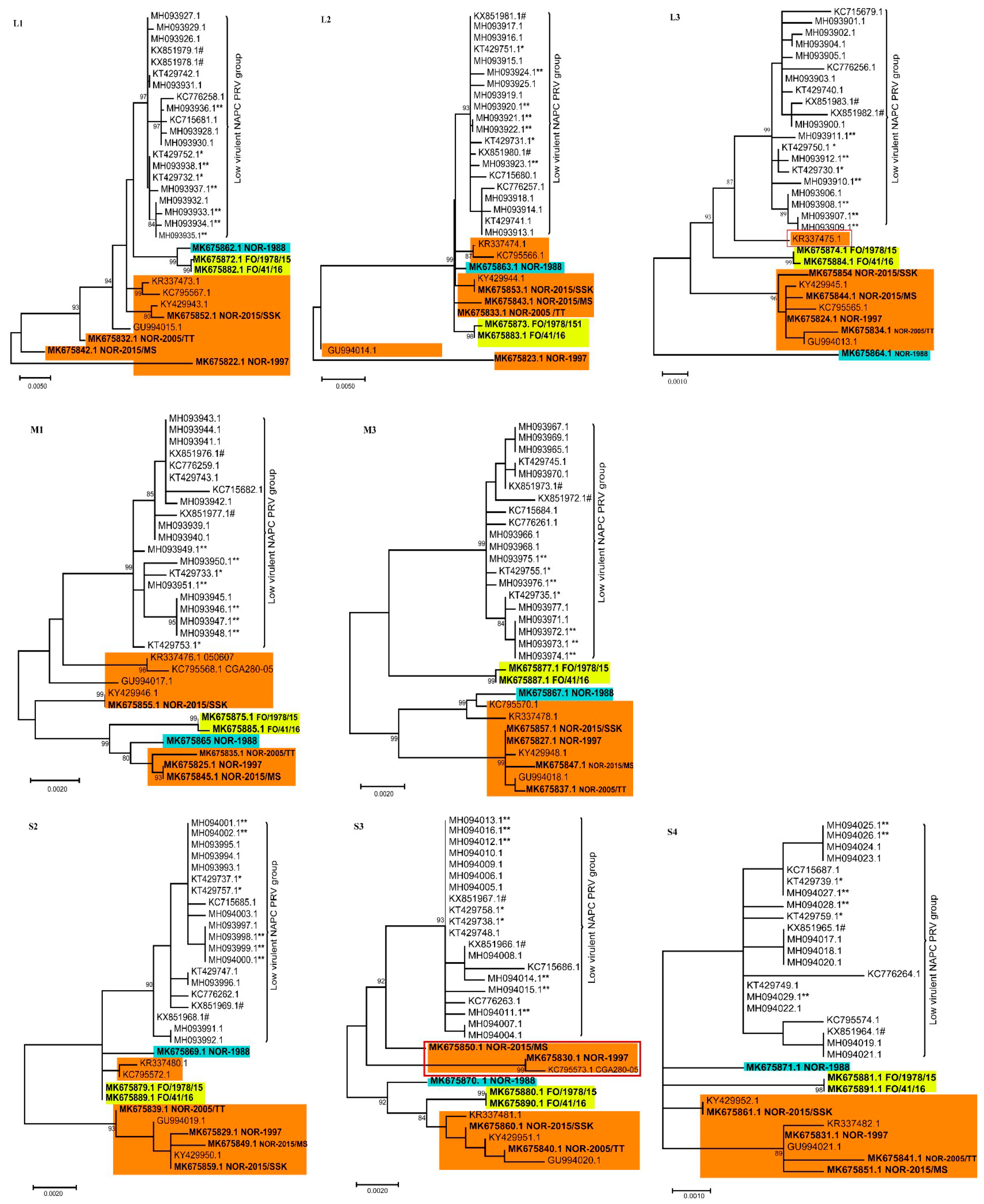
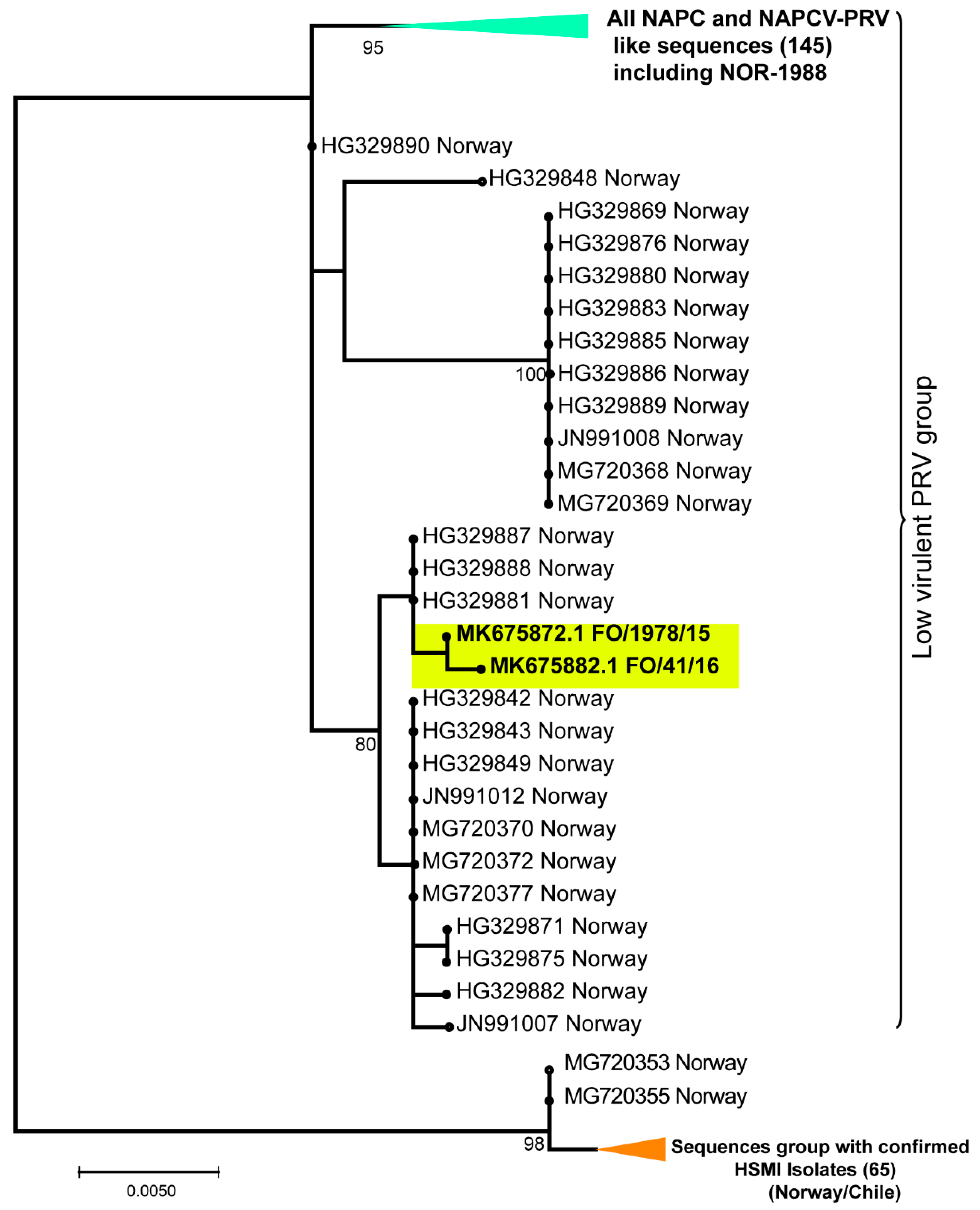
3.2. Phylogenetic Analyses of Individual PRV Genomic Segments Revealed Diversity
3.3. None of the NAPC PRV-1 Isolates Group within the HSMI Clade
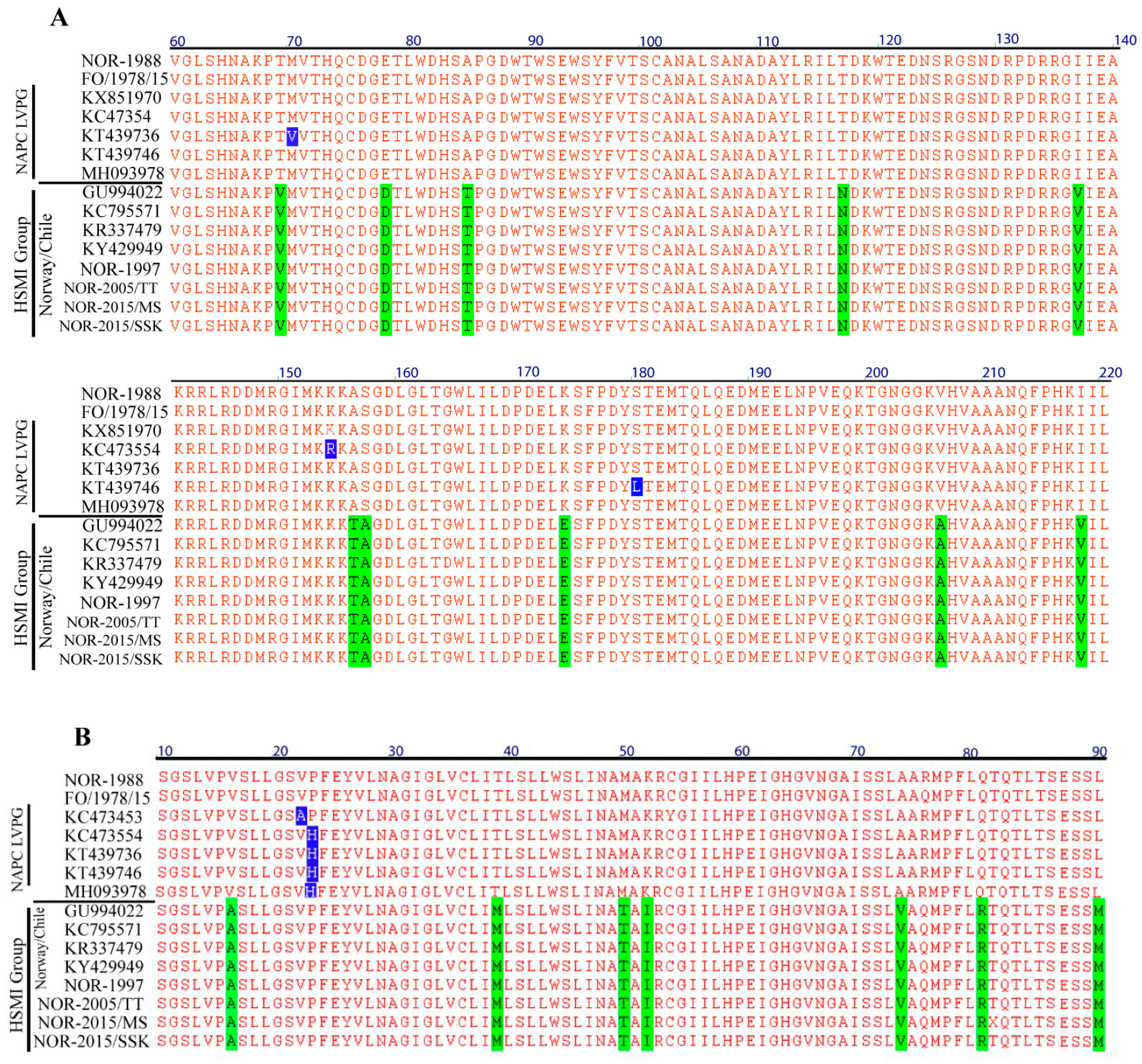
3.4. Amino Acid Residues Unique to HSMI Association are Located to Proteins Encoded by S1 and M2
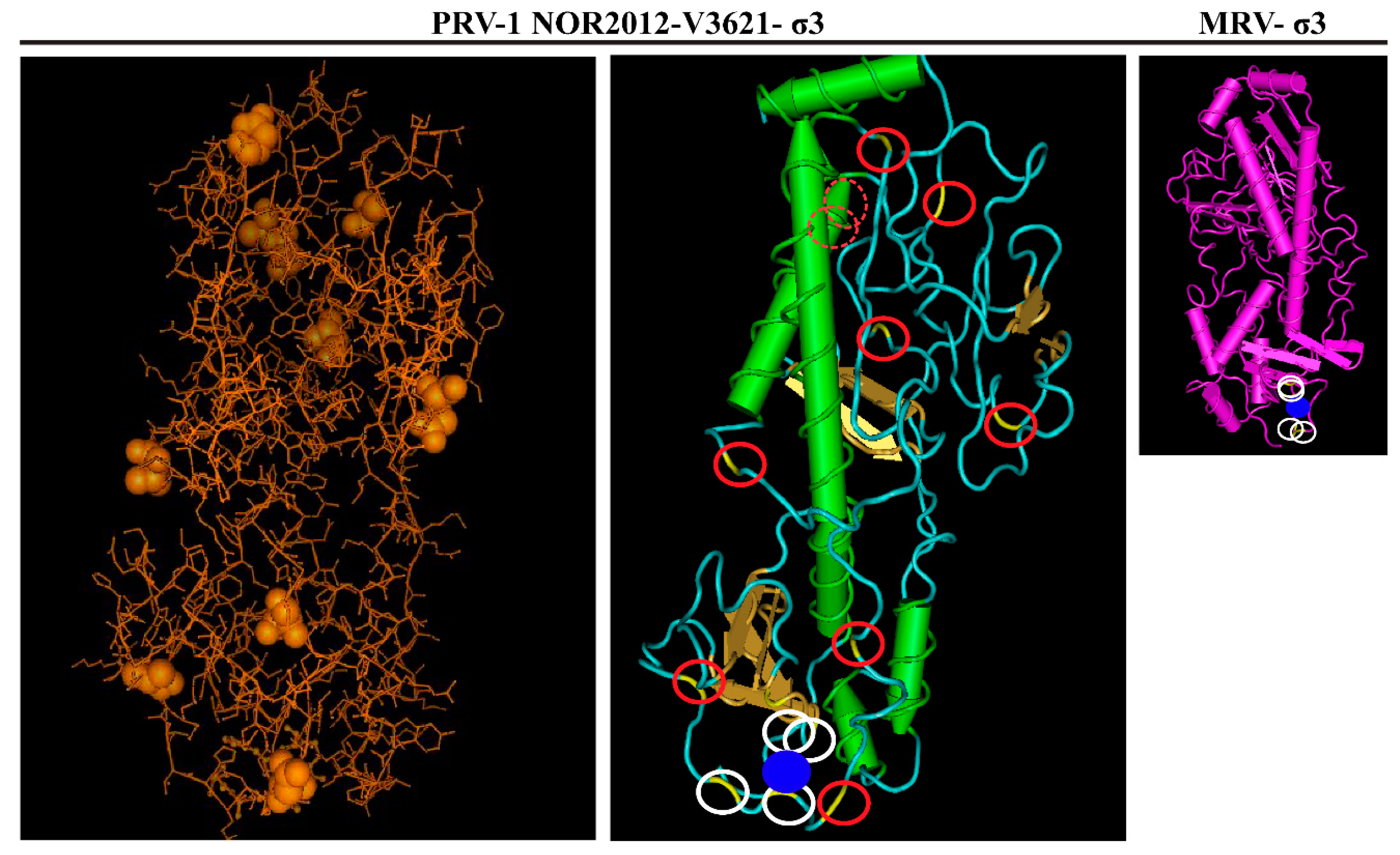
3.5. Predicted Differences between the HSMI Associated and Non-Associated Isolates Locate Mainly to Surface-Exposed Amino Acids in σ3
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Ebert, D.; Bull, J.J. Challenging the trade-off model for the evolution of virulence: Is virulence management feasible? Trends Microbiol. 2003, 11, 15–20. [Google Scholar] [CrossRef]
- Kongtorp, R.T.; Kjerstad, A.; Taksdal, T.; Guttvik, A.; Falk, K. Heart and skeletal muscle inflammation in atlantic salmon, salmo salar l: A new infectious disease. J. Fish Dis. 2004, 27, 351–358. [Google Scholar] [CrossRef] [PubMed]
- Hjeltnes, B.W.C.; Bang Jensen, B.; Haukaas, A. The Fish Health Report 2017; The Norwegian Veterinary Institute: Oslo, Norway, 2018. [Google Scholar]
- Palacios, G.; Lovoll, M.; Tengs, T.; Hornig, M.; Hutchison, S.; Hui, J.; Kongtorp, R.T.; Savji, N.; Bussetti, A.V.; Solovyov, A.; et al. Heart and skeletal muscle inflammation of farmed salmon is associated with infection with a novel reovirus. PLoS ONE 2010, 5, e11487. [Google Scholar] [CrossRef]
- Wessel, O.; Braaen, S.; Alarcon, M.; Haatveit, H.; Roos, N.; Markussen, T.; Tengs, T.; Dahle, M.K.; Rimstad, E. Infection with purified piscine orthoreovirus demonstrates a causal relationship with heart and skeletal muscle inflammation in atlantic salmon. PLoS ONE 2017, 12, e0183781. [Google Scholar] [CrossRef] [PubMed]
- Wessel, O.; Olsen, C.M.; Rimstad, E.; Dahle, M.K. Piscine orthoreovirus (prv) replicates in atlantic salmon (salmo salar l.) erythrocytes ex vivo. Vet. Res. 2015, 46, 26. [Google Scholar] [CrossRef] [PubMed]
- Markussen, T.; Dahle, M.K.; Tengs, T.; Lovoll, M.; Finstad, O.W.; Wiik-Nielsen, C.R.; Grove, S.; Lauksund, S.; Robertsen, B.; Rimstad, E. Sequence analysis of the genome of piscine orthoreovirus (prv) associated with heart and skeletal muscle inflammation (hsmi) in atlantic salmon (salmo salar). PLoS ONE 2013, 8, e70075. [Google Scholar] [CrossRef]
