Simian Varicella Virus Infects Enteric Neurons and α4β7 Integrin-Expressing Gut-Tropic T-Cells in Nonhuman Primates

 , and
, and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Tissue Specimens from Nonhuman Primates
2.2. DNA Extraction from Human Specimens
2.3. DNA Extraction from Rhesus Macaque Intestine
2.4. Quantitative Real-Time PCR
2.5. In Situ Analyses
2.6. Flow Cytometry
2.7. Statistical Analysis
3. Results
3.1. Detection of VZV DNA in Human Intestine Biopsies
3.2. Detection of Viral DNA and RNA in Intestine of Latently SVV-Infected Rhesus Macaques
3.3. SVV Infects T-Cells in Mesenteric Lymph Nodes
3.4. SVV Infects Gut-Tropic T-Cells In Vivo
4. Discussion
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Gershon, A.A.; Breuer, J.; Cohen, J.I.; Cohrs, R.J.; Gershon, M.D.; Gilden, D.; Grose, C.; Hambleton, S.; Kennedy, P.G.E.; Oxman, M.N.; et al. Varicella zoster virus infection. Nat. Rev. Dis. Prim. 2015, 1, 15016. [Google Scholar] [CrossRef] [PubMed]
- Rao, M.; Gershon, M.D. The bowel and beyond: The enteric nervous system in neurological disorders. Nat. Rev. Gastroenterol. Hepatol. 2016, 13, 517–528. [Google Scholar] [CrossRef] [PubMed]
- Gershon, A.A.; Chen, J.; Gershon, M.D. Use of Saliva to Identify Varicella Zoster Virus Infection of the Gut. Clin. Infect. Dis. 2015, 61, 536–544. [Google Scholar] [CrossRef] [PubMed]
- Gershon, A.A.; Chen, J.; Gershon, M.D. A Model of Lytic, Latent, and Reactivating Varicella-Zoster Virus Infections in Isolated Enteric Neurons. J. Infect. Dis. 2008, 197, S61–S65. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.J.; Gershon, A.A.; Li, Z.; Cowles, R.A.; Gershon, M.D. Varicella zoster virus (VZV) infects and establishes latency in enteric neurons. J. Neurovirol. 2011, 17, 578–589. [Google Scholar] [CrossRef] [PubMed]
- Carrascosa, M.F.; Salcines-Caviedes, J.R.; Román, J.G.; Cano-Hoz, M.; Fernández-Ayala, M.; Casuso-Sáenz, E.; Abascal-Carrera, I.; Campo-Ruiz, A.; Martín, M.C.; Díaz-Pérez, A.; et al. Varicella-zoster virus (VZV) infection as a possible cause of Ogilvie’s syndrome in an immunocompromised host. J. Clin. Microbiol. 2014, 52, 2718–2721. [Google Scholar] [CrossRef] [PubMed]
- Pui, J.C.; Furth, E.E.; Minda, J.; Montone, K.T. Demonstration of varicella-zoster virus infection in the muscularis propria and myenteric plexi of the colon in an HIV-positive patient with herpes zoster and small bowel pseudo-obstruction (Ogilvie’s syndrome). Am. J. Gastroenterol. 2001, 96, 1627–1630. [Google Scholar] [CrossRef] [PubMed]
- Ouwendijk, W.J.D.; Verjans, G.M.G.M. Pathogenesis of varicelloviruses in primates. J. Pathol. 2015, 235, 298–311. [Google Scholar] [CrossRef] [PubMed]
- Ouwendijk, W.J.D.; Mahalingam, R.; Traina-Dorge, V.; van Amerongen, G.; Wellish, M.; Osterhaus, A.D.M.E.; Gilden, D.; Verjans, G.M.G.M. Simian varicella virus infection of Chinese rhesus macaques produces ganglionic infection in the absence of rash. J. Neurovirol. 2012, 18, 91–99. [Google Scholar] [CrossRef] [PubMed]
- Ouwendijk, W.J.D.; Mahalingam, R.; de Swart, R.L.; Haagmans, B.L.; van Amerongen, G.; Getu, S.; Gilden, D.; Osterhaus, A.D.M.E.; Verjans, G.M.G.M. T-Cell tropism of simian varicella virus during primary infection. PLoS Pathog. 2013, 9, e1003368. [Google Scholar] [CrossRef] [PubMed]
- FEDERA. Human Tissue and Medical Research: Code of Conduct for Responsible Use (2011). Available online: https://www.federa.org/sites/default/files/images/print_version_of_conduct_english.pdf (accessed on 7 March 2017).
