3. Results
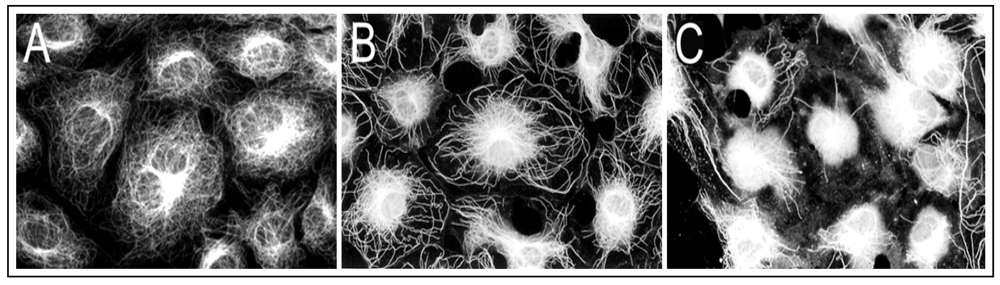
To elucidate As
3+ effects on cellular MT organization, growth arrested RFL6 cells on coverslips were incubated with or without freshly prepared As
3+ solution at final concentrations as indicated. Cellular MT distribution was revealed by immunocytochemistry [
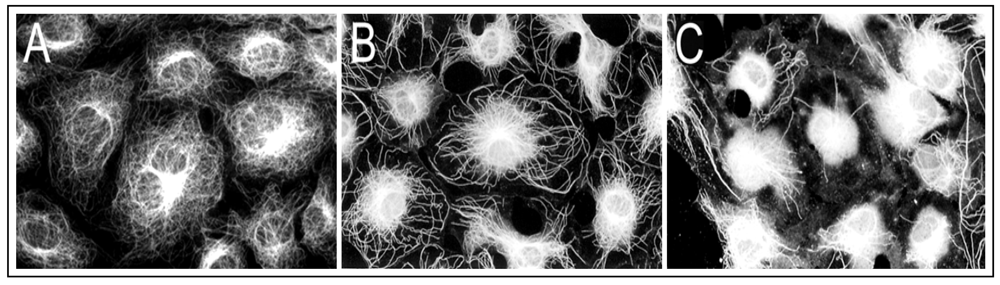
21]. As shown in
Figure 1(A), growth-arrested RFL6 cells contain an intricate, highly organized network of MTs that radiate outward from the MT organizing center (MTOC) in the perinuclear region to the cell periphery. Exposure of cells to 5 µM As
3+ for 24 h induced partial loss of MTs and no-organized tubulins with brightly staining accumulated in the perinuclear area where the MTOC is generally located [
Figure 1(B)]. Treatment of cells with 20 µM As
3+ for 24 h resulted in severe loss of MTs. Instead, mast cells exhibited high-density of depolymerized tubulin mass concentrated in the central area of cells [
Figure 1(C)]. Notably, the maximum doses of As
3+ (20 µM) that disrupted the MT organization in the cell model did not affect cell viability seriously (about 10% decrease) as assayed by trypan blue staining (data not shown). Morphological alterations in MTs were reversible after removal of As
3+. These studies indicated that the MT cytoskeleton is an important cellular target for As
3+ insult.
Figure 1.
As3+ induced disassembly of MTs in RFL6 cells. Growth-arrested RFL6 cells on coverslips were treated without or with As3+ at indicated concentrations for 24 h, and then immunofluorescently stained for visualization of MTs. (A) control; (B) 5 µM As3+; (C) 20 µM As3+.
Figure 1.
As3+ induced disassembly of MTs in RFL6 cells. Growth-arrested RFL6 cells on coverslips were treated without or with As3+ at indicated concentrations for 24 h, and then immunofluorescently stained for visualization of MTs. (A) control; (B) 5 µM As3+; (C) 20 µM As3+.
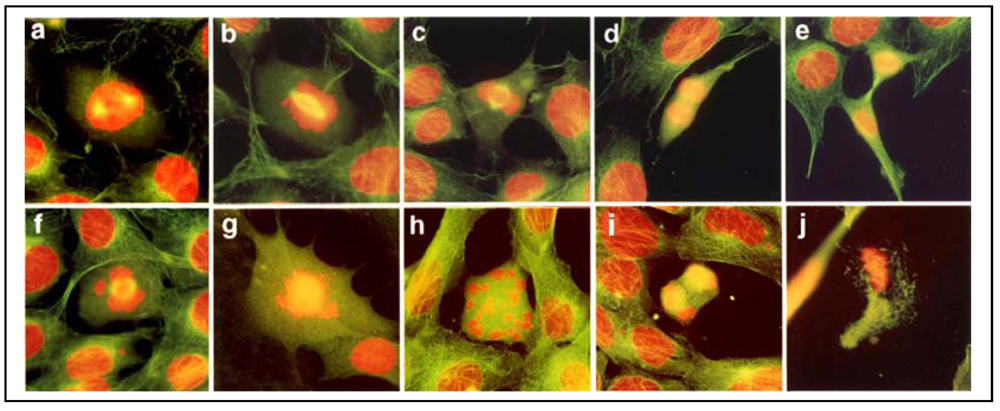
Since spindle MTs are intrinsically involved in chromosomal organization during cell mitosis [
13,
14], As
3+ may perturb spindle MTs inducing abnormal cell division. To test this, we have examined cell division aberrations by fluorescently double-staining spindle MTs and chromosomes
in situ in As
3+-treated RFL6 cells. As shown (
Figure 2), exposure of RFL6 cells to 10 µM As
3+ for 24 h resulted in the defect in or the absence of spindle MTs in 34% of proliferating cells (200 cells counted) accompanied by chromosomal lagging, scattering, clustering,
etc., in 22% of proliferating cells (200 cells counted) [
Figure 2(f–j)], in sharp contrast to those in control cells (only 2% of proliferating cells counted with spindle deficiency and 1.5% of proliferating cells counted with chromosomal disorientations).
Figure 2(a–e) show normal chromosome patterns in different phases of the cell division. These results indicate that chromosomal disorientations are closely linked to As
3+-induced damage to spindle MTs.
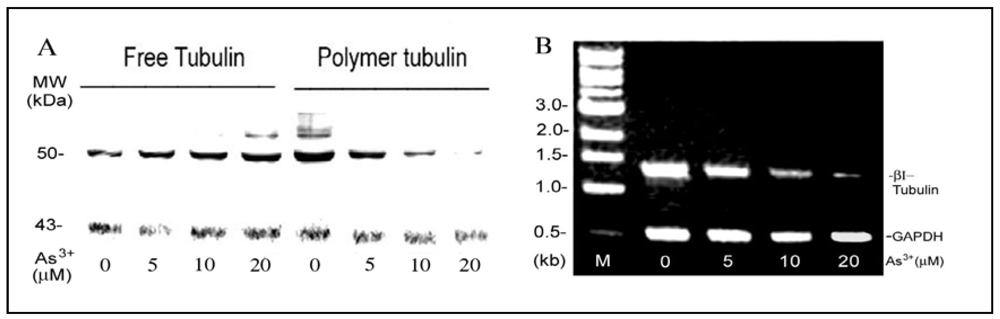
As
3+-induced MT disassembly (
Figure 1) should decrease the polymer MT and increase the free tubulin pool. To test this, As
3+-treated cells were treated with 0.3% NP40 in the PM2G buffer to isolate cytosolic and detergent-resistant cytoskeleton fractions [
21]. Western blot analysis indicated that in comparison to the actin control, RFL6 cells exposed to As
3+ exhibited a dose-dependent decrease in polymer MT proteins and increase in the free tubulin pool [
Figure 3(A)]. The density assays indicated that the 50% decrease in MT polymer proteins and increase in the free tubulin pool occurred in cells exposed to 5 µM As
3+. These results are consistent with morphological loss of MTs and increase of free tubulin mass in these cells as revealed by fluorescent microscopy. Five β-tubulin genes encode highly homologous proteins, class I–V [
26]. The rat class I β-tubulin (βI-tubulin) gene was selected as a sample to elucidate As
3+ modulation of tubulin transcription. As shown by the RT-PCR assay [
Figure 3(B)], the steady-state βI tubulin mRNA levels in cells treated with 5, 10 and 20 µM As
3+ were reduced to 66%, 26% and 6% of the control, respectively. Thus, As
3+ depolymerization of MTs was associated with inhibition of tubulin mRNAs.