- Haatveit, H.M.; Nyman, I.B.; Markussen, T.; Wessel, O.; Dahle, M.K.; Rimstad, E. The non-structural protein muns of piscine orthoreovirus (prv) forms viral factory-like structures. Vet. Res. 2016, 47, 5. [Google Scholar] [CrossRef]
- Haatveit, H.M.; Wessel, O.; Markussen, T.; Lund, M.; Thiede, B.; Nyman, I.B.; Braaen, S.; Dahle, M.K.; Rimstad, E. Viral protein kinetics of piscine orthoreovirus infection in atlantic salmon blood cells. Viruses 2017, 9, 49. [Google Scholar] [CrossRef] [PubMed]
- Wessel, O.; Nyman, I.B.; Markussen, T.; Dahle, M.K.; Rimstad, E. Piscine orthoreovirus (prv) o3 protein binds dsrna. Virus Res. 2015, 198, 22–29. [Google Scholar] [CrossRef]
- Dryden, K.A.; Wang, G.; Yeager, M.; Nibert, M.L.; Coombs, K.M.; Furlong, D.B.; Fields, B.N.; Baker, T.S. Early steps in reovirus infection are associated with dramatic changes in supramolecular structure and protein conformation: Analysis of virions and subviral particles by cryoelectron microscopy and image reconstruction. J. Cell Biol. 1993, 122, 1023–1041. [Google Scholar] [CrossRef]
- McPhillips, T.H.; Ramig, R.F. Extragenic suppression of temperature-sensitive phenotype in reovirus: Mapping suppressor mutations. Virology 1984, 135, 428–439. [Google Scholar] [CrossRef]
- Sandekian, V.; Lemay, G. Amino acids substitutions in sigma1 and mu1 outer capsid proteins of a vero cell-adapted mammalian orthoreovirus are required for optimal virus binding and disassembly. Virus Res. 2015, 196, 20–29. [Google Scholar] [CrossRef] [PubMed]
- Nibert, M.L.; Fields, B.N. A carboxy-terminal fragment of protein mu 1/mu 1c is present in infectious subvirion particles of mammalian reoviruses and is proposed to have a role in penetration. J. Virol. 1992, 66, 6408–6418. [Google Scholar] [PubMed]
- Key, T.; Read, J.; Nibert, M.L.; Duncan, R. Piscine reovirus encodes a cytotoxic, non-fusogenic, integral membrane protein and previously unrecognized virion outer-capsid proteins. J. Gen. Virol. 2013, 94, 1039–1050. [Google Scholar] [CrossRef] [PubMed]
