Do Fire Regime Attributes Affect Soil Biochemical Properties in the Same Way under Different Environmental Conditions?




Abstract
:1. Introduction
2. Materials and Methods
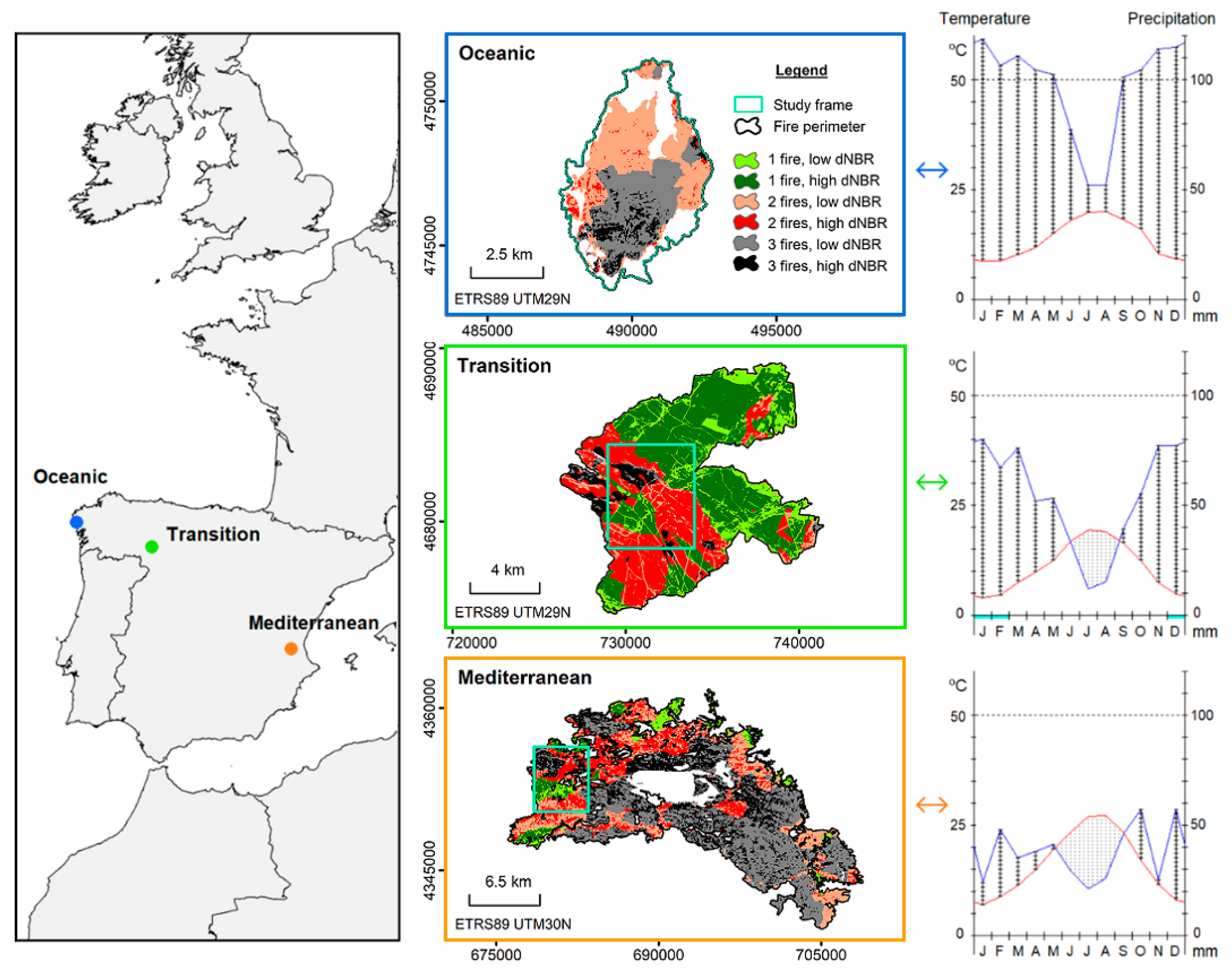
2.1. Study Sites
2.2. Fire Regime Attributes: Frequency and Severity
2.3. Field Sampling: Soil and Ecological Variables
2.4. Analyses of Soil Biochemical Properties: Enzymatic Activities and Microbial Biomass Carbon
2.5. Data Analyses
3. Results
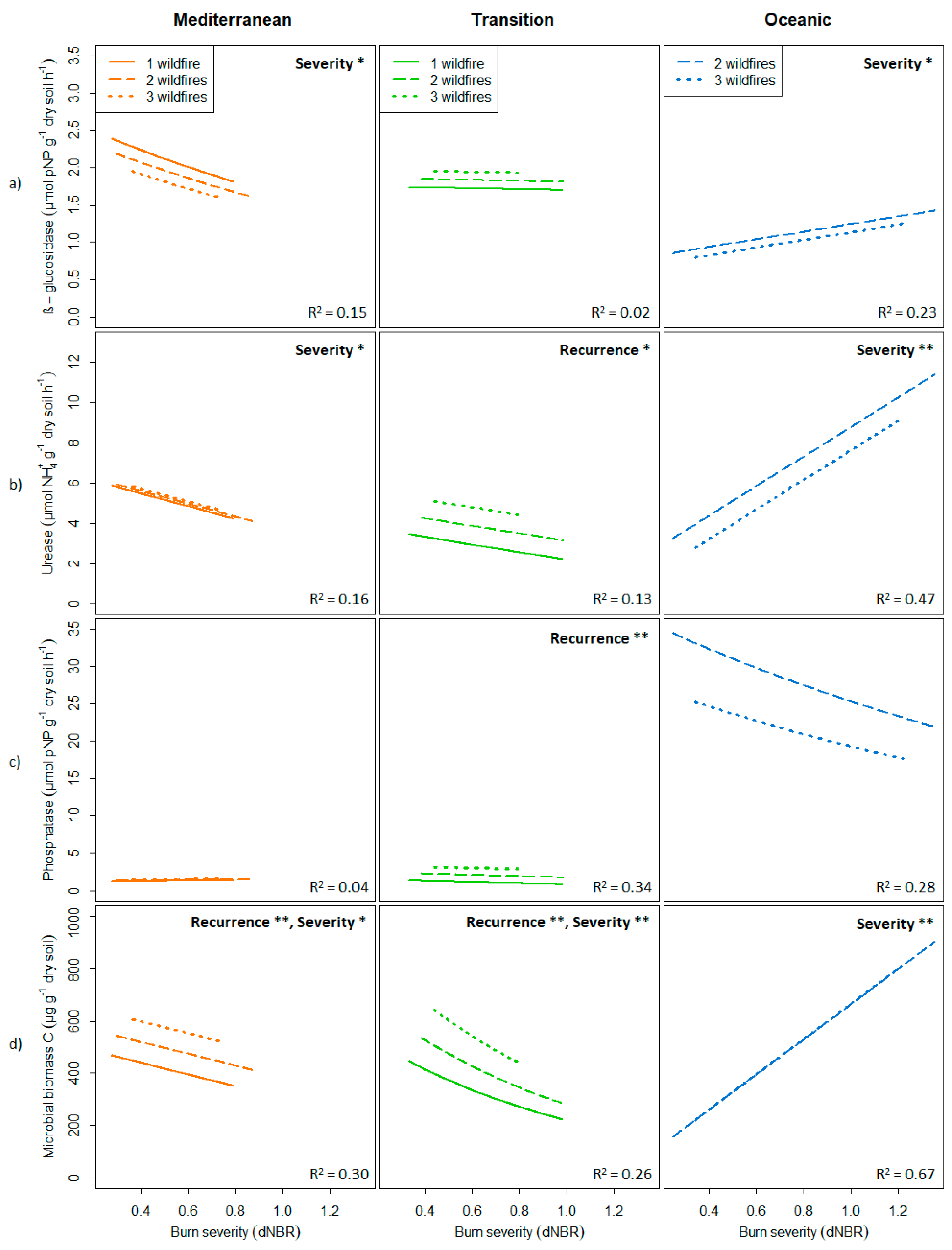
3.1. Relationships between Fire Regime Attributes and Soil Biochemical Properties
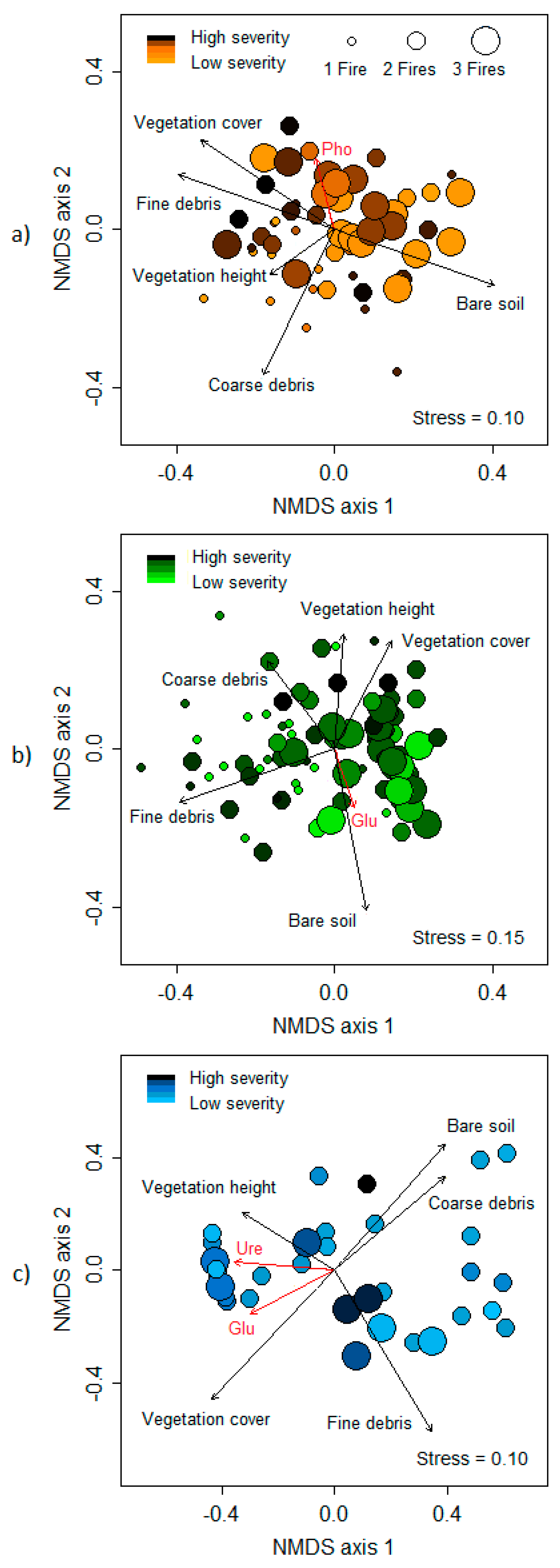
3.2. Impacts of Fire Regime Attributes on Soil Biochemical Properties Through Changes in Ecological Variables
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Moreno, J.M.; Oechel, W.C. The Role of Fire in Mediterranean-Type Ecosystems; Springer: New York, NY, USA, 1994. [Google Scholar]
- Keeley, J.E.; Bond, W.J.; Bradstock, R.A.; Pausas, J.; Rundel, P.W. Fire in Mediterranean Ecosystems: Ecology, Evolution and Management; Cambridge University Press: Cambridge, UK, 2012. [Google Scholar]
- Noss, R.F.; Franklin, J.F.; Baker, W.L.; Schoennagel, P.M.; Moyle, P.B. Managing fire-prone forests in the western United States. Front. Ecol. Environ. 2006, 4, 481–487. [Google Scholar] [CrossRef] [Green Version]
- Mouillot, F.; Rambal, S.; Joffre, R. Simulating climate change impacts on fire frequency and vegetation dynamics in a Mediterranean-type ecosystem. Glob. Chang. Biol. 2002, 8, 423–437. [Google Scholar] [CrossRef]
