A Mouse Model of Enterovirus D68 Infection for Assessment of the Efficacy of Inactivated Vaccine

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cells and Viruses
2.2. Antibodies
2.3. Mouse Infection Experiments
2.4. Determination of Viral Loads in Tissues of Infected Mice
2.5. Histopathological and Immunohistochemical Staining
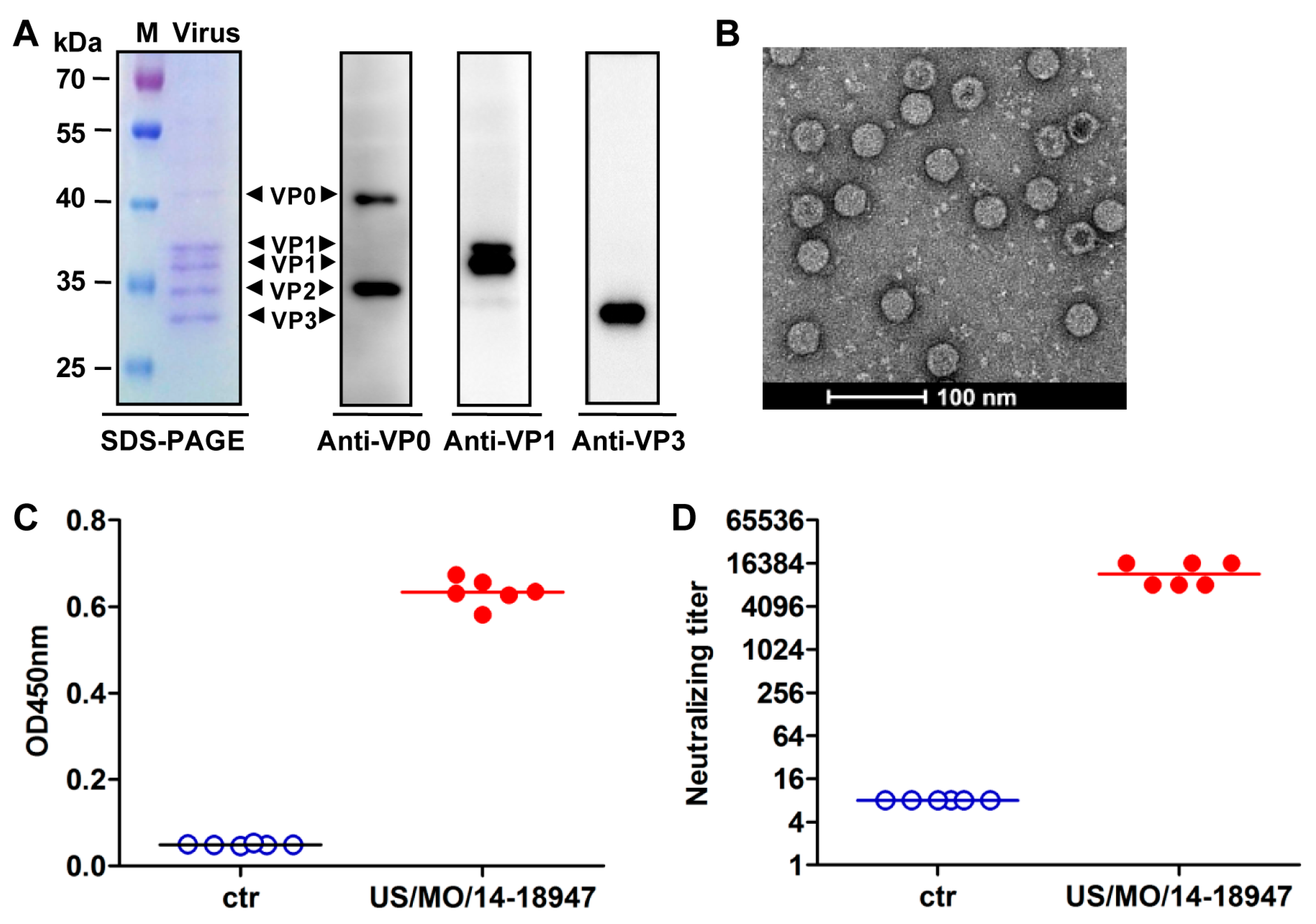
2.6. Preparation of Inactivated EVD68
2.7. Mouse Immunization and Serum Antibody Measurement
2.8. In Vivo Protection Assays
3. Results
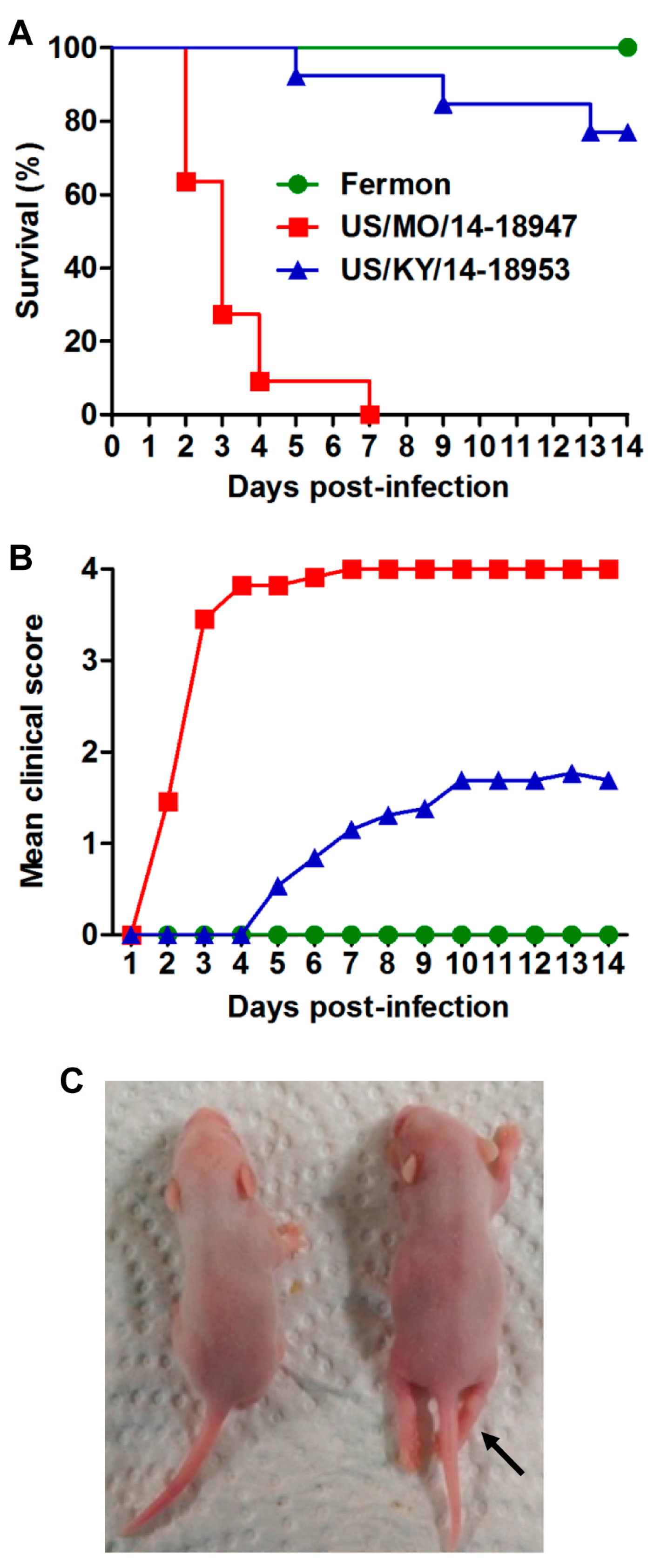
3.1. Comparison of the Virulence of Different EVD68 Strains in Mice
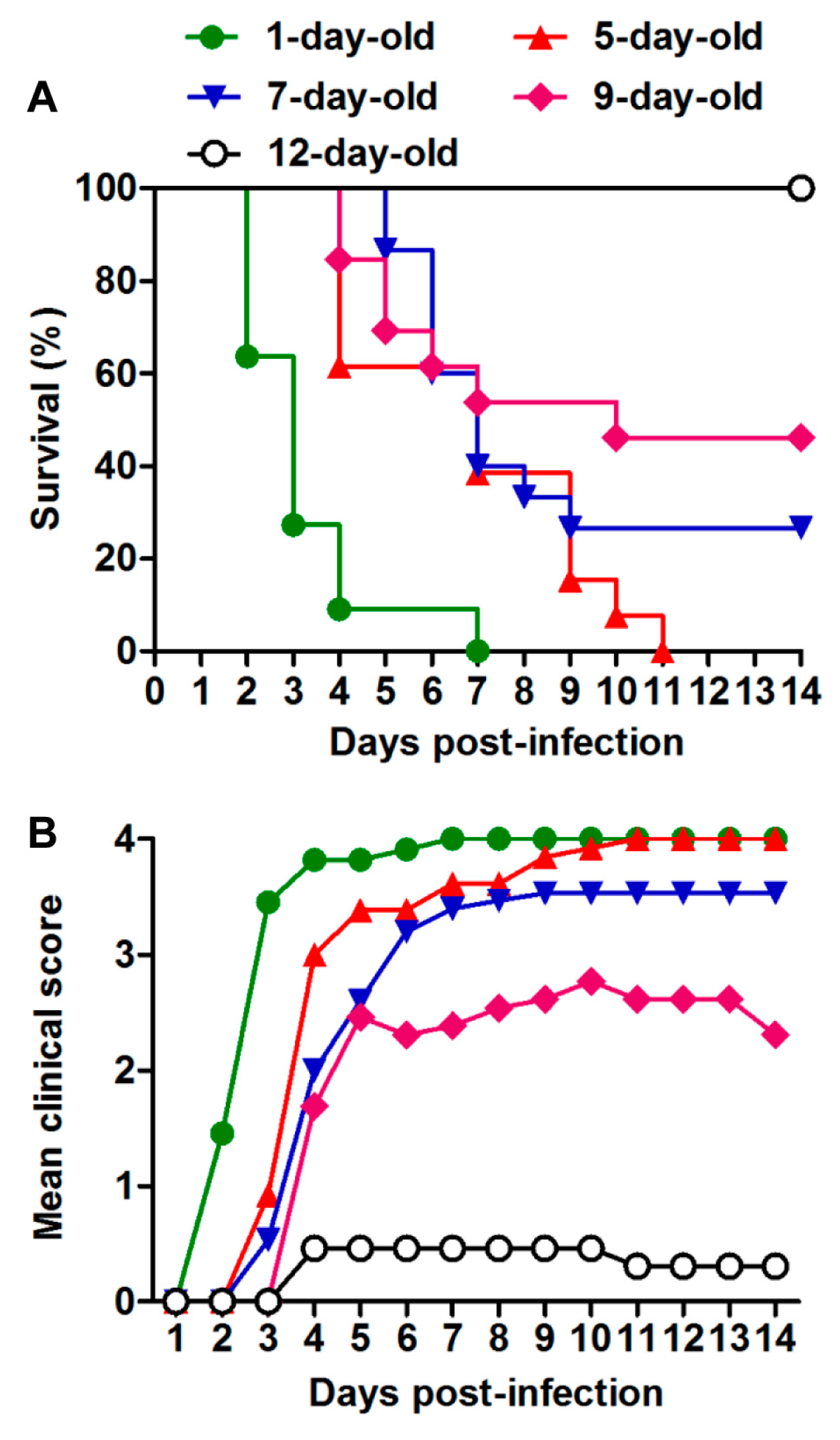
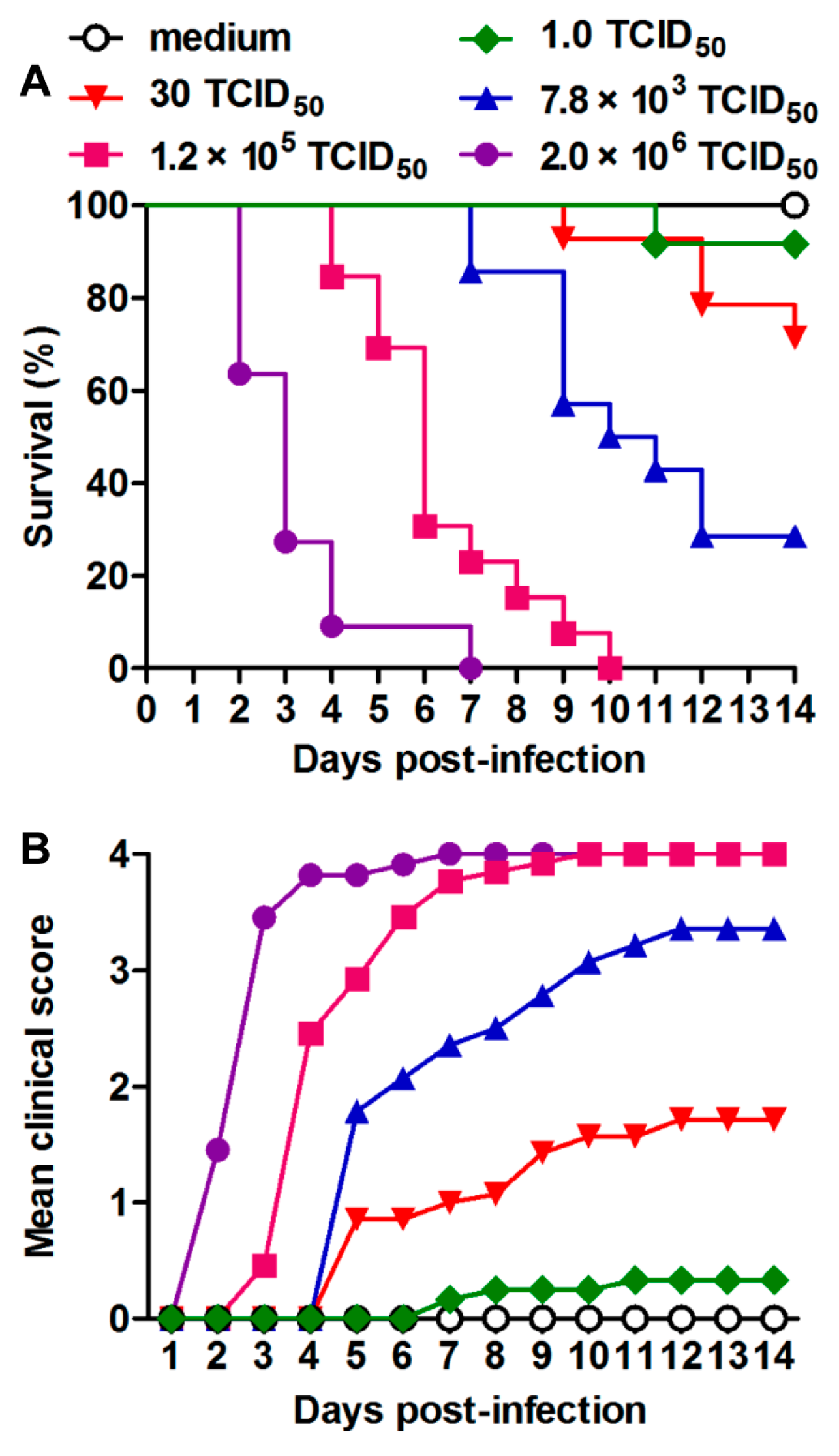
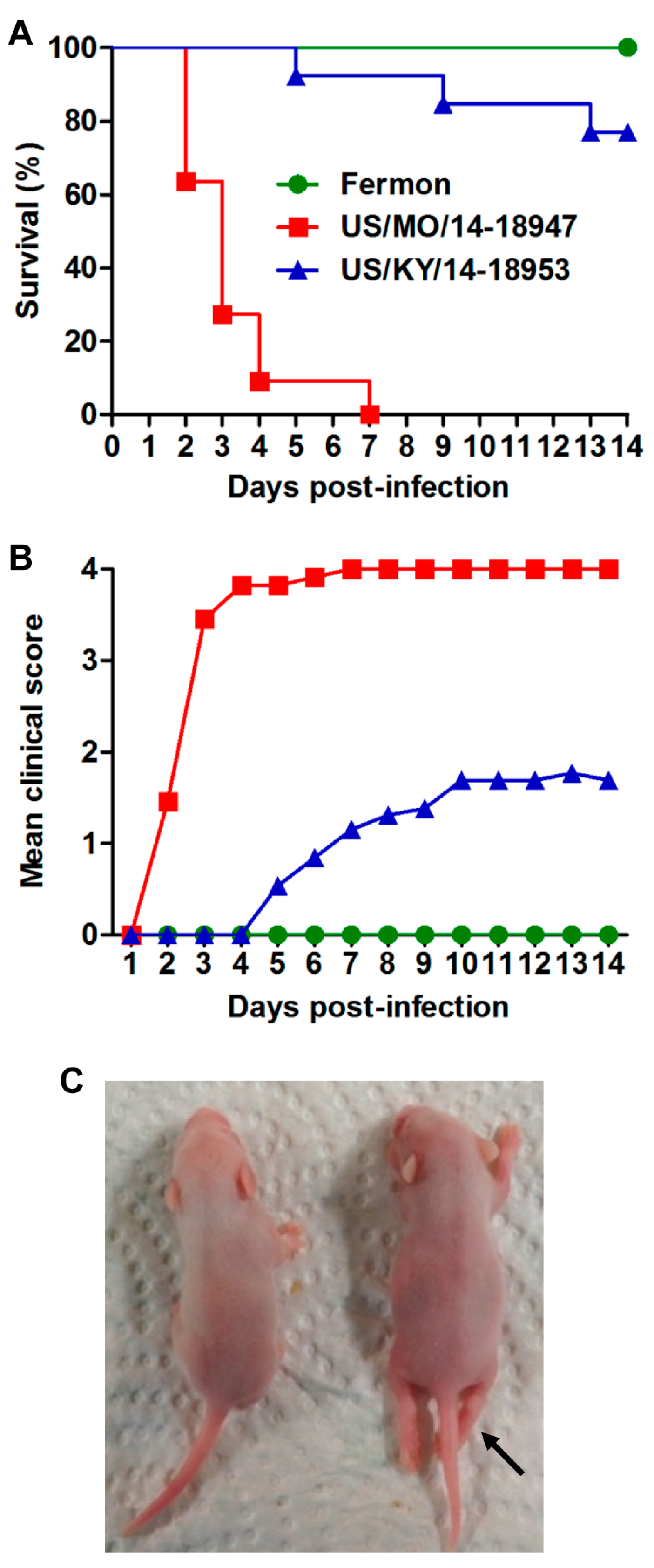
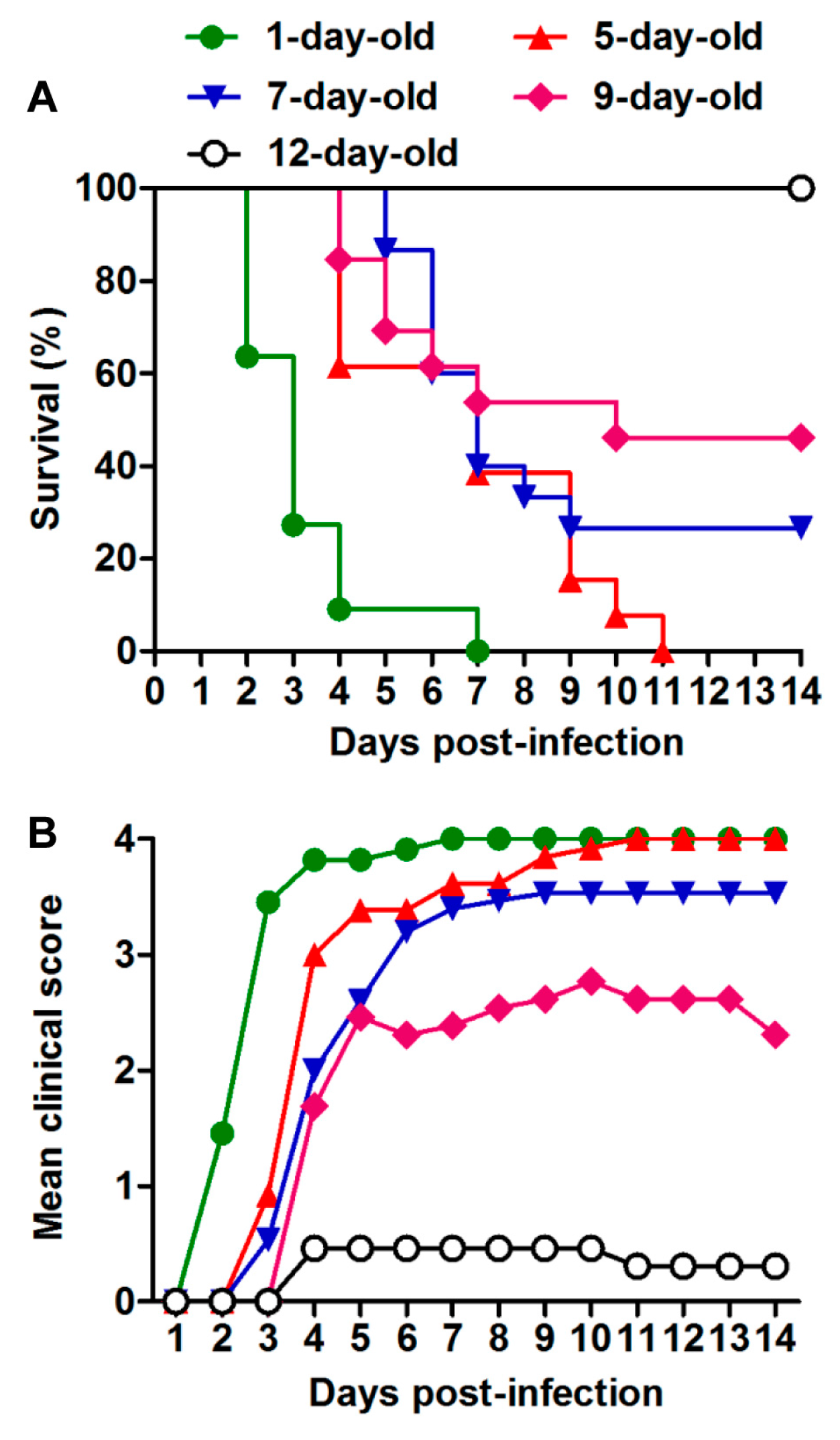
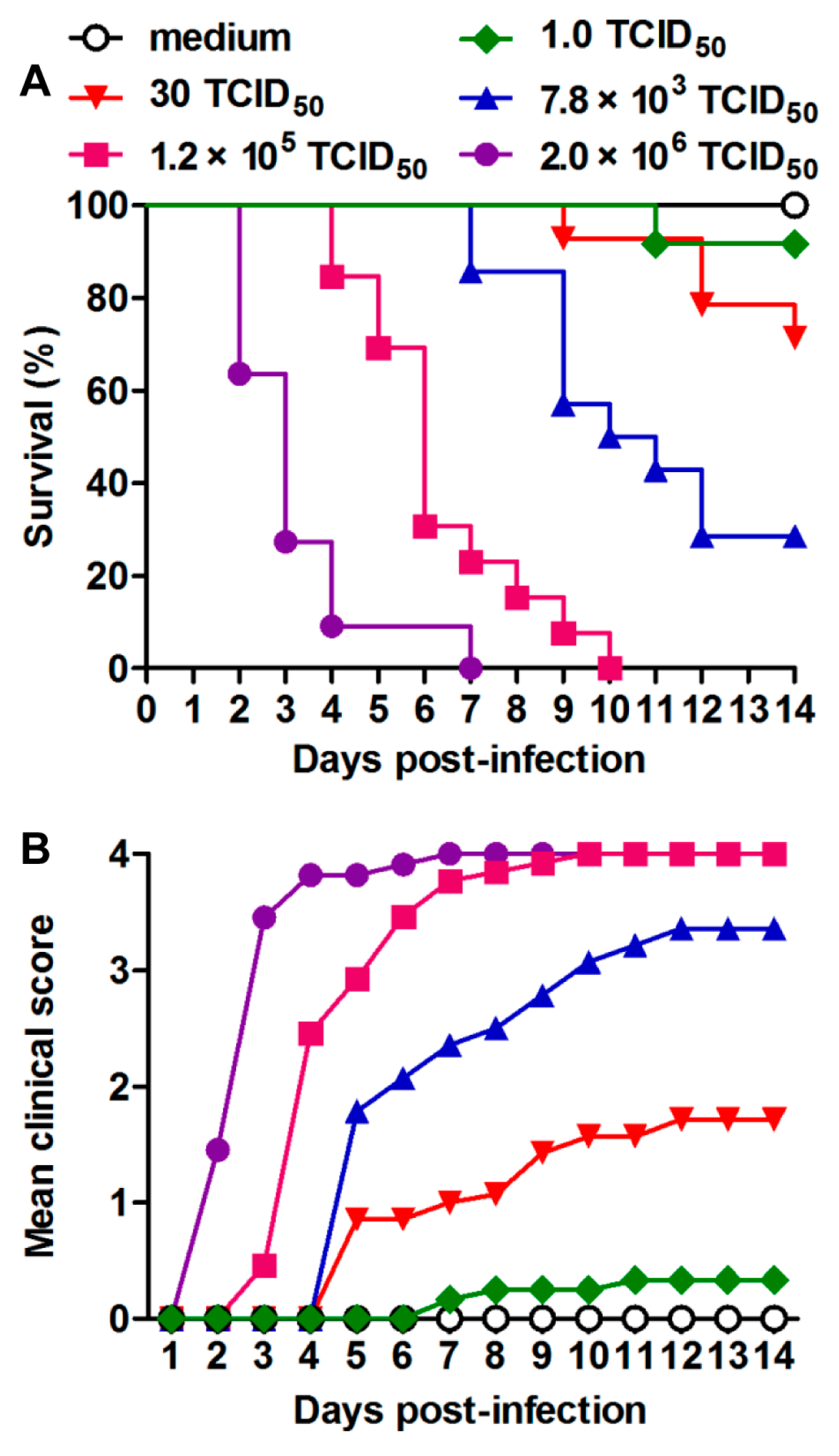
3.2. US/MO/14-18947 Infection Induced Death in Mice in an Age- and Virus Dose-Dependent Manner
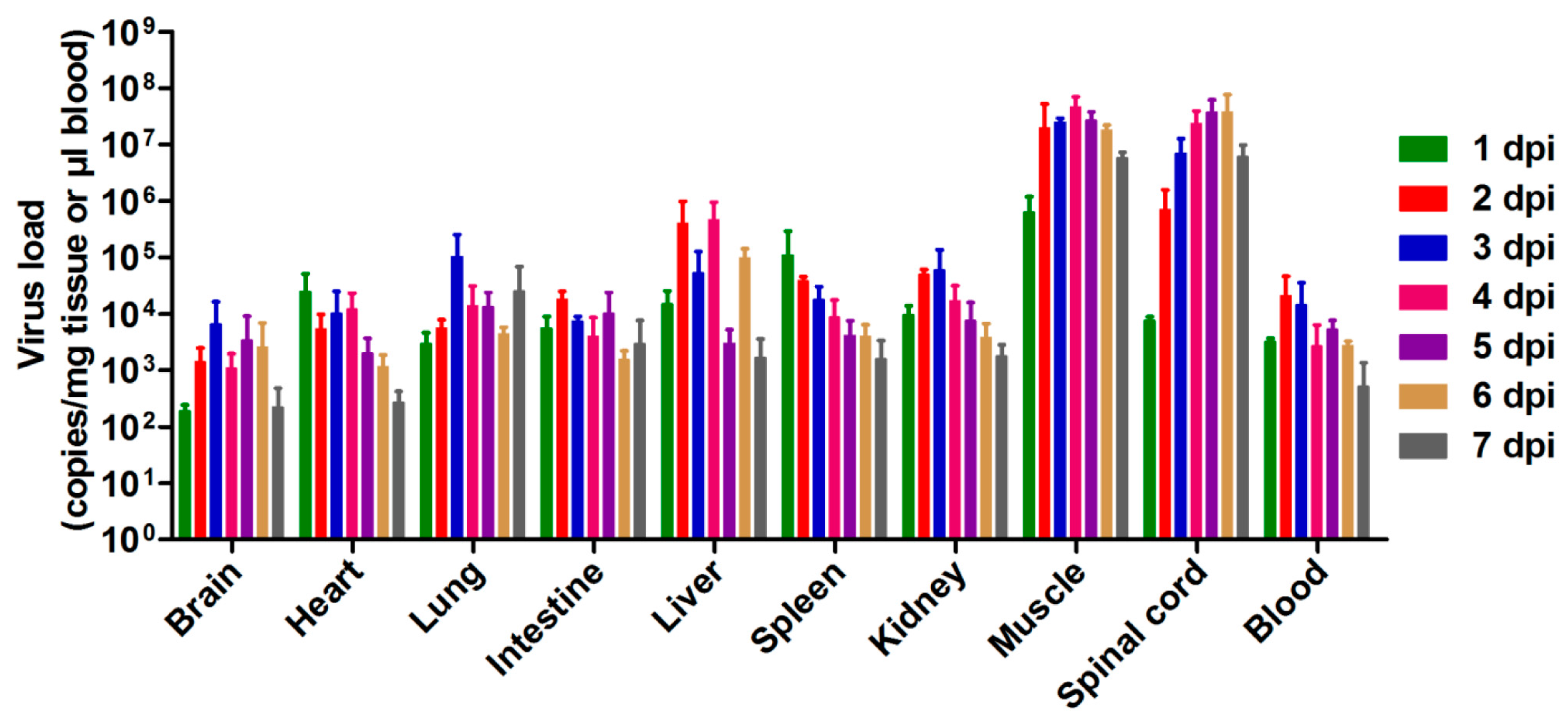
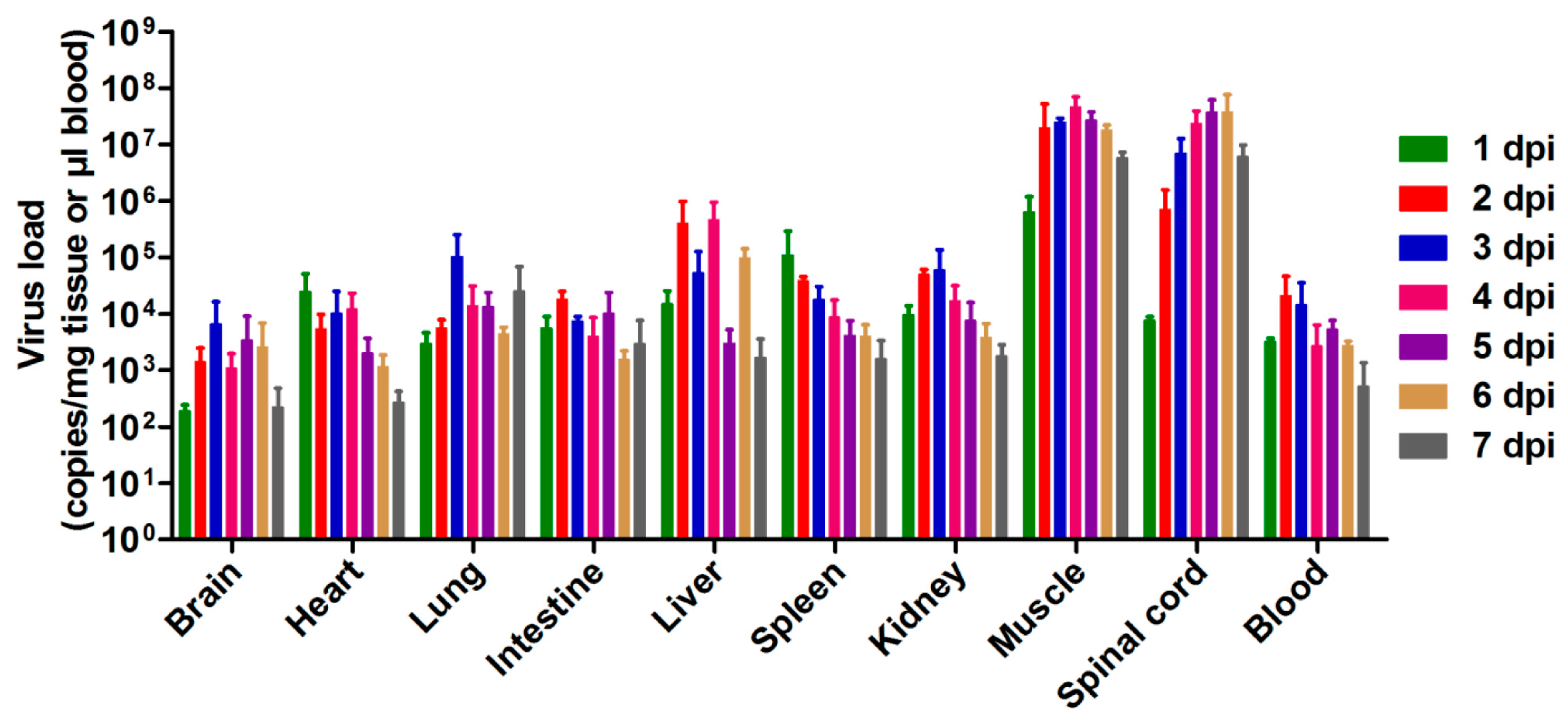
3.3. Tissue Viral Loads in US/MO/14-18947-Infected Mice
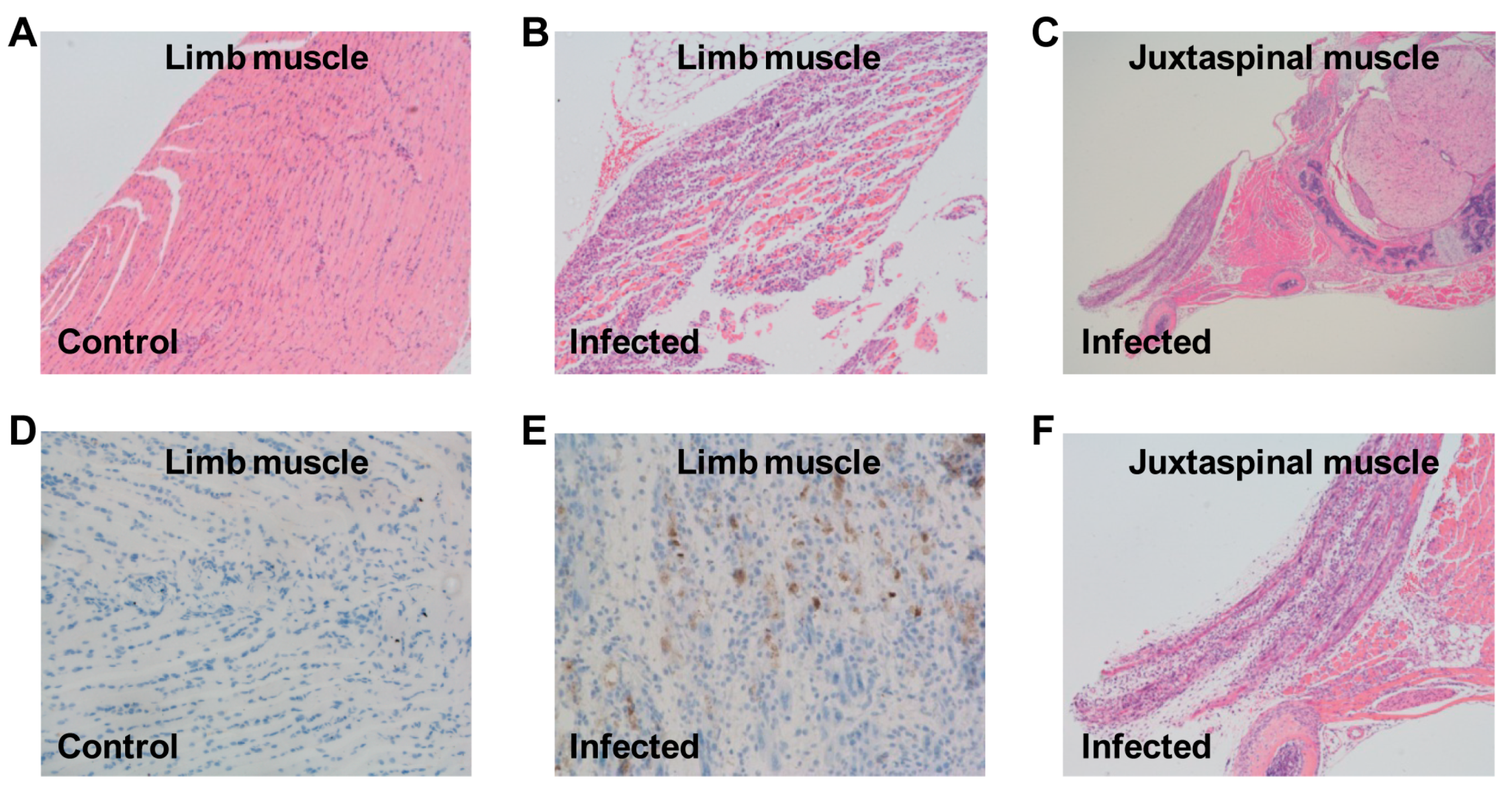
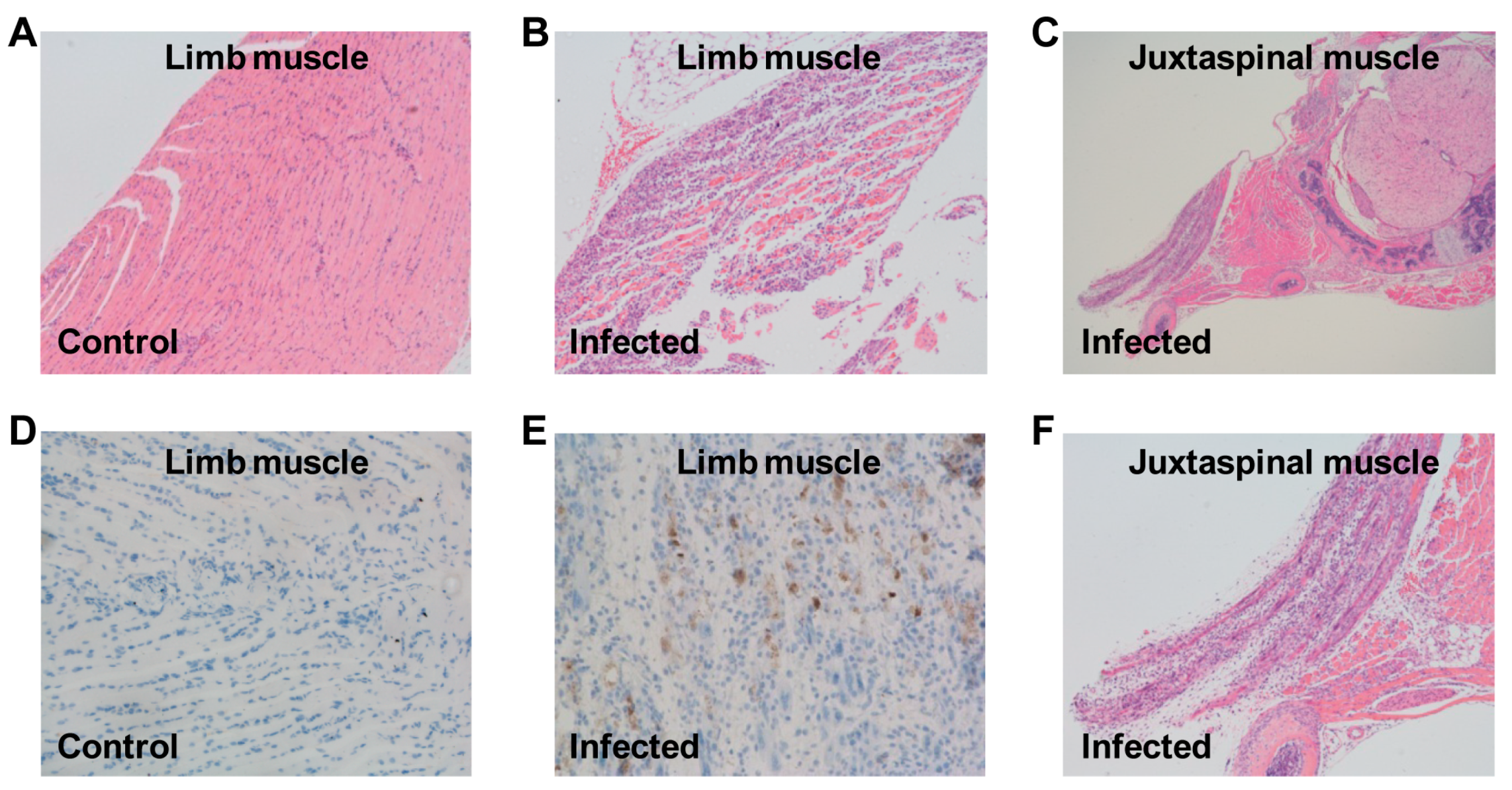
3.4. Pathological Analysis of US/MO/14-18947-Infected Mice
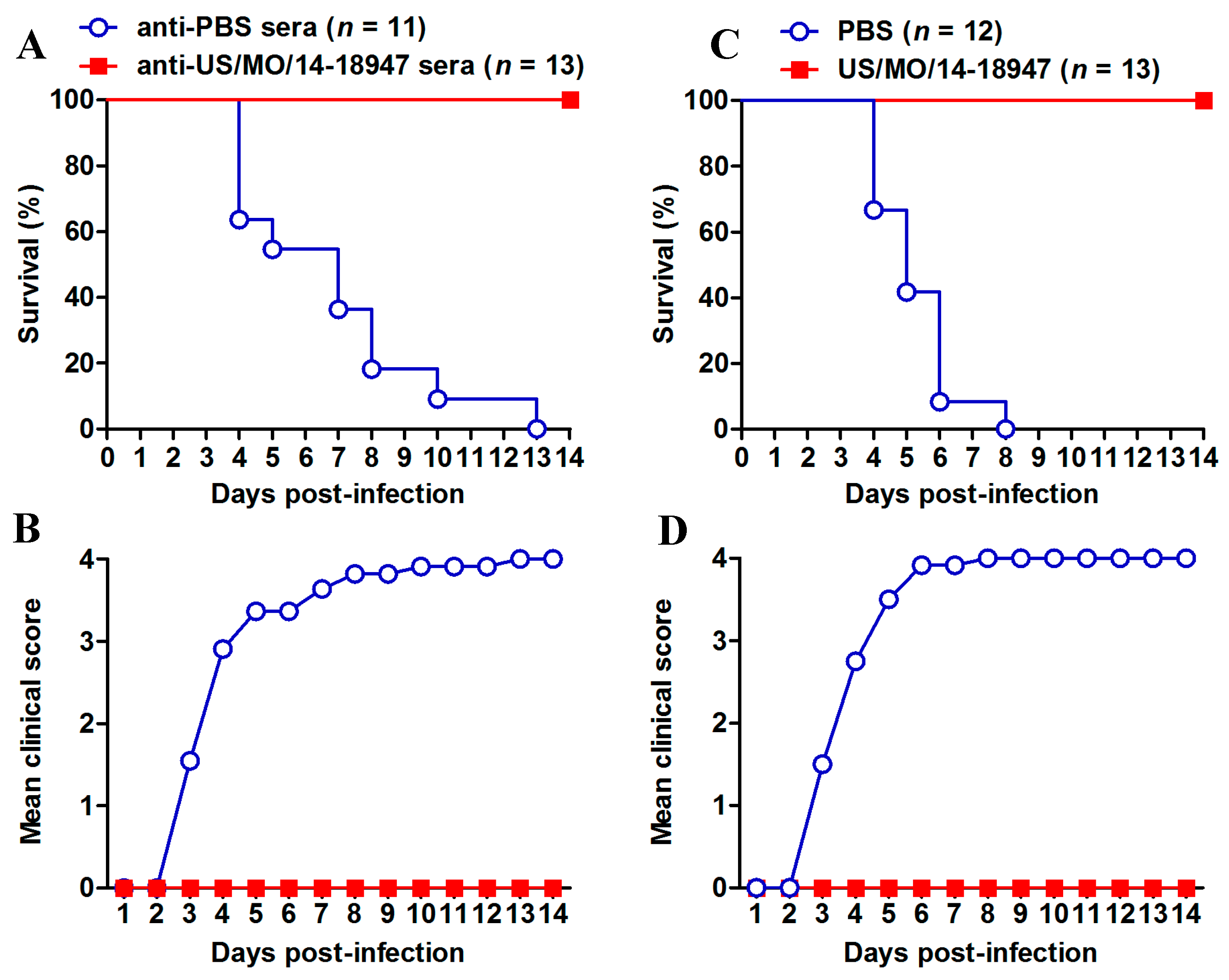
3.5. Evaluation of the Protective Efficacy of Inactivated EVD68 Vaccine in the Mouse Model
4. Discussion
