1. Introduction
According to IPCC projections for the Mediterranean Basin, climate change will increase dryness and aridity due to warmer temperatures and reduced precipitation, mainly in summer [
1,
2] which will induce changes in fire regimes [
3] and increase the distribution of resilient species [
4]. Regional models for southeast (SE) Spain have predicted reduced rainfall and rises in temperature of about 20% and 4.5 °C, respectively, by the year 2100 [
5].
After wildfires, emergency actions and post-fire management are implemented to recover ecosystems and landscapes, and salvage logging is one of the commonest tools implemented in Spain [
6], and also worldwide [
7,
8]. The goal of implementation comprises economic, silvicultural and aesthetic objectives, as well as related benefits for the ecosystem, such as improved seedling recruitment due to mechanical scarification and soil movements [
9,
10]. However, several studies pointed to negative effects on the ecosystem, such as reducing growth and performance of regenerating Maritime pine (
Pinus pinaster Aiton) seedlings related to water stress and reduction of nutrient availability for plants [
11]. Also the abrupt insolation increase promoted by salvage logging increased summer mortality and reduced the growth of survival Aleppo pine (
Pinus halepensis Mill.) seedlings [
12], altered the plant-animal interaction [
13], and reduced carbon sequestration [
14] which could increase restoration costs [
15].
In fire-prone areas, mainly with poor site quality, the objective of adaptive forest management should focus on maintaining and promoting ecosystem protection (biodiversity, soil protection, carbon sink) in a global change scenario, which implies short-term monitoring to predict the forest structure, habitat quality, and carbon storage ratio [
16]. Scientific knowledge of plant species in the seedling stage, post-fire conditions, will help managers in their decision making, such as facilitating post-fire restoration and management tools and actions according to forest characteristics, e.g., tolerance of species in harsh conditions to drought [
17], reducing cost-effective restoration of degraded ecosystems. To reduce the risk of unsuccessful restoration practices, morphological and physiological traits of seedlings that relate to water stress should be known since they act as reliable predictors of survival and field performance [
18]. Mediterranean terrestrial landscapes develop adaptive traits or life strategies to withstand wildfires and drought periods. However, the new scenarios may diminish the resistance and resilience of ecosystems, which will have negative impacts on vegetation productivity [
19], density, and distribution [
20] and, ultimately, will promote desertification [
21]. The dominant trade-offs in fire regeneration strategies among Mediterranean plant species are resprouting and persisting seed banks that are enclosed in fruits (serotiny, aerial seed banks) or buried in soils (soil seed banks) and are, thus, insulated from heating [
22,
23].
The main aim of this research is based on one of the new selected research priorities that comprises the effects of interactions between drivers of global change in ecosystem functioning [
