Gills
The total protein content in gills of the fish exposed to lethal and sub lethal concentrations of copper are presented (
Table 7). In gills of fish exposed to 5.5 mg/L, the total protein content was 13.75 mg, 12.30 mg, 11.76 mg and 11.50 mg and that of gills of fish exposed to 0.55 mg/L was 14.75 mg, 14.50 mg, 13.75 mg and 13.50 mg at the end of every 24h exposure period and up to 96h. A decrease of 7.594%, 17.338%, 20.967% and 22.715% in 5.5 mg/l and 0.874%, 2.554%, 7.594% and 9.274% in 0.5 mg/l was observed from control at the end of 24, 48, 72 and 96h, respectively. The ‘t’ test results show that the decrease in the gills of the fish exposed to 5.5 mg/L is significant (
p<0.01) at the end of 48, 72 and 96h while this decrease is significant only at the end of 96h in gills of fish exposed to 0.55 mg/L.
Muscle
Table 8 shows the protein content in muscle of fish exposed to 5.5 mg/L and 0.55 mg/L of copper. Protein content in muscle of fish exposed to 5.5 mg/l was 22.8 mg, 22.46 mg, 20.84 mg and 20.12 mg and that of fish exposed to 0.55 mg/L was 23.3 mg, 22.95 mg, 21.50 mg and 21.39 mg at the end of 24, 48, 72 and 96h, respectively. A decrease of 2.979%, 4.426%, 11.319% and 14.383% in 5.5 mg/l and 0.8511%, 2.340%, 8.511% and 8.979% in 0.55 mg/L was observed at the end of 24, 48, 72 and 96h, respectively. The results of the ‘t’ test show that the decrease in total protein content in muscle of fish exposed to 5.5 mg/L is significant (
p< 0.05) at the end of 96h and that of fish exposed to 0.55 mg/L is insignificant.
It is evident that copper is highly toxic to the fish and the decrease in biochemical biomarkers viz., total glycogen and total protein content in gills and muscle of fish exposed 5.5 mg/L and 0.55 mg/L concentrations of copper demonstrated a linear and positive correlation with the concentration and duration of exposure indicating that the decrease in biochemical constituents is time and dose dependent.
Total glycogen content of
Esomus danricus was depleted due to copper toxicity. Similar trends were also observed in other investigations on copper toxicity to freshwater fish. Copper was found to impair glycolysis in
Labeo rohita[
8]. A depletion of glycogen and phosphocreatine in the white muscle was observed in brown trout
Salmo trutta exposed to sub-lethal concentration of copper in soft acidic water [
20]. Decrease in glycogen level in liver of
Oreochromis niloticus was reported with an increase in copper concentration in water [
33]. The levels of carbohydrates and glycogen in aquatic organisms reveal their involvement in the endogenous derivation of energy during stress. Rapid depletion of muscle and liver glycogen reserves in order to compensate the energy needs of fish under acute metallic stress was also reported [
39,
40]. A consistent decrease in the tissue glycogen reserves observed in this study suggests impaired glycogenesis. Further, the decline in the glycogen content might be partly due to its utilization in the formation of glycoproteins and glycolipids, which are essential constituents of various cell and other membranes. Decline in the glycogen content of the tissues of
Esomus danricus may be due to its enhanced utilization since glycogen forms the immediate source of energy to meet the energy demands under metallic stress. It might be also due to the prevalence of hypoxic or anoxic conditions, which normally enhances glycogen utilization. Our studies revealed that copper triggers the onset of hypoxia in
Esomus danricus. The enhanced utilization of glycogen and its consequent depletion in the tissues, therefore suggests the initiation of anaerobic glycolytic pathway by increased glycogenolysis as has been suggested [
37].
