Anaerobic Dechlorination by a Humin-Dependent Pentachlorophenol-Dechlorinating Consortium under Autotrophic Conditions Induced by Homoacetogenesis


Abstract
:1. Introduction
2. Materials and Methods
2.1. Humin
2.2. Humin-Dependent PCP-Dechlorinating Culture (HMBC Culture)
2.3. Homoacetogenic Culture (HC Culture)
2.4. Humin-Dependent PCP-Dechlorinating Culture with Acetogenic Activity (M-CO Culture)
2.5. Effect of Conditions on the PCP Dechlorination in the Mixed Consortium M-CO
2.6. Oxidized Humin Preparation
2.7. Chemical and Headspace Analyses
2.8. Microbial Community Structure
3. Results
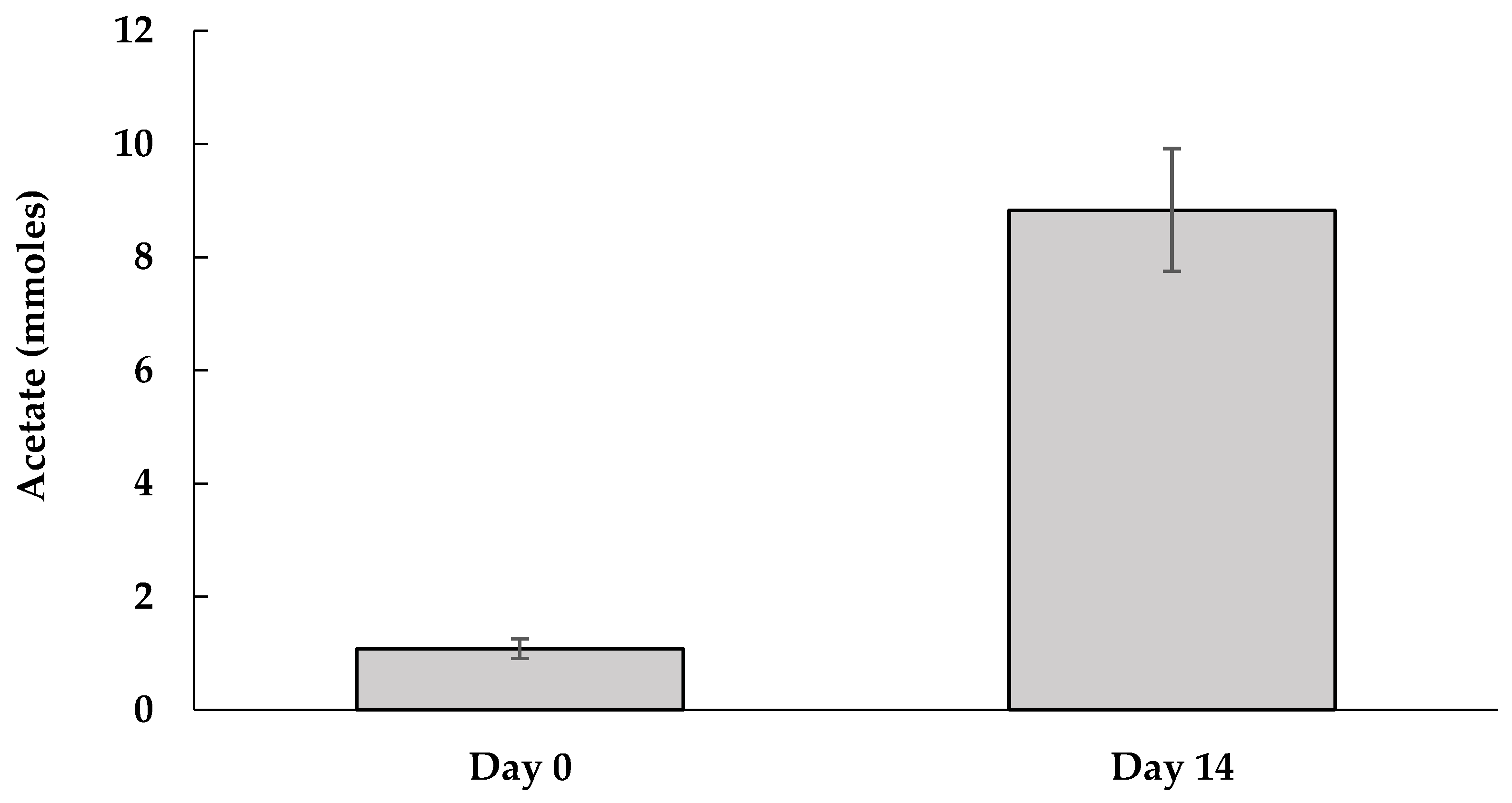
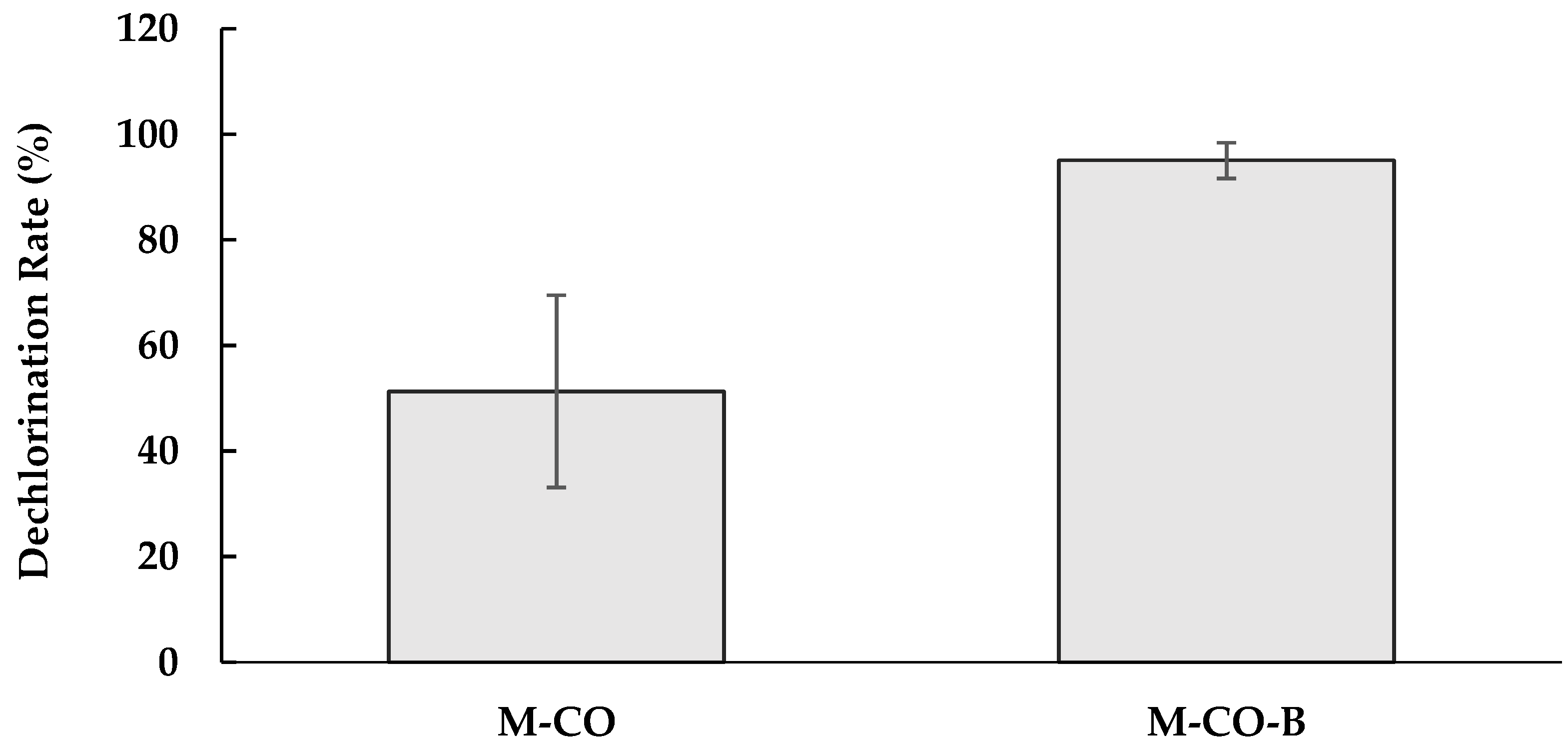
3.1. Dechlorination of PCP by M-CO Culture
3.2. Inhibitory Factors in the M-CO Culture
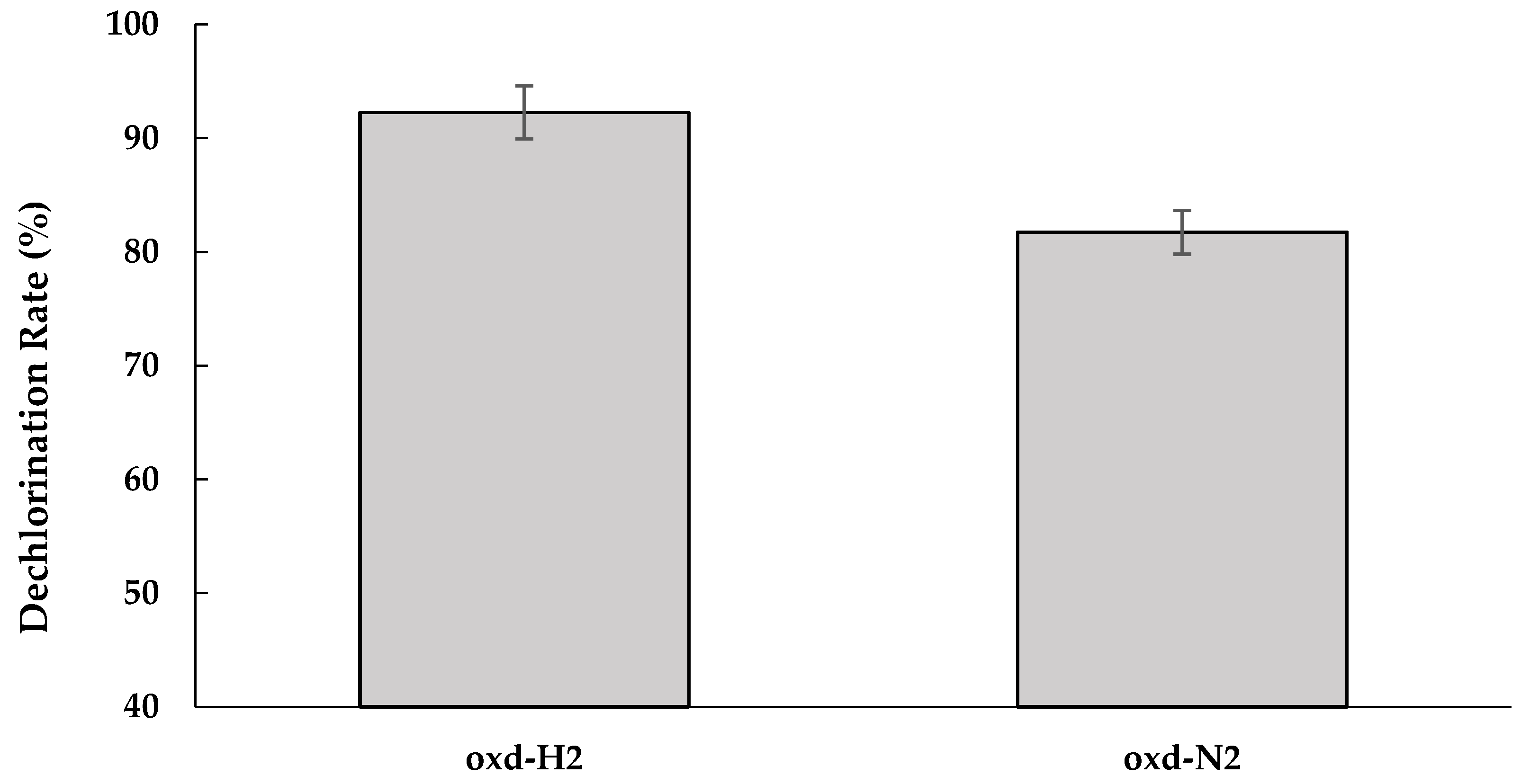
3.3. Role of Humin in Reductive Dechlorination
3.4. Microbial Composition of M-CO Culture
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Bayen, S.; Wurl, O.; Karuppiah, S.; Sivasothi, N.; Hian, K.L.; Obbard, J.P. Persistent organic pollutants in mangrove food webs in Singapore. Chemosphere 2005, 61, 303–313. [Google Scholar] [CrossRef] [PubMed]
- Manz, M.; Wenzel, K.D.; Dietze, U.; Schüürmann, G. Persistent organic pollutants in agricultural soils of central Germany. Sci. Total Environ. 2001, 277, 187–198. [Google Scholar] [CrossRef]
- Weber, K.; Goerke, H. Persistent organic pollutants (POPs) in antarctic fish: Levels, patterns, changes. Chemosphere 2003, 53, 667–678. [Google Scholar] [CrossRef]
- Zhang, Z.L.; Hong, H.S.; Zhou, J.L.; Huang, J.; Yu, G. Fate and assessment of persistent organic pollutants in water and sediment from Minjiang River Estuary, Southeast China. Chemosphere 2003, 52, 1423–1430. [Google Scholar] [CrossRef]