- McDonald, S.M.; Nelson, M.I.; Turner, P.E.; Patton, J.T. Reassortment in segmented rna viruses: Mechanisms and outcomes. Nat. Rev. Microbiol. 2016, 14, 448–460. [Google Scholar] [CrossRef]
- Peyambari, M.; Warner, S.; Stoler, N.; Rainer, D.; Roossinck, M.J. A 1000 year-old rna virus. J. Virol. 2018, 93. [Google Scholar] [CrossRef]
- Vijaykrishna, D.; Mukerji, R.; Smith, G.J.D. Rna virus reassortment: An evolutionary mechanism for host jumps and immune evasion. PLoS Pathog. 2015, 11, e1004902. [Google Scholar] [CrossRef]
- Di Cicco, E.; Ferguson, H.W.; Kaukinen, K.H.; Schulze, A.D.; Li, S.; Tabata, A.; Guenther, O.P.; Mordecai, G.; Suttle, C.A.; Miller, K.M. The same strain of piscine orthoreovirus (prv-1) is involved in the development of different, but related, diseases in atlantic and pacific salmon in british columbia. Facets 2018, 3, 599–641. [Google Scholar] [CrossRef]
- Takano, T.; Nawata, A.; Sakai, T.; Matsuyama, T.; Ito, T.; Kurita, J.; Terashima, S.; Yasuike, M.; Nakamura, Y.; Fujiwara, A.; et al. Full-genome sequencing and confirmation of the causative agent of erythrocytic inclusion body syndrome in coho salmon identifies a new type of piscine orthoreovirus. PLoS ONE 2016, 11, e0165424. [Google Scholar] [CrossRef]
- Hauge, H.; Vendramin, N.; Taksdal, T.; Olsen, A.B.; Wessel, O.; Mikkelsen, S.S.; Alencar, A.L.F.; Olesen, N.J.; Dahle, M.K. Infection experiments with novel piscine orthoreovirus from rainbow trout (oncorhynchus mykiss) in salmonids. PLoS ONE 2017, 12, e0180293. [Google Scholar] [CrossRef]
- Kuehn, R.; Stoeckle, B.C.; Young, M.; Popp, L.; Taeubert, J.E.; Pfaffl, M.W.; Geist, J. Identification of a piscine reovirus-related pathogen in proliferative darkening syndrome (pds) infected brown trout (salmo trutta fario) using a next-generation technology detection pipeline. PLoS ONE 2018, 13, e0206164. [Google Scholar] [CrossRef]
- Fux, R.; Arndt, D.; Langenmayer, M.C.; Schwaiger, J.; Ferling, H.; Fischer, N.; Indenbirken, D.; Grundhoff, A.; Dolken, L.; Adamek, M.; et al. Piscine orthoreovirus 3 is not the causative pathogen of proliferative darkening syndrome (pds) of brown trout (salmo trutta fario). Viruses 2019, 11, 112. [Google Scholar] [CrossRef] [PubMed]