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Imamura, T.; Oshitani, H. Global reemergence of enterovirus D68 as an important pathogen for acute respiratory infections. Rev. Med. Virol. 2015, 25, 102–114. [Google Scholar] [CrossRef] [PubMed]
- Schieble, J.H.; Fox, V.L.; Lennette, E.H. A probable new human picornavirus associated with respiratory diseases. Am. J. Epidemiol. 1967, 85, 297–310. [Google Scholar] [CrossRef] [PubMed]
- Tokarz, R.; Firth, C.; Madhi, S.A.; Howie, S.R.; Wu, W.; Sall, A.A.; Haq, S.; Briese, T.; Lipkin, W.I. Worldwide emergence of multiple clades of enterovirus 68. J. Gen. Virol. 2012, 93, 1952–1958. [Google Scholar] [CrossRef] [PubMed]
- Holm-Hansen, C.C.; Midgley, S.E.; Fischer, T.K. Global emergence of enterovirus D68: A systematic review. Lancet Infect. Dis. 2016, 16, e64–e75. [Google Scholar] [CrossRef]
- Xiang, Z.; Li, L.; Ren, L.; Guo, L.; Xie, Z.; Liu, C.; Li, T.; Luo, M.; Paranhos-Baccala, G.; Xu, W.; et al. Seroepidemiology of enterovirus D68 infection in China. Emerg. Microbes Infect. 2017, 6, e32. [Google Scholar] [CrossRef] [PubMed]
- Messacar, K.; Robinson, C.C.; Pretty, K.; Yuan, J.; Dominguez, S.R. Surveillance for enterovirus D68 in Colorado children reveals continued circulation. J. Clin. Virol. 2017, 92, 39–41. [Google Scholar] [CrossRef] [PubMed]