- Vázquez, A.; Climent, J.M.; Casais, L.; Quintana, J.R. Current and future estimates for the fire frequency and the fire rotation period in the main woodland types of peninsular Spain: A case-study approach. For. Syst. 2015, 24, e031. [Google Scholar] [CrossRef] [Green Version]
- Parks, S.A.; Miller, C.; Abatzoglou, J.T.; Holsinger, L.M.; Parisien, M.A.; Dobrowski, S.Z. How will climate change affect wildland fire severity in the western US? Environ. Res. Lett. 2016, 11, 035002. [Google Scholar] [CrossRef] [Green Version]
- Pausas, J.G.; Llovet, J.; Rodrigo, A.; Vallejo, R. Are wildfires a disaster in the Mediterranean basin?—A review. Int. J. Wildland Fire 2008, 17, 713–723. [Google Scholar] [CrossRef]
- Vilar, L.; Garrido, J.; Echavarría, P.; Martínez-Vega, J.; Martín, M.P. Comparative analysis of CORINE and climate change initiative land cover maps in Europe: Implications for wildfire occurrence estimation at regional and local scales. Int. Appl. Earth Obs. 2019, 78, 102–117. [Google Scholar] [CrossRef]
- Giorgi, F.; Lionello, P. Climate change projections for the Mediterranean region. Glob. Planet. Chang. 2008, 63, 90–104. [Google Scholar] [CrossRef]
- Hinojosa, M.B.; Parra, A.; Laudicina, V.A.; Moreno, J.M. Post-fire soil functionality and microbial community structure in a Mediterranean shrubland subjected to experimental drought. Sci. Total Environ. 2016, 573, 1178–1189. [Google Scholar] [CrossRef]