- Bohle, H.; Bustos, P.; Leiva, L.; Grothusen, H.; Navas, E.; Sandoval, A.; Bustamante, F.; Montecinos, K.; Gaete, A.; Mancilla, M. First complete genome sequence of piscine orthoreovirus variant 3 infecting coho salmon (oncorhynchus kisutch) farmed in southern chile. Genome Announc. 2018, 6. [Google Scholar] [CrossRef]
- Dhamotharan, K.; Vendramin, N.; Markussen, T.; Wessel, O.; Cuenca, A.; Nyman, I.B.; Olsen, A.B.; Tengs, T.; Krudtaa Dahle, M.; Rimstad, E. Molecular and antigenic characterization of piscine orthoreovirus (prv) from rainbow trout (oncorhynchus mykiss). Viruses 2018, 10, 170. [Google Scholar] [CrossRef]
- Kibenge, M.J.; Iwamoto, T.; Wang, Y.; Morton, A.; Godoy, M.G.; Kibenge, F.S. Whole-genome analysis of piscine reovirus (prv) shows prv represents a new genus in family reoviridae and its genome segment s1 sequences group it into two separate sub-genotypes. Virol. J. 2013, 10, 230. [Google Scholar] [CrossRef]
- Siah, A.; Morrison, D.B.; Fringuelli, E.; Savage, P.; Richmond, Z.; Johns, R.; Purcell, M.K.; Johnson, S.C.; Saksida, S.M. Piscine reovirus: Genomic and molecular phylogenetic analysis from farmed and wild salmonids collected on the canada/us pacific coast. PLoS ONE 2015, 10, e0141475. [Google Scholar] [CrossRef]
- Purcell, M.K.; Powers, R.L.; Evered, J.; Kerwin, J.; Meyers, T.R.; Stewart, B.; Winton, J.R. Molecular testing of adult pacific salmon and trout (oncorhynchus spp.) for several rna viruses demonstrates widespread distribution of piscine orthoreovirus in alaska and washington. J. Fish Dis. 2018, 41, 347–355. [Google Scholar] [CrossRef] [PubMed]
- Tucker, S.; Li, S.R.; Kaukinen, K.H.; Patterson, D.A.; Miller, K.M. Distinct seasonal infectious agent profiles in life-history variants of juvenile fraser river chinook salmon: An application of high-throughput genomic screening. PLoS ONE 2018, 13, e0195472. [Google Scholar] [CrossRef]
- Garseth, A.H.; Fritsvold, C.; Opheim, M.; Skjerve, E.; Biering, E. Piscine reovirus (prv) in wild atlantic salmon, salmo salar l., and sea-trout, salmo trutta l., in norway. J. Fish Dis. 2013, 36, 483–493. [Google Scholar] [CrossRef] [PubMed]