Figure 2.
Deficient spindle MTs and cell division aberrations in As3+-treated cells. RFL6 cells on coverslips were incubated in 10% FBS medium without (a–e) or with (f–j) 10 µM As3+ for 24 h. Cell division stages and aberrations were examined by visualization of the in situ chromosomes stained with propidium iodine (red) and spindle MT stained with FITC-conjugated anti-tubulin antibody (green). Note: Cytoplasmic FITC-labeled MTs are green; however, since spindle MTs overlap with DNA staining in red, they appear as yellow structures. a, prophase; b, prometaphase; c, metaphase; d, anaphase; e, telophase; f, chromosome lagging with a defective spindle at metaphase; g, chromosome scattering in the absence of spindle; h, chromosome clustering and scattering in the absence of spindle; i, chromosome lagging with a defective spindle at anaphase; j, apoptosis of a cell with aberrant division.
Figure 2.
Deficient spindle MTs and cell division aberrations in As3+-treated cells. RFL6 cells on coverslips were incubated in 10% FBS medium without (a–e) or with (f–j) 10 µM As3+ for 24 h. Cell division stages and aberrations were examined by visualization of the in situ chromosomes stained with propidium iodine (red) and spindle MT stained with FITC-conjugated anti-tubulin antibody (green). Note: Cytoplasmic FITC-labeled MTs are green; however, since spindle MTs overlap with DNA staining in red, they appear as yellow structures. a, prophase; b, prometaphase; c, metaphase; d, anaphase; e, telophase; f, chromosome lagging with a defective spindle at metaphase; g, chromosome scattering in the absence of spindle; h, chromosome clustering and scattering in the absence of spindle; i, chromosome lagging with a defective spindle at anaphase; j, apoptosis of a cell with aberrant division.
Figure 3.
As3+ effects on expressions of tubulin proteins and mRNAs in the cell model. (A) As3+ enhanced the free tubulin pool as a result of depolymerization of MTs. Growth-arrested RFL6 cells were treated with As3+ at indicated concentrations for 24 h. Cytoskeletal and cytosolic fractions were isolated, and analyzed by Western blot. β-tubulin, 50 kDa; actin, 43 kDa used as a reference. (B) As3+ inhibition of steady-state βI tubulin mRNA levels. Cells were treated with As3+ as indicated above. Total RNA extracted was assessed for steady-state βI tubulin mRNA levels by the RT-PCR. GAPDH mRNA levels were used as an internal control. M indicates a DNA molecular ladder.
Figure 3.
As3+ effects on expressions of tubulin proteins and mRNAs in the cell model. (A) As3+ enhanced the free tubulin pool as a result of depolymerization of MTs. Growth-arrested RFL6 cells were treated with As3+ at indicated concentrations for 24 h. Cytoskeletal and cytosolic fractions were isolated, and analyzed by Western blot. β-tubulin, 50 kDa; actin, 43 kDa used as a reference. (B) As3+ inhibition of steady-state βI tubulin mRNA levels. Cells were treated with As3+ as indicated above. Total RNA extracted was assessed for steady-state βI tubulin mRNA levels by the RT-PCR. GAPDH mRNA levels were used as an internal control. M indicates a DNA molecular ladder.
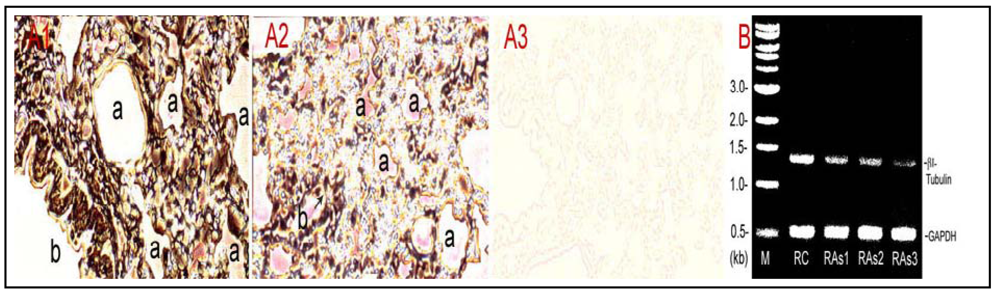
To elucidate As
3+ damage to the lung MTs, a small animal model study was performed to visualize tubulin protein distribution by immunohistochemistry and to assay for steady-state βI tubulin mRNA expression by the RT-PCR in control and As
3+-dosed lungs. As shown [
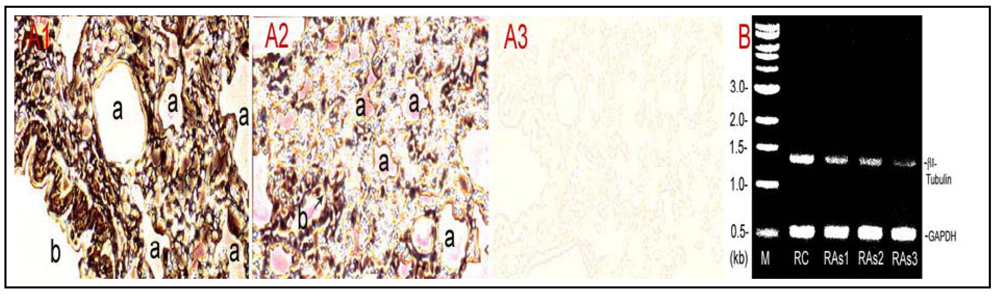
Figure 4(A)], immunoperoxidase staining of the lung tissue indicated markedly weakened tubulin signals,
i.e., peroxidase-labeled brown-black color materials, in As
3+-treated lung tissues [
Figure 4(A2)] in comparison to the control [
Figure 4(A1)].
Figure 4(A3) is a negative control tissue slice from control rats, which was incubated in the absence of the primary antibody against β-tubulin. Notably, As
3+ exposure apparently induced discontinuity of tubulin-rich epithelial cells in small bronchioles (arrow). Furthermore, RT-PCR assays indicated that As
3+-dosed lungs displayed decreased βI tubulin mRNA levels amounting to 53%, 48% and 20% of the control in As
3+-exposed Rat#1, Rat#2 and Rat#3, respectively [
Figure 4(B)]. Thus, As
3+ inhibited pulmonary tubulin expression at protein and mRNA levels
in vivo.
Figure 4.
Inhibition of β-tubulin expression at protein and mRNA levels in As3+-exposed lungs. Rats (≈150 g body weight) were intratracheally instilled with 520–530 µg NaAsO2 in 100 µL physiological saline according to 2.02 mg As/kg body weight once a week for 5 weeks. Control rats received saline only. Rats were killed a week after the last instillation. Lung tissues were processed for immunohistochemical staining and total RNA extraction. (A) β-Tubulin protein distribution. The tubulin protein in control and As3+-dosed lung sections was stained with the antibody against β-tubulin and the streptavidin-HRP system. (A1) control; (A2) As3+ exposed lung; (A3) a negative control without the primary antibody treatment. a, alveolus; b, bronchioles; arrow indicates discontinuity of tubulin-rich epithelial cells in small bronchioles. (B) βI-tubulin mRNA levels revealed by the RT-PCR. RC, control rat; RAs1, As3+ treated Rat#1; RAs2, As3+-treated Rat#2; RAs3, As3+-treated Rat#3 (note: single doses for Rat #1, #2 and #3 were 520, 525 and 530 µg NaAsO2 , respectively, dependent on the body weight). M, a DNA molecular ladder.