24]. We specifically address the interaction between drivers of global change and recent forest management practices in the short term, which characterize post-fire natural regeneration, by sampling experimental plots in burnt and managed (salvage logging six months after fire) Aleppo pine stands located in SE Spain. To check the effects and consequences of intensification of summer droughts and forest management, we recorded several morphological and ecophysiological characteristics of the main tree species and companion species. According to the predicted climate change scenario for the Mediterranean Basin, and specifically for SE Spain by the year 2100 [
5], we experimentally induced a 20% reduction in rainfall. Our purpose was to generate knowledge related to the effects of post-fire forest management in the short term linked to the intensification of summer droughts, to be implemented in adaptive forest management. To achieve this, we simulated scenarios and compared the effectiveness of reproductive strategies related to fire adaption. We characterized the ecosystem recovery recording recruitment of Aleppo pine seedlings and the three main companion species, all of which are representative of plant groups with different fire-adaption strategies. According to the plant trait database for Mediterranean Basin species (BROT) [
25] and including
Stipa tenacissima L. a post-fire resprouter,
Cistus clusii Dunal, an obligate seeder showing persistent soil seed banks whose germination is often stimulated by some fire-related factor (thermal scarification), and
Rosmarinus officinalis L., an obligate seeder with transient seed banks, but with no specific fire adaption, that behaves like a post-fire colonizer of opened insolated areas [
26].
2. Experimental Section
2.1. Study Area
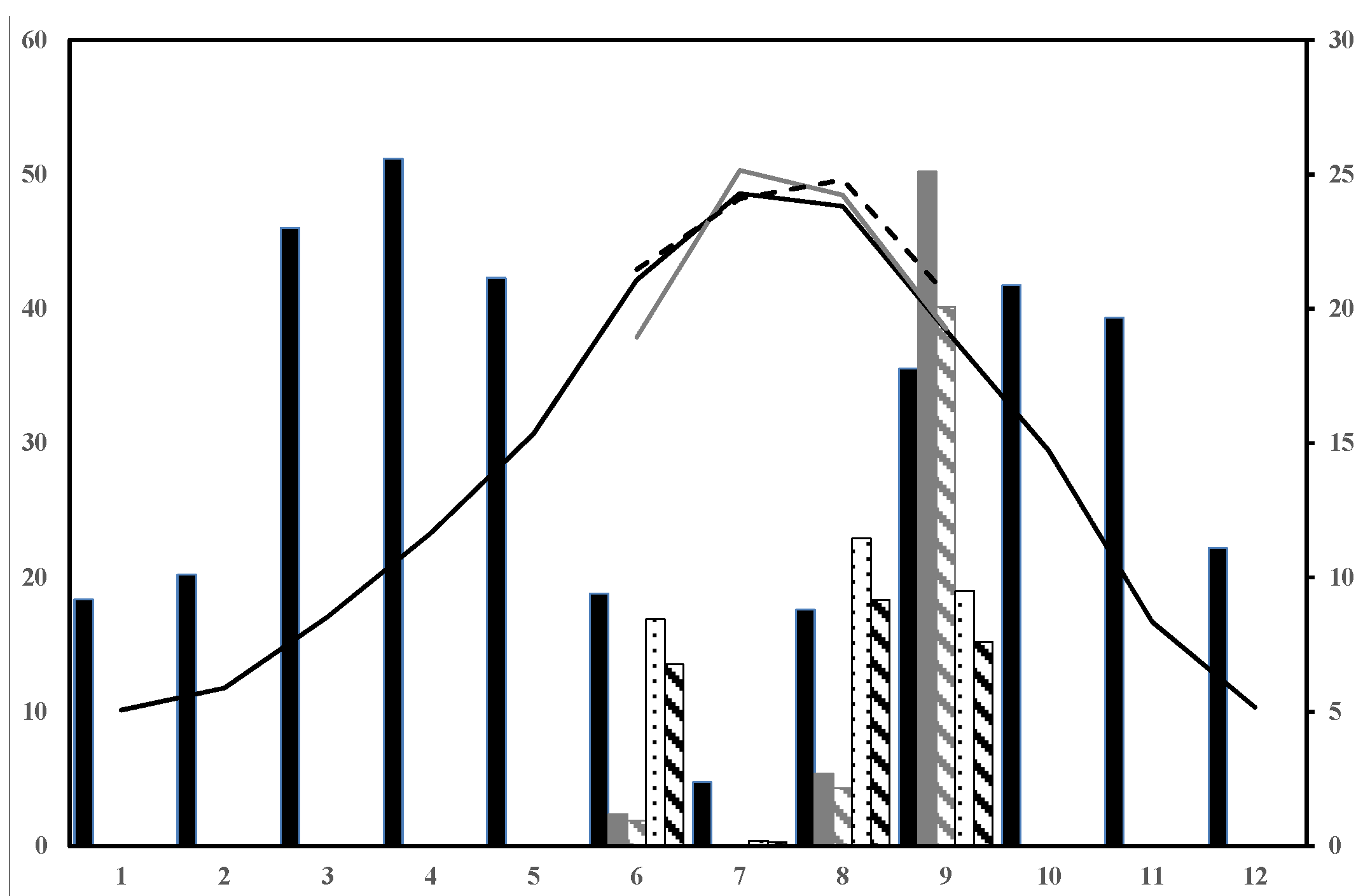
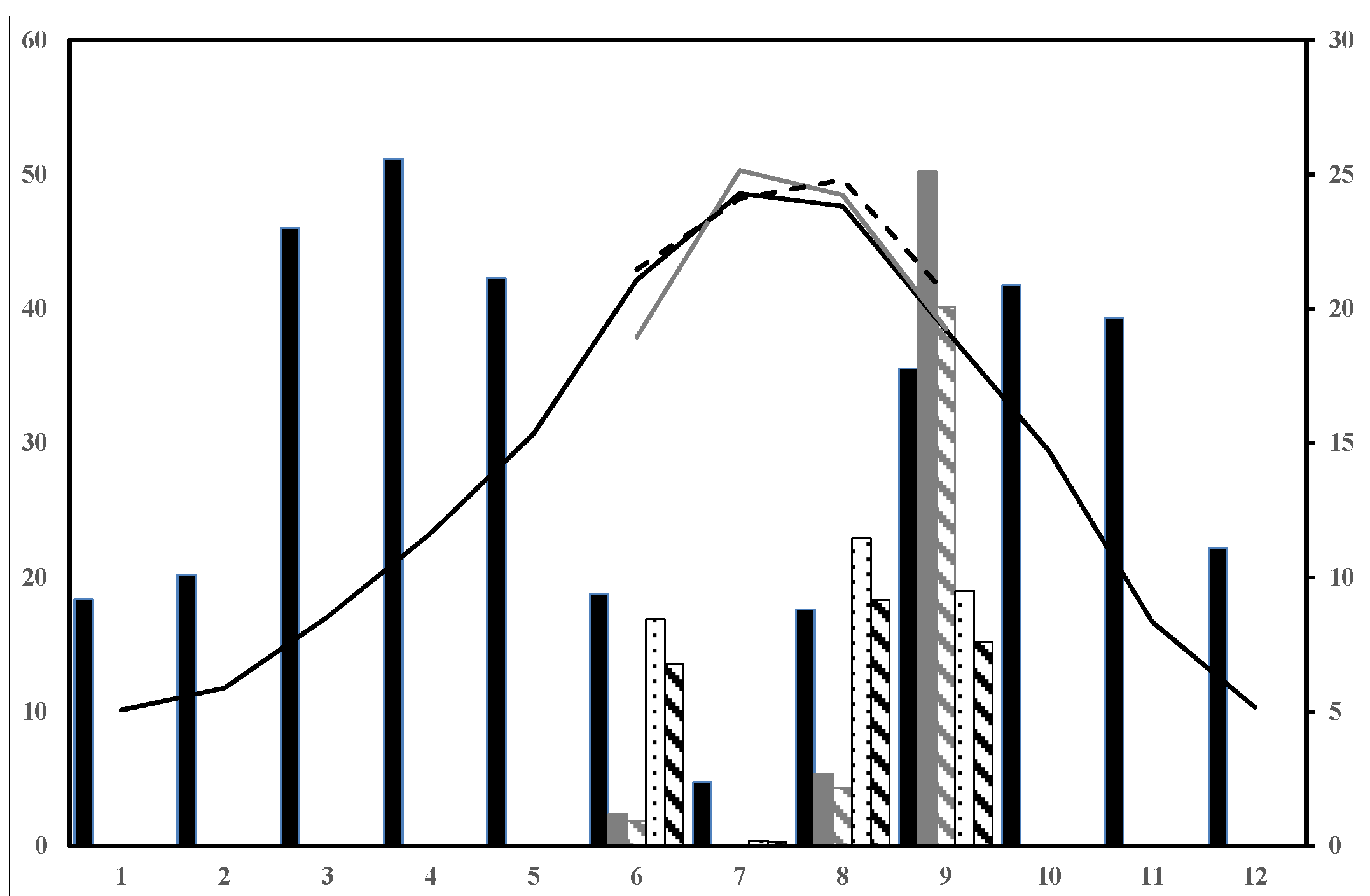
In summer 2009, a mid-high severity fire burnt 968 ha of Aleppo pine stands in the province of Albacete (SE Spain). The average precipitation and annual temperature values were 358.09 mm and 13.59 °C, respectively (based on the 25-year period data (1987–2012) provided by AEMET, the National Meteorological Agency). The drought period went from June to September (
Figure 1) when relative humidity was below 50%. The study period focused on the drought periods of the first two post-fire years (summer 2010 and 2011), which were warmer than the mean averaged values (0.20 °C and 0.63 °C, respectively). Specifically, three of the four months per drought period (June to September) showed higher temperatures than the mean averaged values for the 1987–2012 period, and ranged from 0.05 to 1.34 °C. The exceptions were June 2010 and July 2011, which were colder. The mean precipitation value for summer (76.77 mm) was also higher than that recorded during both periods studied (18.77 and 17.57 mm less in summer 2010 and 2011, respectively). We implemented techniques to induce a 20% reduction in rainfall, which is in accordance with the Mediterranean Basin projections, by promoting the dryer summers that gave 30.37 and 29.41 mm less in the summer of 2010 and 2011, respectively, than the mean averaged values for the 1987–2012 period (
Figure 1). The site is located at the upper mesomediterranean level and shows a dry ombroclimate. Natural potential vegetation corresponds to the series
Bupleuro rigidi-Querceto rotundifoliae sigmetum, a sclerophyllous oak forest [
27]. Before the fire, vegetation was paraclimacic Aleppo pine forests, whose main shrub companion species were Esparto grass (
Stipa tenacissima L.), Rosemary (
Rosmarinus officinalis L.), Rockrose (
Cistus clusii Dunal.), Thyme (
Thymus vulgaris L.), Prickly Juniper (
Juniperus oxycedrus L.), Holm oak (
Quercus ilex L. subsp.
ballota (Desf.) Samp.) and Hermes oak (
Quercus coccifera L.).