- Catalanotti, A.E.; Giuditta, E.; Marzaioli, R.; Ascoli, D.; Esposito, A.; Strumia, S.; Mazzoleni, S.; Rutigliano, F.A. Effects of single and repeated prescribed burns on soil organic C and microbial activity in a Pinus halepensis plantation of Southern Italy. Appl. Soil Ecol. 2017, 125, 108–116. [Google Scholar] [CrossRef]
- Van Wagtendonk, J.W.; Lutz, J.A. Fire Regime Attributes of Wildland Fires in Yosemite National Park, USA. Fire Ecol. 2007, 3, 34–52. [Google Scholar] [CrossRef]
- Bowman, D.M.J.S.; Balch, J.K.; Artaxo, P.; Bond, W.J.; Carlson, J.M.; Cochrane, M.A.; D’Antonio, C.M.; DeFries, R.S.; Doyle, J.C.; Harrison, S.P.; et al. Fire in the Earth system. Science 2009, 324, 481–484. [Google Scholar] [CrossRef]
- Rundel, P.W.; Arroyo, M.T.K.; Cowling, R.M.; Keeley, J.E.; Lamont, B.B.; Pausas, J.G.; Vargas, P. Fire and Plant Diversification in Mediterranean-Climate Regions. Front. Plant Sci. 2018, 9, 851. [Google Scholar] [CrossRef] [PubMed]
- Moya, D.; González-De Vega, S.; García-Orenes, F.; Morugán-Coronado, A.; Arcenegui, V.; Mataix-Solera, J.; Lucas-Borja, M.E.; de las Heras, J. Temporal characterization of soil-plant natural recovery related to fire severity in burned Pinus halepensis Mill. forests. Sci. Total Environ. 2018, 640–641, 42–51. [Google Scholar] [CrossRef] [PubMed]
- Fernández-García, V.; Marcos, E.; Fernández-Guisuraga, J.M.; Taboada, A.; Suárez-Seoane, S.; Calvo, L. Impact of burn severity on soil properties in a Pinus pinaster ecosystem immediately after fire. Int. J. Wildland Fire 2019, 28, 354–364. [Google Scholar] [CrossRef]
- Fernández-García, V.; Miesel, J.; Baeza, M.J.; Marcos, E.; Calvo, L. Wildfire effects on soil properties in fire-prone pine ecosystems: Indicators of burn severity legacy over the medium term after fire. Appl. Soil Ecol. 2019, 135, 147–156. [Google Scholar] [CrossRef] [Green Version]