- Korematsu, S.; Nagashima, K.; Sato, Y.; Nagao, M.; Hasegawa, S.; Nakamura, H.; Sugiura, S.; Miura, K.; Okada, K.; Fujisawa, T. “Spike” in acute asthma exacerbations during enterovirus D68 epidemic in Japan: A nation-wide survey. Allergol. Int. 2018, 67, 55–60. [Google Scholar] [PubMed]
- Dyrdak, R.; Grabbe, M.; Hammas, B.; Ekwall, J.; Hansson, K.E.; Luthander, J.; Naucler, P.; Reinius, H.; Rotzen-Ostlund, M.; Albert, J. Outbreak of enterovirus D68 of the new B3 lineage in Stockholm, Sweden, August to September 2016. Eurosurveillance 2016, 21, 30403. [Google Scholar] [CrossRef] [PubMed]
- Knoester, M.; Scholvinck, E.H.; Poelman, R.; Smit, S.; Vermont, C.L.; Niesters, H.G.; van Leer-Buter, C.C. Upsurge of Enterovirus D68, The Netherlands, 2016. Emerg. Infect. Dis. 2017, 23, 140–143. [Google Scholar] [CrossRef] [PubMed]
- Sejvar, J.J.; Lopez, A.S.; Cortese, M.M.; Leshem, E.; Pastula, D.M.; Miller, L.; Glaser, C.; Kambhampati, A.; Shioda, K.; Aliabadi, N.; et al. Acute Flaccid Myelitis in the United States, August–December 2014: Results of Nationwide Surveillance. Clin. Infect. Dis. 2016, 63, 737–745. [Google Scholar] [CrossRef] [PubMed]
- Messacar, K.; Schreiner, T.L.; van Haren, K.; Yang, M.; Glaser, C.A.; Tyler, K.L.; Dominguez, S.R. Acute flaccid myelitis: A clinical review of US cases 2012–2015. Ann. Neurol. 2016, 80, 326–338. [Google Scholar] [CrossRef] [PubMed]
