Cellular HIV Reservoirs and Viral Rebound from the Lymphoid Compartments of 4′-Ethynyl-2-Fluoro-2′-Deoxyadenosine (EFdA)-Suppressed Humanized Mice

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. NSG-BLT Mice
2.3. Flow Cytometry
2.4. HIV-1 Inoculation and EFdA Treatment
2.5. RNAscope in situ Hybridization and Immunohistochemistry
2.6. Data Analysis
3. Results
3.1. Human T Cells and Macrophages Constitute a Substantial Population of HIV-Susceptible Cells in the Lymphoid Compartments of NSG-BLT Mice
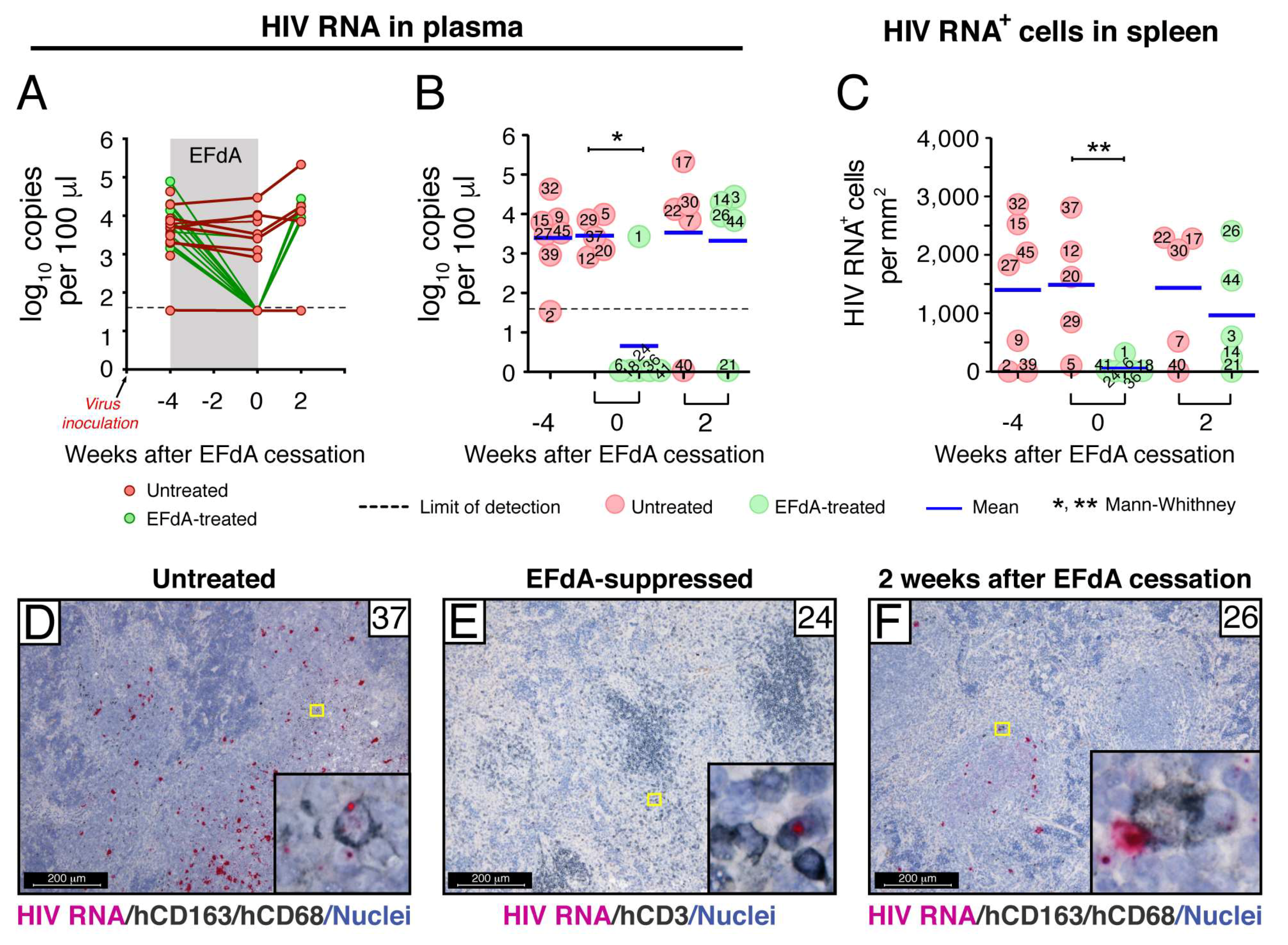
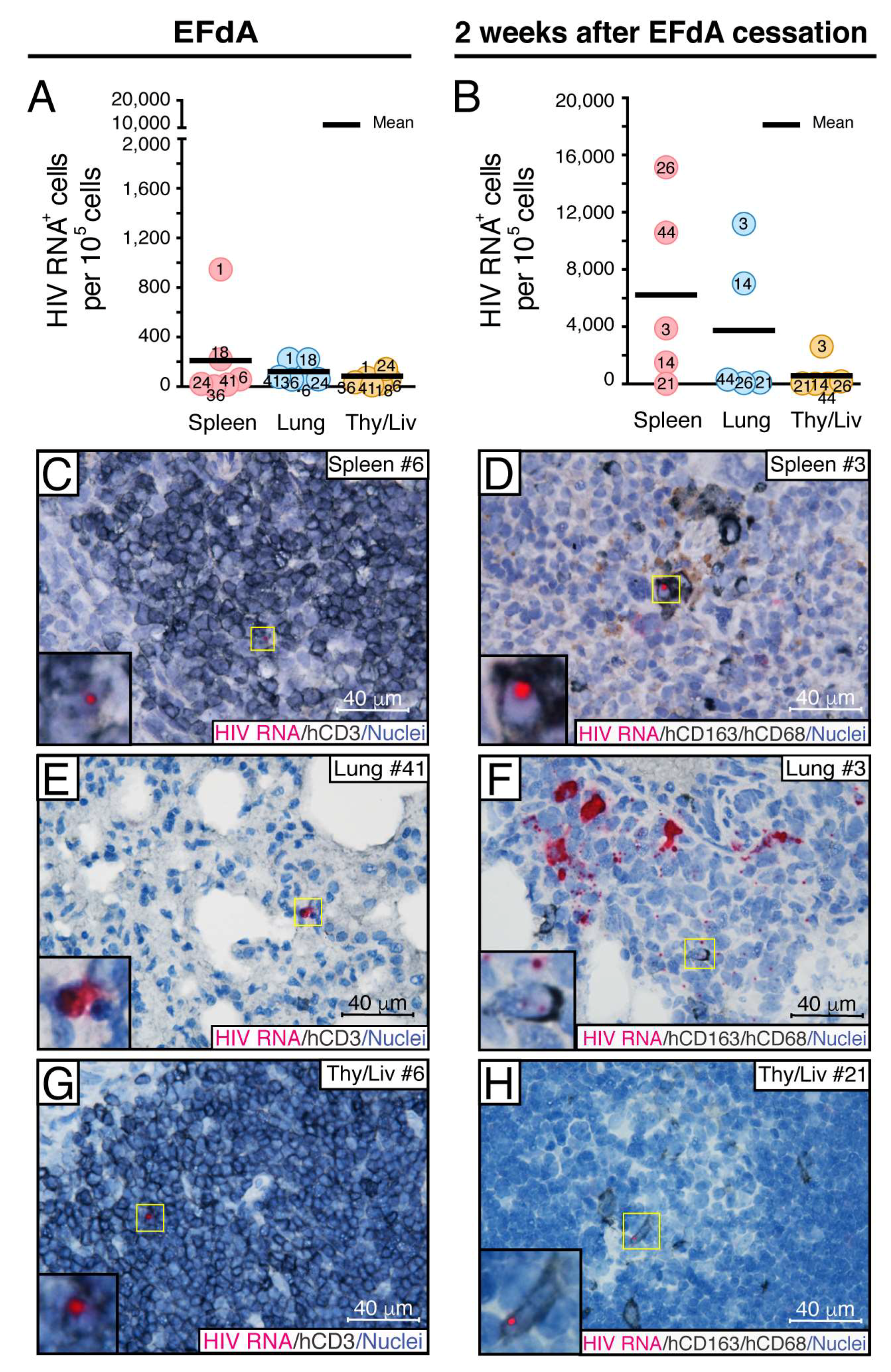
3.2. Fully Suppressive EFdA Treatment Impairs HIV RNA Production in the Lymphoid Tissues of NSG-BLT Mice
3.3. Fully Suppressive EFdA Treatment Does Not Eliminate Cells Harboring HIV DNA
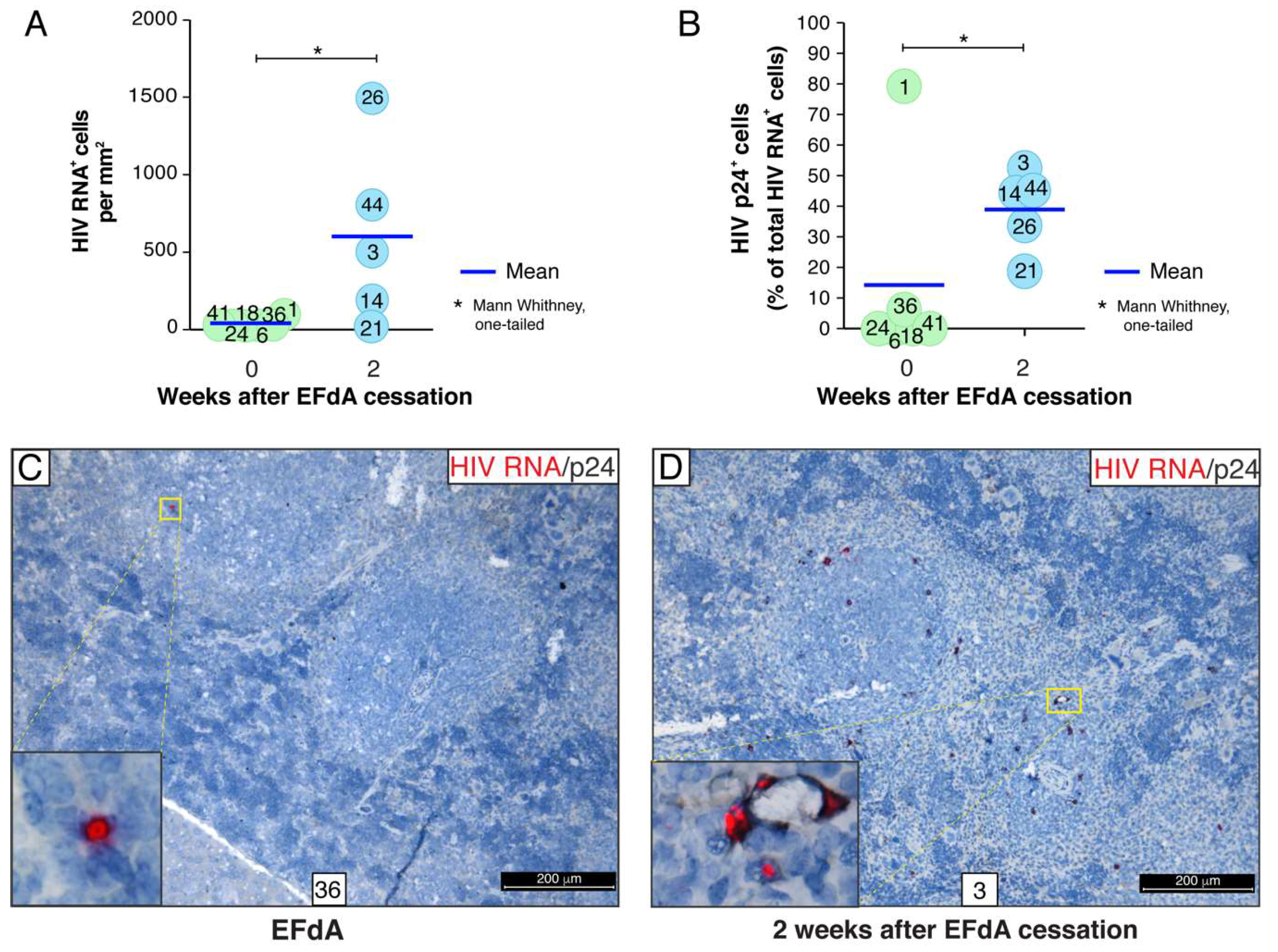
3.4. A Rare Subset of HIV-Infected Cells Expressed HIV RNA and p24 Capsid Protein during Fully Suppressive EFdA Treatment
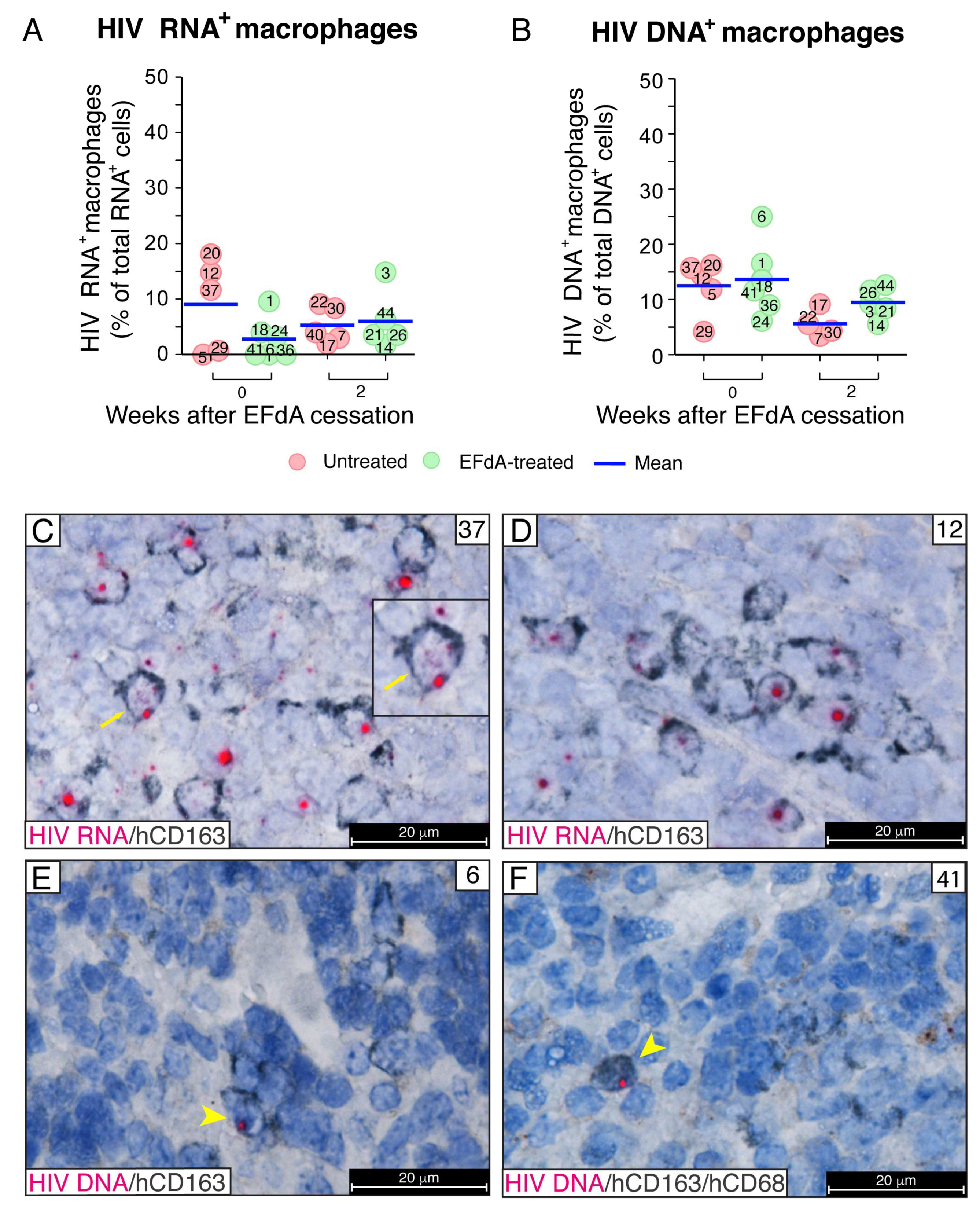
3.5. During Fully Suppressive EFdA Treatment, HIV RNA and Integrated Provirus Were Detected in CD163+ Macrophages
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Johnson, L.F.; Mossong, J.; Dorrington, R.E.; Schomaker, M.; Hoffmann, C.J.; Keiser, O.; Fox, M.P.; Wood, R.; Prozesky, H.; Giddy, J.; et al. Life expectancies of south african adults starting antiretroviral treatment: Collaborative analysis of cohort studies. PLoS Med. 2013, 10, e1001418. [Google Scholar] [CrossRef] [PubMed]
- Nakagawa, F.; May, M.; Phillips, A. Life expectancy living with HIV: Recent estimates and future implications. Curr. Opin. Infect. Dis. 2013, 26, 17–25. [Google Scholar] [CrossRef] [PubMed]
- Lorenzo-Redondo, R.; Fryer, H.R.; Bedford, T.; Kim, E.Y.; Archer, J.; Pond, S.L.K.; Chung, Y.S.; Penugonda, S.; Chipman, J.; Fletcher, C.V.; et al. Persistent HIV-1 replication maintains the tissue reservoir during therapy. Nature 2016, 530, 51–56. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Siliciano, J.D.; Kajdas, J.; Finzi, D.; Quinn, T.C.; Chadwick, K.; Margolick, J.B.; Kovacs, C.; Gange, S.J.; Siliciano, R.F. Long-term follow-up studies confirm the stability of the latent reservoir for HIV-1 in resting CD4+ T cells. Nat. Med. 2003, 9, 727–728. [Google Scholar] [CrossRef] [PubMed]
- Fletcher, C.V.; Staskus, K.; Wietgrefe, S.W.; Rothenberger, M.; Reilly, C.; Chipman, J.G.; Beilman, G.J.; Khoruts, A.; Thorkelson, A.; Schmidt, T.E.; et al. Persistent HIV-1 replication is associated with lower antiretroviral drug concentrations in lymphatic tissues. Proc. Natl. Acad. Sci. USA 2014, 111, 2307–2312. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chomont, N.; El-Far, M.; Ancuta, P.; Trautmann, L.; Procopio, F.A.; Yassine-Diab, B.; Boucher, G.; Boulassel, M.R.; Ghattas, G.; Brenchley, J.M.; et al. HIV reservoir size and persistence are driven by T cell survival and homeostatic proliferation. Nat. Med. 2009, 15, 893–900. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wong, J.K.; Hezareh, M.; Gunthard, H.F.; Havlir, D.V.; Ignacio, C.C.; Spina, C.A.; Richman, D.D. Recovery of replication-competent HIV despite prolonged suppression of plasma viremia. Science 1997, 278, 1291–1295. [Google Scholar] [CrossRef]