- Garseth, A.H.; Biering, E. Little evidence to suggest salmonid freshwater reservoirs of piscine orthoreovirus (prv). J Fish Dis 2018, 41, 1313–1315. [Google Scholar] [CrossRef] [PubMed]
- Lovoll, M.; Alarcon, M.; Bang Jensen, B.; Taksdal, T.; Kristoffersen, A.B.; Tengs, T. Quantification of piscine reovirus (prv) at different stages of atlantic salmon salmo salar production. Dis. Aquat. Org. 2012, 99, 7–12. [Google Scholar] [CrossRef]
- Godoy, M.G.; Kibenge, M.J.T.; Wang, Y.; Suarez, R.; Leiva, C.; Vallejos, F.; Kibenge, F.S.B. First description of clinical presentation of piscine orthoreovirus (prv) infections in salmonid aquaculture in chile and identification of a second genotype (genotype ii) of prv. Virol. J. 2016, 13, 98. [Google Scholar] [CrossRef] [PubMed]
- Marty, G.D.; Morrison, D.B.; Bidulka, J.; Joseph, T.; Siah, A. Piscine reovirus in wild and farmed salmonids in british columbia, canada: 1974-2013. J. Fish Dis. 2015, 38, 713–728. [Google Scholar] [CrossRef]
- Ferguson, H.; Kongtorp, R.; Taksdal, T.; Graham, D.; Falk, K. An outbreak of disease resembling heart and skeletal muscle inflammation in scottish farmed salmon, salmo salar l., with observations on myocardial regeneration. J. Fish Dis. 2005, 28, 119–123. [Google Scholar] [CrossRef] [PubMed]
- Garver, K.A.; Marty, G.D.; Cockburn, S.N.; Richard, J.; Hawley, L.M.; Muller, A.; Thompson, R.L.; Purcell, M.K.; Saksida, S. Piscine reovirus, but not jaundice syndrome, was transmissible to chinook salmon, oncorhynchus tshawytscha (walbaum), sockeye salmon, oncorhynchus nerka (walbaum), and atlantic salmon, salmo salar L. J. Fish Dis. 2016, 39, 117–128. [Google Scholar] [CrossRef]
- Di Cicco, E.; Ferguson, H.W.; Schulze, A.D.; Kaukinen, K.H.; Li, S.; Vanderstichel, R.; Wessel, O.; Rimstad, E.; Gardner, I.A.; Hammell, K.L.; et al. Heart and skeletal muscle inflammation (hsmi) disease diagnosed on a british columbia salmon farm through a longitudinal farm study. PLoS ONE 2017, 12, e0171471. [Google Scholar] [CrossRef]
- Polinski, M.P.; Marty, G.D.; Snyman, H.N.; Garver, K.A. Piscine orthoreovirus demonstrates high infectivity but low virulence in atlantic salmon of pacific canada. Sci. Rep. 2019, 9, 3297. [Google Scholar] [CrossRef] [PubMed]