- Perez, G.; Rosanova, M.T.; Freire, M.C.; Paz, M.I.; Ruvinsky, S.; Rugilo, C.; Ruggieri, V.; Cisterna, D.; Martiren, S.; Lema, C.; et al. Unusual increase of cases of myelitis in a pediatric hospital in Argentina. Archivos Argentinos de Pediatria 2017, 115, 364–369. [Google Scholar] [PubMed]
- Chong, P.F.; Kira, R.; Mori, H.; Okumura, A.; Torisu, H.; Yasumoto, S.; Shimizu, H.; Fujimoto, T.; Hanaoka, N.; Kusunoki, S.; et al. Clinical Features of Acute Flaccid Myelitis Temporally Associated with an Enterovirus D68 Outbreak: Results of a Nationwide Survey of Acute Flaccid Paralysis in Japan, August–December 2015. Clin. Infect. Dis. 2017. [Google Scholar] [CrossRef]
- Ruggieri, V.; Paz, M.I.; Peretti, M.G.; Rugilo, C.; Bologna, R.; Freire, C.; Vergel, S.; Savransky, A. Enterovirus D68 infection in a cluster of children with acute flaccid myelitis, buenos aires, argentina, 2016. Eur. J. Paediatr. Neurol. 2017, 21, 884–890. [Google Scholar] [CrossRef] [PubMed]
- Esposito, S.; Chidini, G.; Cinnante, C.; Napolitano, L.; Giannini, A.; Terranova, L.; Niesters, H.; Principi, N.; Calderini, E. Acute flaccid myelitis associated with enterovirus-D68 infection in an otherwise healthy child. Virol. J. 2017, 14, 4. [Google Scholar] [CrossRef] [PubMed]
- Yoder, J.A.; Lloyd, M.; Zabrocki, L.; Auten, J. Pediatric acute flaccid paralysis: Enterovirus D68-Associated Anterior Myelitis. J. Emerg. Med. 2017, 53, e19–e23. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Sheng, J.; Fokine, A.; Meng, G.; Shin, W.H.; Long, F.; Kuhn, R.J.; Kihara, D.; Rossmann, M.G. Structure and inhibition of EV-D68, a virus that causes respiratory illness in children. Science 2015, 347, 71–74. [Google Scholar] [CrossRef] [PubMed]