Figure 4.
Inhibition of β-tubulin expression at protein and mRNA levels in As3+-exposed lungs. Rats (≈150 g body weight) were intratracheally instilled with 520–530 µg NaAsO2 in 100 µL physiological saline according to 2.02 mg As/kg body weight once a week for 5 weeks. Control rats received saline only. Rats were killed a week after the last instillation. Lung tissues were processed for immunohistochemical staining and total RNA extraction. (A) β-Tubulin protein distribution. The tubulin protein in control and As3+-dosed lung sections was stained with the antibody against β-tubulin and the streptavidin-HRP system. (A1) control; (A2) As3+ exposed lung; (A3) a negative control without the primary antibody treatment. a, alveolus; b, bronchioles; arrow indicates discontinuity of tubulin-rich epithelial cells in small bronchioles. (B) βI-tubulin mRNA levels revealed by the RT-PCR. RC, control rat; RAs1, As3+ treated Rat#1; RAs2, As3+-treated Rat#2; RAs3, As3+-treated Rat#3 (note: single doses for Rat #1, #2 and #3 were 520, 525 and 530 µg NaAsO2 , respectively, dependent on the body weight). M, a DNA molecular ladder.
![Ijerph 09 00474 g004]()
To identify characteristics of the interaction of As
3+ with tubulin proteins, we assessed its effects on the MT polymerization
in vitro. Bovine brain MT proteins containing tubulins and MAPs were purified through two cycles of temperature dependent assembly and disassembly [
17]. MT assembly
in vitro was monitored by turbidity assays at A
350 nm (
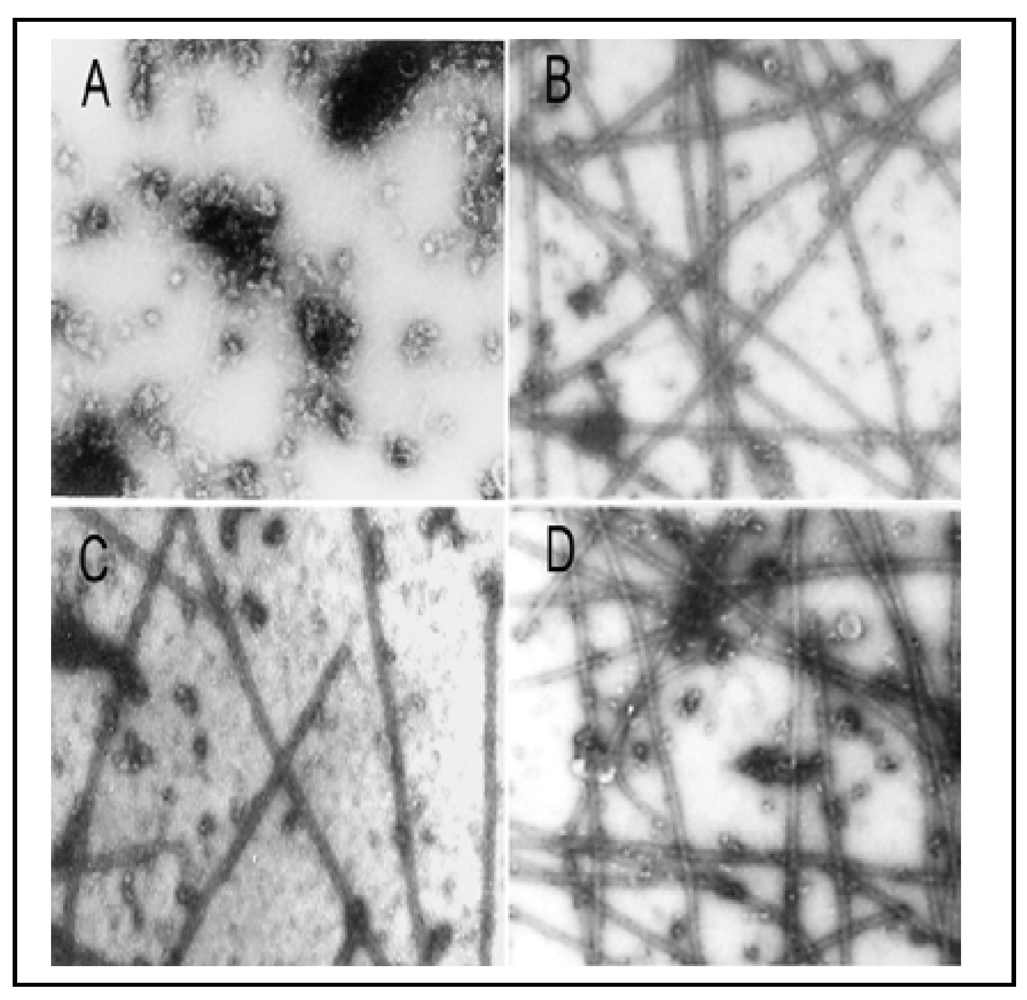
Figure 5) and confirmed by electron microscopy (EM) (
Figure 6) as described [
16,
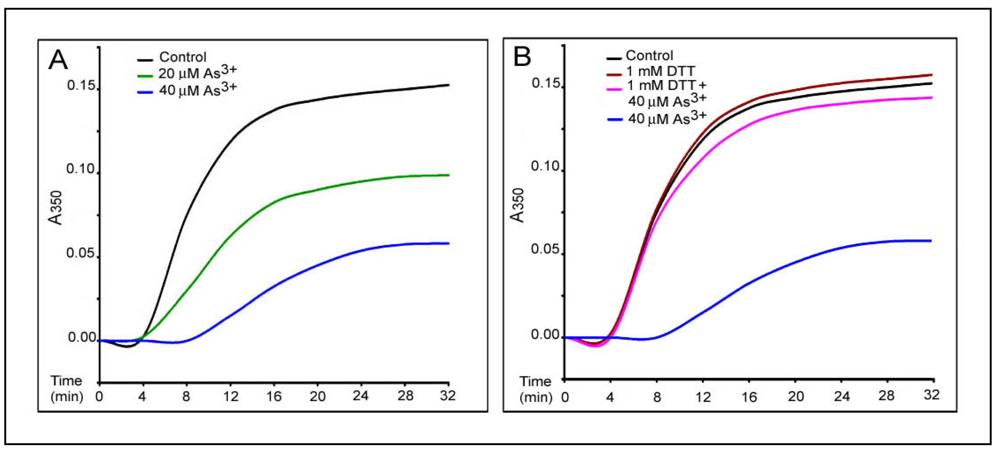
17]. In a typical turbidity assay at 25 °C [the black curve, in
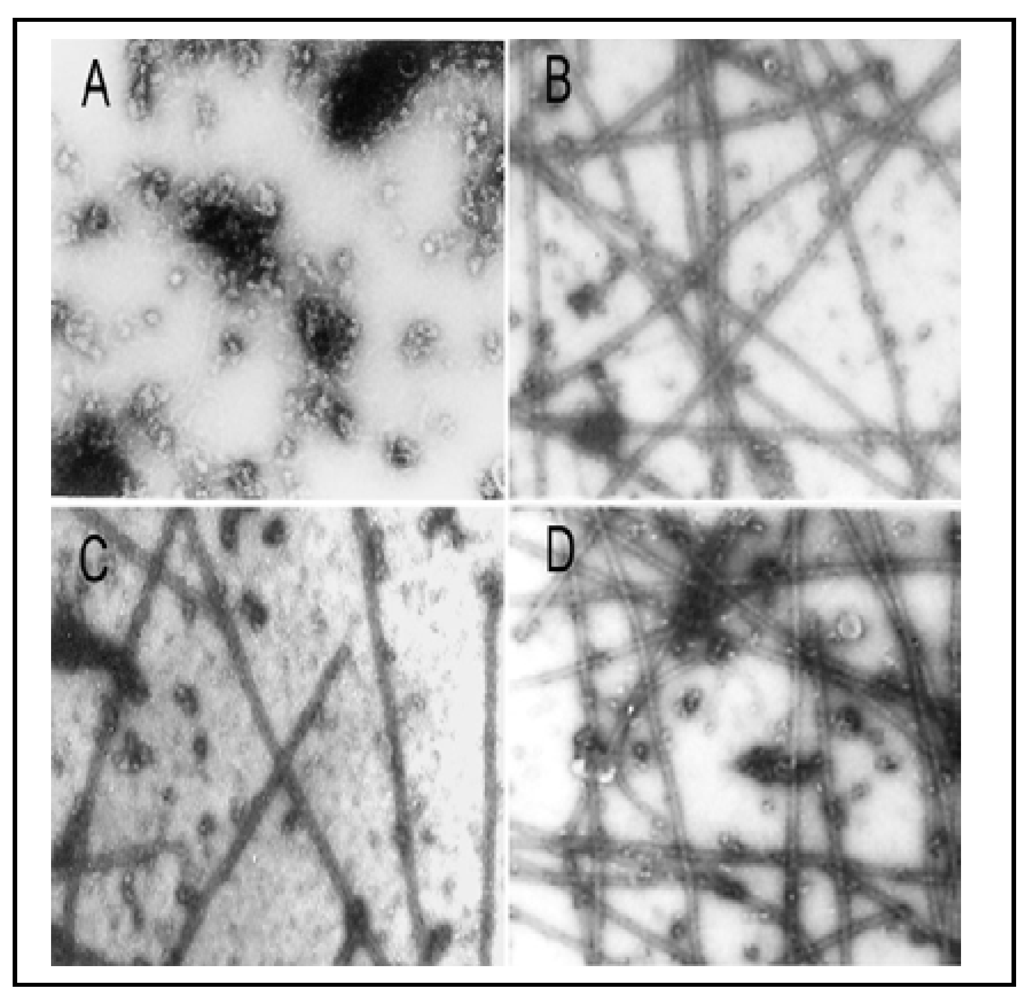
Figure 5(A)], assembly of MT proteins (0.8 mg/mL) progressed steadily with an initial leg (two min) followed by an elongation phase (2–30 min) and a final plateau indicating the reaction equilibrium (30–32 min). EM examination of samples taken at 30 min revealed the typical morphology of
in vitro assembled MTs [
Figure 6(B)]. No assembly of MTs was evident at time 0 [
Figure 6(A)]. Incubation with As
3+ induced a dose-dependent inhibition of MT assembly as evidenced by enhancement of the initial leg, decreases in the elongation rates and the plateau levels. For example, the average elongation rates at 12 min of the reaction (the turbidity values/time) were decreased to 50.0%, and 10.4% of the control at 20 (the green curve) and 40 (the blue curve) µM of As
3+, respectively [
Figure 5(A)]. The inhibition of MT assembly by As
3+ was further confirmed by EM which showed significant reduction in the number of MT fragments in the presence of 40 µM As
3+ [
Figure 6(C)], amounting to 50% of the control [
Figure 6(B)]. Importantly, addition of 1 mM dithiothreitol (DTT), a thiol reducing agent, to the reaction mixture strongly abolished the As
3+ inhibitory effect on MT assembly as shown in the turbidity [the pink curve in
Figure 5(B)] and EM assays [
Figure 6(D)] reaching 95% (the turbidity assay) and 108% (the EM assay) of corresponding controls, respectively. Thus, As
3+ inhibited MT polymerization
in vitro and results provide a critical clue for the direct interaction of As
3+ with tubulin -SH groups.
Figure 5.
As3+-inhibition of MT protein polymerization in vitro as revealed by the turbidity assay. The in vitro polymerization of MT proteins (0.8 mg/mL) in 0.1 M Pipes buffer, pH 6.6, containing 0.15 mM Mg2+, 0.15 mM EGTA and 500 µM GTP was monitored by turbidity readings at A350nm at 25 °C in the absence or presence of As3+ at indicated concentrations. Curves represent the time course of tubulin assembly under different conditions. (A) dose-dependent inhibition by As3+ of MT assembly. the black curve, control; the green curve, 20 µM As3+; the blue curve, 40 µM As3+. (B) DTT effects on As3+ inhibition of MT assembly. DTT (1 mM) was added after preincubation of polymerization mixture in the presence of As3+ for three min. the black curve, control; the brown curve, 1 mM DTT; the blue curve, 40 µM As3+; the pink curve, 40 µM As3+ plus 1 mM DTT.