- Fernández-García, V.; Fulé, P.Z.; Marcos, E.; Calvo, L. The role of fire frequency and severity on the regeneration of Mediterranean serotinous pines under different environmental conditions. For. Ecol. Manag. 2019, 444, 59–68. [Google Scholar] [CrossRef]
- Keesstra, S.; Wittenberg, L.; Maroulis, J.; Sambalino, F.; Malkinson, D.; Cerdà, A.; Pereira, P. The influence of fire history, plant species and post-fire management on soil water repellency in a Mediterranean catchment: The Mount Carmel range, Israel. Catena 2017, 149, 857–866. [Google Scholar] [CrossRef]
- Keeley, J.E. Fire intensity, fire severity and burn severity: A brief review and suggested usage. Int. J. Wildland Fire 2009, 18, 116–126. [Google Scholar] [CrossRef]
- Le Houerou, H.N. Fire and Vegetation in the Mediterranean Basin. In Proceedings of the 13th Tall Timbers Fire Ecology Conference, Tallahassee, FL, USA, 22–23 March 1973; pp. 237–277. [Google Scholar]
- Los incendios forestales en España. Decenio 2001–2010. Área de Defensa contra Incendios Forestales del Ministerio de Agricultura, Alimentación y Medio Ambiente. Available online: https://www.mapa.gob.es/es/desarrollo-rural/estadisticas/ (accessed on 20 December 2019).
- Áreas Ardidas por Tipo de Ocupação do Solo (1996–2014). Instituto da Conservação da Natureza e das Florestas. Available online: http://www2.icnf.pt/portal/florestas/dfci/inc/estat-sgif#tip (accessed on 20 December 2019).