- Strain, M.C.; Little, S.J.; Daar, E.S.; Havlir, D.V.; Gunthard, H.F.; Lam, R.Y.; Daly, O.A.; Nguyen, J.; Ignacio, C.C.; Spina, C.A.; et al. Effect of treatment, during primary infection, on establishment and clearance of cellular reservoirs of HIV-1. J. Infect. Dis. 2005, 191, 1410–1418. [Google Scholar] [CrossRef] [PubMed]
- Barton, K.; Winckelmann, A.; Palmer, S. HIV-1 reservoirs during suppressive therapy. Trends Microbiol. 2016, 24, 345–355. [Google Scholar] [CrossRef]
- Schacker, T.; Little, S.; Connick, E.; Gebhard-Mitchell, K.; Zhang, Z.Q.; Krieger, J.; Pryor, J.; Havlir, D.; Wong, J.K.; Richman, D.; et al. Rapid accumulation of human immunodeficiency virus (HIV) in lymphatic tissue reservoirs during acute and early HIV infection: Implications for timing of antiretroviral therapy. J. Infect. Dis. 2000, 181, 354–357. [Google Scholar] [CrossRef]
- Haase, A.T. Population biology of HIV-1 infection: Viral and CD4+ T cell demographics and dynamics in lymphatic tissues. Annu. Rev. Immunol. 1999, 17, 625–656. [Google Scholar] [CrossRef] [PubMed]
- Pantaleo, G.; Graziosi, C.; Butini, L.; Pizzo, P.A.; Schnittman, S.M.; Kotler, D.P.; Fauci, A.S. Lymphoid organs function as major reservoirs for human immunodeficiency virus. Proc. Natl. Acad. Sci. USA 1991, 88, 9838–9842. [Google Scholar] [CrossRef]
- Haase, A.T.; Henry, K.; Zupancic, M.; Sedgewick, G.; Faust, R.A.; Melroe, H.; Cavert, W.; Gebhard, K.; Staskus, K.; Zhang, Z.Q.; et al. Quantitative image analysis of HIV-1 infection in lymphoid tissue. Science 1996, 274, 985–989. [Google Scholar] [CrossRef]
- Cohen, O.J.; Pantaleo, G.; Lam, G.K.; Fauci, A.S. Studies on lymphoid tissue from HIV-infected individuals: Implications for the design of therapeutic strategies. Springer Semin. Immunopathol. 1997, 18, 305–322. [Google Scholar] [CrossRef] [PubMed]
- Sigal, A.; Kim, J.T.; Balazs, A.B.; Dekel, E.; Mayo, A.; Milo, R.; Baltimore, D. Cell-to-cell spread of HIV permits ongoing replication despite antiretroviral therapy. Nature 2011, 477, 95–98. [Google Scholar] [CrossRef] [PubMed]
- Sigal, A.; Baltimore, D. As good as it gets? The problem of HIV persistence despite antiretroviral drugs. Cell Host Microbe 2012, 12, 132–138. [Google Scholar] [CrossRef] [PubMed]
- Hosmane, N.N.; Kwon, K.J.; Bruner, K.M.; Capoferri, A.A.; Beg, S.; Rosenbloom, D.I.; Keele, B.F.; Ho, Y.C.; Siliciano, J.D.; Siliciano, R.F. Proliferation of latently infected CD4(+) T cells carrying replication-competent HIV-1: Potential role in latent reservoir dynamics. J. Exp. Med. 2017, 214, 959–972. [Google Scholar] [CrossRef] [PubMed]
- Kwon, K.J.; Siliciano, R.F. HIV persistence: Clonal expansion of cells in the latent reservoir. J. Clin. Investig. 2017, 127, 2536–2538. [Google Scholar] [CrossRef]
- Cohn, L.B.; Silva, I.T.; Oliveira, T.Y.; Rosales, R.A.; Parrish, E.H.; Learn, G.H.; Hahn, B.H.; Czartoski, J.L.; McElrath, M.J.; Lehmann, C.; et al. HIV-1 integration landscape during latent and active infection. Cell 2015, 160, 420–432. [Google Scholar] [CrossRef] [PubMed]
- Hiener, B.; Horsburgh, B.A.; Eden, J.S.; Barton, K.; Schlub, T.E.; Lee, E.; von Stockenstrom, S.; Odevall, L.; Milush, J.M.; Liegler, T.; et al. Identification of genetically intact HIV-1 proviruses in specific CD4(+) T cells from effectively treated participants. Cell Rep. 2017, 21, 813–822. [Google Scholar] [CrossRef]
- Gorry, P.R.; Francella, N.; Lewin, S.R.; Collman, R.G. HIV-1 envelope-receptor interactions required for macrophage infection and implications for current HIV-1 cure strategies. J. Leukoc. Biol. 2014, 95, 71–81. [Google Scholar] [CrossRef] [PubMed]