Figure 5.
As3+-inhibition of MT protein polymerization in vitro as revealed by the turbidity assay. The in vitro polymerization of MT proteins (0.8 mg/mL) in 0.1 M Pipes buffer, pH 6.6, containing 0.15 mM Mg2+, 0.15 mM EGTA and 500 µM GTP was monitored by turbidity readings at A350nm at 25 °C in the absence or presence of As3+ at indicated concentrations. Curves represent the time course of tubulin assembly under different conditions. (A) dose-dependent inhibition by As3+ of MT assembly. the black curve, control; the green curve, 20 µM As3+; the blue curve, 40 µM As3+. (B) DTT effects on As3+ inhibition of MT assembly. DTT (1 mM) was added after preincubation of polymerization mixture in the presence of As3+ for three min. the black curve, control; the brown curve, 1 mM DTT; the blue curve, 40 µM As3+; the pink curve, 40 µM As3+ plus 1 mM DTT.
To further assess As
3+ binding to tubulin protein -SH groups, we also examined the competition of As
3+ with N-ethylmaleimide (NEM), a sulfhyryl group reagent [
21], for tubulin binding. “Pure” tubulin free of MAP (1.5 mg/mL) prepared from bovine brain was pretreated with As
3+ at indicated doses for 2 h at 0 °C, mixed with [
3H]NEM (2 µCi/mL), and incubated for an additional 2 h at 37 °C. Proteins were precipitated with 5% TCA and filtered. Radioactivities associated with collected proteins on the membrane were measured by β-counting. As shown (
Figure 7), As
3+ decreased the binding of [
3H]NEM to tubulin with 50% inhibition occurring at 23 µM. This straightforward experiment shows As
3+ binding to tubulin through -SH groups, initiating the cascade of MT disruption events.
Figure 6.
As
3+-inhibition of MT protein polymerization
in vitro as confirmed by EM examination. The
in vitro MT polymerization was carried out as described in
Figure 5. At the plateau level of the MT assembly (30 min after reaction), a 10 µL-drop of the reaction mixture was removed and processed for EM examination. (
A) the absence of polymerized MTs at 0 min; (
B) the typical morphology of
in vitro assembled MTs at 30 min; (
C) 40 µM As
3+; (
D) 40 µM As
3+ plus 1 mM DTT. Photographs with a final magnification of 22,000 × observed under a Philips 300 transmission electron microscope.
Figure 6.
As
3+-inhibition of MT protein polymerization
in vitro as confirmed by EM examination. The
in vitro MT polymerization was carried out as described in
Figure 5. At the plateau level of the MT assembly (30 min after reaction), a 10 µL-drop of the reaction mixture was removed and processed for EM examination. (
A) the absence of polymerized MTs at 0 min; (
B) the typical morphology of
in vitro assembled MTs at 30 min; (
C) 40 µM As
3+; (
D) 40 µM As
3+ plus 1 mM DTT. Photographs with a final magnification of 22,000 × observed under a Philips 300 transmission electron microscope.
Figure 7.
As3+ binding to tubulins through -SH groups as revealed by [3H]NEM binding assay. “Pure” tubulin free of MAP (1.5 mg/mL) prepared from bovine brain was pretreated with As3+ at indicated concentrations for 2 h at 0 °C, mixed with [3H]NEM (2 µCi/mL), and incubated for an additional 2 h at 37 °C. Proteins were precipitated with 5% TCA and filtered. Radioactivities coupled with collected proteins on the membrane were measured by β-counting. The significance of data presented was assessed by ANOVA analysis. * p < 0.05, ** p < 0.01 (n = 3).