Figure 1.
Climogram representing monthly average precipitation (grey bars, left Y axis) and temperature (black line, right Y axis) based on the 25-year period data obtained from the Pozo-Cañada (Albacete, Spain) weather station. Black bars and line: precipitation and mean monthly temperature for 1987–2012; Dark grey bars and line: precipitation and mean monthly temperature for the 2010 drought period months. Bars filled in dark grey diagonal lines: precipitation, including induced drought for the 2010 drought period months. Dashed black line and dotted black bars: precipitation and mean monthly temperature for the 2011 drought period months. Bars filled in black diagonal lines: precipitation including induced drought for the 2011 drought period months.
Figure 1.
Climogram representing monthly average precipitation (grey bars, left Y axis) and temperature (black line, right Y axis) based on the 25-year period data obtained from the Pozo-Cañada (Albacete, Spain) weather station. Black bars and line: precipitation and mean monthly temperature for 1987–2012; Dark grey bars and line: precipitation and mean monthly temperature for the 2010 drought period months. Bars filled in dark grey diagonal lines: precipitation, including induced drought for the 2010 drought period months. Dashed black line and dotted black bars: precipitation and mean monthly temperature for the 2011 drought period months. Bars filled in black diagonal lines: precipitation including induced drought for the 2011 drought period months.
Six months after the fire event, January–February 2010, the Public Administration responsible for forest management carried out salvage logging as a silvicultural tool to facilitate future treatments, such as possible reforestations. Upon our request, five hectares were not treated and dead standing trees were not removed. We selected a study area that underwent two treatments: salvage logging (LOGGING) and control (UNCUT) (30 S; 629485 to 629817 E, 4322126 to 4322247 N; 920 m a.s.l.).
In Spring 2010, we randomly established three 25-metre linear transects, separated from each other by 5 m, in both study areas (LOGGING and UNCUT). Following the line intercept method [
28], we sampled to characterize pine seedling recruitment in spring 2010 and autumn 2011. Pine seedlings and individuals from the selected companion species intercepting lines (e.g.,
S. tenacissima, C. clusii, and
R. officinalis).were counted and their total height recorded.
2.2. Sampling Techniques
To monitor the recruitment and development of the selected species, we set five square plots (2 m × 2 m) in the control area (UNCUT) and ten square paired plots (2 m × 2 m) in the LOGGING area in summer 2010. We included drought as a key factor and randomly set paired plots, i.e., closer than 1 m of a plot no. submitted to induced drought (LOGGINGcontrol). We simulated rainfall exclusion (LOGGINGdrought) in a plot of similar characteristics (for visual details check supplementary material). All the individuals found in the square plots were located and tagged.
Similarly to previous studies [
29,
30], we built rain-exclusion shelters formed by a 2 × 2 m metal frame that supported V-shaped clear methacrylate bands with no UV filter (Barlocast; Faberplast S.L., Madrid, Spain), which covered 20% of the surface to intercept the same percentage of natural water supply by rain. To prevent surface runoff water from entering, we dug trenches (20 cm deep) along the entire shelters to intercept runoff water. Shelters were placed from early May to the end of September to simulate reduced precipitation during a drought period, as predicted for the Mediterranean Basin. To check ecosystem recovery, we selected the species that displayed greater abundance and coverage. The final selection included species with adaption traits to wildfires, a resprouter (
S. tenacissima) and two obligate seeders with different strategies (
C. clusii and
R. officinalis). Samplings were done in post-fire years 1 and 2 (2010 and 2011) at two different ages, before and after the drought period (early May before placing shelters (spring) and end of September after removing them (autumn)).To record soil water availability, morphological growth, and ecophysiological response of selected species, samplings were done in spring 2010, autumn 2010, spring 2011 and autumn 2011.
To estimate water stress in plants during the summer drought, we followed the pressure chamber method [
31] on leaves of
S. tenacissima and terminal shoots of
C. clusii and
R. officinalis using a pressure and a Compact Plant Water Status Console (model 3115P40G4, Soilmoisture Equipment Corp., Santa Barbara, CA, USA). We recorded the predawn leaf water potential (
ΨPD, MPa), which is related to the overall water status of individuals from each species before and after the drought period (early May and end of September) in 2010 and 2011. However, during the second sampling (in 2011) we found
P. halepensis and
Q. coccifera individuals which were included in the water stress sampling.
To characterize the morphological growth and effects of salvage logging and water stress, we recorded the total height (
H, mm) and cover (
COV, m
2) of the selected species by using field measuring tapes (10 m length, 1 mm accuracy). To avoid destructive sampling and to obtain an indirect classic growth measures approach, such as the relative growth ratio and the net assimilation ratio [
32], we calculated the
relative growth in height ratio (
ΔH) as the instantaneous growth in height increment divided by plant height at the beginning of the increment (by assessing growth efficiency). We also calculated the
coverage ratio (
ΔCOV) as the instantaneous change in the plant surface increment divided by the total surface at the beginning (by assessing the net assimilation ratio).
To assess the changes in ecophysiological response one and two years after the fire of the selected species, we monitored four individuals of each species in every monitoring plot. In each one, we carried out instantaneous recordings of gas exchange using a portable system (LI-6400XT; LI-COR, Environmental Division, Lincoln, NE, USA) equipped with a CO
2 and H
2O infrared gas analyzer (6400-07 Needle Chamber; LI-COR, Environmental Division, Lincoln, NE, USA). To characterize the salvage logging and water stress effects on ecophysiological processes according to [
32], we selected the following as target parameters: transpiration ratio (
E, mmol m
−2 s
−1); net photosynthetic ratio (
A, μmol m
−2 s
−1); stomatal conductance (
gs, mol H
2O m
−2 s
−1); water use efficiency, defined as the ratio of the carbon assimilation ratio (net photosynthetic ratio); transpiration ratio (
WUE =
A/
E) [
33]
. Sampling was done under light-saturation conditions for photosynthesis (photosynthetically active radiation (PAR) above 250 μmol m
−2 s
−1), with temperatures ranging from 20 °C to 26 °C, and the CO
2 atmosphere concentration within the 380–395 μmol mol
−1 CO
2 range. Measurements were related to the projected leaf area basis. The methods to characterize water status on plant seedlings were based on a punctual measurement and provided a record of photosynthetic rates at a single time, but were not a time-integrated measure of the water relations in the plant in the long term [
34].