- Gong, Y.N.; Yang, S.L.; Shih, S.R.; Huang, Y.C.; Chang, P.Y.; Huang, C.G.; Kao, K.C.; Hu, H.C.; Liu, Y.C.; Tsao, K.C. Molecular evolution and the global reemergence of enterovirus D68 by genome-wide analysis. Medicine 2016, 95, e4416. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Cao, J.; Zhang, S.; Lee, A.J.; Sun, G.; Larsen, C.N.; Zhao, H.; Gu, Z.; He, S.; Klem, E.B.; et al. Genetic changes found in a distinct clade of Enterovirus D68 associated with paralysis during the 2014 outbreak. Virus Evol. 2016, 2, vew015. [Google Scholar] [CrossRef] [PubMed]
- Patel, M.C.; Wang, W.; Pletneva, L.M.; Rajagopala, S.V.; Tan, Y.; Hartert, T.V.; Boukhvalova, M.S.; Vogel, S.N.; Das, S.R.; Blanco, J.C. Enterovirus D-68 Infection, Prophylaxis, and Vaccination in a Novel Permissive Animal Model, the Cotton Rat (Sigmodon hispidus). PLoS ONE 2016, 11, e0166336. [Google Scholar] [CrossRef] [PubMed]
- Zheng, H.W.; Sun, M.; Guo, L.; Wang, J.J.; Song, J.; Li, J.Q.; Li, H.Z.; Ning, R.T.; Yang, Z.N.; Fan, H.T.; et al. Nasal Infection of Enterovirus D68 Leading to Lower Respiratory Tract Pathogenesis in Ferrets (Mustela putorius furo). Viruses 2017, 9, 104. [Google Scholar] [CrossRef] [PubMed]