- McCune, J.M.; Namikawa, R.; Shih, C.C.; Rabin, L.; Kaneshima, H. Suppression of HIV infection in azt-treated scid-hu mice. Science 1990, 247, 564–566. [Google Scholar] [CrossRef] [PubMed]
- Shultz, L.D.; Ishikawa, F.; Greiner, D.L. Humanized mice in translational biomedical research. Nat. Rev. Immunol. 2007, 7, 118–130. [Google Scholar] [CrossRef]
- Legrand, N.; Weijer, K.; Spits, H. Experimental models to study development and function of the human immune system in vivo. J. Immunol. 2006, 176, 2053–2058. [Google Scholar] [CrossRef] [PubMed]
- Shultz, L.D.; Brehm, M.A.; Garcia-Martinez, J.V.; Greiner, D.L. Humanized mice for immune system investigation: Progress, promise and challenges. Nat. Rev. Immunol. 2012, 12, 786–798. [Google Scholar] [CrossRef] [PubMed]
- Shultz, L.D.; Lyons, B.L.; Burzenski, L.M.; Gott, B.; Chen, X.; Chaleff, S.; Kotb, M.; Gillies, S.D.; King, M.; Mangada, J.; et al. Human lymphoid and myeloid cell development in nod/ltsz-scid il2r gamma null mice engrafted with mobilized human hemopoietic stem cells. J. Immunol. 2005, 174, 6477–6489. [Google Scholar] [CrossRef] [PubMed]
- Honeycutt, J.B.; Wahl, A.; Archin, N.; Choudhary, S.; Margolis, D.; Garcia, J.V. HIV-1 infection, response to treatment and establishment of viral latency in a novel humanized T cell-only mouse (tom) model. Retrovirology 2013, 10, 121. [Google Scholar] [CrossRef]
- Lan, P.; Tonomura, N.; Shimizu, A.; Wang, S.; Yang, Y.G. Reconstitution of a functional human immune system in immunodeficient mice through combined human fetal thymus/liver and cd34+ cell transplantation. Blood 2006, 108, 487–492. [Google Scholar] [CrossRef]
- Melkus, M.W.; Estes, J.D.; Padgett-Thomas, A.; Gatlin, J.; Denton, P.W.; Othieno, F.A.; Wege, A.K.; Haase, A.T.; Garcia, J.V. Humanized mice mount specific adaptive and innate immune responses to ebv and tsst-1. Nat. Med. 2006, 12, 1316–1322. [Google Scholar] [CrossRef] [PubMed]
- Honeycutt, J.B.; Wahl, A.; Baker, C.; Spagnuolo, R.A.; Foster, J.; Zakharova, O.; Wietgrefe, S.; Caro-Vegas, C.; Madden, V.; Sharpe, G.; et al. Macrophages sustain HIV replication in vivo independently of T cells. J. Clin. Investig. 2016, 126, 1353–1366. [Google Scholar] [CrossRef] [Green Version]
- Nixon, C.C.; Mavigner, M.; Silvestri, G.; Garcia, J.V. In vivo models of human immunodeficiency virus persistence and cure strategies. J. Infect. Dis. 2017, 215, S142–S151. [Google Scholar] [CrossRef] [PubMed]
- Stoddart, C.A.; Galkina, S.A.; Joshi, P.; Kosikova, G.; Moreno, M.E.; Rivera, J.M.; Sloan, B.; Reeve, A.B.; Sarafianos, S.G.; Murphey-Corb, M.; et al. Oral administration of the nucleoside efda (4′-ethynyl-2-fluoro-2′-deoxyadenosine) provides rapid suppression of HIV viremia in humanized mice and favorable pharmacokinetic properties in mice and the rhesus macaque. Antimicrob. Agents Chemother. 2015, 59, 4190–4198. [Google Scholar] [CrossRef]
- Tieu, H.V.; Taylor, B.S.; Jones, J.; Wilkin, T.J. Croi 2018: Advances in antiretroviral therapy. Top. Antivir. Med. 2018, 26, 40–53. [Google Scholar]
- Markowitz, M.; Sarafianos, S.G. 4′-ethynyl-2-fluoro-2′-deoxyadenosine, mk-8591: A novel HIV-1 reverse transcriptase translocation inhibitor. Curr. Opin. HIV AIDS 2018, 13, 294–299. [Google Scholar] [CrossRef]