Figure 7.
As3+ binding to tubulins through -SH groups as revealed by [3H]NEM binding assay. “Pure” tubulin free of MAP (1.5 mg/mL) prepared from bovine brain was pretreated with As3+ at indicated concentrations for 2 h at 0 °C, mixed with [3H]NEM (2 µCi/mL), and incubated for an additional 2 h at 37 °C. Proteins were precipitated with 5% TCA and filtered. Radioactivities coupled with collected proteins on the membrane were measured by β-counting. The significance of data presented was assessed by ANOVA analysis. * p < 0.05, ** p < 0.01 (n = 3).
Cellular thiols such as glutathione (GSH) and metallothionein are important in maintaining the integrity of cytoskeletal structures and functions [
21,
22]. Interaction with cellular thiols is the most important biological property of As
3+ [
12,
27]. Thus, we further examined the levels of other cellular thiols such as metallothionein (I and II isoforms) and thiol-related protein such as the heavy chain of γ-GCS, a rate limiting enzyme for GSH synthesis [
28], in As
3+-exposed RFL6 cells. As shown (
Figure 8), treatment of cells with As
3+ at indicated concentrations induced a dose-dependent elevation of cellular levels of metallothionein [
Figure 8(A)] and the γ-GCS heavy chain [
Figure 8(B)]. Thus, RFL6 cells exhibited a sensitive response to As
3+ exposure in terms of altered cellular thiol homeostasis.
Figure 8.
As3+ enhanced expression of metallothionein and γ-GCS. Growth-arrested RFL6 cells were treated with As3+ at indicated concentrations for 24 h and processed for Western blot analysis. (A) metallothionein (I and II); (B) γ-GCS heavy chain; actin as an internal control.
Figure 8.
As3+ enhanced expression of metallothionein and γ-GCS. Growth-arrested RFL6 cells were treated with As3+ at indicated concentrations for 24 h and processed for Western blot analysis. (A) metallothionein (I and II); (B) γ-GCS heavy chain; actin as an internal control.
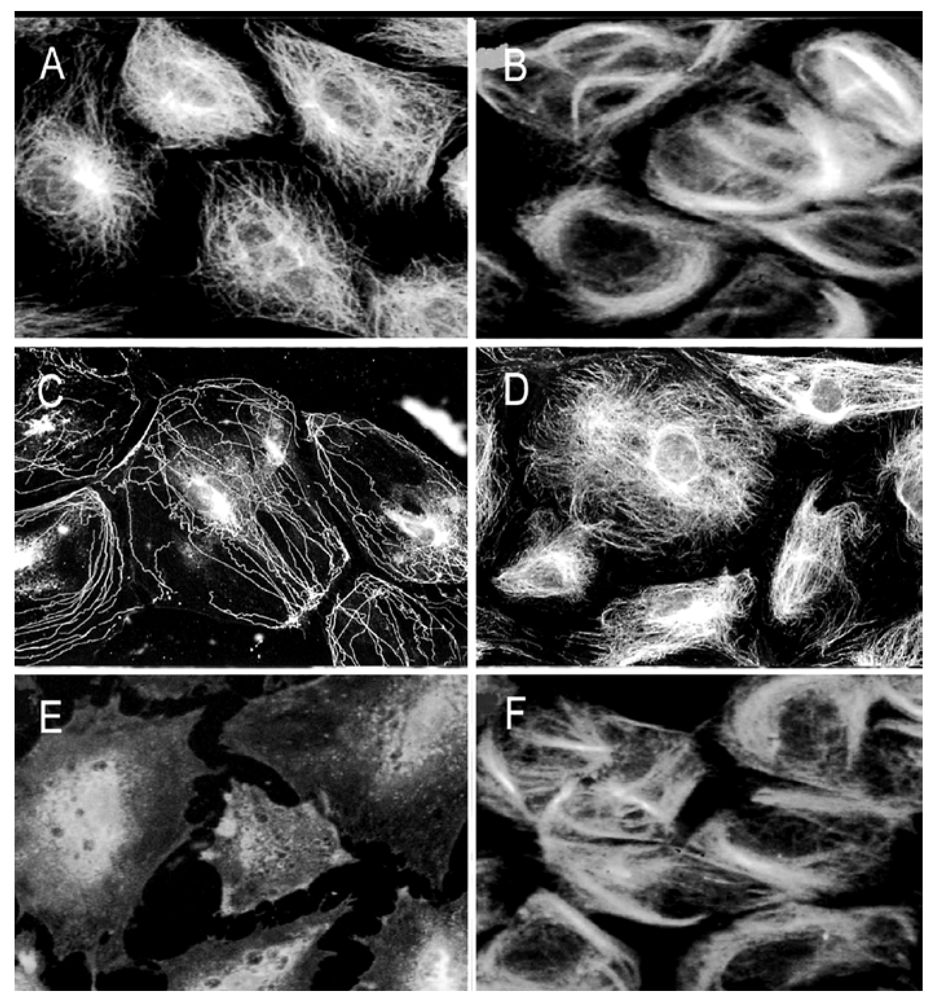
Taxol is an MT stabilizing drug [
13]. To assess taxol protective effect on As
3+-induced MT injury, we examined MT organization in As
3+-treated RFL6 cells in the presence or absence of taxol. In comparison to the control [
Figure 9(A)], treatment of cells with taxol at 20 µM induced MT bundling [
Figure 9(B)]. As expected, 20 µM As
3+ exposure for 24 h induced severe loss of cellular MTs [
Figure 9(C)]. In contrast, co-incubation with 20 µM taxol prevented As
3+-induced MT disruption [
Figure 9(D)]. In addition, taxol also blocked disassembly of MTs induced by NEM (25 µM), a sulfhydryl group binding agent [
21], which was used as a positive control in this experiment [comparing
Figure 9(F), NEM + Taxol with
Figure 9(E), NEM alone]. Thus, taxol antagonizes disassembly of MTs by thiol-bound agents such as As
3+ and NEM.