- Garver, K.A.; Johnson, S.C.; Polinski, M.P.; Bradshaw, J.C.; Marty, G.D.; Snyman, H.N.; Morrison, D.B.; Richard, J. Piscine orthoreovirus from western north america is transmissible to atlantic salmon and sockeye salmon but fails to cause heart and skeletal muscle inflammation. PLoS ONE 2016, 11, e0146229. [Google Scholar] [CrossRef]
- Finstad, O.W.; Dahle, M.K.; Lindholm, T.H.; Nyman, I.B.; Lovoll, M.; Wallace, C.; Olsen, C.M.; Storset, A.K.; Rimstad, E. Piscine orthoreovirus (prv) infects atlantic salmon erythrocytes. Vet. Res. 2014, 45, 35. [Google Scholar] [CrossRef] [PubMed]
- Lovoll, M.; Wiik-Nielsen, J.; Grove, S.; Wiik-Nielsen, C.R.; Kristoffersen, A.B.; Faller, R.; Poppe, T.; Jung, J.; Pedamallu, C.S.; Nederbragt, A.J.; et al. A novel totivirus and piscine reovirus (prv) in atlantic salmon (salmo salar) with cardiomyopathy syndrome (cms). Virol. J. 2010, 7, 309. [Google Scholar] [CrossRef]
- Li, H.; Durbin, R. Fast and accurate short read alignment with burrows-wheeler transform. Bioinformatics 2009, 25, 1754–1760. [Google Scholar] [CrossRef]
- Koboldt, D.C.; Zhang, Q.; Larson, D.E.; Shen, D.; McLellan, M.D.; Lin, L.; Miller, C.A.; Mardis, E.R.; Ding, L.; Wilson, R.K. Varscan 2: Somatic mutation and copy number alteration discovery in cancer by exome sequencing. Genome Res. 2012, 22, 568–576. [Google Scholar] [CrossRef] [PubMed]
- Hall, T.A. Bioedit: A User-Friendly Biological Sequence Alignment Editor and Analysis Program for Windows 95/98/nt. In Nucleic Acids Symposium Series; Information Retrieval Ltd.: London, UK, 1999; pp. 95–98. [Google Scholar]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. Mega x: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Hillis, D.M.; Bull, J.J. An empirical test of bootstrapping as a method for assessing confidence in phylogenetic analysis. Syst. Biol. 1993, 42, 182–192. [Google Scholar] [CrossRef]
- Jones, D.T. Protein secondary structure prediction based on position-specific scoring matrices. J. Mol. Biol. 1999, 292, 195–202. [Google Scholar] [CrossRef]
