Nitrogen and Carbon Biogeochemistry in Forest Sites along an Indirect Urban–Rural Gradient in Southeastern Michigan


Abstract
:1. Introduction
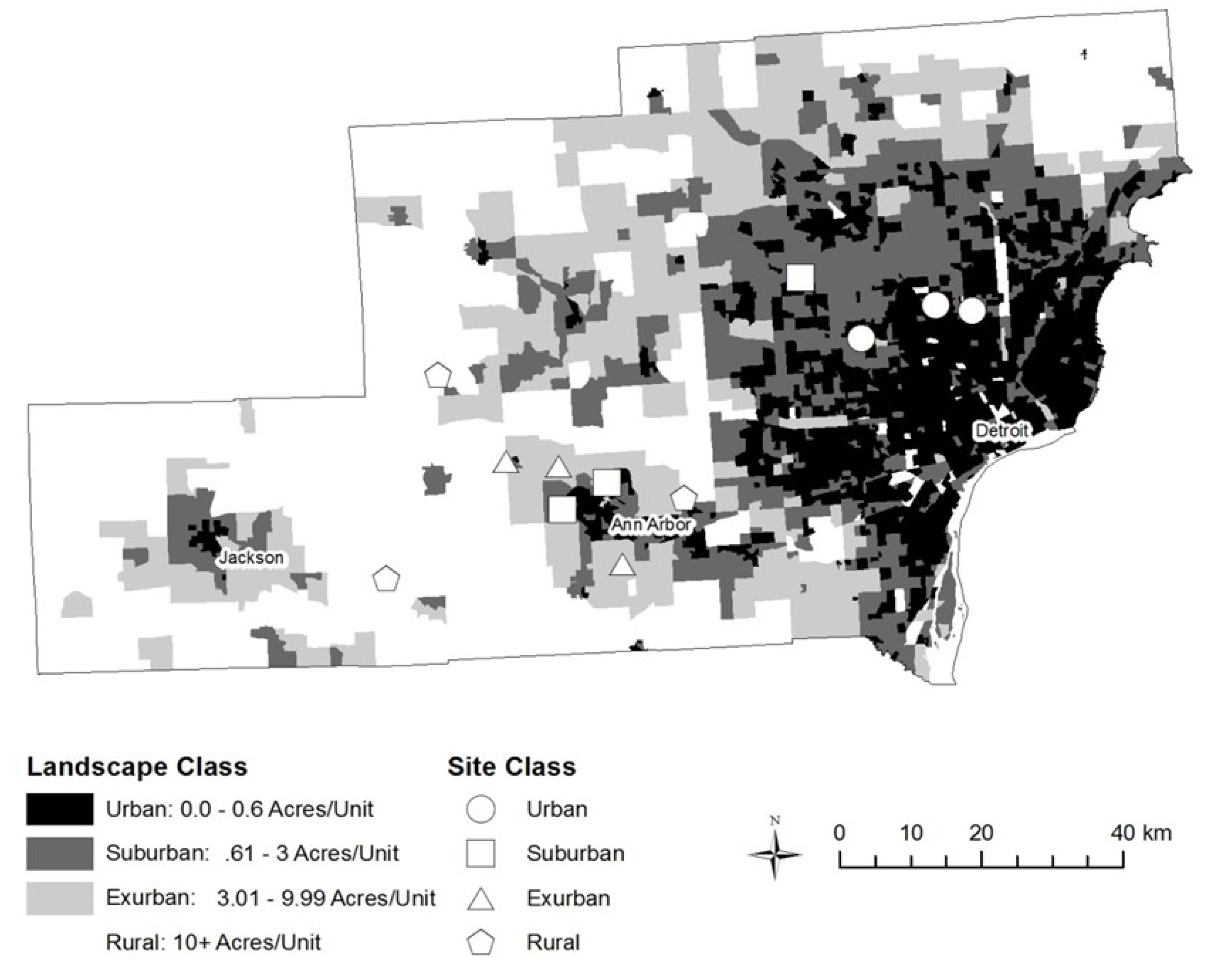
2. Experimental Section

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Housing Density | Site Name | Site Code | Size (acres) | Plot mean Elevation (m) | Designated Preserve | Oldest date of surrounding development |
|---|---|---|---|---|---|---|
| Urban | Suarez Friendship Woods | SFW | 37.5 | 631 | n/a | 1921 |
| Urban | Tenhave Woods at Quickstad Park | TW | 20 | 662 | 1955 | 1918 |
| Urban | Valley Woods | VW | n/a | 670 | 1972 | 1951 |
| Suburban | West Bloomfield Woods | WBW | 162 | 943 | 1988 | 1938 |
| Suburban | Saginaw Woods | SAG | 80 | 987 | 1903 | 1925 |
| Suburban | Black Pond Woods | BPW | 34 | 861 | 1991 | 1948 |
| Exurban | Smith Woods | SW | 17 | 874 | 2006 | 1958 |
| Exurban | Pittsfield Preserve | PP | 535 | 833 | 2002 | 1954 |
| Exurban | Osborne Mill | OSM | 39 | 814 | 1981 | 1928 |
| Rural | E.S. George Reserve | ESG | 1300 | 957 | 1930 | 1920 |
| Rural | Lefurge Woods Preserve | LW | 325 | 813 | 1995 | 1968 |
| Rural | Nan Weston Nature Preserve | NWN | 249 | 940 | 1983 | 1970 |
2.1. Sampling Design
2.2. Analysis of Field Samples
2.3. Statistical Methods
3. Results and Discussion
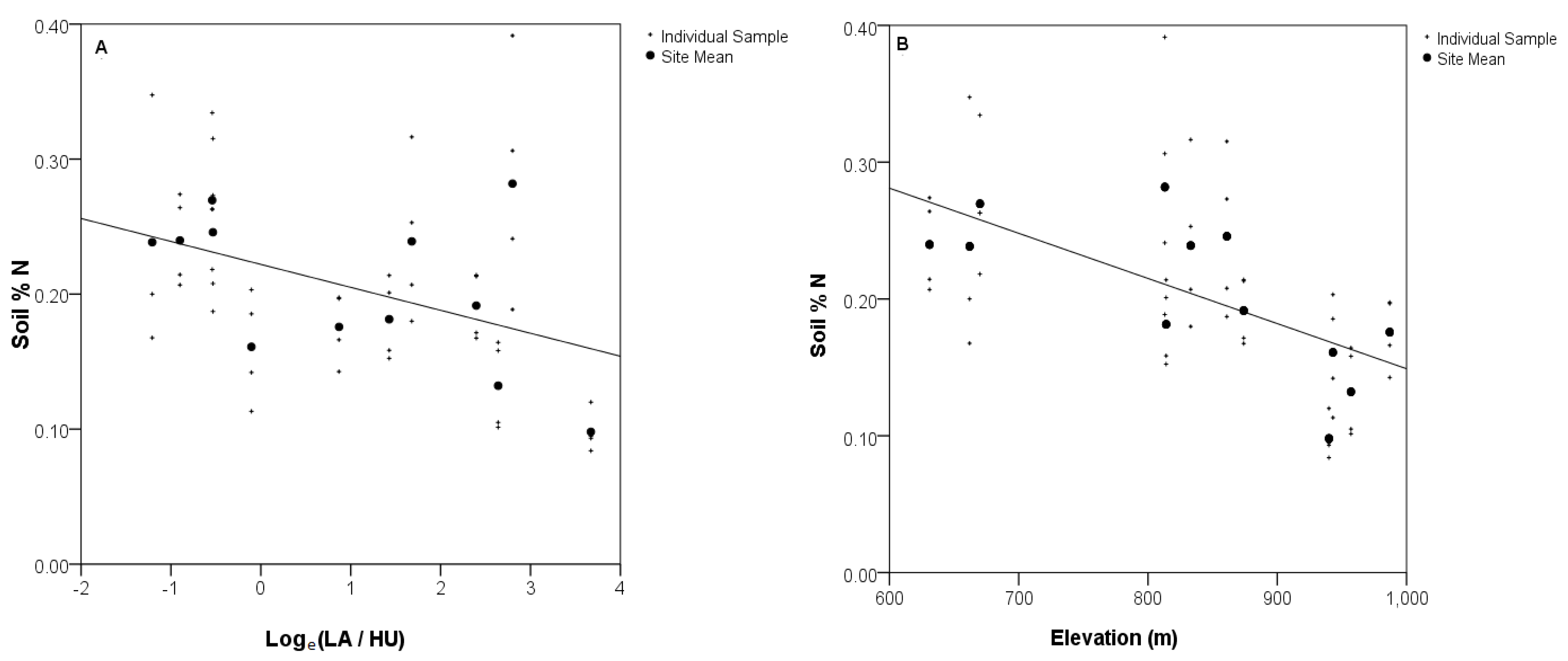
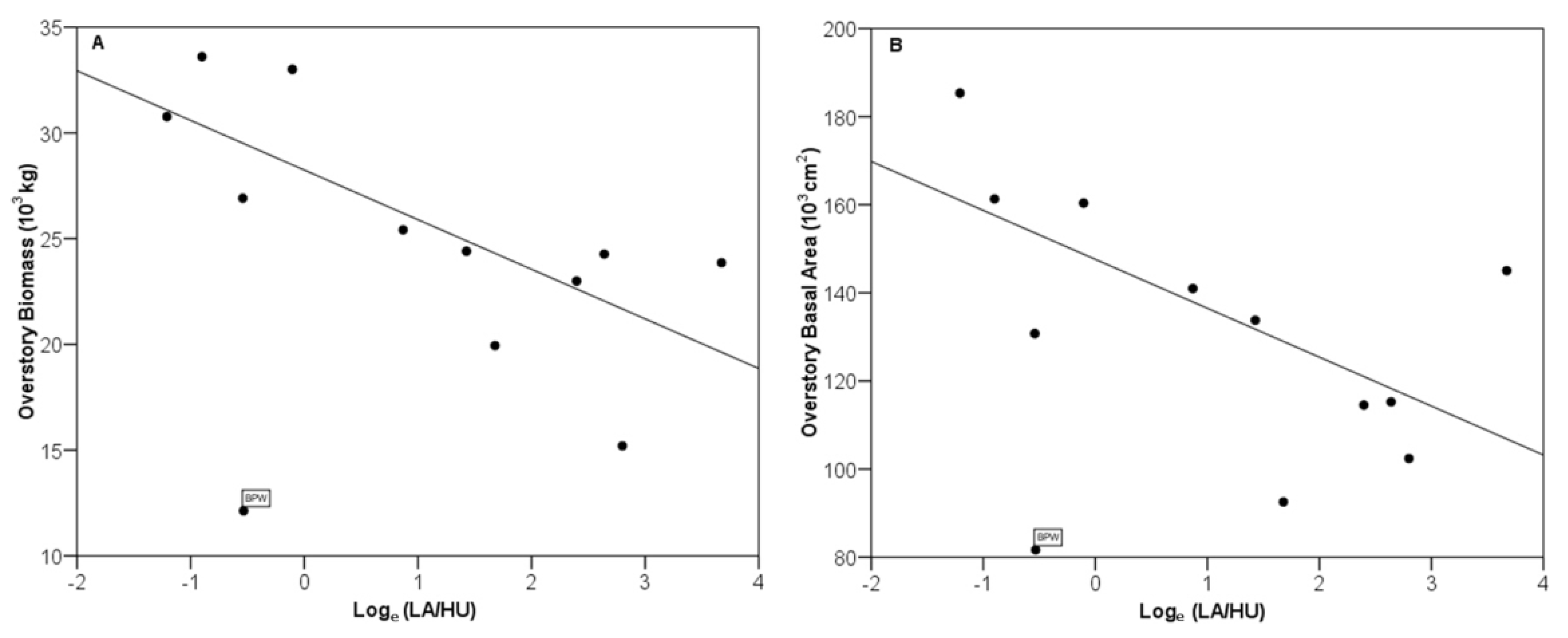
3.1. Relationship between Soil N and Surrounding Housing Density

| Independent variable(s) | Coefficients (p) | R2 | Model significance (p) |
|---|---|---|---|
| Loge LA/HU | −0.02 (0.09) | 0.27 | 0.08 |
| % Sand | −1.3 × 10−3 (0.17) | 0.09 | 0.17 |
| Loge LA/HU × % Sand | −3.3 × 10−4 (0.01) | 0.47 | 0.01 |
| Elevation | 3.3 × 10−4 (0.01) | 0.52 | 0.01 |
| Elevation | 3.8 × 10−4 (<0.01) | 0.82 | <0.01 |
| % Sand | −1.8 × 10−3 (<0.01) | ||
| Wetness | 0.03 (0.01) | 0.72 | 0.01 |
| % Sand | −2.2 × 10−3 (0.01) | ||
| Aspect | −1.2 × 10−4 (0.199) |
| Independent Variable (s) | Coefficients ( p) | Model AIC
(smaller is better form) | χ2 Fit Test:
improvement over null (p value) |
|---|---|---|---|
| Intercept Only (null) | ------------ | −131.47 | ------------ |
| Loge LA/HU | −0.02 (0.01) | −141.31 | <0.01 |
| % Sand | −1.0 × 10−3 (0.06) | −133.03 | 0.16 |
| Loge LA/HU × % Sand | 3.4 × 10−4 (<0.01) | −144.63 | <0.01 |