2.3. Statistical Analysis
General linear models (GLM) were used to assess an overall model including significant variables and their interactions. We included the calculated parameters (ΨPD, ΔH, ΔCOV, E, A, and WUE) and considered the following to be categorical factors: sampling period (SAMPLING: spring 2010, autumn 2010, spring 2011, autumn 2011), post-fire management (TREATMENT: salvage logging, salvage logging plus induced drought, uncut) and species (SPECIES: S. tenacissima, R. officinalis and C. clusii). Since initial size of individuals can influence growth and modify the effects of induced drought effects which could be additive, we included an analysis for three different periods: initial stage (post-fire year 1, before summer and immediately after salvage logging), mid-stage, after one drought period (post-fire year 1 and immediately after the drought period), and a final stage (post-fire year 2 and immediately after the drought period). To obtain increments, we calculated growth in the first summer (differences from autumn and spring 2010), the second summer (differences from autumn and spring 2011), and from the whole study period (differences from autumn 2011 and spring 2010).
Kruskal—Wallis tests were used to check the mean significant differences among the independent variables. Significant differences between groups were tested by MANOVA using Tukey’s honest significant difference test, which corrects the experiment-wise error ratio by reducing the probability of making a type I error at the nominal level of significance. Values were log transformed to maximize Pearson’s correlation coefficient to achieve normality assumptions and homoscedasticity, however, values were showed untransformed, along with a standard error (±SE). Statistical analyses were performed by the Statgraphics CENTURION XV software using a critical p < 0.05.
To gain a better understanding, we conducted a principal components analysis (PCA), which included all the recorded morpho- and ecophysiological variables, using the mean values of the variables for species and treatments. The missing values were estimated covariance list wise (based on complete cases) and creating a standardized correlation matrix. The purpose of the PCA was to reduce the variables used by employing their linear combinations, and it accounted for most of the variability contained in the original data.
3. Results
3.1. Seedling Recruitment, Water Stress, and Survival
According to the line intercept method, Aleppo pine recruitment was very poor, although there were more pine seedlings in UNCUT (560 ± 100 seedlings ha−1) than in LOGGING area (280 ± 70 seedlings ha−1), but not significantly so. In addition, a few Q. coccifera individuals were found in UNCUT area but none in LOGGING area. The one-way ANOVA indicated that the pine seedlings that grew in UNCUT were significantly taller (44.0 ± 1.0 mm) than those in LOGGING (39.5 ± 0.5 mm). From the recordings obtained in the linear transects, we found a low P. halepensis seedling survival ratio in the UNCUT area (<15%), however, in the LOGGING area no P. halepensis seedlings were recorded two years after the fire, so null survival rate was obtained.
We found a small number of deceased individuals in the experimental areas after drought periods. Hence the GLM analysis corroborated that record, and showed the significance of factors treatment, species, and their interaction, but no significant effect on seedling survival was found. For the selected species, we obtained very high survival ratios for S. tenacissima and R. officinalis (>96%) and lower, but not significant, differences for C. clusii (90.85 ± 2.28%).
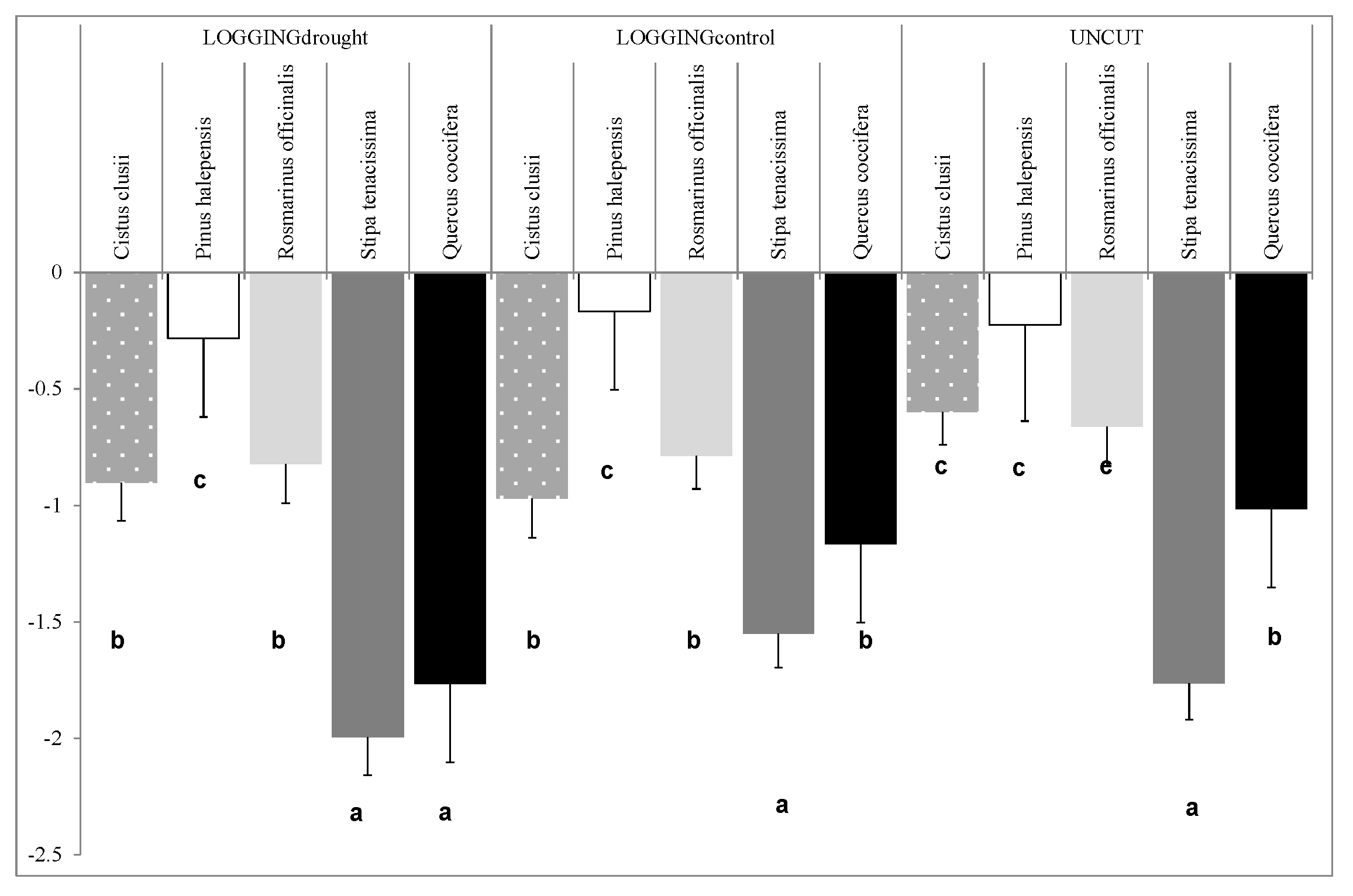
For the predawn leaf water potential (
ΨPD), we ran a MANOVA after including post-fire management, species, and sampling period as factors. The
F-test values showed no significant differences for silvicultural treatments or induced drought in post-fire year 1. However, we found significant differences for reproductive strategies, and obtained significantly lower values for resprouter
S. tenacissima (−2.115 ± 0.101 MPa) than for obligate seeders
R. officinalis (−1.010 ± 0.102 MPa) and
C. clusii (−1.311 ± 0.061 MPa), which agrees with other studies in Mediterranean species: resprouter strategies for coping with drought periods are based on avoiding water deficit, with high gradients of water tension to pull water up from soil to leaves, which allows quick recovery after water stress [
35]. For the sampling done in post-fire year 2, the
F-tests in the ANOVA test identified both post-fire management (sum of squares = 0.578;
F-ratio = 6.22;
p-value < 0.01) and species (sum of squares = 17.374;
F-ratio = 186.81;
p-value < 0.001) as significant main effects, in addition to their interaction (sum of squares = 2.853;
F-ratio = 15.34;
p-value < 0.001). After including the results for
P. halepensis and
Q. coccifera recorded for 2011, we found significant differences for resprouters and obligate seeders, and seeders obtained lower values for each treatment (
Figure 2). The
ΨPD values for
Q. coccifera differed significantly per treatment, with the highest in UNCUT and the lowest in LOGGINGdrought. No differences were observed for the other resprouter species,