- Holliger, C.; Gaspard, S.; Glod, G.; Heijman, C.; Schumacher, W.; Schwarzenbach, R.P.; Vazquez, F. Contaminated environments in the subsurface and bioremediation: Organic contaminants. FEMS Microbiol. Rev. 1997, 20, 517–523. [Google Scholar] [CrossRef]
- Lovley, D.R.; Chapelle, F.H. Deep subsurface microbial processes. Rev. Geophys. 1995, 33, 365–381. [Google Scholar] [CrossRef]
- Adrian, L.; Löffler, F.E. Organohalide-Respiring Bacteria; Springer: Berlin/Heidelberg, Germany, 2016; ISBN 978-3-662-49875-0. [Google Scholar]
- Nicholson, D.K.; Woods, S.L.; Istok, J.D.; Peek, D.C. Reductive Dechlorination of Chlorophenols by a Pentachlorophenol-Acclimated Methanogenic Consortium. Appl. Environ. Microbiol. 1992, 58, 2280–2286. [Google Scholar]
- Strycharz, S.M.; Gannon, S.M.; Boles, A.R.; Franks, A.E.; Nevin, K.P.; Lovley, D.R. Reductive dechlorination of 2-chlorophenol by Anaeromyxobacter dehalogenans with an electrode serving as the electron donor. Environ. Microbiol. Rep. 2010, 2, 289–294. [Google Scholar] [CrossRef]
- Watanabe, I. Pentachlorophenol (PCP) decomposing activity of field soils treated annually with PCP. Soil Biol. Biochem. 1978, 10, 71–75. [Google Scholar] [CrossRef]
- Holliger, C.; Schraa, G. Physiological meaning and potential for application of reductive dechlorination by anaerobic bacteria. FEMS Microbiol. Rev. 1994, 15, 297–305. [Google Scholar] [CrossRef]
- Dolfing, J.; Novak, I. The Gibbs free energy of formation of halogenated benzenes, benzoates and phenols and their potential role as electron acceptors in anaerobic environments. Biodegradation 2014, 26, 15–27. [Google Scholar] [CrossRef] [Green Version]
- Dercová, K.; Sejáková, Z.; Skokanová, M.; Barančíková, G.; Makovníková, J. Bioremediation of soil contaminated with pentachlorophenol (PCP) using humic acids bound on zeolite. Chemosphere 2007, 66, 783–790. [Google Scholar] [CrossRef]