- Roy, A.; Kucukural, A.; Zhang, Y. I-tasser: A unified platform for automated protein structure and function prediction. Nat. Protoc. 2010, 5, 725–738. [Google Scholar] [CrossRef]
- Kongtorp, R.T.; Taksdal, T.; Lyngøy, A. Pathology of heart and skeletal muscle inflammation (hsmi) in farmed atlantic salmon salmo salar. Dis. Aquat. Org. 2004, 59, 217–224. [Google Scholar] [CrossRef]
- Wenske, E.A.; Chanock, S.J.; Krata, L.; Fields, B.N. Genetic reassortment of mammalian reoviruses in mice. J. Virol. 1985, 56, 613–616. [Google Scholar] [PubMed]
- Svinti, V.; Cotton, J.A.; McInerney, J.O. New approaches for unravelling reassortment pathways. BMC Evol. Biol. 2013, 13, 1. [Google Scholar] [CrossRef]
- Thete, D.; Danthi, P. Protein mismatches caused by reassortment influence functions of the reovirus capsid. J. Virol. 2018, 92. [Google Scholar] [CrossRef]
- Mabrouk, T.; Lemay, G. The sequence similarity of reovirus sigma-3 protein to picornaviral proteases is unrelated to its role in mu-1 viral protein cleavage. Virology 1994, 202, 615–620. [Google Scholar] [CrossRef]
- Bergeron, J.; Mabrouk, T.; Garzon, S.; Lemay, G. Characterization of the thermosensitive ts453 reovirus mutant: Increased dsrna binding of sigma 3 protein correlates with interferon resistance. Virology 1998, 246, 199–210. [Google Scholar] [CrossRef]
- Imani, F.; Jacobs, B.L. Inhibitory activity for the interferon-induced protein kinase is associated with the reovirus serotype 1 sigma 3 protein. Proc. Natl. Acad. Sci. USA 1988, 85, 7887–7891. [Google Scholar] [CrossRef]
- Mikalsen, A.B.; Haugland, O.; Rode, M.; Solbakk, I.T.; Evensen, O. Atlantic salmon reovirus infection causes a cd8 t cell myocarditis in atlantic salmon (salmo salar l.). PLoS ONE 2012, 7, e37269. [Google Scholar] [CrossRef]
- Dahle, M.K.; Wessel, O.; Timmerhaus, G.; Nyman, I.B.; Jorgensen, S.M.; Rimstad, E.; Krasnov, A. Transcriptome analyses of atlantic salmon (salmo salar l.) erythrocytes infected with piscine orthoreovirus (prv). Fish Shellfish Immunol. 2015, 45, 780–790. [Google Scholar] [CrossRef]