| Elevation | −2.7 × 10−4 (<0.01) | −137.37 | 0.02 |
| Elevation | −3.3 × 10−4 (<0.01) | −148.09 | <0.01 |
| Sand | −1.5 × 10−3 (<0.01) | ||
| Wetness | 0.01 (<0.01) | −138.92 | 0.01 |
| % Sand | −1.6 × 10−3 (<0.01) | ||
| Aspect | -9.9 × 10−5 (0.12) |
3.2. Relationship between Soil C/N Ratio and Surrounding Housing Density
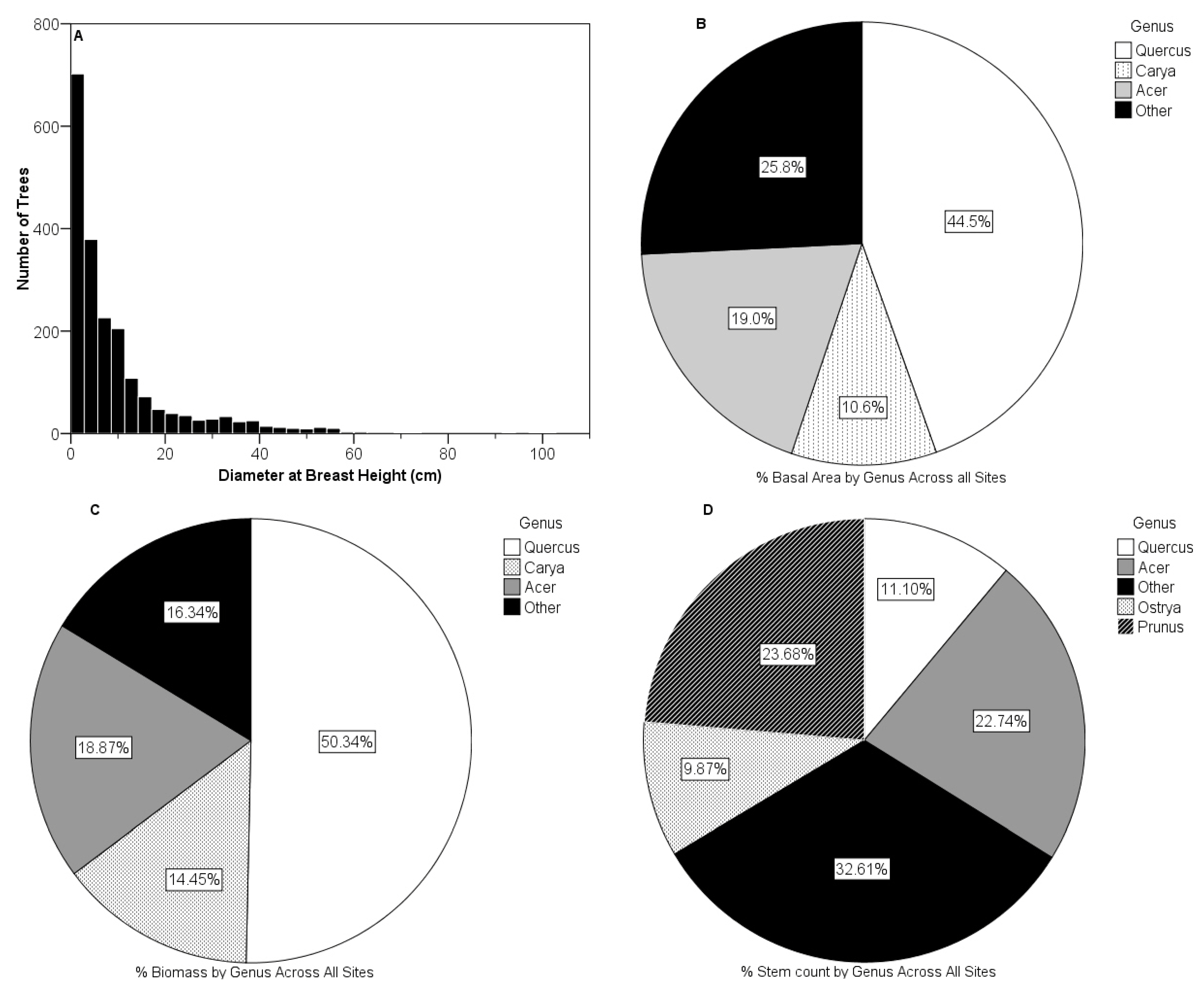
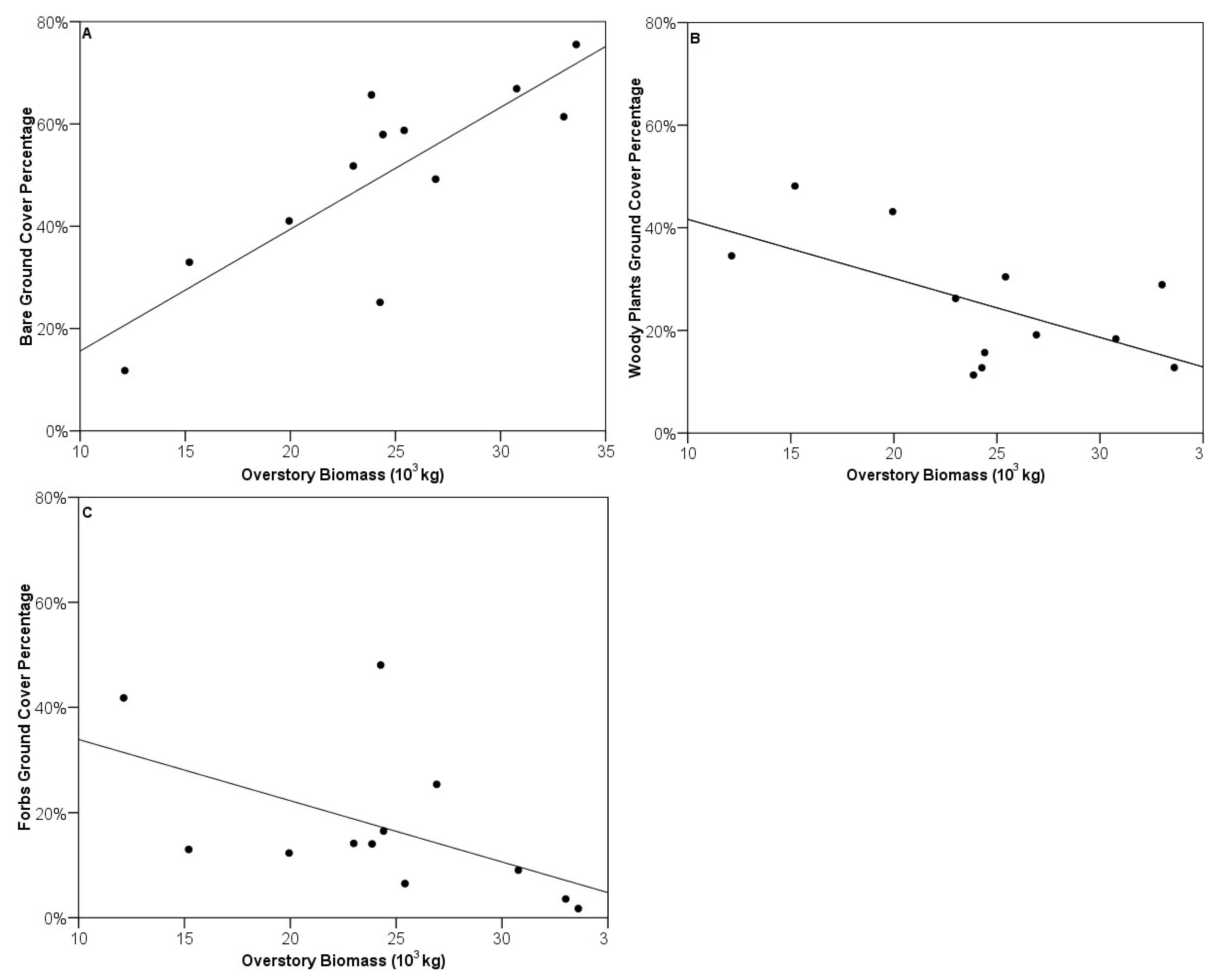
3.3. Vegetative Composition of Sites
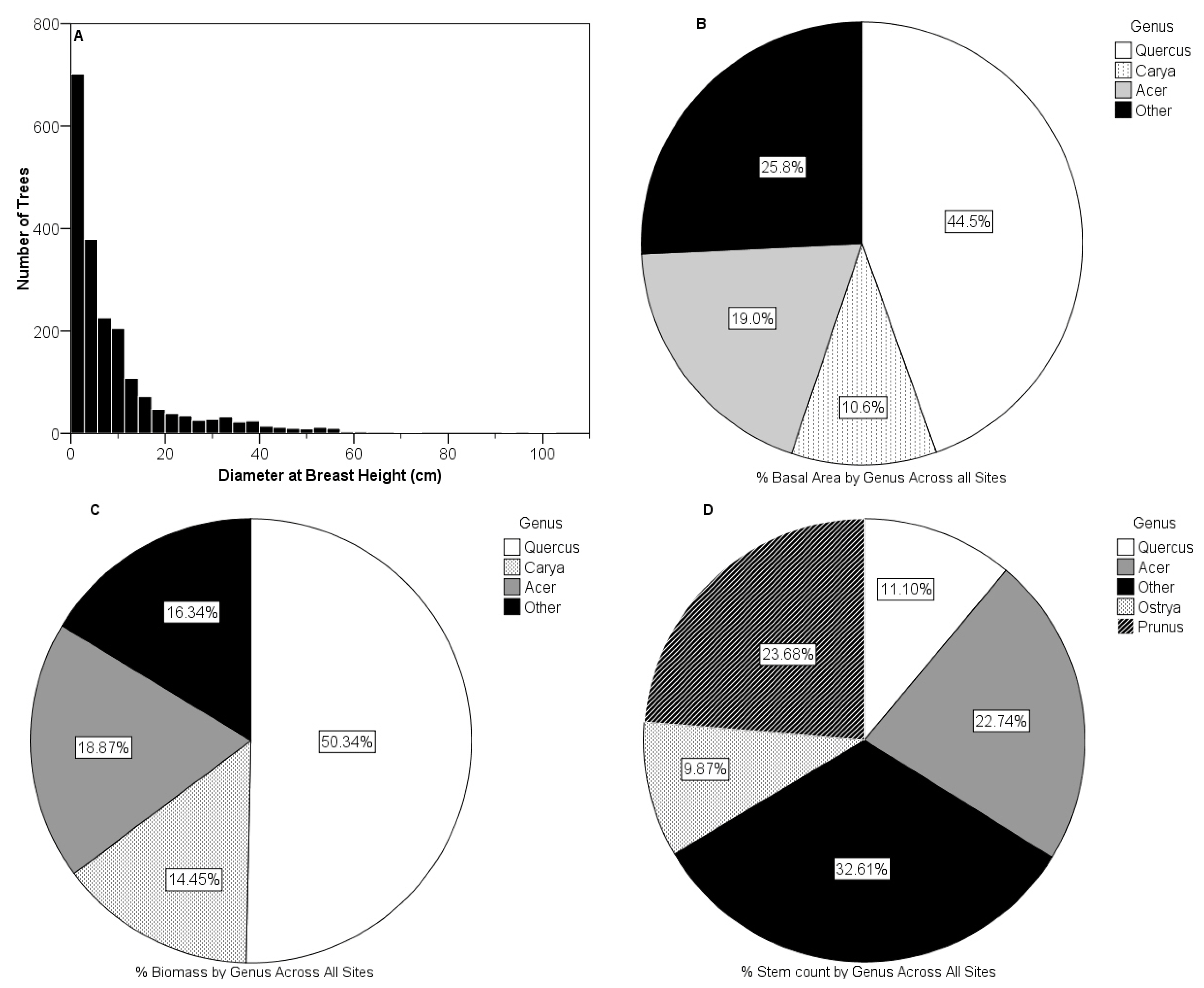

3.4. Relationship between Foliar N and Surrounding Housing Density
3.5. Soil N Content and Housing Density in the Surrounding Landscape
| Density | Site | Loge (LA/HU) | Soil % C | Soil % N | Soil C/N | Trees (#) | Biomass (kg) | Basal Area (103 cm2) | Foliar % C | Foliar % N | Foliar C/N |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Urban | SFW | −0.9 | 3.20% | 0.24% | 13.3 | 134 | 33,602 | 161,315 | 45.90% | 2.10% | 21.7 |
| Urban | TW | −1.21 | 3.28% | 0.24% | 13.8 | 167 | 30,769 | 185,351 | 44.40% | 2.50% | 17.5 |
| Urban | VW | −0.54 | 3.22% | 0.27% | 11.9 | 166 | 26,913 | 130,756 | 42.70% | 2.20% | 19.6 |
| Sub. | BPW | −0.53 | 3.35% | 0.25% | 13.6 | 79 | 12,129 | 81,655 | 43.30% | 2.10% | 20.3 |
| Sub. | SAG | 0.87 | 2.31% | 0.18% | 13.1 | 162 | 25,415 | 140,988 | 44.10% | 2.20% | 20.4 |
| Sub. | WBW | −0.1 | 2.25% | 0.16% | 14.3 | 195 | 33,008 | 160,385 | 44.80% | 2.00% | 22.9 |
| Ex. | OSM | 1.43 | 2.62% | 0.19% | 14.2 | 230 | 24,406 | 133,792 | 43.90% | 2.20% | 19.7 |
| Ex. | PP | 1.68 | 2.89% | 0.24% | 12.1 | 286 | 19,943 | 92,536 | 42.50% | 2.00% | 21.6 |
| Ex. | SW | 2.4 | 3.07% | 0.19% | 16 | 153 | 22,996 | 114,536 | 43.80% | 1.90% | 22.7 |
| Rural | ESG | 2.64 | 1.70% | 0.13% | 12.8 | 104 | 24,269 | 115,243 | 43.30% | 2.30% | 18.9 |
| Rural | LW | 2.8 | 3.19% | 0.27% | 11.7 | 157 | 15,200 | 102,401 | 44.20% | 2.30% | 19.6 |
| Rural | NWN | 3.67 | 1.41% | 0.10% | 14.4 | 194 | 23,861 | 145,049 | 43.90% | 2.00% | 22.2 |
| MEAN | -- | 2.71% | 0.20% | 13.4 | 169 | 24,376 | 130,334 | 43.90% | 2.14% | 20.6 |
| Genus | # of Samples | Mean % N | Mean C/N Ratio |
|---|---|---|---|
| Acer | 42 | 1.94 | 23.85 |
| Aesculus | 1 | 2.28 | 19.90 |
| Carpinus | 7 | 1.88 | 23.64 |
| Carya | 22 | 2.02 | 20.92 |
| Cornus | 5 | 2.01 | 20.93 |
