Absence of A3Z3-Related Hypermutations in the env and vif Proviral Genes in FIV Naturally Infected Cats

 , , and
, , and
Abstract
1. Introduction
2. Materials and Methods
2.1. Samples and DNA Extraction
2.2. A3Z3, env and vif Amplifications and Sequencing
2.3. Genotype and Haplotype Analyses
2.4. Hypermutations Analyses
3. Results
3.1. PCR Amplification and Sequencing
3.2. Identification of A3Z3 Haplotypes
3.3. Detection of Hypermutations
4. Discussion
Author Contributions
Funding
Acknowledgments
Appendix A

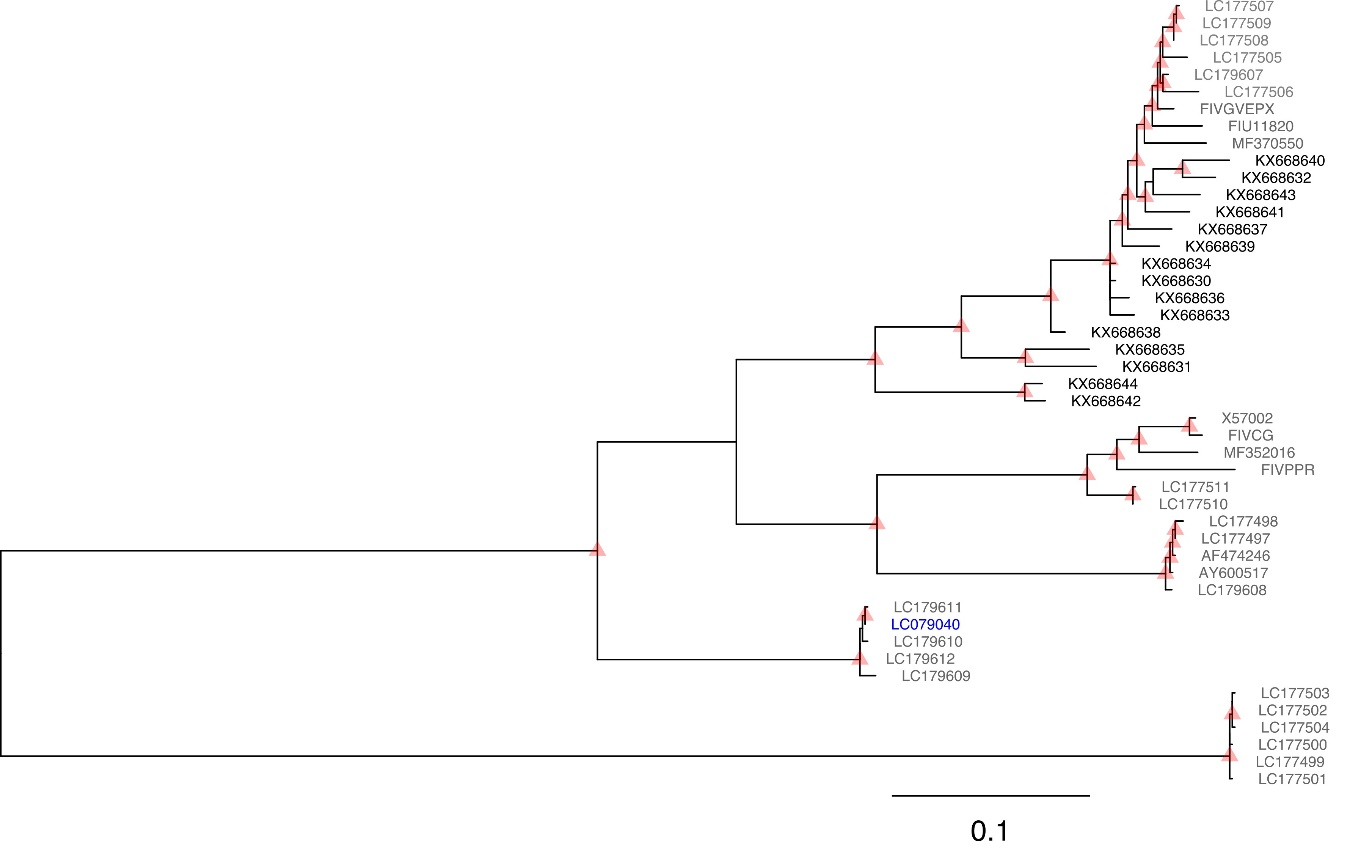{kind=link}
| Region | Primer Name | Sequence (5′–3′) | Size |
|---|---|---|---|
| Exon 3 A3Z3 | A3H2F | TCATCCCCAATGGCACCCACAGC | 590 pb |
| A3H3R | TCAAACTCTGAGACGGAGGAGGAG | ||
| External primers | VIF_FIV_PF | CTTCCTGAAGGGGATGAGTG | 3.1 kb |
| ENV_PR | CCTARTTCTTGCATAGCRAAAGC | ||
| Internal primers vif | VIF_FIV_PF | CTTCCTGAAGGGGATGAGTG | 756 pb |
| VIF_FIV_PR | ATCTCTTCCATTCATAGYTCTCC | ||
| Internal primers env | ENV2-3_PF | GAATGAGACTATAACAGGAC | 831 pb |
| ENV2-3_PR | CAAGACCAATTTCCAGCAAT |

References
- Pecon-Slattery, J.; Troyer, J.L.; Johnson, W.E.; O’Brien, S.J. Evolution of feline immunodeficiency virus in Felidae: Implications for human health and wildlife ecology. Vet. Immunol. Immunopathol. 2008, 123, 32–44. [Google Scholar] [CrossRef] [PubMed]
- Troyer, R.M.; Thompson, J.; Elder, J.H.; Vandewoude, S. Accessory Genes Confer a High Replication Rate to Virulent Feline. J. Virol. 2013, 87, 7940–7951. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Kenyon, J.C.; Lever, A.M.L. The molecular biology of feline immunodeficiency virus (FIV). Viruses 2011, 3, 2192–2213. [Google Scholar] [CrossRef] [PubMed]
- Tomonagat, K.; Mikami, T. Molecular biology of the feline immunodeficiency virus auxiliary genes. J. Gen. Virol. 1996, 77, 1611–1621. [Google Scholar] [CrossRef] [PubMed]
- Gabuzda, D.H.; Lawrence, K.; Langhoff, E.; Terwilliger, E.; Dorfman, T.; Haseltine, W.A.; Sodroski, J. Role of vif in Replication of Human Immunodeficiency Virus Type 1 in CD4+ T Lymphocytes. J. Virol. 1992, 66, 6489–6495. [Google Scholar] [PubMed]
- Sova, P.; Volsky, D.J. Efficiency of Viral DNA Synthesis during Infection of Permissive and Nonpermissive Cells with vif-Negative Human Immunodeficiency Virus Type 1 Cp. J. Virol. 1993, 67, 6322–6326. [Google Scholar] [PubMed]
- Harris, R.S.; Liddament, M.T. Retroviral restriction by APOBEC proteins. Nat. Rev. Immunol. 2004, 4, 868–877. [Google Scholar] [CrossRef] [PubMed]
- Münk, C.; Beck, T.; Zielonka, J.; Hotz-Wagenblatt, A.; Chareza, S.; Battenberg, M.; Thielebein, J.; Cichutek, K.; Bravo, I.G.; O’Brien, S.J.; et al. Functions, structure, and read-through alternative splicing of feline APOBEC3 genes. Genome Biol. 2008, 9, R48. [Google Scholar] [CrossRef] [PubMed]