S. tenacissima, and an obligate seeder,
P. halepensis for UNCUT and LOGGINGcontrol, which were higher than in LOGGINGdrought. The other obligate seeder species,
C. clusii and
R. officinalis, showed no significant differences in the treated area, but had higher values in UNCUT.
Figure 2.
Predawn leaf water potential (ΨPD, MPa) after the drought period 2 years after the fire (autumn 2011), with post-fire management and species as significant factors. Letters indicate significant differences between species within treatments by using Tukey’s HSD and p-value < 0.05.
Figure 2.
Predawn leaf water potential (ΨPD, MPa) after the drought period 2 years after the fire (autumn 2011), with post-fire management and species as significant factors. Letters indicate significant differences between species within treatments by using Tukey’s HSD and p-value < 0.05.
3.2. Morphological Responses to Drought and Salvage Logging
We studied the effects on productivity, recorded as height and coverage growth. We checked that initial H and COV did not differ between the selected areas to reinforce the ecological meaning of their responses to the treatments. Indeed, values from the initial stage for H submitted to a multifactor ANOVA, showed significant differences for interspecies (141.95 ± 5.45 mm for C. clusii; 91.14 ± 5.31 mm for R. officinalis; 191.83 ± 5.858 mm for S. tenacissima), independently of treatment. Also, COV values were not influenced by treatment immediately after salvage logging, but merely showed significantly higher values for the resprouter S. tenacissima (0.112 ± 0.009 m2) than obligate seeders species R. officinalis and C. clusii (0.004 ± 0.001 and 0.005 ± 0.001 m2, respectively).
However, the last recording taken two years after the fire indicated that the two study factors, post-fire management (p-value < 0.001) and species (p-value < 0.001), and their interaction (p-value < 0.001), had a significant influence on both variables H and COV. We found that the S. tenacissima individuals were the tallest in UNCUT (659.47 ± 30.60 mm), but the tallest ones in the treated areas were C. clussi (360.22 ± 27.81 mm in LOGGINGdrought and 412.42 ± 23.22 mm in LOGGINGcontrol).
The COV values for resprouter S. tenacissima showed significant differences to those for obligate seeders species, and were significantly higher in UNCUT (0.385 ± 0.020 m2 for S. tenacissima, 0.024 ± 0.002 m2 for R. officinalis and 0.022 ± 0.002 m2 for C. clusii) than in the LOGGING area (0.037 ± 0.011, 0.018 ± 0.009 and 0.014 ± 0.009 m2, respectively for the species in LOGGINGcontrol; 0.026 ± 0.012, 0.007 ± 0.001 and 0.008 ± 0.001 m2 respectively for the species in LOGGINGdrought).
Since initial size can influence growth, we included instantaneous growth for initial size. We also ran multifactor ANOVAs for ΔH and ΔCOV for the initial, mid and final stages. For ΔH in the initial stage, we found that factors treatment (sum of squares = 1.70; F-ratio = 9.31; p-value < 0.001), species (sum of squares = 7.77; F-ratio = 3.88; p-value < 0.001), and their interaction (sum of squares = 2.40; F-ratio = 0.60; p-value < 0.001), were significant. Thus after the summer of post-fire year 1, we obtained a significantly higher ΔH for resprouter than obligate seeders. However, the ratio indicated that resprouter S. tenacissima grew significantly taller in the LOGGING area (both control and drought, 0.67 ± 0.04 and 0.48 ± 0.04, respectively) than in UNCUT (0.16 ± 0.07). Nevertheless, obligate seeder species displayed the exact opposite as a significantly higher growth ratio was obtained for C. clusii in the UNCUT area (0.24 ± 0.07) and a significantly lower one was observed for R. officinalis in the area with induced rainfall exclusion (LOGGINGdrought, 0.08 ± 0.03). There were no significant differences for the other factors. We obtained significant differences for ΔCOV based on factors treatments and species. The highest ratio went to obligate seeders, specifically for R. officinalis, which was independent of treatment (0.98 ± 0.08), while C. clusii increased more in the UNCUT area (1.17 ± 0.19) than in the LOGGING area (0.42 ± 0.09). The resprouter increased the least (0.34 ± 0.08), but was maximized in the UNCUT areas (0.76 ± 0.19) rather than in LOGGING (0.14 ± 0.11).
The results varied when we considered variations for year 2 (by adding two dry periods with induced rainfall exclusion). For both variables ΔH and ΔCOV, the F-tests showed a significant influence for all the factors and their interaction. During this period, we once again found a significantly higher ΔH of resprouter than obligate seeders. Conversely, the most marked growth (height) was found in the UNCUT area (0.61 ± 0.05). Seeders R. officinalis and C. clusii not only gave a significantly higher ratio in UNCUT (0.20 ± 0.05 and 0.09 ± 0.05, respectively), but also underwent a marked decrease in LOGGINGdrought (−0.02 ± 0.04 and −0.04 ± 0.05, respectively). For ΔCOV, the highest increase went to the resprouter in the UNCUT areas (1.11 ± 0.13), and no significant differences appeared in the LOGGING areas. Obligate seeders obtained higher values in the UNCUT areas than in LOGGINGcontrol, while those in LOGGINGdrought obtained the lowest values for both species.