| Fagus | 7 | 2.45 | 19.00 |
| Fraxinus | 22 | 2.22 | 19.74 |
| Hammamelis | 5 | 1.74 | 26.17 |
| Malus | 6 | 1.53 | 28.71 |
| Ostrya | 15 | 1.98 | 22.23 |
| Prunus | 39 | 2.26 | 19.92 |
| Quercus | 24 | 2.16 | 20.67 |
| Rhamnus | 7 | 3.31 | 12.16 |
| Robinea | 1 | 2.95 | 13.60 |
| Sassafras | 2 | 2.62 | 16.46 |
| Tilia | 11 | 2.59 | 16.45 |
| Ulmus | 24 | 2.15 | 19.92 |
| Viburnum | 1 | 2.36 | 18.14 |
3.6. The Effects of Time and Agricultural Abandonment
4. Conclusions
Acknowledgments
Author Contributions
Conflict of Interest
References
- Blair, R.B.; Launer, A.E. Butterfly diversity and human land use: Species assemblages along an urban gradient. Biol. Cons. 1997, 80, 113–125. [Google Scholar] [CrossRef]
- Alberti, M. The effects of urban patterns on ecosystem function. Int. Reg. Sci. Rev. 2005, 28, 168. [Google Scholar] [CrossRef]
- Duguay, S.; Eigenbrod, F.L.; Fahrig, L. Effects of surrounding urbanization on non-native flora in small forest patches. Landsc. Ecol. 2007, 22, 589–599. [Google Scholar] [CrossRef]
- Donnelly, R.; Marzluff, J.M. Importance of reserve size and landscape context to urban bird conservation. Conserv. Biol. 2004, 18, 733. [Google Scholar] [CrossRef]
- Piessens, K.; Honnay, O.; Hermy, M. The role of fragment area and isolation in the conservation of heathland species. Biol. Conserv. 2005, 122, 61–69. [Google Scholar] [CrossRef]
- Burger, J.; Gochfeld, M. Effects of ecotourists on bird behaviour at Loxahatchee National Wildlife Refuge, Florida. Environ. Conserv. 1998, 25, 13–21. [Google Scholar] [CrossRef]
- Matlack, G.R. Sociological edge effects: Spatial distribution of human impact in suburban forest fragments. Environ. Manag. 1993, 17, 829. [Google Scholar] [CrossRef]
- Pouyat, R.V.; Pataki, D.E.; Belt, K.T.; Horn, J.; Band, L.E. Effects of Urban Land-Use Change on Biogeochemical Cycles. In Terr. Ecosyst. Chang. World; Canadell, J.G., Pataki, D.E., Pitelka, L.F., Eds.; Springer-Verlag: Berlin, Germany, 2007; pp. 45–88. [Google Scholar]
- McKinney, M.L. Urbanization, biodiversity, and conservation. Bioscience 2002, 52, 883–890. [Google Scholar] [CrossRef]
- Grimm, N.B.; Foster, D.; Groffman, P.; Grove, J.M.; Hopkinson, C.S.; Nadelhoffer, K.J.; Pataki, D.E.; Peters, D.P.C. The changing landscape: Ecosystem responses to urbanization and pollution across climatic and societal gradients. Front. Ecol. Environ. 2008, 6, 264–272. [Google Scholar] [CrossRef]