- Hixon, A.M.; Yu, G.; Leser, J.S.; Yagi, S.; Clarke, P.; Chiu, C.Y.; Tyler, K.L. A mouse model of paralytic myelitis caused by enterovirus D68. PLoS Pathog. 2017, 13, e1006199. [Google Scholar] [CrossRef] [PubMed]
- Ku, Z.; Shi, J.; Liu, Q.; Huang, Z. Development of murine monoclonal antibodies with potent neutralization effects on enterovirus 71. J. Virol. Methods 2012, 186, 193–197. [Google Scholar] [CrossRef] [PubMed]
- Cai, Y.; Liu, Q.; Huang, X.; Li, D.; Ku, Z.; Zhang, Y.; Huang, Z. Active immunization with a Coxsackievirus A16 experimental inactivated vaccine induces neutralizing antibodies and protects mice against lethal infection. Vaccine 2013, 31, 2215–2221. [Google Scholar] [CrossRef] [PubMed]
- Reed, L.J.; Muench, H. A simple method of estimating fifty per cent endpoints. Am. J. Epidemiol. 1938, 27, 493–497. [Google Scholar] [CrossRef]
- Zhang, C.; Zhang, X.; Zhang, W.; Dai, W.; Xie, J.; Ye, L.; Wang, H.; Chen, H.; Liu, Q.; Gong, S.; et al. Enterovirus D68 virus-like particles expressed in Pichia pastoris potently induce neutralizing antibody responses and confer protection against lethal viral infection in mice. Emerg. Microbes Infect. 2018, 7, 3. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Ku, Z.; Liu, Q.; Wang, X.; Chen, T.; Ye, X.; Li, D.; Jin, X.; Huang, Z. High-yield production of recombinant virus-like particles of enterovirus 71 in pichia pastoris and their protective efficacy against oral viral challenge in mice. Vaccine 2015, 33, 2335–2341. [Google Scholar] [CrossRef] [PubMed]