- Certini, G. Effects of fire on properties of forest soils: A review. Oecologia 2005, 143, 1–10. [Google Scholar] [CrossRef]
- Caon, L.; Vallejo, V.R.; Ritsema, C.J.; Geissen, V. Effects of wildfire on soil nutrients in Mediterranean ecosystems. Earth-Sci. Rev. 2014, 139, 47–58. [Google Scholar] [CrossRef]
- Hedo, J.; Lucas-Borja, M.E.; Wic, C.; Andrés-Abellán, M.; de las Heras, J. Soil microbiological properties and enzymatic activities of long-term post-fire recovery in dry and semiarid Aleppo pine (Pinus halepensis M.) forest stands. Solid Earth 2015, 6, 243–252. [Google Scholar] [CrossRef] [Green Version]
- Muñoz-Rojas, M.; Erickson, T.E.; Martini, D.; Dixon, K.W.; Merritt, D. Soil physicochemical and microbiological indicators of short, medium and long term post-fire recovery in semi-arid ecosystems. Ecol. Indic. 2016, 63, 14–22. [Google Scholar] [CrossRef]
- Santín, C.; Doerr, S. Fire effects on soils: The human dimension. Philos. Trans. R. Soc. B 2016, 371, 20150171. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eugenio, M.; Verkaik, I.; Lloret, F.; Espelta, J.M. Recruitment and growth decline in Pinus halepensis populations after recurrent wildfires in Catalonia (NE Iberian Peninsula). For. Ecol. Manag. 2006, 231, 47–54. [Google Scholar]
- DeLuca, T.H.; Sala, A. Frequent fire alters nitrogen transformations in Ponderosa pine stands of the inland northwest. Ecology 2006, 87, 2511–2522. [Google Scholar] [CrossRef]
- Boerner, R.E.J.; Brinkman, J.A. Fire frequency and soil enzyme activity in southern Ohio oak-hickory forests. Appl. Soil Ecol. 2003, 23, 137–146. [Google Scholar] [CrossRef]
- Williams, R.J.; Hallgren, S.W.; Wilson, G.W.T. Frequency of prescribed burning in an upland oak forest determines soil and litter properties and alters the soil microbial community. For. Ecol. Manag. 2012, 265, 241–247. [Google Scholar] [CrossRef]
- Dooley, S.; Treseder, K.K. The effect of fire on microbial biomass: A meta-analysis of field studies. Biogeochemistry 2012, 109, 49–61. [Google Scholar] [CrossRef] [Green Version]
- Vega, J.A.; Fontúrbel, T.; Merino, A.; Fernández, C.; Ferreiro, A.; Jiménez, E. Testing the ability of visual indicators of soil burn severity to reflect changes in soil chemical and microbial properties in pine forests and shrubland. Plant Soil 2013, 369, 73–91. [Google Scholar] [CrossRef]
- Holden, S.R.; Rogers, B.M.; Treseder, K.K.; Randerson, J. Fire severity influences the response of soil microbes to a boreal forest fire. Environ. Res. Lett. 2016, 11, 035004. [Google Scholar] [CrossRef]
- Tabatabai, M.A. Soil enzymes. In Methods of Soil Analysis; Part 2: Microbial and Biochemical Properties; Weaver, R.W., Angle, J.S., Bottomley, P.J., Bezdicek, D.F., Smith, S., Tabatabai, M.A., Wollum, A.G., Eds.; ASA/SSSA: Madison, WI, USA, 1994; pp. 775–833. [Google Scholar]
- Pourreza, M.; Hosseini, S.M.; Sinegani, A.A.S.; Matinizadeh, M.; Dick, W.A. Soil microbial activity in response to fire severity in Zagros oak (Quercus brantii Lindl.) forests, Iran, after one year. Geoderma 2014, 213, 95–102. [Google Scholar] [CrossRef]
- BRENDA. The Comprehensive Enzyme Information System. Available online: https://www.brenda-enzymes.org/index.php (accessed on 20 December 2019).
- Bonde, T.A.; Schnürer, J.; Rosswall, T. Microbial biomass as a fraction of potentially mineralizable nitrogen in soils from long-term field experiments. Soil Biol. Biochem. 1988, 20, 447–452. [Google Scholar] [CrossRef]
- Sollins, P.; Homann, P.; Caldwell, B.A. Stabilization and destabilization of soil organic matter: Mechanisms and controls. Geoderma 1996, 74, 65–105. [Google Scholar] [CrossRef]
- Key, C.H.; Benson, N.C. Landscape Assessment (LA) Sampling and Analysis Methods; General Technical Report, RMRS-GTR-164-CD; U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station: Fort Collins, CO, USA, 2006.