- Forman, R.T.T. Some general principles of landscape and regional ecology. Landsc. Ecol. 1995, 10, 133–142. [Google Scholar] [CrossRef]
- Chen, L.; Fu, B.; Zhao, W. Source-sink landscape theory and its ecological significance. Front. Biol. China 1995, 3, 131–136. [Google Scholar] [CrossRef]
- Turner, B.L.; Meyer, W.B. Environmental Change, the Human Factor. In Humans as Components of Ecosystems; McDonnell, M.J., Pickett, S.T.A., Eds.; Springer-Verlag: New York, NY, USA, 1993; pp. 40–49. [Google Scholar]
- Thompson, F.R., III; DeGraaf, R.M. Conservation approaches for woody, early successional communities in the eastern United States. Wildl. Soc. Bull. 2001, 29, 483–494. [Google Scholar]
- Lepers, E.; Lambin, E.F.; Janetos, A.C.; DeFries, R.; Achard, F. A synthesis of information on rapid land-cover change for the period 1981–2000. BioScience 2005, 55, 115–124. [Google Scholar] [CrossRef]
- Foley, J.A.; DeFries, R.; Asner, G.P.; Barford, C.; Bonan, G.; Carpenter, S.R.; Chapin, F.S.; Coe, M.T.; Daily, G.C.; Gibbs, H.K.; Helkowski, J.H.; Holloway, T.; Howard, E.A.; Kucharik, C.J.; Monfreda, C.; Patz, J.A.; Prentice, I.C.; Ramankutty, N.; Snyder, P.K. Global consequences of land use. Science 2005, 309, 570. [Google Scholar] [CrossRef]
- Golubiewski, N.E. Urbanization increases grassland carbon pools: Effects of landscaping in Colorado’s front range. Ecol. Appl. 2006, 16, 555–571. [Google Scholar] [CrossRef]
- Botkin, D.B.; Beveridge, C.E. Cities as environments. Urban. Ecosyst. 1997, 1, 3–19. [Google Scholar] [CrossRef]
- Jenerette, G.D.; Wu, J.; Grimm, N.B.; Hope, D. Points, Patches, and regions, scaling soil biogeochemical patterns in an urbanized arid ecosystem. Glob. Chang. Biol. 2006, 12, 1532–1544. [Google Scholar] [CrossRef]
- Gregg, J.W.; Jones, C.G.; Dawson, T.E. Urbanization effects on tree growth in the vicinity of New York City. Nature 2003, 424, 183–187. [Google Scholar] [CrossRef]
- McDonnell, M.J.; Pickett, S.T.A.; Pouyat, R.V. The Application of the Ecological Gradient Paradigm to the Study of Urban Effects. In Humans as Components of Ecosystems; McDonnell, M.J., Pickett, S.T.A., Eds.; Springer-Verlag: New York, NY, USA, 1993; pp. 175–189. [Google Scholar]