For ΔH in the final stage, the F-tests in the multifactor ANOVA test identified only the post-fire management and species interaction to be a significant factor (sum of squares= 9.04; F-ratio = 2.91; p-value = 0.02). In fact the growth ratio of height in S. tenacissima and C. clusii did not significantly differ in the LOGGING area (1.54 ± 0.19 and 1.58 ± 0.20 the former; 1.64 ± 0.18 and 1.65 ± 0.15 the latter in LOGGINGdrought and LOGGINGcontrol, respectively), but was higher than UNCUT (0.96 ± 0.20 and 1.35 ± 0.21, respectively). Yet the ratio for R. officinalis was significantly higher in UNCUT (1.80 ± 0.20) than in LOGGINGcontrol (1.75 ± 0.14), and LOGGINGdrought gave the significantly lowest value (1.22 ± 0.14). For ΔCOV, only factor species was identified by the F-tests as being a significant factor (sum of squares = 670.74; F-ratio = 335.37; p-value < 0.001). Obligate seeders gave a higher growth ratio of coverage (5.93 ± 0.77 and 6.88 ± 0.69 for R. officinalis and C. clusii, respectively) than resprouters (2.63 ± 0.83).
Higher
ΔH and
ΔCOV have been related to the root:shoot ratio, higher in resprouters [
36], which was related to increased water potential and leaf conductance after fire in resprouters. Resprouters also allocated a bigger proportion of biomass to stems, while obligate seeders with drought tolerance, described as post-fire seeders such as
R. officinalis, increased the root biomass in young individuals to achieve a better soil water supply to photosynthetic organs [
37]. Some studies have focused on differences in fire regeneration strategies and have concluded that the key aspect of physiological responses to drought is age (juvenile or mature individuals) since seedlings of obligate seeders are better adapted to water stress under post-fire conditions than seedlings of resprouter species [
38].
3.3. Ecophysiological Responses to Drought and Salvage Logging
The multifactor ANOVA test decomposed the variability of
A and
E into contributions due to factors treatment, species, and interaction, which showed a statistically significant effect on both variables at the confidence level (
Table 1). Both variables obtained similar results, and gave the highest values in the UNCUT area of both, which were significantly lower for obligate seeders than for resprouters. We found no significant differences in the LOGGINGcontrol area when we compared values
A and
E for species, while the lowest values were recorded for
C. clusii and the highest for
S. tenacissima in the other experimental areas (UNCUT and LOGGINGdrought) (
Figure 3).
Table 1.
F-tests identified post-fire management, induced drought, species and the interaction for the net photosynthesis ratio (A, μmol m−2 s−1), transpiration ratio (E, mmol m−2 s−1), stomatal conductance (gs, mol H2O m−2 s−1) and water use efficiency (WUE= A/E) as significant factors (marked with asterisks) by using a critical p-value<0.05. Asterisk indicates the significant influence of the factor.
Table 1.
F-tests identified post-fire management, induced drought, species and the interaction for the net photosynthesis ratio (A, μmol m−2 s−1), transpiration ratio (E, mmol m−2 s−1), stomatal conductance (gs, mol H2O m−2 s−1) and water use efficiency (WUE= A/E) as significant factors (marked with asterisks) by using a critical p-value<0.05. Asterisk indicates the significant influence of the factor.
| Net photosynthesis ratio | Transpiration ratio | Stomatal conductance | Water use efficiency |
|---|
| (A, μmol m−2 s−1) | (E, mmol m−2 s−1) | (gs, mol H2O m−2 s−1) | (WUE= A/E) |
|---|
| Sum of Squares | F-Ratio | p-Value | Sum of Squares | F-Ratio | p-Value | Sum of Squares | F-Ratio | p-Value | Sum of Squares | F-Ratio | p-Value |
| TREATMENT | 157708 | 27.44 | <0.001* | 1.222 | 33.10 | <0.001* | 1.11 | 11.29 | <0.001* | 0.081 | 6.31 | <0.01* |
| SPECIES | 91772 | 15.97 | <0.001* | 0.638 | 17.28 | <0.001* | 0.03 | 0.34 | 0.715 | 0.015 | 1.16 | 0.32 |
| TREATMENT × SPECIES | 107081 | 9.32 | <0.001* | 0.761 | 10.30 | <0.001* | 6.66 | 33.93 | <0.001** | 0.066 | 2.55 | 0.04* |
We also ran a multifactor ANOVA for
gs and
WUE, and checked the significant influence of factors post-fire treatment and the post-fire treatment × species interaction. Both seeder species showed similar patterns for
gs, and no significant differences were found in the LOGGING area, but
gs was higher in the UNCUT area. However, resprouter
S. tenacissima showed the lowest
gs values in the UNCUT area (−0.163 ± 0.026 mol H
2O m
−2 s
−1), but the highest ones in LOGGINGcontrol (0.234 ± 0.020 mol H
2O m
−2 s
−1). Conversely, obligate seeder species presented less significant
WUE in the UNCUT and LOGGINGcontrol areas, which were higher in LOGGINGdrought (0.478 ± 0.015 and 0.417 ± 0.014 for
C. clusii and
R. officinalis, respectively). We observed the lowest WUE value for the resprouter in the UNCUT area (0.377 ± 0.024), but no significant differences were obtained in the areas treated after the fire (0.441 ± 0.018 and 0.420 ± 0.013 corresponding to the LOGGINGcontrol and LOGGINGdrought areas). Differences were not marked, which may indicate that drought conditions were not extreme [
37].
Figure 3.
Multifactor ANOVA test which includes the mean value and standard error according to the significant factors using Tukey’s HSD for the net photosynthesis ratio (A, μmol m−2 s−1), transpiration ratio (E, mmol m−2 s−1), stomatal conductance (gs, mol H2O m−2 s−1) and water use efficiency (WUE= A/E) as significant factors with a critical p-value of <0.05. Letters indicate significant differences by groups including species interacting treatments.
Figure 3.
Multifactor ANOVA test which includes the mean value and standard error according to the significant factors using Tukey’s HSD for the net photosynthesis ratio (A, μmol m−2 s−1), transpiration ratio (E, mmol m−2 s−1), stomatal conductance (gs, mol H2O m−2 s−1) and water use efficiency (WUE= A/E) as significant factors with a critical p-value of <0.05. Letters indicate significant differences by groups including species interacting treatments.