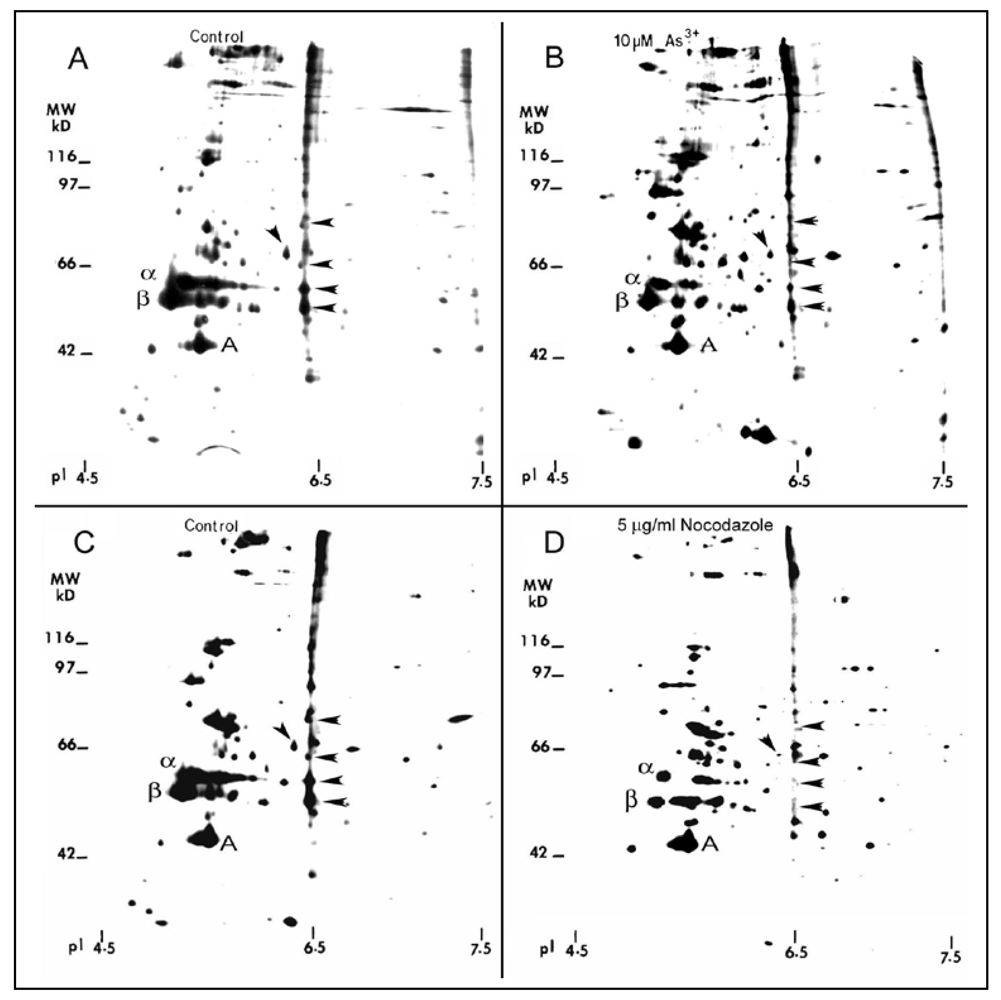
As
3+ damage to the MT organization may be facilitated by alterations in MAPs which are important in maintaining the structural and functional integrity of MTs [
29]. Because MTs are very sensitive to Ca
2+, proteins released by Ca
2+ extraction of the detergent resistant MTs of cells, by definition, contain tubulins and MAPs [
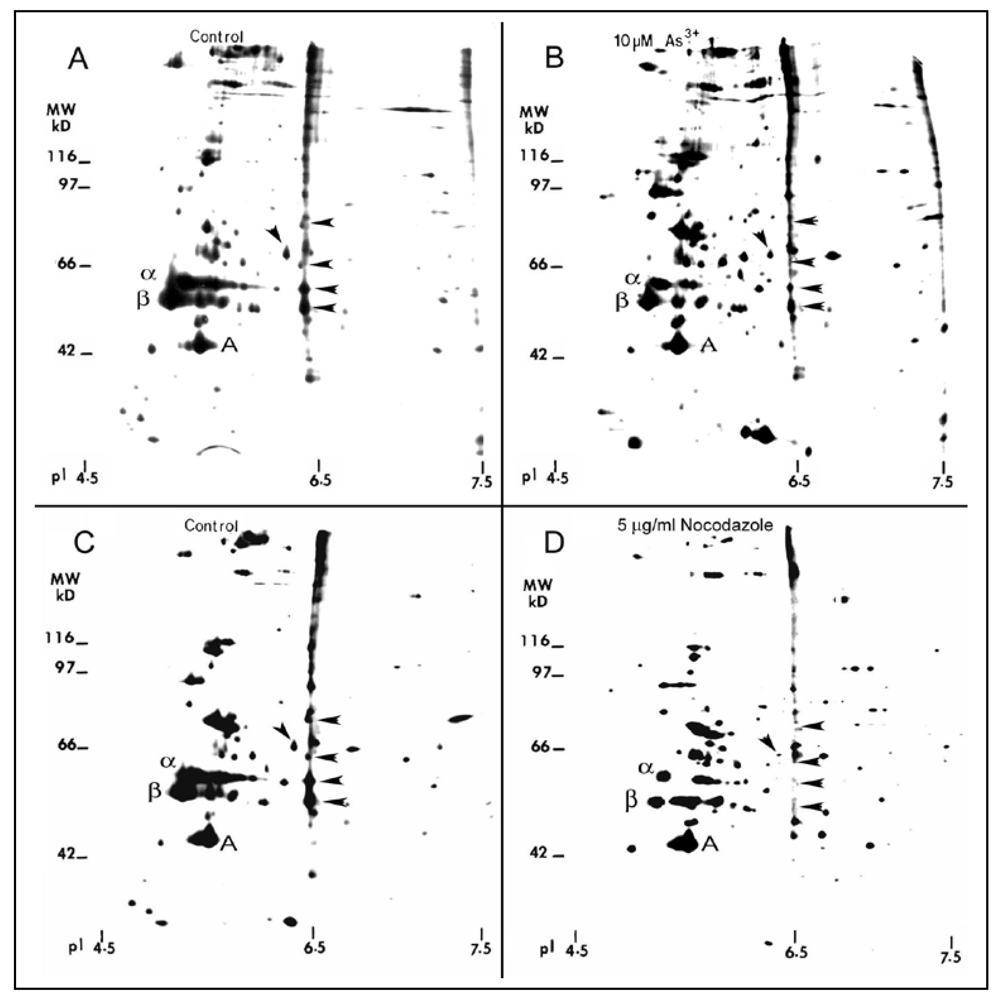
25]. As analyzed by two-dimension (2-D) gels, quiescent RFL6 cells treated with 5 µg/mL nocodazole, a MT disassembly agent, a positive control in this study, for 3 h, displayed marked reduction of MT proteins in association with disappearing several MAPs such as 50/6.5, 53/6.5, 60/6.4, 62/6.3 and 86/6.5 (apparent MW in kDa/isoelectric point) indicated by arrowheads [compare
Figure 10(D) with
Figure 10(C)]. Similarly, cells incubated with 10 µM As
3+ for 24 h exhibited a decrease in α and β tubulins and those MAPs sensitive to nocodazole [arrowheads in
Figure 10(B)] in comparison to the control in the absence of As
3+ treatment [
Figure 10(A)]. Note that the internal control of actin was not significantly changed. Quantitative densitometric analysis indicated that the integrated intensities of MAPs 50/6.5, 53/6.5, 60/6.4, 62/6.3 and 86/6.5 were decreased to 44%, 49%, 15%, 48%, and 12% of the control, respectively. In view of the critical role of MAPs in the MT stability [
29], decreases in the levels of several MAPs would contribute to MT instability favoring their disassembly.
Figure 9.
Taxol antagonistic effects on As3+-induced MT injury. Growth–arrested RFL6 cells on coverslips were treated with As3+, NEM, or their combinations with taxol for 24 h, and then stained with an anti-β tubulin antibody-FITC to visualize MTs. (A) control cells; (B) cells treated with 20 µM taxol; (C) cells treated with 20 µM As3+; (D) cells treated with 20 µM As3+ + 20 µM taxol; (E) cells treated with 25 µM NEM; (F) cells treated with 25 µM NEM + 20 µM taxol.
Figure 9.
Taxol antagonistic effects on As3+-induced MT injury. Growth–arrested RFL6 cells on coverslips were treated with As3+, NEM, or their combinations with taxol for 24 h, and then stained with an anti-β tubulin antibody-FITC to visualize MTs. (A) control cells; (B) cells treated with 20 µM taxol; (C) cells treated with 20 µM As3+; (D) cells treated with 20 µM As3+ + 20 µM taxol; (E) cells treated with 25 µM NEM; (F) cells treated with 25 µM NEM + 20 µM taxol.
Figure 10.
2-D gel profile of tubulins, MAPs, and other cytoskeletal proteins in As3+-treated cells. Growth-arrested RFL6 cells were labeled with [35S]methionine (50 μCi/mL) in the absence (A) or presence (B) of 10 µM As3+ for 24 h. Cells treated without (C) or with (D) 5 µg/mL nocodazole, an MT depolymerization agent, at the last 3 h of the metabolic labeling, were used as a positive control. The MT protein-rich extracts with identical redioactivity from control and treated cells were processed for 2-D gel analysis (α, β and A indicate α tubulin, β tubulin and actin, respectively). For MAP nomenclature, e.g., MAP56/6.5, the numbers refer to their apparent molecular weight (MW in kDa) and isoelectric point (pI), respectively. Protein quantitation was measured by the 1 D Scan EX software. Arrowheads indicate altered MAPs.