- McKinney, M.L. Urbanization as a major cause of biotic homogenization. Biol. Conserv. 2006, 127, 247–260. [Google Scholar] [CrossRef]
- Groffman, P.M.; Law, N.L.; Belt, K.T.; Band, L.E.; Fisher, G.T. Nitrogen fluxes and retention in urban watershed ecosystems. Ecosystems 2004, 7, 393–403. [Google Scholar]
- Hansen, A.; Knight, R.; Marzluff, J.; Powell, S.; Brown, K.; Gude, P.; et al. Effects of exurban development on biodiversity: Patterns, mechanisms, and research needs. Ecol. Appl. 2005, 15, 1893. [Google Scholar] [CrossRef]
- Brown, D.G.; Johnson, K.M.; Loveland, T.R.; Theobald, D.M. Rural land use trends in the conterminous United States, 1950–2000. Ecol. Appl. 2005, 15, 1851–1863. [Google Scholar] [CrossRef]
- Theobald, D.M. Landscape patterns of exurban growth in the USA from 1980 to 2020. Ecol. Soc. 2005, 10, 32. [Google Scholar]
- Forys, E.; Allen, C. The impacts of sprawl on biodiversity: The ant fauna of the Lower Florida Keys. Ecol. Soc. 2005, 10, 1–15. [Google Scholar]
- Russell, E.W.B. Discovery of the Subtle. In Hum. Compon. Ecosyst; McDonnell, M.J., Pickett, S.T.A., Eds.; Springer-Verlag: New York, NY, USA, 1993; pp. 81–90. [Google Scholar]
- Vitousek, P.M.; Mooney, H.A.; Lubchenco, J.; Melillo, J.M. Human domination of earth’s ecosystems. Science 1997, 277, 494–499. [Google Scholar] [CrossRef]
- Vitousek, P.M.; Aber, J.D.; Howarth, R.W.; Likens, G.E.; Matson, P.A.; Schindler, D.W.; Schlesinger, W.H.; Tilman, D.G. Human alteration of the global nitrogen cycle: Sources and consequences. Ecol. Appl. 1997, 7, 737–750. [Google Scholar]
- Gruber, N.; Galloway, J.N. An Earth-system perspective of the global nitrogen cycle. Nature 2008, 451, 293–296. [Google Scholar] [CrossRef]
- Cape, N.; Tang, Y.S.; van Dijk, N.; Love, L.; Sutton, M.A.; Palmer, S.C.F. Concentrations of ammonia and nitrogen dioxide at roadside verges, and their contribution to nitrogen deposition. Environ. Pollut. 2004, 132, 469–478. [Google Scholar] [CrossRef]
- Hawbaker, T.J.; Radeloff, V.C.; Hammer, R.B.; Clayton, M.B. Road density and landscape pattern in relation to housing density, and ownership, land cover, and soils. Landsc. Ecol. 2005, 20, 609–625. [Google Scholar] [CrossRef]