- Stoddart, C.A.; Maidji, E.; Galkina, S.A.; Kosikova, G.; Rivera, J.M.; Moreno, M.E.; Sloan, B.; Joshi, P.; Long, B.R. Superior human leukocyte reconstitution and susceptibility to vaginal HIV transmission in humanized nod-scid il-2rγ(-/-) (nsg) blt mice. Virology 2011, 417, 154–160. [Google Scholar] [CrossRef] [PubMed]
- Haltiner, M.; Kempe, T.; Tjian, R. A novel strategy for constructing clustered point mutations. Nucleic Acids Res. 1985, 13, 1015–1025. [Google Scholar] [CrossRef] [Green Version]
- Koyanagi, Y.; Miles, S.; Mitsuyasu, R.T.; Merrill, J.E.; Vinters, H.V.; Chen, I.S. Dual infection of the central nervous system by aids viruses with distincT cellular tropisms. Science 1987, 236, 819–822. [Google Scholar] [CrossRef]
- Cann, A.J.; Zack, J.A.; Go, A.S.; Arrigo, S.J.; Koyanagi, Y.; Green, P.L.; Koyanagi, Y.; Pang, S.; Chen, I.S. Human immunodeficiency virus type 1 T-cell tropism is determined by events prior to provirus formation. J. Virol. 1990, 64, 4735–4742. [Google Scholar] [PubMed]
- Joshi, P.; Maidji, E.; Stoddart, C.A. Inhibition of heat shock protein 90 prevents HIV rebound. J. Biol. Chem. 2016, 291, 10332–10346. [Google Scholar] [CrossRef]
- Deleage, C.; Wietgrefe, S.W.; Del Prete, G.; Morcock, D.R.; Hao, X.P.; Piatak, M., Jr.; Bess, J.; Anderson, J.L.; Perkey, K.E.; Reilly, C.; et al. Defining HIV and siv reservoirs in lymphoid tissues. Pathog. Immun. 2016, 1, 68–106. [Google Scholar] [CrossRef]
- Leal, S.; Diniz, C.; Sa, C.; Goncalves, J.; Soares, A.S.; Rocha-Pereira, C.; Fresco, P. Semiautomated computer-assisted image analysis to quantify 3,3′-diaminobenzidine tetrahydrochloride-immunostained small tissues. Anal. Biochem. 2006, 357, 137–143. [Google Scholar] [CrossRef] [PubMed]
- Rosok, B.; Brinchmann, J.E.; Voltersvik, P.; Olofsson, J.; Bostad, L.; Asjo, B. Correlates of latent and productive HIV type-1 infection in tonsillar CD4(+) T cells. Proc. Natl. Acad. Sci. USA 1997, 94, 9332–9336. [Google Scholar] [CrossRef] [PubMed]
- Chun, T.W.; Nickle, D.C.; Justement, J.S.; Meyers, J.H.; Roby, G.; Hallahan, C.W.; Kottilil, S.; Moir, S.; Mican, J.M.; Mullins, J.I.; et al. Persistence of HIV in gut-associated lymphoid tissue despite long-term antiretroviral therapy. J. Infect. Dis. 2008, 197, 714–720. [Google Scholar] [CrossRef] [PubMed]
- Yukl, S.A.; Gianella, S.; Sinclair, E.; Epling, L.; Li, Q.; Duan, L.; Choi, A.L.; Girling, V.; Ho, T.; Li, P.; et al. Differences in HIV burden and immune activation within the gut of HIV-positive patients receiving suppressive antiretroviral therapy. J. Infect. Dis. 2010, 202, 1553–1561. [Google Scholar] [CrossRef] [PubMed]
- Gunthard, H.F.; Havlir, D.V.; Fiscus, S.; Zhang, Z.Q.; Eron, J.; Mellors, J.; Gulick, R.; Frost, S.D.; Brown, A.J.; Schleif, W.; et al. Residual human immunodeficiency virus (HIV) type 1 rna and DNA in lymph nodes and HIV rna in genital secretions and in cerebrospinal fluid after suppression of viremia for 2 years. J. Infect. Dis. 2001, 183, 1318–1327. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Flanagan, J.; Su, N.; Wang, L.C.; Bui, S.; Nielson, A.; Wu, X.; Vo, H.T.; Ma, X.J.; Luo, Y. Rnascope: A novel in situ rna analysis platform for formalin-fixed, paraffin-embedded tissues. J. Mol. Diagn. 2012, 14, 22–29. [Google Scholar] [CrossRef]