- Schink, B. Microbially driven redox reactions in anoxic environments: Pathways, Energetics, and biochemical consequences. Eng. Life Sci. 2006, 6, 228–233. [Google Scholar] [CrossRef]
- Lovley, D.R. Microbial Fe(III) reduction in subsurface environments. FEMS Microbiol. Rev. 1997, 20, 305–313. [Google Scholar] [CrossRef]
- Zhang, C.; Katayama, A. Humin as an electron mediator for microbial reductive dehalogenation. Environ. Sci. Technol. 2012, 46, 6575–6583. [Google Scholar] [CrossRef]
- Zhang, D.; Zhang, C.; Li, Z.; Suzuki, D.; Komatsu, D.D.; Tsunogai, U.; Katayama, A. Electrochemical stimulation of microbial reductive dechlorination of pentachlorophenol using solid-state redox mediator (humin) immobilization. Bioresour. Technol. 2014, 164, 232–240. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.; Zhang, C.; Xiao, Z.; Suzuki, D.; Katayama, A. Humin as an electron donor for enhancement of multiple microbial reduction reactions with different redox potentials in a consortium. J. Biosci. Bioeng. 2015, 119, 188–194. [Google Scholar] [CrossRef]
- Zhang, C.; Zhang, D.; Xiao, Z.; Li, Z.; Suzuki, D.; Katayama, A. Characterization of humins from different natural sources and the effect on microbial reductive dechlorination of pentachlorophenol. Chemosphere 2015, 131, 110–116. [Google Scholar] [CrossRef]
- Zhang, C.; Li, Z.; Suzuki, D.; Ye, L.; Yoshida, N.; Katayama, A. A humin-dependent Dehalobacter species is involved in reductive debromination of tetrabromobisphenol A. Chemosphere 2013, 92, 1343–1348. [Google Scholar] [CrossRef]
- Xiao, Z.; Awata, T.; Zhang, D.; Zhang, C.; Li, Z.; Katayama, A. Enhanced denitrification of Pseudomonas stutzeri by a bioelectrochemical system assisted with solid-phase humin. J. Biosci. Bioeng. 2016, 122, 85–91. [Google Scholar] [CrossRef]