- Aber, J.D.; Nadelhoffer, K.J.; Steudler, P.; Melillo, J.M. Nitrogen saturation in northern forest ecosystems. BioScience 1989, 39, 378–386. [Google Scholar] [CrossRef]
- Aber, J.D.; Magill, A.; Mcnulty, S.G.; Boone, R.D.; Nadelhoffer, K.J.; Downs, M.; Hallet, R. Forest biogeochemistry and primary production altered by nitrogen saturation. Water Air Soil Pollut. 1995, 85, 1665–1670. [Google Scholar] [CrossRef]
- Schlesinger, W.H. Biogeochemistry, 2nd ed.; Academic Press: London, UK, 1997. [Google Scholar]
- Freedman, B.; Love, S.; O’Neil, B. Tree species composition, structure, and carbon storage in stands of urban forest of varying character in Halifax, Nova Scotia. Can. Field-Nat. 1996, 110, 675. [Google Scholar]
- Pouyat, R.V. Litter decomposition and nitrogen mineralization in oak stands along an urban-rural land use gradient. Urban Ecosyst. 1997, 1, 117–131. [Google Scholar] [CrossRef]
- Baker, L.A.; Xu, Y.; Hope, D.; Lauver, L.; Edmonds, J. Nitrogen mass balance for the Phoenix-Cap ecosystem. Ecosystems 2001, 4, 582. [Google Scholar] [CrossRef]
- Groffman, P.M.; Pouyat, R.V.; Cadenasso, M.L.; Zipperer, W.C.; Szlavecz, K.; Yesilonis, I.D.; Band, L.E.; Brush, G.S. Land use context and natural soil controls on plant community composition and soil nitrogen and carbon dynamics in urban and rural forests. For. Ecol. Manag. 2006, 236, 177–192. [Google Scholar] [CrossRef]
- Fissore, C.; Hobbie, S.E.; King, J.Y.; McFadden, J.P.; Nelson, K.C.; Baker, L.A. The residential landscape: Fluxes of elements and the role of household decisions. Urban Ecosyst. 2012, 15, 1–18. [Google Scholar]
- Raciti, S.M.; Groffman, P.M.; Jenkins, J.C.; Pouyat, R.V.; Fahey, T.J.; Pickett, S.T.A.; Cadenasso, M.L. Accumulation of carbon and nitrogen in residential soils with different land-use histories. Ecosystems 2011, 14, 287–297. [Google Scholar] [CrossRef]
- Nowak, D.J.; Crane, D.E. Carbon storage and sequestration by urban trees in the USA. Environ. Pollut. 2002, 116, 381–389. [Google Scholar] [CrossRef]
- Galloway, J.N. Acid deposition: perspectives in time and space. Water AirSoil Pollut. 1995, 85, 15–24. [Google Scholar] [CrossRef]
- Liu, X.; Rogers, J. Indicator: Human Population Growth and Distribution in Southeast Michigan. 2007. Available online: http://www.epa.gov/med/grosseile_site/indicators/sos/hpg.pdf (accessed on 14 July 2008).