- Ouwendijk, W.J.D.; Choe, A.; Nagel, M.A.; Gilden, D.; Osterhaus, A.D.M.E.; Cohrs, R.J.; Verjans, G.M.G.M. Restricted varicella-zoster virus transcription in human trigeminal ganglia obtained soon after death. J. Virol. 2012, 86, 10203–10206. [Google Scholar] [CrossRef] [PubMed]
- Van Velzen, M.; Ouwendijk, W.J.D.; Selke, S.; Pas, S.D.; van Loenen, F.B.; Osterhaus, A.D.M.E.; Wald, A.; Verjans, G.M.G.M. Longitudinal study on oral shedding of herpes simplex virus 1 and varicella-zoster virus in individuals infected with HIV. J. Med. Virol. 2013, 85, 1669–1677. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Flanagan, J.; Su, N.; Wang, L.C.; Bui, S.; Nielson, A.; Wu, X.; Vo, H.T.; Ma, X.J.; Luo, Y. RNAscope: A novel In Situ RNA analysis platform for formalin-fixed, paraffin-embedded tissues. J. Mol. Diagn. 2012, 14, 22–29. [Google Scholar] [CrossRef] [PubMed]
- Cohrs, R.J.; Gilden, D.H. Prevalence and abundance of latently transcribed varicella-zoster virus genes in human ganglia. J. Virol. 2007, 81, 2950–2956. [Google Scholar] [CrossRef] [PubMed]
- Messaoudi, I.; Barron, A.; Wellish, M.; Engelmann, F.; Legasse, A.; Planer, S.; Gilden, D.; Nikolich-Zugich, J.; Mahalingam, R. Simian varicella virus infection of rhesus macaques recapitulates essential features of varicella zoster virus infection in humans. PLoS Pathog. 2009, 5. [Google Scholar] [CrossRef] [PubMed]