3.4. Morpho-Ecophysiological Effects of Induced Drought and Salvage Logging
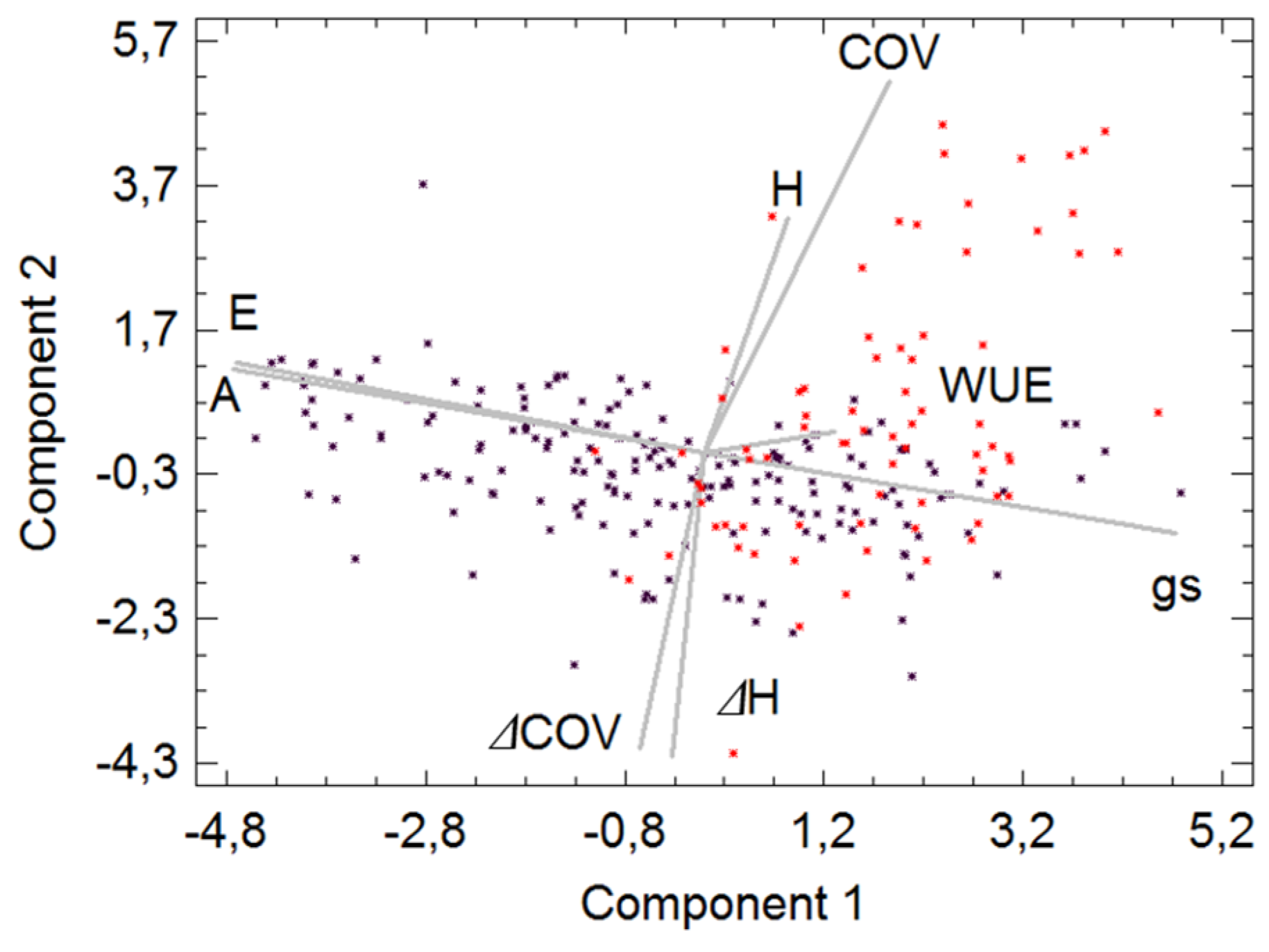
Finally, a PCA (
Figure 4) including the recorded variables was carried out. The variables selected reflected differences in morphological and physiological traits between the studied drought and post-fire treatment scenarios. We found a three-component solution (eigenvalues > 1), however, we simplified to a two-component solution in spite of losing explained variance but improving the significance. The first component accounted for 39.38%, while the second explained 20.82% and 60.20% of the variability contained in the original data accumulated. The main variation trend, marked by the first PCA component, ecophysiological water use variables, was evidenced by the relationship of
A,
E and
gs. The PCA highlighted was related to a positive trade-off between
A and
E, but a negative one with
gs, and evidenced a clear separation of resprouters and seeders, with the latter linked to this component. It implied greater resistance to water stress, but less annual growth. The second component indicated a link between the morphological traits. The PCA displayed an association between species, with resprouters associated with a larger size of individuals and higher annual growth ratios.
Figure 4.
A principal component analysis including the recorded variables: total height (H, mm), total cover (COV, m2), relative growth in height ratio (ΔH), coverage ratio (ΔCOV), transpiration ratio (E, mmol m−2 s−1), net photosynthetic ratio (A, μmol m−2 s−1), stomatal conductance (gs, mol H2O m−2 s−1) and water use efficiency (WUE). Black dots: standardized values for obligate seeders (R. officinalis and C. clusii); Red dots: standardized values for resprouter (S. tenacisiima). The first component accounted for 39.38% and the second 20.82%, both accumulated 60.20% of the total variability contained in the original data.
Figure 4.
A principal component analysis including the recorded variables: total height (H, mm), total cover (COV, m2), relative growth in height ratio (ΔH), coverage ratio (ΔCOV), transpiration ratio (E, mmol m−2 s−1), net photosynthetic ratio (A, μmol m−2 s−1), stomatal conductance (gs, mol H2O m−2 s−1) and water use efficiency (WUE). Black dots: standardized values for obligate seeders (R. officinalis and C. clusii); Red dots: standardized values for resprouter (S. tenacisiima). The first component accounted for 39.38% and the second 20.82%, both accumulated 60.20% of the total variability contained in the original data.
4. Discussion
The literature describes that studied species displayed differences in root architecture according to reproductive strategies [
37,
39] and developed adaptations to arid and fire-prone environments, such as the ability to maintain humidity in leaves, a rigid cell wall to reduce transpiration, stomatal control, and protection from photosynthetic activity [
35,
40]. However, knowledge on their response and tolerance to silvicultural treatments in the post-fire seedling stage is still lacking.
Our results agreed with previous studies describing greater water stress in the seedlings of seeder species than in resprouters because of the size of their root systems since it influences the availability of deeper soil water reserves in summer [
36]. According to [
38], resprouters lose more turgor at lower water potentials than seeders. The inter and intraspecific competition of seeder species, mainly in young stages after fire showing high seedling densities, can also diminish water availability which agreed with the results found in this study and others developed in SE Spain [
41]. In addition, the effects of low water availability were more important during drought periods for seedlings of
S. tenacissima and
C.clusii [
41]. Other resprouter species, such as oaks, suffered lower water stress during droughts due to their high root:shoot ratio and the evolutive ecophysiological adaptations [
42], similarly to that described for
Q. coccifera in our study.
Drought increased water use efficiency, reducing net photosynthesic rate and CO
2 assimilation in the Mediterranean species studied. Nevertheless, some authors reported that seeders maintained higher gas exchange ratios, stomatal conductance, and transpiration but lower water-use efficiency than resprouters [
38], but our results showed that post-fire management interacted significantly. According to [
43],
R. officinalis and other Mediterranean obligate seeder species reduced photosynthetic productivity and leaf area reduction due to the intensification of summer droughts which agreed with the results found for the logged area. In addition, we recorded lower stomatal conductance of seeders in the three treatments considered which could be due to the higher root:shoot ratio of resprouters [
36]. Hence the differences found in this article could be explained by water stress due to differences related to various factors, such as simulated drought and response of species but others should be studied, mainly inter- and intraspecific competition, soil water content, and the size of root systems. Furthermore, those species adapted to drought periods, and recovered productivity after summer and maintain stomatal conductance at a low leaf water potential in relation to a quick stomatal response [
44]. Our results disagreed with [
45], since
WUE for
S. tenaccisima was negatively related to individual size and drought conditions, being lower in higher tussocks and higher in logged areas including drought induction. This could imply differences in interspecific competition, showing seeders lower survival rates, since our results showed that
E values were also higher for resprouters under drought conditions in areas where dead standing trees remained. Seeders occurring in logged areas were more sensitive to drought conditions because the gas exchange ratios of seeders dropped as the water potential lowered, which could be induced by stomatal closure [
38].