- Mayor, A.G.; Goirán, S.B.; Vallejo, V.R.; Bautista, S. Variation in soil enzyme activity as a function of vegetation amount, type, and spatial structure in fire-prone Mediterranean shrublands. Sci. Total Environ. 2016, 573, 1209–1216. [Google Scholar] [CrossRef]
- Bai, X.; Zeng, Q.; Fakher, A.; Dong, Y.; An, S. Characteristics of soil enzyme activities and microbial biomass carbon and nitrogen under different vegetation zones on the Loess Plateau, China. Arid Land Res. Manag. 2018, 32, 438–454. [Google Scholar] [CrossRef]
- Atlas Climático Digital de la Península Ibérica. Available online: http://www.opengis.uab.es/wms/iberia/mms/index.htm (accessed on 20 December 2019).
- Jones, A.; Montanarella, L.; Jones, R. Soil atlas of Europe; European Commission: Brussels, Belgium, 2005. [Google Scholar]
- Worldwide Bioclimatic Classification System. Phytosociological Research Center, Spain. Available online: http://www.globalbioclimatics.org (accessed on 20 December 2019).
- Fernández-García, V.; Marcos, E.; Fulé, P.Z.; Reyes, O.; Santana, V.; Calvo, L. Fire regimes shape diversity and traits of vegetation under different climatic conditions. Sci. Total Environ. 2020, 716, 137137. [Google Scholar] [CrossRef]
- Fernández-García, V.; Quintano, C.; Taboada, A.; Marcos, E.; Calvo, L.; Fernández-Manso, A. Remote sensing applied to the study of fire regime attributes and their influence on post-fire greenness recovery in pine ecosystems. Remote Sens. 2018, 10, 733. [Google Scholar] [CrossRef] [Green Version]
- Fernández-García, V.; Santamarta, M.; Fernández-Manso, A.; Quintano, C.; Marcos, E.; Calvo, L. Burn severity metrics in fire-prone pine ecosystems along a climatic gradient using Landsat imagery. Remote Sens. Environ. 2018, 206, 205–217. [Google Scholar] [CrossRef]
- Peoples, M.S.; Koide, R.T. Considerations in the storage of soil samples for enzyme activity analysis. Appl. Soil Ecol. 2012, 62, 98–102. [Google Scholar] [CrossRef]
- Moreira, R.S.; Chiba, M.K.; Nunes, S.B.; De Maria, I.C. Air-drying pretreatment effect on soil enzymatic activity. Plant Soil Environ. 2017, 63, 98–102. [Google Scholar]
- Kandeler, E.; Gerber, H. Short-term assay of soil urease activity using colorimetric determination of ammonium. Biol. Fert. Soils 1988, 6, 68–72. [Google Scholar] [CrossRef]
- Vance, E.D.; Brookes, P.C.; Jenkinson, D.S. An extraction method for measuring soil microbial biomass C. Soil Biol. Biochem. 1987, 19, 703–707. [Google Scholar] [CrossRef]
- Joergensen, J.G. The fumigation-extraction method to estimate soil microbial biomass: Calibration of the kEC value. Soil Biol. Biochem. 1996, 28, 25–31. [Google Scholar] [CrossRef]
- Zuur, A.F.; Ieno, E.N.; Walker, N.; Saveliev, A.A.; Smith, G.M. Mixed Effects Models and Extensions in Ecology with R; Springer: New York, NY, USA, 2009. [Google Scholar]
- R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing. Available online: http://www.R-project.org/ (accessed on 20 December 2019).
- Vegan: Community Ecology Package. R Package Version 2. Available online: https://CRAN.R-project.org/package=vegan (accessed on 20 December 2019).
- Nlme: Linear and Nonlinear Mixed Effects Models. R Package Version 3. Available online: https://CRAN.R-project.org/package=nlme (accessed on 20 December 2019).
- Venables, W.N.; Ripley, B.D. Modern Applied Statistics with S, 4th ed.; Springer: New York, NY, USA, 2002. [Google Scholar]
- R2glmm: Computes R Squared for Mixed (Multilevel) Models. R Package Version 0.1.2. Available online: https://CRAN.R-project.org/package=r2glmm (accessed on 20 December 2019).