- Zerboni, L.; Ku, C.-C.; Jones, C.D.; Zehnder, J.L.; Arvin, A.M. Varicella-zoster virus infection of human dorsal root ganglia in vivo. Proc. Natl. Acad. Sci. USA 2005, 102, 6490–6495. [Google Scholar] [CrossRef] [PubMed]
- Ku, C.-C.; Zerboni, L.; Ito, H.; Graham, B.S.; Wallace, M.; Arvin, A.M. Varicella-zoster virus transfer to skin by T Cells and modulation of viral replication by epidermal cell interferon-alpha. J. Exp. Med. 2004, 200, 917–925. [Google Scholar] [CrossRef] [PubMed]
- Abendroth, A.; Morrow, G.; Cunningham, A.L. Varicella-Zoster Virus Infection of Human Dendritic Cells and Transmission to T Cells: Implications for Virus Dissemination in the Host. J. Virol. 2001, 75, 6183–6192. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Xu, H.; Gill, A.F.; Pahar, B.; Kempf, D.; Rasmussen, T.; Lackner, A.A.; Veazey, R.S. Monitoring α4β7 integrin expression on circulating CD4+ T cells as a surrogate marker for tracking intestinal CD4+ T-cell loss in SIV infection. Mucosal Immunol. 2009, 2, 518–526. [Google Scholar] [CrossRef] [PubMed]