- Pabst, R.; Gehrke, I. Is the bronchus-associated lymphoid tissue (balt) an integral structure of the lung in normal mammals, including humans? Am. J. Respir. Cell Mol. Biol. 1990, 3, 131–135. [Google Scholar] [CrossRef] [PubMed]
- Maidji, E.; Somsouk, M.; Rivera, J.M.; Hunt, P.W.; Stoddart, C.A. Replication of cmv in the gut of HIV-infected individuals and epithelial barrier dysfunction. PLoS Pathog. 2017, 13, e1006202. [Google Scholar] [CrossRef] [PubMed]
- Bianconi, E.; Piovesan, A.; Facchin, F.; Beraudi, A.; Casadei, R.; Frabetti, F.; Vitale, L.; Pelleri, M.C.; Tassani, S.; Piva, F.; et al. An estimation of the number of cells in the human body. Ann. Hum. Biol. 2013, 40, 463–471. [Google Scholar] [CrossRef]
- Michailidis, E.; Marchand, B.; Kodama, E.N.; Singh, K.; Matsuoka, M.; Kirby, K.A.; Ryan, E.M.; Sawani, A.M.; Nagy, E.; Ashida, N.; et al. Mechanism of inhibition of HIV-1 reverse transcriptase by 4′-ethynyl-2-fluoro-2′-deoxyadenosine triphosphate, a translocation-defective reverse transcriptase inhibitor. J. Biol. Chem. 2009, 284, 35681–35691. [Google Scholar] [CrossRef] [PubMed]
- Geijtenbeek, T.B.; Kwon, D.S.; Torensma, R.; van Vliet, S.J.; van Duijnhoven, G.C.; Middel, J.; Cornelissen, I.L.; Nottet, H.S.; KewalRamani, V.N.; Littman, D.R.; et al. Dc-sign, a dendritic cell-specific HIV-1-binding protein that enhances trans-infection of T cells. Cell 2000, 100, 587–597. [Google Scholar] [CrossRef]
- DeLucia, D.C.; Rinaldo, C.R.; Rappocciolo, G. Inefficient HIV-1 trans infection of CD4(+) T cells by macrophages from HIV-1 nonprogressors is associated with altered membrane cholesterol and dc-sign. J. Virol. 2018, 92, e00092-18. [Google Scholar] [CrossRef] [PubMed]
- Geijtenbeek, T.B.; Torensma, R.; van Vliet, S.J.; van Duijnhoven, G.C.; Adema, G.J.; van Kooyk, Y.; Figdor, C.G. Identification of dc-sign, a novel dendritic cell-specific icam-3 receptor that supports primary immune responses. Cell 2000, 100, 575–585. [Google Scholar] [CrossRef]
- Real, F.; Sennepin, A.; Ganor, Y.; Schmitt, A.; Bomsel, M. Live imaging of HIV-1 transfer across T cell virological synapse to epithelial cells that promotes stromal macrophage infection. Cell Rep. 2018, 23, 1794–1805. [Google Scholar] [CrossRef] [PubMed]
- Piguet, V.; Sattentau, Q. Dangerous liaisons at the virological synapse. J. Clin. Investig. 2004, 114, 605–610. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feldmann, J.; Schwartz, O. HIV-1 virological synapse: Live imaging of transmission. Viruses 2010, 2, 1666–1680. [Google Scholar] [CrossRef] [PubMed]
- Arrighi, J.F.; Pion, M.; Garcia, E.; Escola, J.M.; van Kooyk, Y.; Geijtenbeek, T.B.; Piguet, V. Dc-sign-mediated infectious synapse formation enhances x4 HIV-1 transmission from dendritic cells to T cells. J. Exp. Med. 2004, 200, 1279–1288. [Google Scholar] [CrossRef]
- Denton, P.W.; Estes, J.D.; Sun, Z.; Othieno, F.A.; Wei, B.L.; Wege, A.K.; Powell, D.A.; Payne, D.; Haase, A.T.; Garcia, J.V. Antiretroviral pre-exposure prophylaxis prevents vaginal transmission of HIV-1 in humanized blt mice. PLoS Med. 2008, 5, e16. [Google Scholar] [CrossRef] [PubMed]
- Brainard, D.M.; Seung, E.; Frahm, N.; Cariappa, A.; Bailey, C.C.; Hart, W.K.; Shin, H.S.; Brooks, S.F.; Knight, H.L.; Eichbaum, Q.; et al. Induction of robusT cellular and humoral virus-specific adaptive immune responses in human immunodeficiency virus-infected humanized blt mice. J. Virol. 2009, 83, 7305–7321. [Google Scholar] [CrossRef]