- Madhun, A.S.; Isachsen, C.H.; Omdal, L.M.; Einen, A.C.B.; Maehle, S.; Wennevik, V.; Niemela, E.; Svasand, T.; Karlsbakk, E. Prevalence of piscine orthoreovirus and salmonid alphavirus in sea-caught returning adult atlantic salmon (salmo salar l.) in northern norway. J. Fish Dis. 2018, 41, 797–803. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| PRV Isolate | Associated Disease Status | Tissue Origin of Sequence | Accession Numbers |
|---|---|---|---|
| NOR-1988 | Healthy | Plasma | MK675862–MK675871 |
| NOR-1997 | Unresolved, suspicion of ISA | Plasma | MK675822–MK675831 |
| NOR-2005/TT | CMS | Heart | MK675832–MK675841 |
| NOR-2015/SSK | HSMI | Plasma | MK675852–MK675861 |
| NOR-2015/MS | HSMI-suspected | Plasma | MK675842–MK675851 |
| FO/1978/15 | Healthy | Head kidney | MK675872–MK675881 |
| FO/41/16 | Healthy | Head kidney | MK675882–MK675891 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dhamotharan, K.; Tengs, T.; Wessel, Ø.; Braaen, S.; Nyman, I.B.; Hansen, E.F.; Christiansen, D.H.; Dahle, M.K.; Rimstad, E.; Markussen, T. Evolution of the Piscine orthoreovirus Genome Linked to Emergence of Heart and Skeletal Muscle Inflammation in Farmed Atlantic Salmon (Salmo salar). Viruses 2019, 11, 465. https://doi.org/10.3390/v11050465
Dhamotharan K, Tengs T, Wessel Ø, Braaen S, Nyman IB, Hansen EF, Christiansen DH, Dahle MK, Rimstad E, Markussen T. Evolution of the Piscine orthoreovirus Genome Linked to Emergence of Heart and Skeletal Muscle Inflammation in Farmed Atlantic Salmon (Salmo salar). Viruses. 2019; 11(5):465. https://doi.org/10.3390/v11050465
Chicago/Turabian StyleDhamotharan, Kannimuthu, Torstein Tengs, Øystein Wessel, Stine Braaen, Ingvild B. Nyman, Elisabeth F. Hansen, Debes H. Christiansen, Maria K. Dahle, Espen Rimstad, and Turhan Markussen. 2019. "Evolution of the Piscine orthoreovirus Genome Linked to Emergence of Heart and Skeletal Muscle Inflammation in Farmed Atlantic Salmon (Salmo salar)" Viruses 11, no. 5: 465. https://doi.org/10.3390/v11050465
APA StyleDhamotharan, K., Tengs, T., Wessel, Ø., Braaen, S., Nyman, I. B., Hansen, E. F., Christiansen, D. H., Dahle, M. K., Rimstad, E., & Markussen, T. (2019). Evolution of the Piscine orthoreovirus Genome Linked to Emergence of Heart and Skeletal Muscle Inflammation in Farmed Atlantic Salmon (Salmo salar). Viruses, 11(5), 465. https://doi.org/10.3390/v11050465