Figure 10.
2-D gel profile of tubulins, MAPs, and other cytoskeletal proteins in As3+-treated cells. Growth-arrested RFL6 cells were labeled with [35S]methionine (50 μCi/mL) in the absence (A) or presence (B) of 10 µM As3+ for 24 h. Cells treated without (C) or with (D) 5 µg/mL nocodazole, an MT depolymerization agent, at the last 3 h of the metabolic labeling, were used as a positive control. The MT protein-rich extracts with identical redioactivity from control and treated cells were processed for 2-D gel analysis (α, β and A indicate α tubulin, β tubulin and actin, respectively). For MAP nomenclature, e.g., MAP56/6.5, the numbers refer to their apparent molecular weight (MW in kDa) and isoelectric point (pI), respectively. Protein quantitation was measured by the 1 D Scan EX software. Arrowheads indicate altered MAPs.
4. Discussion
Our previous studies have shown As
3+ perturbation of the cytoskeletal organization and cellular glutathione homeostasis in Swiss 3T 3 cells [
21]. The lung is a major target organ for arsenic. In this report, we further evaluated As
3+ effects on the organization of MTs, a critical element of the cytoskeleton,
in vitro, in cultured rat fetal lung fibroblasts and in lungs of the animal model. RFL6 cells were selected as an
in vitro model in this study because they display a typical MT morphology sensitive to As
3+ injury (
Figure 1). Results indicated that As
3+ disrupted MT assembly by targeting tubulin sulfhytryl groups and MAPs, two major stabilizers of MTs. Our findings showing As
3+-elicited biochemical pathology of MTs are summarized in
Table 1.
Table 1.
Summary for As3+-elicited MT Biochemical Pathology.
Table 1.
Summary for As3+-elicited MT Biochemical Pathology.
| Figure# | Stressor/antagonist | System | MT biochemical pathology by As3+ |
|---|
| 1 | As3+ 5 µM, 20 µM. | MTs in RFL6 cells. | Disassembly of MTs. |
| 2 | As3+ 10 µM. | Spindle MTs and chromosomal patterns in RFL6 cells. | Spindle MT deficiency, cell division aberration, chromosome clustering, scattering and lagging. |
| 3 | As3+ 5 µM, 10 µM, 20 µM. | Tubulin proteins and mRNA in RFL6 cells. | Increasing free tubulin pool, decreasing polymer tubulins, inhibiting βI tubulin mRNA. |
| 4 | As3+ intratracheal instillation (2.02 mg As/kg body in the form of NaAsO2 in 100 μL physiological saline) once a week for 5 weeks. | β-tubulin protein and mRNA in the lung of rats. | Inhibition of lung β-tubulin protein and mRNA levels. |
| 5 | As3+ 20 µM, 40 µM, DTT 1 mM. | Bovine brain tubulin polymerization in vitro. | Inhibition of MT polymerization in vitro and DTT abolishing As3+ effects assessed by turbidimetry. |
| 6 | As3+ 40 µM, DTT 1 mM. | Bovine brain tubulin polymerization in vitro. | Inhibition of MT polymerization in vitro and DTT abolishing As3+ effects conformed by EM. |
| 7 | As3+ 10 µM, 20 µM, 40 µM, [3H]NEM (2 µCi/mL). | NEM binding to pure tubulins free of MAPs in vitro. | Blocking NEM binding to tubulins. |
| 8 | As3+ 5 µM, 10 µM, 20 µM. | Metallothionein and γ-GCS proteins in RFL6 cells. | Enhancement of γ-GCS and metallothionein expression. |
| 9 | As3+ 20 µM, NEM 25 µM, taxol 20 µM. | MTs in RFL6 cells. | Taxol antagonized As3+ and NEM-induced MT disassembly. |
| 10 | As3+ 10 µM, nocodazole 5 µg/mL. | Tubulins and MAPs in RFL6 cells. | Suppression of tubulin and MAP expression, nocodazole as a positive control. |
Arsenic is a metalloid element existing in inorganic and organic forms and in different oxidative states (−3, 0, +3 and +5) which distribute in water, soil and air from nature and anthropogenic sources. High-level exposure to arsenic is involved in workers in metal smelting, wood preservation and semiconductor industries [
1]. The acute inhalation of arsenic dust or fume resulted in gastrointestinal effects, and central and peripheral nervous disorders such as vomiting, diarrhea, bloody urine, anuria, shock, convulsion coma and death [
1,
30]. Chronic exposure to arsenic in humans, by inhalation and oral routes via contaminated water, food and air, are strongly associated with anemia, peripheral neuropathy, lesions and carcinogenesis in the lung, liver, bladder, skin, kidney,
etc. [
1]. As
3+ is 60-times more toxic than As
5+ [
31]. As
3+ has a high affinity for sulfhydryl groups in proteins, such as enzymes, receptors or coenzymes. Binding of As
3+ to critical thiol groups of proteins may be important mechanism for As
3+ toxicity [
12,
27]. Our previous and present studies showing As
3+ depolymerization of MTs indicated that tubilin, a critical cell structure protein, is a target for As
3+ insult.
A dimeric tubulin molecule contains 20 cysteine residues (12 for α and eight for β), most of which are biochemically accessible. Maintenance of certain tubulin -SH groups in the reduced form is crucial for MT polymerization since their loss induces MT disassembly [
32,
33]. One such “assembly-critical” -SH group, for example Cys
239, has been identified in β-tubulin [
33,
34]. Cys
239 mediates the lateral interaction of tubulin molecules required for MT assembly [
33]. Thus, tubulin -SH groups are of a critical stabilizer for MTs. In this report, to identify the mechanism for As
3+ depolymerization of MTs, we performed the
in vitro MT polymerization assay using purified tubulins isolated from bovine brain. As
3+ incubation with tubulin proteins inhibited MT assembly
in vitro and DTT, a thiol-reducing agent, reversed the As
3+ suppression of MT assembly (
Figure 5 and
Figure 6) supporting As
3+ interaction with tubulin -SH groups. One should be noted. As
3+ concentrations used in this
in vitro MT assembly study were higher than those used in cell culture studies. This is most likely due to: (1) low concentrations of MT proteins in cultured cells, and (2) more abundant MAPs in the bovine brain tissue compared to non-neural cells, e.g., fibroblasts [
35]. As
3+ doses used in present studies,
i.e., 5–20 µM (0.374–1.498 ppm) for RFL6 cells, 10–40 µM (0.748–2.996 ppm) for
in vitro tubulin polymerization assays, and 2.02 ppm for the rat animal model, are relevant to human exposure situation. The As content in the well drinking water of some areas in India reaches 3.4 ppm [
36]. Furthermore, using the
in vitro [
3H]NEM binding competition assay (
Figure 7) we demonstrated again As
3+ binding to tubulin -SH groups. Decreases in [
3H]NEM association with tubulins in the protein mixture preincubated with As
3+ provided more direct evidence for As
3+ targeting -SH groups of MT proteins initiating the MT injury cascade. More importantly, As exposure stimulates the formation of the reactive oxygen species (ROS) and activation of oxidative stress signaling such as the NF-
kB pathway in the biological system [
37] which act as an amplification system to further facilitate oxidation of sulfhydryl groups on tubulins and cell injuries. As reported, cells treated with hydrogen peroxide exhibited disruption of the MT cytoskeleton [
38].