- Ku, C.-C.; Padilla, J.A.; Grose, C.; Butcher, E.C.; Arvin, A.M. Tropism of varicella-zoster virus for human tonsillar CD4(+) T lymphocytes that express activation, memory, and skin homing markers. J. Virol. 2002, 76, 11425–11433. [Google Scholar] [CrossRef] [PubMed]
- Sen, N.; Mukherjee, G.; Sen, A.; Bendall, S.C.; Sung, P.; Nolan, G.P.; Arvin, A.M. Single-Cell Mass Cytometry Analysis of Human Tonsil T Cell Remodeling by Varicella Zoster Virus. Cell Rep. 2014, 8, 633–645. [Google Scholar] [CrossRef] [PubMed]
- Berlin, C.; Berg, E.L.; Briskin, M.J.; Andrew, D.P.; Kilshaw, P.J.; Holzmann, B.; Weissman, I.L.; Hamann, A.; Butcher, E.C. Alpha 4 beta 7 integrin mediates lymphocyte binding to the mucosal vascular addressin MAdCAM-1. Cell 1993, 74, 185–195. [Google Scholar] [CrossRef]
- Briskin, M.; Winsor-Hines, D.; Shyjan, A.; Cochran, N.; Bloom, S.; Wilson, J.; McEvoy, L.M.; Butcher, E.C.; Kassam, N.; Mackay, C.R.; et al. Human mucosal addressin cell adhesion molecule-1 is preferentially expressed in intestinal tract and associated lymphoid tissue. Am. J. Pathol. 1997, 151, 97–110. [Google Scholar] [PubMed]
- Weller, T.H.; Witton, H.M.; Bell, E.J. The etiologic agents of varicella and herpes zoster: Isolation, propagation, and cultural characteristics in vitro. J. Exp. Med. 1958, 108, 843–868. [Google Scholar] [CrossRef] [PubMed]