- Cen, S.; Guo, F.; Niu, M.; Saadatmand, J.; Deflassieux, J.; Kleiman, L. The Interaction between HIV-1 Gag and APOBEC3G. J. Biol. Chem. 2004, 279, 33177–33184. [Google Scholar] [CrossRef] [PubMed]
- Luo, K.; Liu, B.; Xiao, Z.; Yu, Y.; Yu, X.; Gorelick, R.; Yu, X. Amino-Terminal Region of the Human Immunodeficiency Virus Type 1 Nucleocapsid Is Required for Human APOBEC3G Packaging. J. Virol. 2004, 78, 11841–11852. [Google Scholar] [CrossRef] [PubMed]
- Wedekind, J.E.; Dance, G.S.C.; Sowden, M.P.; Smith, H.C. Messenger RNA editing in mammals: New members of the APOBEC family seeking roles in the family business. Trends Genet. 2003, 19, 207–216. [Google Scholar] [CrossRef]
- Mangeat, B.; Turelli, P.; Caron, G.; Friedli, M. Broad antiretroviral defence by human APOBEC3G through lethal editing of nascent reverse transcripts. Nature 2003, 424, 99–103. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Yang, B.; Pomerantz, R.J.; Zhang, C.; Arunachalam, S.C.; Gao, L. The cytidine deaminase CEM15 induces hypermutation in newly synthesized HIV-1 DNA. Nature 2003, 424, 94–98. [Google Scholar] [CrossRef] [PubMed]
- Conticello, S.G.; Harris, R.S.; Neuberger, M.S. The Vif Protein of HIV Triggers Degradation of the Human Antiretroviral DNA Deaminase APOBEC3G. Curr. Biol. 2003, 13, 2009–2013. [Google Scholar] [CrossRef] [PubMed]
- Yu, X.; Yu, Y.; Liu, B.; Luo, K.; Kong, W.; Mao, P.; Yu, X.-F. Induction of APOBEC3G Ubiquitination and Degradation by an HIV-1 Vif-Cul5-SCF Complex. Science 2003, 1056, 3–8. [Google Scholar] [CrossRef] [PubMed]
- Malim, M.H.; Bieniasz, P.D. HIV restriction factors and mechanisms of evasion. Cold Spring Harb. Perspect. Med. 2012, 2, a006940. [Google Scholar] [CrossRef] [PubMed]
- Jäger, S.; Kim, D.Y.; Hultquist, J.F.; Shindo, K.; LaRue, R.S.; Kane, J.; Franks-skiba, K.; Cimermancic, P.; Burlingame, A.; Sali, A.; et al. Vif hijacks CBF—β to degrade APOBEC3G and promote HIV–1 infection. Nature 2013, 481, 371–375. [Google Scholar] [CrossRef] [PubMed]
- Gu, Q.; Zhang, Z.; Cano, L.; Franco, C.; Häussinger, D. Feline Immunodeficiency Virus Vif N-Terminal Residues Selectively. J. Virol. 2016, 90, 10545–10557. [Google Scholar] [CrossRef] [PubMed]
- De Castro, F.L.; Junqueira, D.M.; de Medeiros, R.M.; da Silva, T.R.; Costenaro, J.G.; Knak, M.B.; de Matos Almeida, S.E.; Campos, F.S.; Roehe, P.M.; Franco, A.C. Analysis of single-nucleotide polymorphisms in the APOBEC3H gene of domestic cats (Felis catus) and their association with the susceptibility to feline immunodeficiency virus and feline leukemia virus infections. Infect. Genet. Evol. 2014, 27, 389–394. [Google Scholar] [CrossRef] [PubMed]