- Denton, P.W.; Krisko, J.F.; Powell, D.A.; Mathias, M.; Kwak, Y.T.; Martinez-Torres, F.; Zou, W.; Payne, D.A.; Estes, J.D.; Garcia, J.V. Systemic administration of antiretrovirals prior to exposure prevents rectal and intravenous HIV-1 transmission in humanized blt mice. PLoS ONE 2010, 5, e8829. [Google Scholar] [CrossRef] [PubMed]
- Shimizu, S.; Hong, P.; Arumugam, B.; Pokomo, L.; Boyer, J.; Koizumi, N.; Kittipongdaja, P.; Chen, A.; Bristol, G.; Galic, Z.; et al. A highly efficient short hairpin rna potently down-regulates ccr5 expression in systemic lymphoid organs in the hu-blt mouse model. Blood 2010, 115, 1534–1544. [Google Scholar] [CrossRef] [PubMed]
- Denton, P.W.; Olesen, R.; Choudhary, S.K.; Archin, N.M.; Wahl, A.; Swanson, M.D.; Chateau, M.; Nochi, T.; Krisko, J.F.; Spagnuolo, R.A.; et al. Generation of HIV latency in humanized blt mice. J. Virol. 2012, 86, 630–634. [Google Scholar] [CrossRef] [PubMed]
- Marsden, M.D.; Kovochich, M.; Suree, N.; Shimizu, S.; Mehta, R.; Cortado, R.; Bristol, G.; An, D.S.; Zack, J.A. HIV latency in the humanized blt mouse. J. Virol. 2012, 86, 339–347. [Google Scholar] [CrossRef]
- Long, B.R.; Stoddart, C.A. Alpha interferon and HIV infection cause activation of human T cells in nsg-blt mice. J. Virol. 2012, 86, 3327–3336. [Google Scholar] [CrossRef]
- Chateau, M.L.; Denton, P.W.; Swanson, M.D.; McGowan, I.; Garcia, J.V. Rectal transmission of transmitted/founder HIV-1 is efficiently prevented by topical 1% tenofovir in blt humanized mice. PLoS ONE 2013, 8, e60024. [Google Scholar] [CrossRef] [PubMed]
- Stoddart, C.A.; Galkina, S.A.; Joshi, P.; Kosikova, G.; Long, B.R.; Maidji, E.; Moreno, M.E.; Rivera, J.M.; Sanford, U.R.; Sloan, B.; et al. Efficacy of broadly neutralizing monoclonal antibody pg16 in HIV-infected humanized mice. Virology 2014, 462–463, 115–125. [Google Scholar] [CrossRef]
- Veselinovic, M.; Charlins, P.; Akkina, R. Modeling HIV-1 mucosal transmission and prevention in humanized mice. Methods Mol. Biol. 2016, 1354, 203–220. [Google Scholar]
- Shanmugasundaram, U.; Kovarova, M.; Ho, P.T.; Schramm, N.; Wahl, A.; Parniak, M.A.; Garcia, J.V. Efficient inhibition of HIV replication in the gastrointestinal and female reproductive tracts of humanized blt mice by efda. PLoS ONE 2016, 11, e0159517. [Google Scholar] [CrossRef] [PubMed]
- Honeycutt, J.B.; Liao, B.; Nixon, C.C.; Cleary, R.A.; Thayer, W.O.; Birath, S.L.; Swanson, M.D.; Sheridan, P.; Zakharova, O.; Prince, F.; et al. T cells establish and maintain cns viral infection in HIV-infected humanized mice. J. Clin. Investig. 2018, 128, 2862–2876. [Google Scholar] [CrossRef] [PubMed]
- Ganusov, V.V.; De Boer, R.J. Do most lymphocytes in humans really reside in the gut? Trends Immunol. 2007, 28, 514–518. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pabst, R.; Tschernig, T. Bronchus-associated lymphoid tissue: An entry site for antigens for successful mucosal vaccinations? Am. J. Respir. Cell Mol. Biol. 2010, 43, 137–141. [Google Scholar] [CrossRef] [PubMed]
- Bienenstock, J.; Johnston, N.; Perey, D.Y. Bronchial lymphoid tissue. Ii. Functional characterisitics. Lab. Investig. 1973, 28, 693–698. [Google Scholar] [PubMed]