- Gershon, A.A.; Chen, J.; Davis, L.; Krinsky, C.; Cowles, R.; Reichard, R.; Gershon, M. Latency of varicella zoster virus in dorsal root, cranial, and enteric Ganglia in vaccinated children. Trans. Am. Clin. Climatol. Assoc. 2012, 123, 17–35. [Google Scholar] [PubMed]
- Gulani, J.; Koch, A.; Chappell, M.G.; Christensen, C.L.; Facemire, P.; Singh, V.K.; Ossetrova, N.I.; Srinivasan, V.; Holt, R.K. Cercopithecine Herpesvirus 9 (Simian Varicella Virus) Infection after Total-Body Irradiation in a Rhesus Macaque (Macaca mulatta). Comp. Med. 2016, 66, 150–153. [Google Scholar] [PubMed]
- Kolappaswamy, K.; Mahalingam, R.; Traina-Dorge, V.; Shipley, S.T.; Gilden, D.H.; Kleinschmidt-Demasters, B.K.; McLeod, C.G.; Hungerford, L.L.; DeTolla, L.J. Disseminated Simian Varicella Virus Infection in an Irradiated Rhesus Macaque (Macaca mulatta). J. Virol. 2006, 81, 411–415. [Google Scholar] [CrossRef] [PubMed]
- Zerboni, L.; Arvin, A. Neuronal Subtype and Satellite Cell Tropism Are Determinants of Varicella-Zoster Virus Virulence in Human Dorsal Root Ganglia Xenografts In Vivo. PLoS Pathog. 2015, 11, e100498. [Google Scholar] [CrossRef]