- Ku, Z.; Ye, X.; Huang, X.; Cai, Y.; Liu, Q.; Li, Y.; Su, Z.; Huang, Z. Neutralizing antibodies induced by recombinant virus-like particles of enterovirus 71 genotype C4 inhibit infection at pre- and post-attachment steps. PLoS ONE 2013, 8, e57601. [Google Scholar] [CrossRef] [PubMed]
- Greninger, A.L.; Naccache, S.N.; Messacar, K.; Clayton, A.; Yu, G.; Somasekar, S.; Federman, S.; Stryke, D.; Anderson, C.; Yagi, S.; et al. A novel outbreak enterovirus D68 strain associated with acute flaccid myelitis cases in the USA (2012–14): A retrospective cohort study. Lancet Infect. Dis. 2015, 15, 671–682. [Google Scholar] [CrossRef]
- Maloney, J.A.; Mirsky, D.M.; Messacar, K.; Dominguez, S.R.; Schreiner, T.; Stence, N.V. MRI findings in children with acute flaccid paralysis and cranial nerve dysfunction occurring during the 2014 enterovirus D68 outbreak. AJNR Am. J. Neuroradiol. 2015, 36, 245–250. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.C.; Guo, M.S.; Lin, F.H.; Hsiao, K.N.; Chang, K.H.; Chou, A.H.; Wang, Y.C.; Chen, Y.C.; Yang, C.S.; Chong, P.C. Purification and characterization of enterovirus 71 viral particles produced from vero cells grown in a serum-free microcarrier bioreactor system. PLoS ONE 2011, 6, e20005. [Google Scholar] [CrossRef] [PubMed]