- Tschernig, T.; Pabst, R. Bronchus-associated lymphoid tissue (balt) is not present in the normal adult lung but in different diseases. Pathobiology 2000, 68, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Chen, P.; Hubner, W.; Spinelli, M.A.; Chen, B.K. Predominant mode of human immunodeficiency virus transfer between T cells is mediated by sustained env-dependent neutralization-resistant virological synapses. J. Virol. 2007, 81, 12582–12595. [Google Scholar] [CrossRef] [PubMed]
- McDonald, D.; Wu, L.; Bohks, S.M.; KewalRamani, V.N.; Unutmaz, D.; Hope, T.J. Recruitment of HIV and its receptors to dendritic cell-T cell junctions. Science 2003, 300, 1295–1297. [Google Scholar] [CrossRef] [PubMed]
- Rothenberger, M.K.; Keele, B.F.; Wietgrefe, S.W.; Fletcher, C.V.; Beilman, G.J.; Chipman, J.G.; Khoruts, A.; Estes, J.D.; Anderson, J.; Callisto, S.P.; et al. Large number of rebounding/founder HIV variants emerge from multifocal infection in lymphatic tissues after treatment interruption. Proc. Natl. Acad. Sci. USA 2015, 112, E1126–E1134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jung, A.; Maier, R.; Vartanian, J.P.; Bocharov, G.; Jung, V.; Fischer, U.; Meese, E.; Wain-Hobson, S.; Meyerhans, A. Recombination: Multiply infected spleen cells in HIV patients. Nature 2002, 418, 144. [Google Scholar] [CrossRef]
- Estes, J.D.; Kityo, C.; Ssali, F.; Swainson, L.; Makamdop, K.N.; Del Prete, G.Q.; Deeks, S.G.; Luciw, P.A.; Chipman, J.G.; Beilman, G.J.; et al. Defining total-body aids-virus burden with implications for curative strategies. Nat. Med. 2017, 23, 1271–1276. [Google Scholar] [CrossRef] [PubMed]
- Baxter, A.E.; Niessl, J.; Fromentin, R.; Richard, J.; Porichis, F.; Charlebois, R.; Massanella, M.; Brassard, N.; Alsahafi, N.; Delgado, G.G.; et al. Single-cell characterization of viral translation-competent reservoirs in HIV-infected individuals. Cell Host Microbe 2016, 20, 368–380. [Google Scholar] [CrossRef] [PubMed]
- Imamichi, H.; Natarajan, V.; Adelsberger, J.W.; Rehm, C.A.; Lempicki, R.A.; Das, B.; Hazen, A.; Imamichi, T.; Lane, H.C. Lifespan of effector memory CD4+ T cells determined by replication-incompetent integrated HIV-1 provirus. AIDS 2014, 28, 1091–1099. [Google Scholar] [CrossRef] [Green Version]
- Pollack, R.A.; Jones, R.B.; Pertea, M.; Bruner, K.M.; Martin, A.R.; Thomas, A.S.; Capoferri, A.A.; Beg, S.A.; Huang, S.H.; Karandish, S.; et al. Defective HIV-1 proviruses are expressed and can be recognized by cytotoxic t lymphocytes, which shape the proviral landscape. Cell Host Microbe 2017, 21, 494–506.e4. [Google Scholar] [CrossRef]
- Baxter, A.E.; O’Doherty, U.; Kaufmann, D.E. Beyond the replication-competent HIV reservoir: Transcription and translation-competent reservoirs. Retrovirology 2018, 15, 18. [Google Scholar] [CrossRef] [PubMed]
- Baxter, A.E.; Niessl, J.; Fromentin, R.; Richard, J.; Porichis, F.; Massanella, M.; Brassard, N.; Alsahafi, N.; Routy, J.P.; Finzi, A.; et al. Multiparametric characterization of rare HIV-infected cells using an rna-flow fish technique. Nat. Protoc. 2017, 12, 2029–2049. [Google Scholar] [CrossRef] [PubMed]
- Koenig, S.; Gendelman, H.E.; Orenstein, J.M.; Dal Canto, M.C.; Pezeshkpour, G.H.; Yungbluth, M.; Janotta, F.; Aksamit, A.; Martin, M.A.; Fauci, A.S. Detection of aids virus in macrophages in brain tissue from aids patients with encephalopathy. Science 1986, 233, 1089–1093. [Google Scholar] [CrossRef] [PubMed]