- Dilly, O.; Pompili, L.; Benedetti, A. Soil micro-biological indicators separated land use practices in contrast to abiotic soil properties at the 50 km scale under summer warm Mediterranean climate in northern Italy. Ecol. Indic. 2018, 84, 298–303. [Google Scholar] [CrossRef]
- Albiach, M.R.; Bonmatí, M.; Canet, R.; García, C.; García, A.; Gíl, F.; González, S.; Hernández, M.T.; Jiménez de Ridder, P.; Leirós, M.C.; et al. Sobre las enzimas del suelo y sus técnicas de medida. Edafología 2006, 13, 117–125. [Google Scholar]
- Lombao, A.; Barreiro, A.; Carballas, T.; Fontúrbel, M.T.; Martín, A.; Vega, J.A.; Fernández, C.; Díaz-Raviña, M. Changes in soil properties after a wildfire in Fragas do Eume Natural Park (Galicia, NW Spain). Catena 2015, 135, 409–418. [Google Scholar] [CrossRef]
- DeBano, L.F. The effect of fire on soil properties. In Proceedings-Management and Productivity of Western-Montane Forest Soils; Harvey, A.E., Neuenschwander, L.F., Compilers, Eds.; General Technical Report; INT-280; U.S. Department of Agriculture, Forest Service, Intermountain Research Station: Ogden, UT, USA, 1991; pp. 151–156. [Google Scholar]
- Parkin, T.B. Spatial variability of microbial processes in soil—A review. J. Environ. Q. 1993, 22, 409–417. [Google Scholar] [CrossRef]
- Trasar-Cepeda, C.; Leirós, M.C.; Gil-Sotres, F. Modification of biochemical properties by soil use. J. Soil Sci. Plant Nut. 2008, 8, 53–60. [Google Scholar] [CrossRef] [Green Version]
- Carballas, T.; Martín, A.; Díaz-Raviña, M. Efectos de los incendios forestales sobre los suelos de Galicia. In Efecto de Los Incendios Forestales Sobre Los Suelos de España: El Estado de la Cuestión Visto por Los Científicos Españoles; Cerdá, A., Mataix-Solera, J., Eds.; Cátedra de Divulgación de la Ciencia, Universitat de Valencia: Valencia, Spain, 2009; pp. 271–301. [Google Scholar]
- Miesel, J.R.; Boerner, R.E.; Skinner, C.N. Soil nitrogen mineralization and enzymatic activities in fire and fire surrogate treatments in California. Can. J. Soil Sci. 2011, 91, 935–946. [Google Scholar] [CrossRef]
- Cerón Rincón, L.; Melgarejo Múñoz, L. Enzimas del suelo: Indicadores de salud y calidad. Acta Biol. Colomb. 2005, 10, 5–18. [Google Scholar]
- González-Pérez, J.A.; González-Vila, F.J.; Almnedros, G.; Knicker, H. The effect of fire on soil organic matter—A review. Environ. Int. 2004, 30, 855–870. [Google Scholar] [CrossRef] [PubMed]
- Merino, A.; Fonturbel, M.T.; Fernández, C.; Chávez-Vergara, B.; García-Oliva, F.; Vega, J.A. Inferring changes in soil organic matter in post-wildfire soil burn severity levels in a temperate climate. Sci. Total Environ. 2018, 627, 622–632. [Google Scholar] [CrossRef] [PubMed]
- Kim, E.J.; Oh, J.E.; Chang, Y.S. Effects of forest fire on the level and distribution of PCDD/Fs and PAHs in soil. Sci. Total Environ. 2003, 311, 177–189. [Google Scholar] [CrossRef]
- López-Poma, R.; Bautista, S. Plant regeneration functional groups modulate the response to fire of soil enzyme activities in a Mediterranean shrubland. Soil Biol. Biochem. 2014, 79, 5–13. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Response Variable | Predictor | Mediterranean | Transition | Oceanic | Average | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| R2 | UC | LC | R2 | UC | LC | R2 | UC | LC | R2 ± SE | ||