- Schnobrich, M.R.; Chaplin, B.P.; Semmens, M.J.; Novak, P.J. Stimulating hydrogenotrophic denitrification in simulated groundwater containing high dissolved oxygen and nitrate concentrations. Water Res. 2007, 41, 1869–1876. [Google Scholar] [CrossRef]
- Fang, Y.; Hozalski, R.M.; Clapp, L.W.; Novak, P.J.; Semmens, M.J. Passive dissolution of hydrogen gas into groundwater using hollow-fiber membranes. Water Res. 2002, 36, 3533–3542. [Google Scholar] [CrossRef]
- Xiao, Z.; Awata, T.; Zhang, D.; Katayama, A. Denitrification of Pseudomonas stutzeri coupled with CO2 reduction by Sporomusa ovata with hydrogen as electron donor assisted with solid-phase humin. J. Biosci. Bioeng. 2016, 122, 307–313. [Google Scholar] [CrossRef]
- Ragsdale, S.W.; Pierce, E. Acetogenesis and the Wood-Ljungdahl pathway of CO2 fixation. Biochim. Biophys. Acta Proteins Proteom. 2008, 1784, 1873–1898. [Google Scholar] [CrossRef]
- Nevin, K.P.; Hensley, S.A.; Franks, A.E.; Summers, Z.M.; Ou, J.; Woodard, T.L.; Snoeyenbos-West, O.L.; Lovley, D.R. Electrosynthesis of organic compounds from carbon dioxide is catalyzed by a diversity of acetogenic microorganisms. Appl. Environ. Microbiol. 2011, 77, 2882–2886. [Google Scholar] [CrossRef]
- Luijten, M.L.G.C.; Roelofsen, W.; Langenhoff, A.A.M.; Schraa, G.; Stams, A.J.M. Hydrogen threshold concentrations in pure cultures of halorespiring bacteria and at a site polluted with chlorinated ethenes. Environ. Microbiol. 2004, 6, 646–650. [Google Scholar] [CrossRef]
- Schuchmann, K.; Müller, V. Energetics and Application of Heterotrophy in Acetogenic Bacteria. Appl. Environ. Microbiol. 2016, 82, 4056–4069. [Google Scholar] [CrossRef] [Green Version]
- Yoshida, N.; Yoshida, Y.; Handa, Y.; Kim, H.K.; Ichihara, S.; Katayama, A. Polyphasic characterization of a PCP-to-phenol dechlorinating microbial community enriched from paddy soil. Sci. Total Environ. 2007, 381, 233–242. [Google Scholar] [CrossRef]