There was also depletion in the total protein content in gills and muscle of
Esomus danricus exposed to copper. Protein content can be taken as a biomarker of copper level. The decrease in the tissue proteins observed in the gills and muscle (
Tables 5–
8) could be partly due to their utilization in cell repair and tissue organization with the formation of lipoproteins, which are important cellular constituents, occurring in cell membranes and cell organelles present in cytoplasm. Direct utilization of proteins as immediate source of energy to meet the energy demands also cannot be ruled out [
42]. The effects of metals may result from their binding with biological constituents of the body such as lipids, amino acids, enzymes and proteins [
6]. The depletion in tissue proteins may be due to impaired or low rate of protein synthesis under metallic stress [
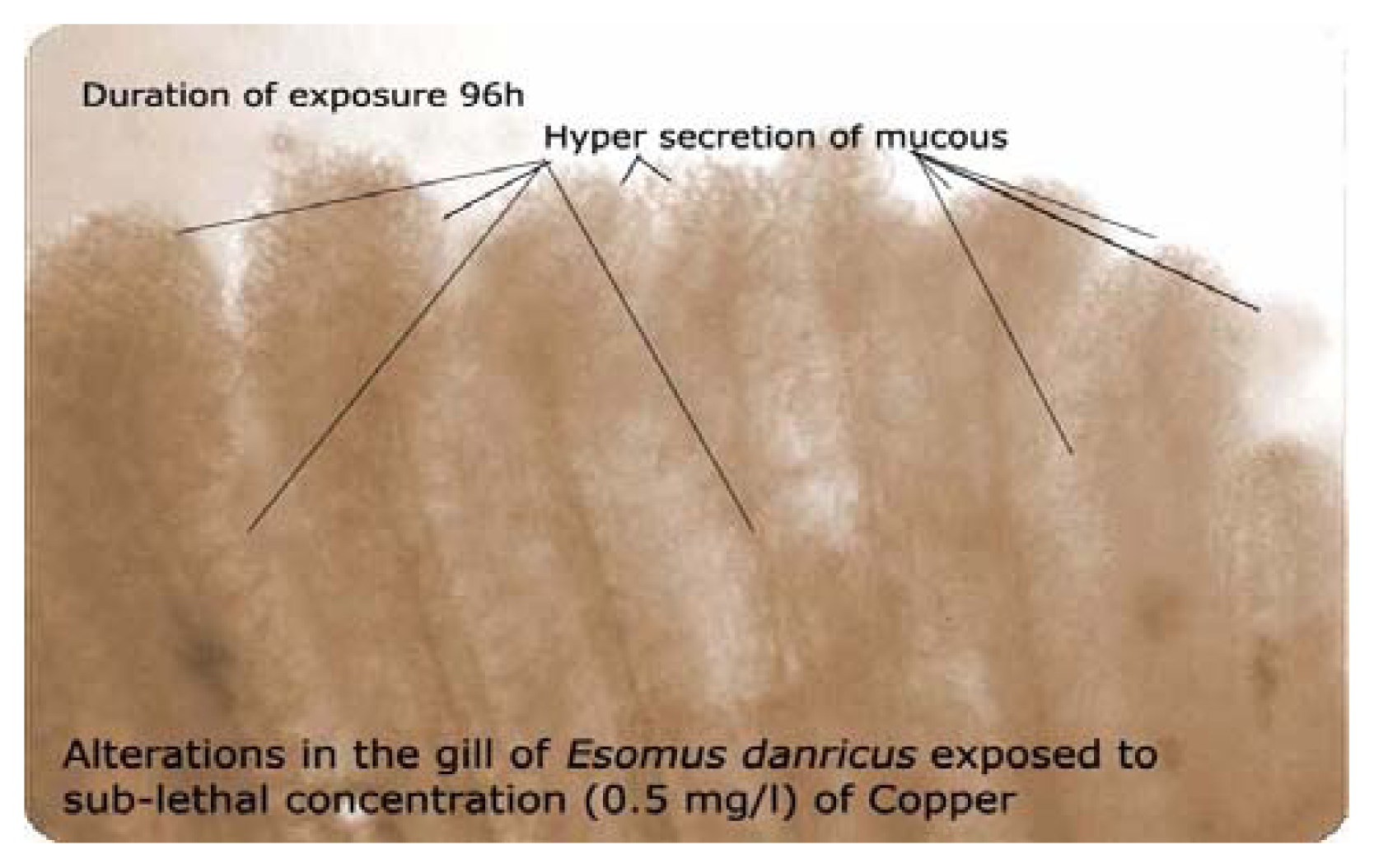
43] or due to their utilization in the formation of mucoproteins, which are eliminated in the form of mucous. The depletion in protein may result in further modification of enzyme activity (stimulation or inhibition). Changes in protein content may also modify signal transfer in cells. Also attack on proteins can lead to the modification of amino acids, oxidation of sulphydryl groups, leading to conformational changes, cross linking, peptide bond cleavage as well as carbohydrate modification in glycoproteins. It can be substantiated that glycogen and protein contents under metallic stress in fish can be effectively employed as biomarkers for rapid assessment of heavy metal toxicity in bio monitoring of aquatic environments.