- Southeastern Michigan Council of Governments. Demographics Data Library. Available online: http://www.semcog.org/Demographic.aspx (accessed on 14 July 2008).
- McDonnell, M.J.; Pickett, S.T.A. Ecosystem structure and function along urban-rural gradients: an unexploited opportunity for ecology. Ecology 1990, 71, 1232–1237. [Google Scholar] [CrossRef]
- U.S. Census Bureau. Census 2000. Available online: http://www.census.gov/main/www/cen2000.html (accessed on 15 May 2008).
- Michigan Center for Geographic Information. Michigan Geographic Data Library. Available online: http://www.mcgi.state.mi.us/mgdl/ (accessed on 22 May 2008).
- Wilcox, G.; Macleod, D. CARL: Conservation and Recreation Lands Spatial Data Layer. Available online: http://glaro.ducks.org/carl/ (accessed on 22 May 2008).
- Taylor, J. Avian Populations in Human Dominated Landscapes: An Analysis of Spatio-Temporal Dynamics at the Urban Rural Interface. Ph.D. Thesis, University of Michigan School of Natural Resources and Environment, Ann Arbor, MI, USA, 2008. [Google Scholar]
- Bonham, C.D. Measurements for Terrestrial Vegetation; John Wiley and Sons: New York, NY, USA, 1989. [Google Scholar]
- Stohlgren, T.J.; Bull, K.A.; Otsuki, Y. Comparison of rangeland vegetation sampling techniques in the central grasslands. J. Range. Manag. 1998, 51, 164–172. [Google Scholar] [CrossRef]
- Balmford, A.; Green, M.J.B.; Murray, M.G. Using higher-taxon richness as a surrogate for species richness: I. Regional tests. Proc. Biol. Sci. 1996, 263, 1267–1274. [Google Scholar] [CrossRef]
- Balmford, A.; Jayasuriya, A.H.M.; Green, M.J.B. Using higher-taxon richness as a surrogate for species richness: II. Local applications. Proc. Biol. Sci. 1996, 263, 1571–1575. [Google Scholar] [CrossRef]
- Pregitzer, K.S.; Burton, A.J.; Zak, D.R.; Talhelm, A.F. Simulated chronic nitrogen deposition increases carbon storage in Northern Temperate forests. Glob. Chang. Biol. 2008, 14, 142–153. [Google Scholar]
- Gee, G.W.; Bauder, J.W. Particle size analysis. In Methods of Soil Analysis, Part 1, Physical and Mineralogical Methods, 2nd ed.; Klute, A., Ed.; American Society of Agronomy: Madison, WI, USA, 1986; pp. 383–411. [Google Scholar]
- Boerner, R.E.J.; Kost, J.A. Biomass equations for flowering dogwood, Cornus. florida L. Castanea 1986, 51, 153–155. [Google Scholar]
- Chifflot, V.; Bertoni, G.; Cabanettes, A.; Gavaland, A. Beneficial effects of intercropping on the growth and nitrogen status of young wild cherry and hybrid walnut trees. Agrofor. Syst. 2006, 66, 13–21. [Google Scholar] [CrossRef]
- Harrington, R.A.; Brown, B.J.; Reich, P.B. Ecophysiology of exotic and native shrubs in Southern Wisconsin. Oecologia. 1989, 80, 356–367. [Google Scholar] [CrossRef]
- Jenkins, M.A.; Jose, S.; White, P.S. Impacts of an exotic disease and vegetation change on foliar calcium cycling in Appalachian forests. Ecol. Appl. 2007, 17, 869–881. [Google Scholar] [CrossRef]
- Martin, J.G.; Kloeppel, B.D.; Schaefer, T.L.; Kimbler, D.L.; McNulty, S.G. Aboveground biomass and nitrogen allocation of ten deciduous southern Appalachian tree species. Can. J. For. Res. 1998, 28, 1648–1659. [Google Scholar]
- Perala, D.A.; Alban, D. Allometric Biomass Estimators for Aspen-Dominated Ecosystems in the Upper Great Lakes; Research Paper NC-314; USA. Department of Agriculture, Forest Service, North Central Forest Experiment Station: St. Paul, M.N, USA, 1993. [Google Scholar]
- Singh, B. Biomass production and nutrient dynamics in three clones of Populus. deltoides planted on Indogangetic plains. Plant Soil 1998, 203, 15–26. [Google Scholar] [CrossRef]
- Ter-Mikaelian, M.T.; Korzukhin, M.D. Biomass equations for sixty-five North American tree species. For. Ecol. Manag. 1997, 97, 1–24. [Google Scholar] [CrossRef]
- Williams, R.A.; McClenahen, J.R. Notes: Biomass prediction equations for seedlings, sprouts, and saplings of ten central hardwood species. For. Sci. 1984, 30, 523–527. [Google Scholar]
- Guerold, F. Influence of taxonomic determination level on several community indices. Water Res. 2000, 34, 487–492. [Google Scholar] [CrossRef]
- USDA Forest Service Forest Inventory and Analysis National Program. Available online: http://fia.fs.fed.us/ (accessed on 12 December 2008).
- Beven, K.; Kirby, M.J. A physically based, variable contributing area model of basin hydrology. Hydrol. Sci. Bull. 1979, 24, 43–69. [Google Scholar] [CrossRef]
- Kelejian, H.; Prucha, I.R. On the asymptotic distribution of the Moran I test statistic with applications. J. Econ. 2001, 104, 219–257. [Google Scholar] [CrossRef]
- Moran, P.A.P. Notes on continuous stochastic phenomena. Biometrika. 1950, 37, 17–23. [Google Scholar]
- STIS. Version 1.62. Terraseer: Ann Arbor, MI, USA, 2008. Available online: www.terraseer.com (accessed on 15 October 2008).
- SPSS. Version 15.0. SPSS: Chicago, IL, USA, 2007. Available online: www.spss.com (accessed on 1 June 2008).