- Zerboni, L.; Sobel, R.A.; Lai, M.; Triglia, R.; Steain, M.; Abendroth, A.; Arvin, A. Apparent expression of varicella-zoster virus proteins in latency resulting from reactivity of murine and rabbit antibodies with human blood group a determinants in sensory neurons. J. Virol. 2012, 86, 578–583. [Google Scholar] [CrossRef] [PubMed]
- Ouwendijk, W.J.D.; Flowerdew, S.E.; Wick, D.; Horn, A.K.E.; Sinicina, I.; Strupp, M.; Osterhaus, A.D.M.E.; Verjans, G.M.G.M.; Hüfner, K. Immunohistochemical detection of intra-neuronal VZV proteins in snap-frozen human ganglia is confounded by antibodies directed against blood group A1-associated antigens. J. Neurovirol. 2012, 18, 172–180. [Google Scholar] [CrossRef] [PubMed]
- Gan, L.; Wang, M.; Chen, J.J.; Gershon, M.D.; Gershon, A.A. Infected peripheral blood mononuclear cells transmit latent varicella zoster virus infection to the guinea pig enteric nervous system. J. Neurovirol. 2014, 20, 442–456. [Google Scholar] [CrossRef] [PubMed]
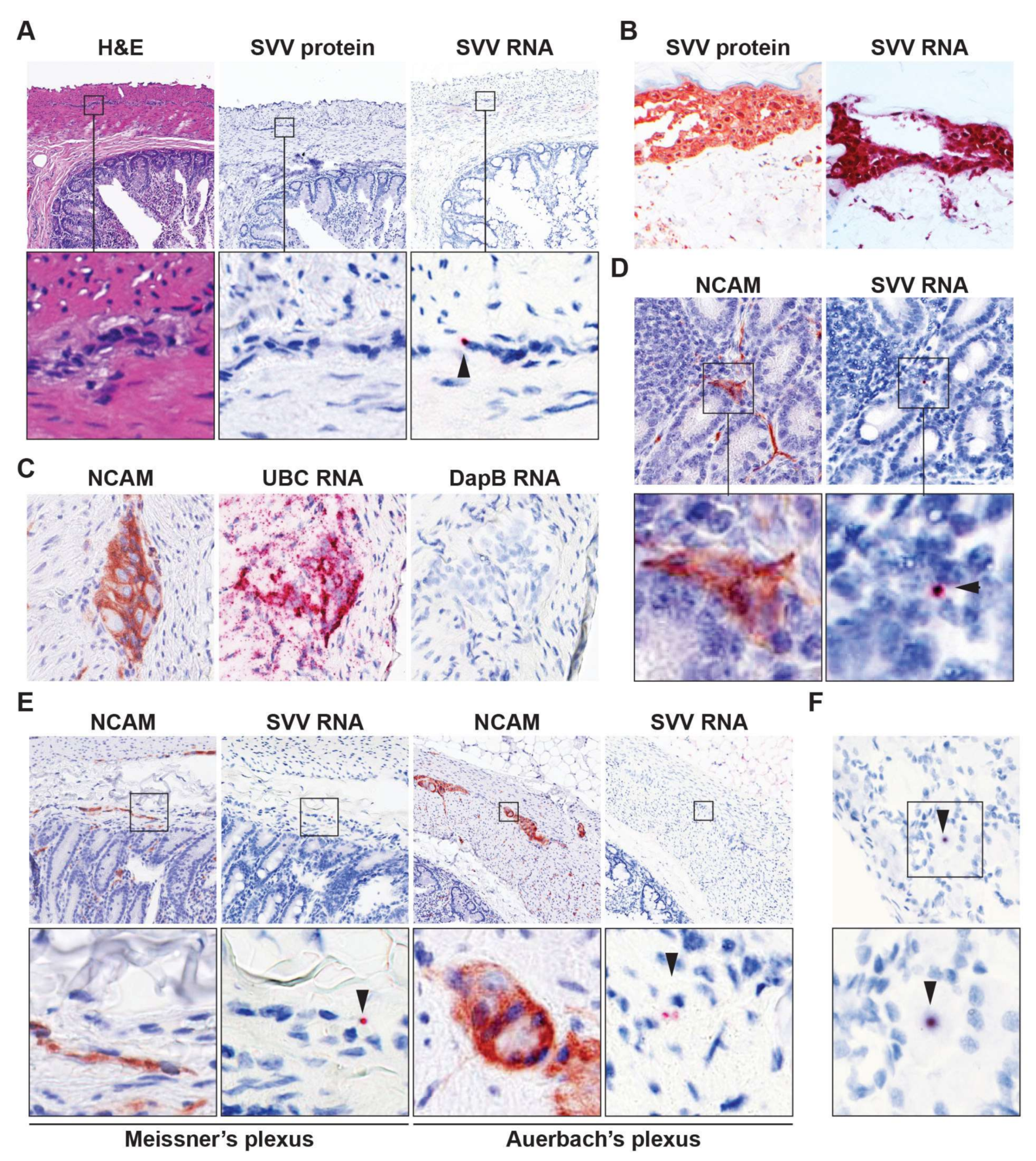



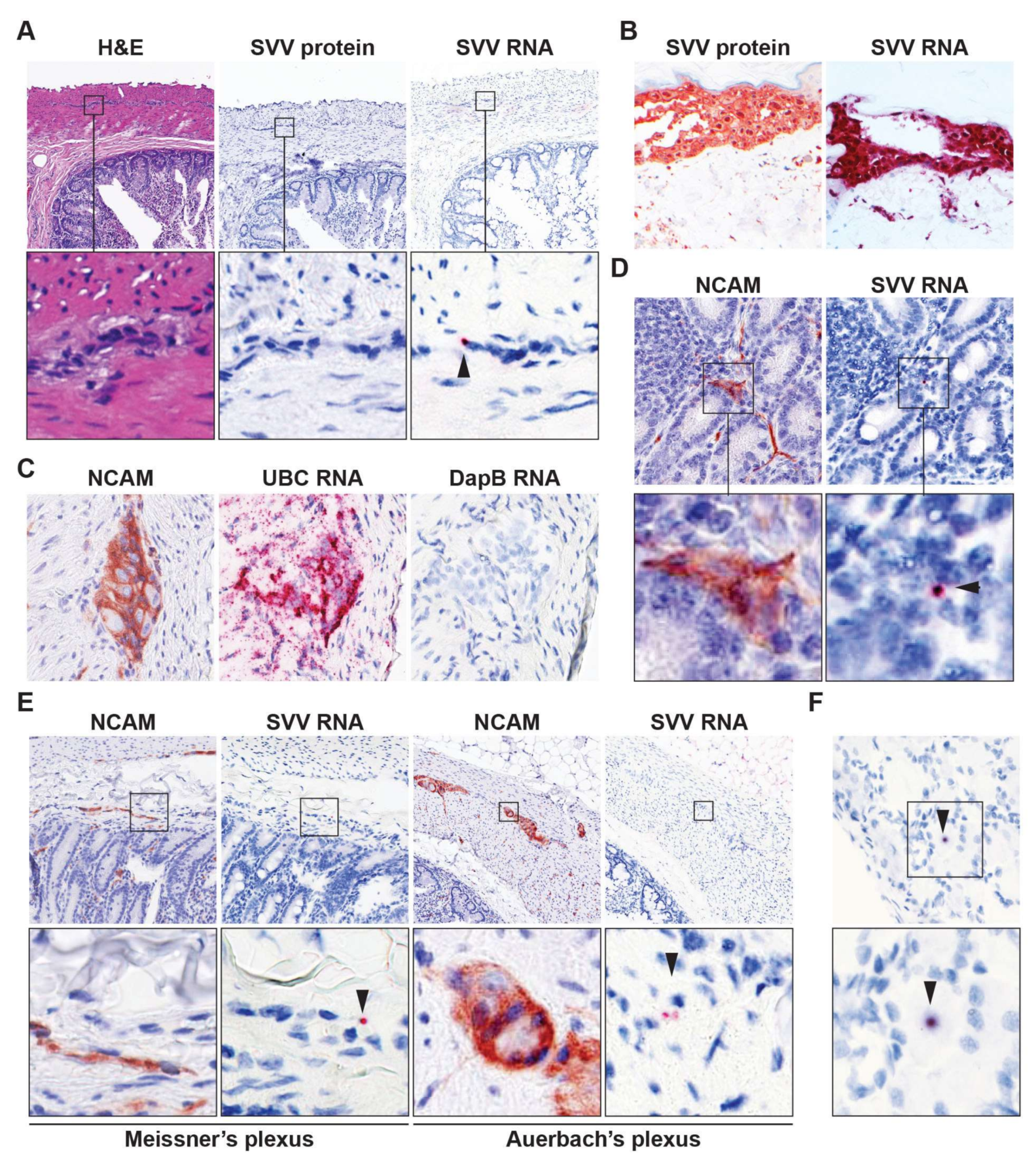{kind=link}
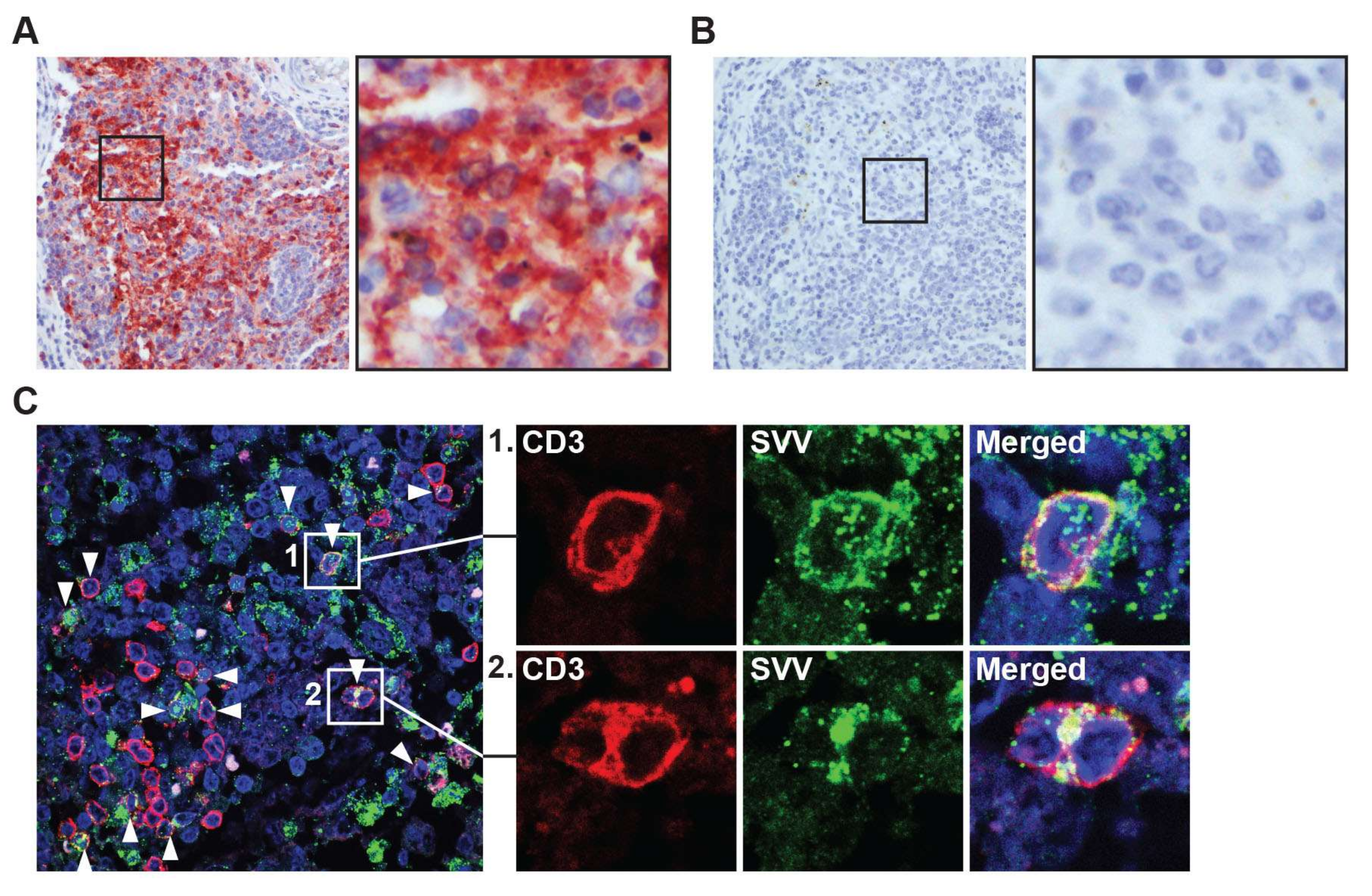{kind=link}
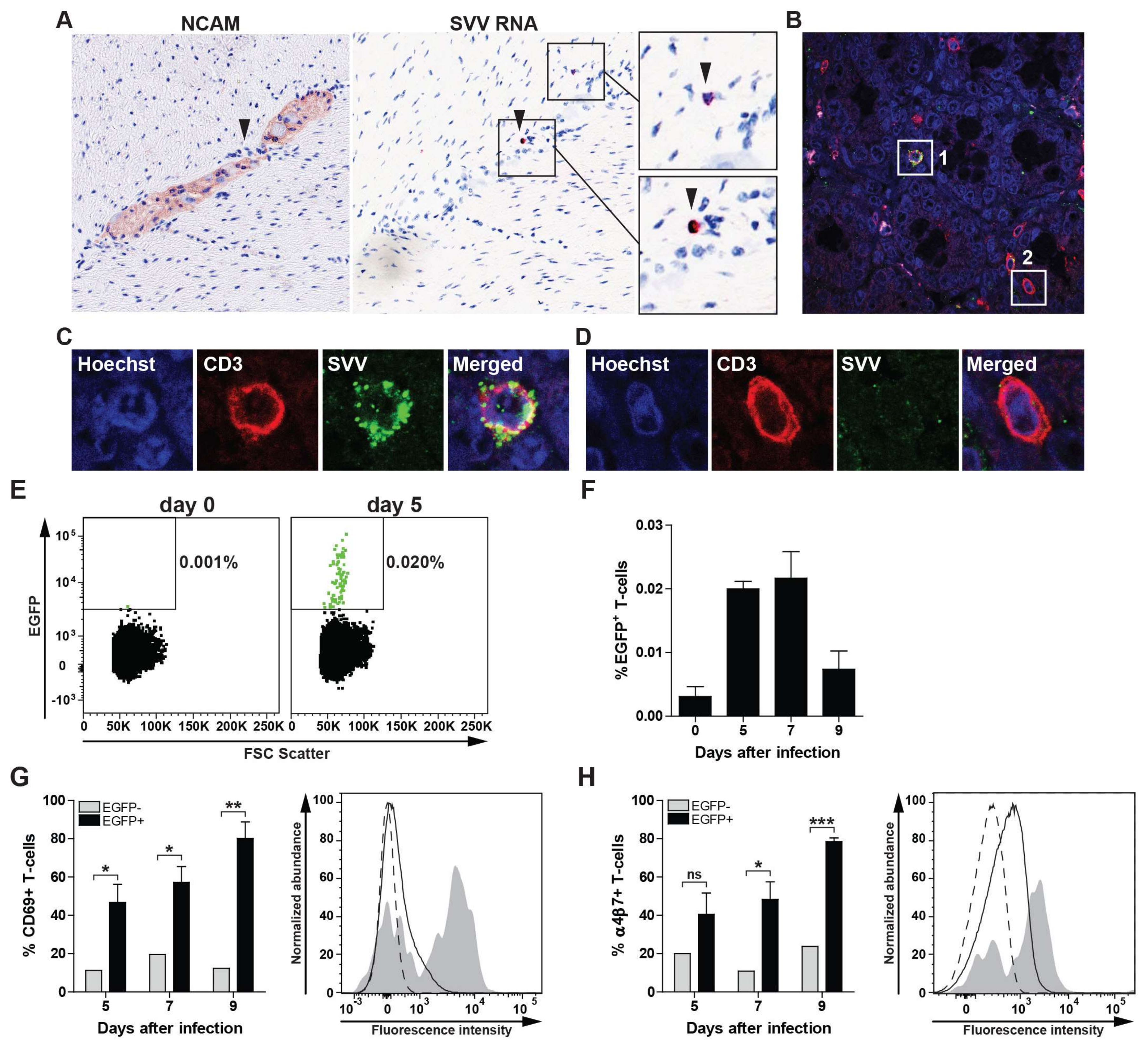{kind=link}
| Patient ID | Gender 1 | Age 2 | Disease at Time of Intestinal Biopsy | DNA 3 | |