- Widdel, F.; Kohring, G.-W.; Mayer, F. Studies on Dissimilatory Sulfate-Reducing Bacteria that Decompose Fatty Acids III. Characterization of the Filamentous Gliding Desulfonemalimicolagen. nov. sp. nov. and Desulfonemamagnumsp. nov. Arch. Microbiol. 1983, 134, 286–294. [Google Scholar] [CrossRef]
- Takahashi, S.; Tomita, J.; Nishioka, K.; Hisada, T.; Nishijima, M. Development of a prokaryotic universal primer for simultaneous analysis of Bacteria and Archaea using next-generation sequencing. PLoS ONE 2014, 9, e105592. [Google Scholar] [CrossRef]
- Edgar, R.C. Search and clustering orders of magnitude faster than BLAST. Bioinformatics 2010, 26, 2460–2461. [Google Scholar] [CrossRef] [Green Version]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Peña, A.G.; Goodrich, J.K.; Gordon, J.I.; et al. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef] [Green Version]
- McDonald, D.; Price, M.N.; Goodrich, J.; Nawrocki, E.P.; Desantis, T.Z.; Probst, A.; Andersen, G.L.; Knight, R.; Hugenholtz, P. An improved Greengenes taxonomy with explicit ranks for ecological and evolutionary analyses of bacteria and archaea. ISME J. 2012, 6, 610–618. [Google Scholar] [CrossRef]
- Elfantroussi, S.; Naveau, H.; Agathos, S.N. Anaerobic dechlorinating bacteria. Biotechnol. Prog. 1998, 14, 167–188. [Google Scholar] [CrossRef]
- Diekert, G.; Wohlfarth, G. Metabolism of homoacetogens. Antonie Van Leeuwenhoek 1994, 66, 209–221. [Google Scholar] [CrossRef]
- Holliger, C.; Wohlfarth, G.; Diekert, G. Reductive dechlorination in the energy metabolism of anaerobic bacteria. FEMS Microbiol. Rev. 1998, 22, 383–398. [Google Scholar] [CrossRef]