- Yoshikawa, R.; Izumi, T.; Yamada, E.; Nakano, Y.; Misawa, N.; Ren, F.; Carpenter, M.A.; Ikeda, T.; Munk, C.; Harris, R.S.; et al. A Naturally Occurring Domestic Cat APOBEC3 Variant Confers Resistance to Feline Immunodeficiency Virus Infection. J. Virol. 2016, 90, 474–485. [Google Scholar] [CrossRef] [PubMed]
- Cano-Ortiz, L.; Junqueira, D.M.; Comerlato, J.; Costa, C.S.; Zani, A.; Duda, N.B.; Tochetto, C.; dos Santos, R.N.; da Costa, F.V.A.; Roehe, P.M.; et al. Phylodynamics of the Brazilian feline immunodeficiency virus. Infect. Genet. Evol. 2017, 55, 166–171. [Google Scholar] [CrossRef] [PubMed]
- Kearse, M.; Moir, R.; Wilson, A.; Stones-Havas, S.; Cheung, M.; Sturrock, S.; Buxton, S.; Cooper, A.; Markowitz, S.; Duran, C.; et al. Geneious Basic: An integrated and extendable desktop software platform for the organization and analysis of sequence data. Bioinformatics 2012, 28, 1647–1649. [Google Scholar] [CrossRef] [PubMed]
- Long, J.C.; Williams, R.C.; Urbanek, M. An E-M algorithm and testing strategy for multiple-locus haplotypes. Am. J. Hum. Genet. 1995, 56, 799–810. [Google Scholar] [PubMed]
- Rose, P.P.; Korber, B.T. Detecting hypermutations in viral sequences with an emphasis on G→A hypermutation. Bioinformatics 2000, 16, 400–401. [Google Scholar] [CrossRef] [PubMed]
- Piantadosi, A.; Humes, D.; Chohan, B.; McClelland, R.S.; Overbaugh, J. Analysis of the Percentage of Human Immunodeficiency Virus Type 1 Sequences That Are Hypermutated and Markers of Disease Progression in a Longitudinal Cohort, Including One Individual with a Partially Defective Vif. J. Virol. 2009, 83, 7805–7814. [Google Scholar] [CrossRef] [PubMed]
- Pace, C.; Keller, J.; Nolan, D.; James, I.; Gaudieri, S.; Moore, C.; Mallal, S. Population level analysis of human immunodeficiency virus type 1 hypermutation and its relationship with APOBEC3G and vif genetic variation. J. Virol. 2006, 80, 9259–9269. [Google Scholar] [CrossRef] [PubMed]
- Knoepfel, S.A.; Di, F.; Däumer, M.; Thielen, A.; Metzner, K.J. In-depth analysis of G-to-A hypermutation rate in HIV-1 env DNA induced by endogenous APOBEC3 proteins using massively parallel sequencing. J. Virol. Methods 2011, 171, 329–338. [Google Scholar] [CrossRef] [PubMed]
- Eyzaguirre, L.M.; Charurat, M.; Redfield, R.R.; Blattner, W.A.; Carr, J.K.; Sajadi, M.M. Elevated hypermutation levels in HIV-1 natural viral suppressors. Virology 2013, 443, 306–312. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Fourati, S.; Lambert-Niclot, S.; Soulie, C.; Wirden, M.; Malet, I.; Valantin, M.A.; Tubiana, R.; Simon, A.; Katlama, C.; Carcelain, G.; et al. Differential impact of APOBEC3-driven mutagenesis on HIV evolution in diverse anatomical compartments. AIDS 2014, 28, 487–491. [Google Scholar] [CrossRef] [PubMed]