- Chong, P.; Guo, M.S.; Lin, F.H.; Hsiao, K.N.; Weng, S.Y.; Chou, A.H.; Wang, J.R.; Hsieh, S.Y.; Su, I.J.; Liu, C.C. Immunological and biochemical characterization of coxsackie virus A16 viral particles. PLoS ONE 2012, 7, e49973. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Pooled Antisera | Neutralization Titer Against | ||||
|---|---|---|---|---|---|
| EVD68/ Fermon | EVD68/ US/MO/14-18947 | EVD68/ US/KY/14-18953 | EV71/ G082 | CVA16/ SZ05 | |
| Anti-PBS (control) | <32 | <32 | <32 | <32 | <32 |
| Anti-US/MO/14-18947 | 128 | 8192 | 256 | <32 | <32 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, C.; Zhang, X.; Dai, W.; Liu, Q.; Xiong, P.; Wang, S.; Geng, L.; Gong, S.; Huang, Z. A Mouse Model of Enterovirus D68 Infection for Assessment of the Efficacy of Inactivated Vaccine. Viruses 2018, 10, 58. https://doi.org/10.3390/v10020058
Zhang C, Zhang X, Dai W, Liu Q, Xiong P, Wang S, Geng L, Gong S, Huang Z. A Mouse Model of Enterovirus D68 Infection for Assessment of the Efficacy of Inactivated Vaccine. Viruses. 2018; 10(2):58. https://doi.org/10.3390/v10020058
Chicago/Turabian StyleZhang, Chao, Xueyang Zhang, Wenlong Dai, Qingwei Liu, Pei Xiong, Shuxia Wang, Lanlan Geng, Sitang Gong, and Zhong Huang. 2018. "A Mouse Model of Enterovirus D68 Infection for Assessment of the Efficacy of Inactivated Vaccine" Viruses 10, no. 2: 58. https://doi.org/10.3390/v10020058