- Brown, D.; Mattapallil, J.J. Gastrointestinal tract and the mucosal macrophage reservoir in HIV infection. Clin. Vaccine Immunol. 2014, 21, 1469–1473. [Google Scholar] [CrossRef] [PubMed]
- Zalar, A.; Figueroa, M.I.; Ruibal-Ares, B.; Bare, P.; Cahn, P.; de Bracco, M.M.; Belmonte, L. Macrophage HIV-1 infection in duodenal tissue of patients on long term haart. Antivir. Res. 2010, 87, 269–271. [Google Scholar] [CrossRef] [PubMed]
- Arainga, M.; Su, H.; Poluektova, L.Y.; Gorantla, S.; Gendelman, H.E. HIV-1 cellular and tissue replication patterns in infected humanized mice. Sci. Rep. 2016, 6, 23513. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Honeycutt, J.B.; Thayer, W.O.; Baker, C.E.; Ribeiro, R.M.; Lada, S.M.; Cao, Y.; Cleary, R.A.; Hudgens, M.G.; Richman, D.D.; Garcia, J.V. HIV persistence in tissue macrophages of humanized myeloid-only mice during antiretroviral therapy. Nat. Med. 2017, 23, 638–643. [Google Scholar] [CrossRef] [PubMed]
- Williams, D.W.; Engle, E.L.; Shirk, E.N.; Queen, S.E.; Gama, L.; Mankowski, J.L.; Zink, M.C.; Clements, J.E. Splenic damage during siv infection: Role of t-cell depletion and macrophage polarization and infection. Am. J. Pathol. 2016, 186, 2068–2087. [Google Scholar] [CrossRef]
- Baxter, A.E.; Russell, R.A.; Duncan, C.J.; Moore, M.D.; Willberg, C.B.; Pablos, J.L.; Finzi, A.; Kaufmann, D.E.; Ochsenbauer, C.; Kappes, J.C.; et al. Macrophage infection via selective capture of HIV-1-infected CD4+ T cells. Cell Host Microbe 2014, 16, 711–721. [Google Scholar] [CrossRef] [PubMed]
- Bracq, L.; Xie, M.; Lambele, M.; Vu, L.T.; Matz, J.; Schmitt, A.; Delon, J.; Zhou, P.; Randriamampita, C.; Bouchet, J.; et al. T cell-macrophage fusion triggers multinucleated gianT cell formation for HIV-1 spreading. J. Virol. 2017. [Google Scholar] [CrossRef] [PubMed]
- McDonald, D. Dendritic cells and HIV-1 trans-infection. Viruses 2010, 2, 1704–1717. [Google Scholar] [CrossRef]
- Wang, L.; Eng, E.T.; Law, K.; Gordon, R.E.; Rice, W.J.; Chen, B.K. Visualization of HIV T cell virological synapses and virus-containing compartments by three-dimensional correlative light and electron microscopy. J. Virol. 2017, 91. [Google Scholar] [CrossRef] [PubMed]
- Jolly, C.; Kashefi, K.; Hollinshead, M.; Sattentau, Q.J. HIV-1 cell to cell transfer across an env-induced, actin-dependent synapse. J. Exp. Med. 2004, 199, 283–293. [Google Scholar] [CrossRef] [PubMed]









© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Maidji, E.; Moreno, M.E.; Rivera, J.M.; Joshi, P.; Galkina, S.A.; Kosikova, G.; Somsouk, M.; Stoddart, C.A. Cellular HIV Reservoirs and Viral Rebound from the Lymphoid Compartments of 4′-Ethynyl-2-Fluoro-2′-Deoxyadenosine (EFdA)-Suppressed Humanized Mice. Viruses 2019, 11, 256. https://doi.org/10.3390/v11030256
Maidji E, Moreno ME, Rivera JM, Joshi P, Galkina SA, Kosikova G, Somsouk M, Stoddart CA. Cellular HIV Reservoirs and Viral Rebound from the Lymphoid Compartments of 4′-Ethynyl-2-Fluoro-2′-Deoxyadenosine (EFdA)-Suppressed Humanized Mice. Viruses. 2019; 11(3):256. https://doi.org/10.3390/v11030256
Chicago/Turabian StyleMaidji, Ekaterina, Mary E. Moreno, Jose M. Rivera, Pheroze Joshi, Sofiya A. Galkina, Galina Kosikova, Ma Somsouk, and Cheryl A. Stoddart. 2019. "Cellular HIV Reservoirs and Viral Rebound from the Lymphoid Compartments of 4′-Ethynyl-2-Fluoro-2′-Deoxyadenosine (EFdA)-Suppressed Humanized Mice" Viruses 11, no. 3: 256. https://doi.org/10.3390/v11030256