- Land, A.M.; Ball, T.B.; Luo, M.; Pilon, R.; Sandstrom, P.; Embree, J.E.; Wachihi, C.; Kimani, J.; Plummer, F.A. Human Immunodeficiency Virus (HIV) Type 1 Proviral Hypermutation Correlates with CD4 Count in HIV-Infected Women from Kenya. J. Virol. 2008, 82, 8172–8182. [Google Scholar] [CrossRef] [PubMed]
- Colson, P.; Ravaux, I.; Tamalet, C.; Glazunova, O.; Baptiste, E.; Chabriere, E.; Wiedemann, A.; Lacabaratz, C. HIV infection en route to endogenization: Two cases. Clin. Microbiol. Infect. 2014, 20, 1280–1288. [Google Scholar] [CrossRef] [PubMed]
| Polymorphism | Codons | Genotype | Animals N = 30 (%) |
|---|---|---|---|
| A65S | TCA/TCA TCA/GCA GCA/GCA GCA/ATA | TT TG GG GA | 5 (16.7%) 18 (60%) 5 (16.7%) 2 (6.7%) |
| R68Q | CAG/CGG CGG/CGG | AG GG | 10 (33.3%) 20 (66.7%) |
| A94T | ACG/GCG GCG/GCG | AG GG | 10 (33.3%) 20 (66.7%) |
| V96I | ATG/GTC GTC/GTC | AG GG | 4 (13.3%) 26 (86.7%) |
| Number [20] | Haplotype | Frequency (N) | Frequency in the Same Population (N) [19] |
|---|---|---|---|
| I | GGGG | 0.5 (30) | 0.55045871 (120) |
| II | TGGG | 0.23 (14) | 0.26605505 (58) |
| III | TAAG | 0.17 (10) | 0.09174312 (20) |
| IV | TGGA | 0.07 (4) | 0.0412844 (9) |
| V | AGGG | 0.3 (2) | 0.03669725 (8) |
| VI | GGGA | 0 (0) | 0.00917431 (2) |
| VII | TAGG | 0 (0) | 0.00458716 (1) |
| Total | 1 (60) | 1 (218) |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cano-Ortiz, L.; Maletich Junqueira, D.; Comerlato, J.; Zani, A.; Santos Costa, C.; Michel Roehe, P.; Franco, A.C. Absence of A3Z3-Related Hypermutations in the env and vif Proviral Genes in FIV Naturally Infected Cats. Viruses 2018, 10, 296. https://doi.org/10.3390/v10060296
Cano-Ortiz L, Maletich Junqueira D, Comerlato J, Zani A, Santos Costa C, Michel Roehe P, Franco AC. Absence of A3Z3-Related Hypermutations in the env and vif Proviral Genes in FIV Naturally Infected Cats. Viruses. 2018; 10(6):296. https://doi.org/10.3390/v10060296
Chicago/Turabian StyleCano-Ortiz, Lucía, Dennis Maletich Junqueira, Juliana Comerlato, André Zani, Cristina Santos Costa, Paulo Michel Roehe, and Ana Cláudia Franco. 2018. "Absence of A3Z3-Related Hypermutations in the env and vif Proviral Genes in FIV Naturally Infected Cats" Viruses 10, no. 6: 296. https://doi.org/10.3390/v10060296
APA StyleCano-Ortiz, L., Maletich Junqueira, D., Comerlato, J., Zani, A., Santos Costa, C., Michel Roehe, P., & Franco, A. C. (2018). Absence of A3Z3-Related Hypermutations in the env and vif Proviral Genes in FIV Naturally Infected Cats. Viruses, 10(6), 296. https://doi.org/10.3390/v10060296