- Fox, J. Linear Mixed Models. In Appendix: An R and S-PLUS Companion to Applied Regression; Sage Publication: Thousand Oaks, CA, USA, 2002. [Google Scholar]
- Steenbergen, M.R.; Jones, B.S. Modeling multilevel data structures. Am. J. Polit. Sci. 2002, 46, 218–237. [Google Scholar] [CrossRef]
- Vonesh, E.F.; Vernon, M.; Chinchilli, P.K. Goodness-of-fit in generalized nonlinear mixed-effects models. Biometrics 1996, 52, 572–587. [Google Scholar] [CrossRef]
- Hoaglin, D.; Welsch, R.E. The Hat Matrix in regression and ANOVA. Am. Stat. 1978, 32, 17–22. [Google Scholar]
- Szlavecz, K.; Placella, S.A.; Pouyat, R.V.; Groffman, P.M.; Csuzdi, C.; Yesilonis, I. Invasive earthworm species and nitrogen cycling in remnant forest patches. Appl. Soil. Ecol. 2006, 32, 54–62. [Google Scholar] [CrossRef]
- Pavao-Zuckerman, M.A.; Coleman, D.C. Decomposition of chestnut oak (Quercus prinus) leaves and nitrogen mineralization in an urban environment. Biol. Fertil. Soils 2005, 41, 343–349. [Google Scholar] [CrossRef]
- Hope, D.; Zhu, W.; Gries, C.; Oleson, J.; Kaye, J.; Grimm, N.; Baker, L.A. Spatial variation in soil inorganic nitrogen across an arid urban ecosystem. Urban. Ecosyst. 2005, 8, 251–273. [Google Scholar] [CrossRef]
- Driscoll, C.T.; Whitall, D.; Aber, J.; Boyer, E.; Castro, M.; Cronan, C.; Goodale, C.L.; Groffman, P.; Hopkinson, C.; Lambert, K.; Lawrence, G.; Ollinger, S. Nitrogen pollution in the northeastern United States: Sources, Effects, and Management Options. Bioscience 2003, 53, 367–374. [Google Scholar]
- Bohlen, P.J.; Groffman, P.M.; Driscoll, C.T.; Fahey, T.J.; Siccama, T.G. Plant-Soil-Microbial Interactions in a Northern Hardwood Forest. Ecology 2001, 82, 965–978. [Google Scholar]
- Finzi, A.C.; Van Breemen, N.; Canham, C.D. Canopy Tree-Soil Interactions within temperate forests: Species Effects on Soil Carbon and Nitrogen. Ecol. Appl. 1998, 8, 440–446. [Google Scholar]
- McFarland, J.; Ruess, R.; Kielland, K.; Pregitzer, K.; Hendrick, R. Glycine mineralization in situ closely correlates with soil carbon availability across six North American forest ecosystems. Biogeochemistry 2010, 99, 175–191. [Google Scholar] [CrossRef]
- Ollinger, S.V.; Smith, M.L.; Martin, M.E.; Hallett, R.A.; Goodale, C.L.; Aber, J.D. Regional variation in foliar chemistry and N cycling among forests of diverse history and composition. Ecology 2002, 83, 339–355. [Google Scholar]
- McNeil, B.E.; Read, J.M.; Driscoll, C.T. Foliar nitrogen responses to elevated atmospheric nitrogen deposition in nine temperate forest canopy species. Environ. Sci. Technol. 2007, 41, 5191–5197. [Google Scholar] [CrossRef]
- Holland, E.; Braswell, B.; Sulzman, J.; Lamarque, J. Nitrogen deposition onto the United States and western Europe: Synthesis of observations and models. Ecol. Appl. 2005, 15, 38–57. [Google Scholar] [CrossRef]
- Brown, D.G. Land use and forest cover on private parcels in the Upper Midwest USA, 1970 to 1990. Landsc. Ecol. 2003, 18, 777–790. [Google Scholar] [CrossRef]
- Compton, J.E.; Boone, R.D.; Motzkin, G.; Foster, D.R. Soil carbon and nitrogen in a pine-oak sand plain in central Massachusetts: Role of vegetation and land-use history. Oecologia 1998, 116, 536–542. [Google Scholar] [CrossRef]
- Kalisz, P.J. Soil properties of steep appalachian old fields. Ecology 1986, 67, 1011–1023. [Google Scholar] [CrossRef]
- Currie, W.S. The responsive C and N biogeochemistry of the temperate forest floor. Trends. Ecol. Evolut. 1999, 14, 316–320. [Google Scholar] [CrossRef]
- Knops, J.M.H.; Tilman, D. Dynamics of soil nitrogen and carbon accumulation for 61 years after agricultural abandonment. Ecology 2000, 81, 88–98. [Google Scholar] [CrossRef]
- Burgi, M.; Turner, M.G. Factors and processes shaping land cover and land cover changes along the Wisconsin River. Ecosystems 2002, 5, 184–201. [Google Scholar] [CrossRef]
- Foster, D.R. Land-Use History (1730–1990) and vegetation dynamics in central New England, USA. J. Ecol. 1992, 80, 753–772. [Google Scholar] [CrossRef]
- Le Toan, T.; Beaudoin, A.; Riom, J.; Guyon, D. Relating forest biomass to SAR data. Trans. Geosci. Remote. Sens. 1992, 30, 403–411. [Google Scholar] [CrossRef]
- Zak, D.R.; Pregitzer, K.S. Spatial and temporal variability of nitrogen cycling in northern Lower Michigan. For. Sci. 1990, 36, 367–380. [Google Scholar]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Kahan, A.Y.; Currie, W.S.; Brown, D.G. Nitrogen and Carbon Biogeochemistry in Forest Sites along an Indirect Urban–Rural Gradient in Southeastern Michigan. Forests 2014, 5, 643-665. https://doi.org/10.3390/f5040643
Kahan AY, Currie WS, Brown DG. Nitrogen and Carbon Biogeochemistry in Forest Sites along an Indirect Urban–Rural Gradient in Southeastern Michigan. Forests. 2014; 5(4):643-665. https://doi.org/10.3390/f5040643
Chicago/Turabian StyleKahan, Ari Y., William S. Currie, and Daniel G. Brown. 2014. "Nitrogen and Carbon Biogeochemistry in Forest Sites along an Indirect Urban–Rural Gradient in Southeastern Michigan" Forests 5, no. 4: 643-665. https://doi.org/10.3390/f5040643
APA StyleKahan, A. Y., Currie, W. S., & Brown, D. G. (2014). Nitrogen and Carbon Biogeochemistry in Forest Sites along an Indirect Urban–Rural Gradient in Southeastern Michigan. Forests, 5(4), 643-665. https://doi.org/10.3390/f5040643