Enzyme Biomarkers
Table 9 illustrates the activity of superoxide dismutase and catalase in the viscera of the fish exposed to lethal and a sub lethal concentration of CuSO4.5H2O, respectively. The results show that there was significant decrease (
p<0.001) in the activity of superoxide dismutase in the viscera of the fish exposed to lethal and sub lethal concentrations of copper for 96h. There was significant (
p<0.001) decrease in the Catalase activity in viscera of the fish exposed to lethal and sub-lethal concentrations of CuSO
4.5H
2O for both 24 and 96h. From the results it is evident that there is decrease in the antioxidant enzymes activity in the fish exposed to lethal and sub-lethal concentrations of CuSO
4.5H
2O.
This decrease increased with duration of exposure and concentration of the metal. Appreciable decline in SOD activity in the viscera of silver fish,
Esomus danricus was found under the impact of Cu toxicity when exposed to lethal and sub lethal concentrations for 96h. The decrease in the activity of the enzyme SOD could be due to its inhibition by excess production of ROS, decreased uptake in the diet, alteration in the gene expression or due to protein precipitation [
45,
46].
Catalase activity declined significantly in the viscera of silver fish,
Esomus danricus exposed to lethal and sub lethal concentrations for both 24 and 96h. The decrease in the Catalase activity could be due to its inactivation by Superoxide radical or due to decrease in the rate of reaction as a result of excess production of H
2O
2. Earlier reports attributed the inhibition of catalase activity due to Superoxide radical and demonstrated the synergism between the SOD and Catalase [
47]. A decrease in the activity of catalase is due to excess free radical production by Cu, which results in accumulation of O
2−, and H
2O
2, which in addition to direct generation by Cu is also produced by dismutation of O
2− by superoxide dismutase. The H
2O
2 produced reacts with O
2− to form more reactive ROS like OH, which damages the cellular components like DNA, proteins and lipids. Our studies also revealed a significant decrease in protein profiles of the fish, which substantiates the findings of the present study. A consistent decrease in the antioxidant enzyme levels was observed in the study suggesting that there is impaired antioxidant defense mechanism due to excess generation of oxy radicals by Cu. Further, decline in antioxidant levels might also be due to overproduction of ROS which creates a net increase in the amount of oxygen free radicals present in the cell. Antioxidants like SOD act as primary preventive inhibitor by catalyzing the conversion of superoxide anion (O
2−) to H
2O
2 and O
2 and scavenge the O
2 produced by rapid dismutation reaction. Catalase functions to rapidly dismutase H
2O
2 to water and oxygen [
48]. Therefore any significant reduction in these antioxidant levels results in lipid peroxidation, as normal levels of antioxidants produced could not quench the excess free radicals generated. The findings of the present study clearly demonstrated that enzymatic biomarkers like SOD and CAT in fish can be effectively used for rapid assessment of copper toxicity in biomonitoring of aquatic environment.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}