|---|---|---|---|---|---|
| HSV-1 | VZV | ||||
| 1 | F | 72 | Small intestine B-cell lymphoma | – | 2 |
| 2 | F | 55 | Small intestine neuroendocrine tumor | – | 107 |
| 3 | F | 53 | Colonic adenocarcinoma | – | – |
| 4 | F | 57 | Small intestine B-cell lymphoma | – | – |
| 5 | F | 51 | Pancreatic carcinoma | – | – |
| 6 | M | 44 | Small intestine gastrointestinal stromal tumor | – | – |
| 7 | F | 40 | Colonic adenocarcinoma | – | – |
| 8 | F | 65 | Bladder adenocarcinoma | – | – |
| 9 | M | 48 | Metastatic melanoma | – | – |
| 10 | M | 61 | Small intestine neuroendocrine tumor | – | – |
| 11 | F | 64 | Ovarian papillary serous carcinoma | – | 423 |
| 12 | F | 60 | Colonic adenocarcinoma | – | – |
| 13 | M | 58 | Small intestine neuroendocrine tumor | – | – |
| 14 | M | 55 | Small intestine neuroendocrine tumor | – | – |
| 15 | M | 73 | Metastatic carcinoma (unknown origin) | – | – |
| 16 | F | 75 | Adnexal carcinosarcoma | – | 93 |
| 17 | M | 32 | Peutz-Jeghers syndrome | – | 30 |
| 18 | M | 68 | Carcinoid small intestine | – | – |
| Animal ID | Tissue 1 | SVV DNA 2 |
|---|---|---|
| 9021 | Colon + small intestine | 14 |
| Colon | 11 | |
| Duodenum | – | |
| Ileum | – | |
| Jejunum | – | |
| Caecum | – | |
| 2207 | Colon + small intestine | 290 |