| β-glucosidase | Model | 0.15 | 0.35 | 0.03 | 0.02 | 0.15 | 0.00 | 0.23 | 0.50 | 0.05 | 0.13 ± 0.06 |
| Frequency | 0.05 | 0.22 | 0.00 | 0.02 | 0.13 | 0.00 | 0.02 | 0.22 | 0.00 | 0.03 ± 0.01 | |
| Severity | 0.12 | 0.31 | 0.01 | 0.00 | 0.07 | 0.00 | 0.22 | 0.48 | 0.03 | 0.11 ± 0.06 | |
| Urease | Model | 0.16 | 0.36 | 0.04 | 0.13 | 0.30 | 0.04 | 0.47 | 0.68 | 0.26 | 0.25 ± 0.11 |
| Frequency | 0.01 | 0.11 | 0.00 | 0.12 | 0.27 | 0.02 | 0.04 | 0.26 | 0.00 | 0.06 ± 0.03 | |
| Severity | 0.14 | 0.34 | 0.02 | 0.04 | 0.15 | 0.00 | 0.46 | 0.67 | 0.23 | 0.21 ± 0.13 | |
| Phosphatase | Model | 0.04 | 0.21 | 0.00 | 0.34 | 0.50 | 0.20 | 0.28 | 0.54 | 0.08 | 0.22 ± 0.09 |
| Frequency | 0.03 | 0.18 | 0.00 | 0.34 | 0.49 | 0.19 | 0.09 | 0.34 | 0.00 | 0.15 ± 0.09 | |
| Severity | 0.01 | 0.12 | 0.00 | 0.03 | 0.13 | 0.00 | 0.11 | 0.36 | 0.00 | 0.05 ± 0.03 | |
| Microbial biomass C | Model | 0.30 | 0.50 | 0.14 | 0.26 | 0.42 | 0.12 | 0.67 | 0.81 | 0.51 | 0.41 ± 0.13 |
| Frequency | 0.20 | 0.40 | 0.05 | 0.19 | 0.35 | 0.07 | 0.00 | 0.15 | 0.00 | 0.13 ± 0.07 | |
| Severity | 0.10 | 0.28 | 0.04 | 0.14 | 0.30 | 0.04 | 0.63 | 0.78 | 0.45 | 0.29 ± 0.17 | |
| Response Variable | Predictor | Mediterranean | Transition | Oceanic | |||
|---|---|---|---|---|---|---|---|
| F | P | F | P | F | P | ||
| Ecological variables | Frequency | 4.37 | 0.02 | 5.28 | <0.01 | 0.53 | 0.59 |
| Severity | 1.86 | 0.16 | 3.20 | 0.04 | 0.14 | 0.86 | |
| Variable | Mediterranean | Transition | Oceanic | |||
|---|---|---|---|---|---|---|
| R2 | P | R2 | P | R2 | P | |
| Frequency | 0.17 | 0.02 | 0.25 | <0.01 | 0.12 | 0.13 |
| Severity | 0.04 | 0.42 | 0.08 | 0.05 | 0.02 | 0.71 |
| β-glucosidase | 0.02 | 0.68 | 0.11 | 0.02 | 0.20 | 0.02 |
| Urease | 0.04 | 0.41 | 0.03 | 0.29 | 0.23 | 0.02 |
| Phosphatase | 0.18 | 0.02 | 0.06 | 0.11 | 0.05 | 0.48 |
| Microbial biomass C | 0.10 | 0.08 | 0.06 | 0.10 | 0.03 | 0.63 |
| Bare soil | 0.93 | <0.01 | 0.82 | <0.01 | 0.63 | <0.01 |
| Fine plant debris | 0.88 | <0.01 | 0.81 | <0.01 | 0.79 | <0.01 |
| Coarse plant debris | 0.85 | <0.01 | 0.37 | <0.01 | 0.47 | <0.01 |
| Vegetation cover | 0.83 | <0.01 | 0.85 | <0.01 | 0.71 | <0.01 |
| Vegetation height | 0.20 | <0.01 | 0.41 | <0.01 | 0.26 | <0.01 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fernández-García, V.; Marcos, E.; Reyes, O.; Calvo, L. Do Fire Regime Attributes Affect Soil Biochemical Properties in the Same Way under Different Environmental Conditions? Forests 2020, 11, 274. https://doi.org/10.3390/f11030274
Fernández-García V, Marcos E, Reyes O, Calvo L. Do Fire Regime Attributes Affect Soil Biochemical Properties in the Same Way under Different Environmental Conditions? Forests. 2020; 11(3):274. https://doi.org/10.3390/f11030274
Chicago/Turabian StyleFernández-García, Víctor, Elena Marcos, Otilia Reyes, and Leonor Calvo. 2020. "Do Fire Regime Attributes Affect Soil Biochemical Properties in the Same Way under Different Environmental Conditions?" Forests 11, no. 3: 274. https://doi.org/10.3390/f11030274
APA StyleFernández-García, V., Marcos, E., Reyes, O., & Calvo, L. (2020). Do Fire Regime Attributes Affect Soil Biochemical Properties in the Same Way under Different Environmental Conditions? Forests, 11(3), 274. https://doi.org/10.3390/f11030274