Our results revealed that the main species comprising the plant community were under higher water stress in the areas where early salvage logging was carried out. The transpiration values were found lower in those individuals located in areas where dead wood was removed, which could be due to stomatal closure and a fall in photosynthetic activity (mainly for obligate seeders). Protection of dead standing trees for young seedlings implies lower surface solar radiation and temperature extremes, increased soil moisture and reduced water deficit (mainly for seeders) provided by standing dead trees and coarse woody debris [
8,
12]. In addition, intense drought periods reduce transpiration which promotes stomatal closure and those effects may be seasonally accumulative. The wood removal also has a detrimental effect on the soil characteristics and on the ecophysiological performance and growth of naturally regenerating pine seedlings [
11,
46].
Consequently, the ecosystem’s response was modified by short-term silvicultural treatments, which may alter vegetation distribution patterns, influence vulnerability and resilience, and could even depend on fire regimes and soil characteristics in the long-term [
46,
47]. Therefore, post-fire management, plant community and ecosystem structure will increase their relevance in predicted scenarios with more frequent and severe drought periods and wildfires [
1]. According to our findings, this situation was critical in the first months after a fire since plant coverage prevents soil erosion, flooding and runoff [
48], while it can significantly affect growth and ecophysiological variables during longer periods [
41]. Some morphological or ecophysiological parameters can also vary depending on forest management [
49], intraspecific competition (seedling density, spatial distribution, and size of individuals) [
50], and seasonality [
45].
We found that the predawn leaf water potential measured in a Scholander chamber was related to morpho- and ecophysiological processes which were directly influenced by water stress and soil water content [
44]. Therefore, attenuating the water stress of plants in response to soil water depletion may be crucial in Mediterranean species, mainly in climate change scenarios since variations in the leaf water potential influence survival, growth and ecophysiological variations, depend on the species and age of individuals [
38]. The short-term survival and ecophysiological results indicate changes in community composition if salvage logging is carried out six months after fire because differential drought resistance between obligate seeders and resprouters induced differential survival [
36] and inferred morpho- and ecophysiological processes after severe summer droughts, similarly to IPPC projections predictions,
i.e., water use efficiency usually increases during drought which is a poor strategy in habitats with high intra- and interspecific competition for water since it will be used by competitors [
45].
Other studies included climate change projections which have been proved to introduce great uncertainties with regard to sustaining current management practices in Mediterranean terrestrial ecosystems [
28]. To cope with them, adaptive restoration approaches have been suggested, such as improving quality of plant production in forest nurseries, techniques for optimizing rain use efficiency in plantations, and exploring native plant species, including provenances (genotypes) that show resilience to fire and water use efficiency [
18]. Another study included in the post-fire management, interactions between species (facilitation/competition) [
51] and compensation of mortality with the higher density of seeders. In that study, they achieved the pre-fire balance between obligate seeders and resprouters in the community [
36] including changes in the interactions that depend on the scenario; e.g. environmental conditions: water availability or disturbances (fire events) and effects of forest restoration. Therefore, those ecological interactions and effects of fire severity should be taken into account in future researches to counterbalance changes in the Mediterranean terrestrial ecosystems [
28]. Consequently, tools implemented in adaptive forest management have to include effects in the long-term [
52], according to projections, but also include the variability of interaction in the short-term balance, mainly due to fluctuating environmental conditions [
51]. In these scenarios, small seedlings, under effects of salvage logging and induced water stress, displayed differences in inter- and intra-specific competence. It suggested stronger effects under the most likely climate change scenario, that of increased drought and fire regimes in the Mediterranean Region [
45]. These trends, which have been recorded for other Mediterranean plant communities under extreme drought conditions [
53], have confirmed that intensification of summer droughts reduces net primary productivity and the development of individuals, but negative effects can be mitigated by applying proper forest management that has been adapted to the given scenario [
54,
55].
5. Conclusions
The post-fire regeneration strategies (
i.e., resprouting
vs. seeding) are adaptions to Mediterranean conditions but display different physiological behaviors [
37], which permit co-existence in fire- and drought-prone Mediterranean ecosystems [
20]. In this study, they developed different responses to water availability, seeders adapted to deal with high water limitation and highly variable conditions, but take advantage of periods with high water availability. Conversely, resprouter species are better adapted to more stable water availability conditions as they maintain a deep root system, but resist water shortages in summer [
38].
Knowledge on evolutionary adaptation to different regeneration microsites (post-fire gaps prone to drought (seeders) and sheltered conditions under mature vegetation (obligate resprouters) have to be included in adaptive restoration approaches, including dynamic vegetation models which aim to predict biodiversity and community composition related to land use change, climate change, and forest management. In this way, silvicultural treatments such as salvage logging, applied as emergency actions after fire, may imply distinct effects from those that focused on the goals. In our case study, early dead wood removal (first winter after fire) diminished the vigor and growth of pine seedlings of scarce recruitment, presumably because post-fire wood removal increases water stress and reduces nutrient availability in contrast with the amelioration of microsite/microclimate conditions by the presence of residual burnt wood and decomposition of woody debris, similar to Maritime pine stands in SE Spain [
11]. Simultaneously under extreme drought events, the main tree species, but also companion species (including both resprouters and obligate seeders), were affected, and productivity could be lower due to water stress, reducing plant growth in coverage which affects the ecosystem by reducing soil protection in the short term [
48]. Salvage logging is a post-fire management tool in which time of application should be taken into account, especially in areas with no economic benefits from timber but which include non-use benefits. In fact, a delay could prevent changes in the ecological succession stage. However, it should be corroborated and studied in depth, implementing more scenarios, including induced drought and increase of temperature related to different post-fire management. Also a wider seedling density range should be included to assess the suitability of how adaptive management tools would improve the recovery of burned ecosystems.






{kind=link}
{kind=link}
{kind=link}
{kind=link}