| Colon | – | |
| Duodenum | 12 | |
| Ileum | 14 | |
| Jejunum | – | |
| Caecum | 110 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ouwendijk, W.J.D.; Van Veen, S.; Mehraban, T.; Mahalingam, R.; Verjans, G.M.G.M. Simian Varicella Virus Infects Enteric Neurons and α4β7 Integrin-Expressing Gut-Tropic T-Cells in Nonhuman Primates. Viruses 2018, 10, 156. https://doi.org/10.3390/v10040156
Ouwendijk WJD, Van Veen S, Mehraban T, Mahalingam R, Verjans GMGM. Simian Varicella Virus Infects Enteric Neurons and α4β7 Integrin-Expressing Gut-Tropic T-Cells in Nonhuman Primates. Viruses. 2018; 10(4):156. https://doi.org/10.3390/v10040156
Chicago/Turabian StyleOuwendijk, Werner J. D., Suzanne Van Veen, Tamana Mehraban, Ravi Mahalingam, and Georges M. G. M. Verjans. 2018. "Simian Varicella Virus Infects Enteric Neurons and α4β7 Integrin-Expressing Gut-Tropic T-Cells in Nonhuman Primates" Viruses 10, no. 4: 156. https://doi.org/10.3390/v10040156
APA StyleOuwendijk, W. J. D., Van Veen, S., Mehraban, T., Mahalingam, R., & Verjans, G. M. G. M. (2018). Simian Varicella Virus Infects Enteric Neurons and α4β7 Integrin-Expressing Gut-Tropic T-Cells in Nonhuman Primates. Viruses, 10(4), 156. https://doi.org/10.3390/v10040156