Our previous studies have shown As
3+-induced a biphasic response in cellular glutathione (GSH),
i.e., an initial decrease followed by a full restoration and overshooting. Inhibition of cellular GSH by buthionine sulfoximine (BSO), an inhibitor of GSH biosynthesis, apparently increased cell sensitivity to As
3+ toxicity [
21]. Since As
3+ targets cellular thiols we also examined other thiol-containing molecules such as metallothionein as well as γ-GCS, a rate limiting GSH synthetase [
28], in cell response to As
3+. In this report, we illustrated As
3+ enhanced expression of metallothionein and γ-GCS at the protein levels accompanied by depolymerization of MT. The metallothionein molecule with 20 Cys residues can bind six As
3+ while GSH binds As
3+ to form As(SG)3 complex which favors transferring of As
3+ to other thiol containing components [
39,
40]. Increased expression of γ-GCS as an upstream event is required for downstream enhancement of cellular GSH in cells treated with As
3+ [
21]. The metallothionein gene contains multiple copies of the metal responsive element (MRE) in its promoter region. MRE-binding transcription factor-1 (MTF-1), a zinc (Zn) finger protein, interacts with the MRE transactivating the metallothionein gene [
41]. Zn also binds to tubulin proteins stimulating MT assembly
in vitro. Tubulins isolated from brains of Zn-deficient animals showed an impaired ability for polymerization of MTs [
42]. As
3+ upregulated methallothioneins possibly by its binding to sulfhydryls of tubulins or other proteins releasing bound Zn ions that in turn enhance MTF-1 affinity for MRE containing genes such as metallothionein [
41]. The NF-E2-related factor-2 (Nrf2) regulates antioxidant response element (ARE)-mediated expression of γ-GCS heavy subunit and other xenobiotic metabolizing enzymes. Nrf2 binds to the cysteine-rich keap1 protein in the actin filaments as an inactive form in the cytoplasm. Upon oxidative stress such as As
3+ exposure, modification of cysteine residues in the Keap1 protein results in the Nrf2 release and nuclear localization for ARE binding and transactivation of the genes [
43]. Elevation of cellular metallothionein and γ-GCS/GSH may be a critical detoxification mechanism against As
3+ toxicity.
Tubulin heterodimer binds two GTP molecules, one exchangeable in β-tubulin and another one nonexchangeale in α-tubulin. Upon polymerization, GTP bound to β tubulin is hydrolyzed to GDP by intrinsic tubulin GTPase activity. The resulting GDP-tubulin exhibits a reduced affinity for its neighbors and undergoes disassembly [
35]. Thus, MTs exhibit dynamic equilibrium, growing and shrinking by the addition or loss of tubulin dimers from the ends of protofilaments. As
3+ has been reported to inactivate the GTP binding site on tubulins, thus inhibiting the MT assembly
in vitro [
44]. The hydrolysis of GTP to GDP in β-tubulin increases the curvature of protofilaments, destabilizing MTs [
45]. This suggests that modulating the curvature status of MTs can change its stability. Taxol binding to the β-tubulin subunit, specifically to peptides 1–31, 217–231 and 297–293 [
46], has been shown to straighten the GDP protofilament and slow down the transition of protofilaments from straight to a curved configuration [
45]. Taxol stabilization of MTs has been reported to resist depolymerization by Ca
2+, cold temperature, dilution,
etc. [
47]. Here, we go on to demonstrate taxol antagonism on disassembly of MTs by sulfhydryl-directed agents such as As
3+ and NEM. Presumably, As
3+ binding to tubulin -SH-groups may enhance the curvature of protofilaments, thus instabilizing MTs.
In addition to the tubulin dimer, MTs also contain several minor proteins,
i.e., MAPs. The procedures for metabolic labeling, extraction with detergent and Ca
2+, analysis of MAPs and other related proteins on 2-D gels have been used in different cells in response to different agents [
25,
48]. As shown in our studies, As
3+ induced disassembly of MTs. Autoradiograms of proteins present in the MT fraction under As
3+ treatment conditions were compared with its “control” profile derived from cells without As
3+ treatment to identify subtle changes in protein molecular species. A protein that is present in cells with intact MT but changed or absent when intact MT no longer exist in such cells (such as nocodazole and As
3+ treatments) fulfilling the operational definition of MAPs. Because of its high resolution, the quantitative 2-D gel electrophoresis has been the most powerful technique used for identifying unknown MAP proteins. With this 2-D gel system, several MAP proteins have been identified [
25,
48]. However, only five of them were decreased by >50% in cells exposed to 10 μM As
3+ [
Figure 10(B)], indicating that not all MAPs were equally susceptible to this trivalent metalloid element. MAPs exist only in minute amounts in non-neuronal cells. Based on their apparent MW and pI on 2-D gels, none of these MAPs (
Figure 10) appeared to resemble any existing MAP reported in the literature. This is not surprising since most MAPs identified so far, such as MAP1, MAP2, MAP3, Tau family and 210 kD protein (MAP4),
etc., are from either the brain tissue or cells of neural origin [
29,
49,
50,
51]. Notably, several MAP, e.g., MAP-2 and Tau proteins isolated from the brain tissue, stimulate MT assembly and enhance MT stability both
in vitro and
in vivo [
29,
50]. Thus, results in giving for inhibition of MAPs by As
3+ would lead to the enhancement of MT instability.
Immunohistochemical studies indicated severe loss of tubulin synthesis and distribution in As
3+-dosed lungs [
Figure 4(A2)]. Since absorbed As
3+ is distributed throughout the body [
5,
6], all types of lung cells are targets for As
3+ insult including interstitial fibroblasts. In view of MT functions, loss of MTs/tubulins, a cell structural protein, may be a critical basis for As-induced lung pathology such as bronchiectasis, bronchitis, COPD,
etc. Synthesis of tubulin is subjected to a novel feedback autoregulation,
i.e., the level of the free tubulin pool directly controls the level of tubulin expression [
52]. Thus, As
3+ disrupts MTs increasing the free tubulin pool, as a result, inhibiting new tubulin synthesis. Our studies have shown As
3+ downregulation of tubulin synthesis occuring at the mRNA level,
i.e., marked reduction of steady-state βI-tubulin expression in
in vitro and
in vivo models [
Figure 3(B) and
Figure 4(B)]. As reported, the binding of tubulin subunits to the nascent N-terminal tetrapeptide (Met-Arg-Glu-Ile) of β-tubulin triggers the adjacent ribosome to activate an RNase that degrades the polysome bound tubulin mRNA, inducing its destabilization [
53]. In addition to its effects on tubulin mRNA stability, As
3+ may also reduce tubulin mRNA initiation (synthesis) leading to low levels of steady-state tubulin mRNAs in lung cells and tissues.
MTs play a fundamental role in regulation of cell division. Mitosis requires precise execution of a number of MT-based processes such as reduplication and splitting of the centrosome, assembly of a bipolar spindle, attachment of spindle MTs to chromosomes through kinetochores, tension generation on kinetochores by the dynamics of bipolar spindle MTs, and finally pulling each chromatid to one of the spindle poles [
54]. An error in any step of such interdependent subcellular processes could perturb chromosomal organization, resulting in failure of mitosis. In this study we have observed cell division aberrations in the forms of chromosome lagging, scattering and clustering associated with spindle MT injury in the cell model following As
3+ exposure (
Figure 2). These characteristic chromosomal disorientations are also evident in cells treated with colchicine, an MT disassembly reagent [
55]. Disassembly of all spindle MTs by As
3+ may induce chromosomal scattering and clustering at metaphase while partial injure of spindle MT (such as disconnection of a few spindle fibers with chromosomes and/or monopole or multipoles,
etc.) may lead to chromosomal lagging at anaphase/telophase. The latter appears to result from escaping the mitotic block (
i.e., passing through metaphase) as seen [
Figure 2(i)]. Importantly, lagging chromosomes are believed to induce aneuploidy [
56], which is in turn closely related to tumorigenesis [
57]. Oxidation and methylation status of As displayed various effects on the mitotic apparatus [
58]. As
3+-resulted in production of aneuploid cells was associated with disruption of both the MT assembly and the spindle formation [
59]. Thus, As
3+-induced disassembly of spindle MTs and aberrations in the cell division have a critical biological significance for human tumor pathogenesis.






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}