- Adam, P.S.; Borrel, G.; Gribaldo, S. Evolutionary history of carbon monoxide dehydrogenase/acetyl-CoA synthase, one of the oldest enzymatic complexes. Proc. Natl. Acad. Sci. USA 2018, 115, E1166–E1173. [Google Scholar] [CrossRef]
- Müller, V.; Frerichs, J. Acetogenic Bacteria. eLS 2013, 1–9. [Google Scholar] [CrossRef]
- Rainey, F.A. Oxobacter. In Bergey’s Manual of Systematics of Archaea and Bacteria (online); John Wiley & Sons, Ltd.: Chichester, UK, 2015; pp. 1–4. ISBN 9781118960608. [Google Scholar]
- Baena, S.; Patel, B.K.C. Caloramator. In Bergey’s Manual of Systematics of Archaea and Bacteria (online); John Wiley & Sons, Ltd.: Chichester, UK, 2015; pp. 1–10. ISBN 9781118960608. [Google Scholar]
- Esquivel-Elizondo, S.; Delgado, A.G.; Krajmalnik-Brown, R. Evolution of microbial communities growing with carbon monoxide, hydrogen, and carbon dioxide. FEMS Microbiol. Ecol. 2017, 93, 1–12. [Google Scholar] [CrossRef]
- Kuever, J. The family desulfovibrionaceae. In The Prokaryotes: Deltaproteobacteria and Epsilonproteobacteria; Rosenberg, E., DeLong, E.F., Lory, S., Stackebrandt, E., Thompson, F., Eds.; Springer: Berlin/Heidelberg, Germany, 2014; pp. 107–133. [Google Scholar]
- Kim, B.-R.; Shin, J.; Guevarra, R.B.; Lee, J.H.; Kim, D.W.; Seol, K.-H.; Lee, J.-H.; Kim, H.B.; Isaacson, R.E. Deciphering Diversity Indices for a Better Understanding of Microbial Communities. J. Microbiol. Biotechnol. 2017, 27, 2089–2093. [Google Scholar] [CrossRef] [Green Version]
- Battino, R. The Ostwald coefficient of gas solubility. Fluid Phase Equilibria 1984, 15, 231–240. [Google Scholar] [CrossRef]
- Ferry, J.G. CO Dehydrogenase of Methanogens. In Acetogenesis; Chapman and Hall Microbiology Series; Drake, H.I., Ed.; Springer: Boston, MA, USA, 1994; pp. 539–556. [Google Scholar]
- Oren, A. There Must be an Acetogen Somewhere. Front. Microbiol. 2012, 3, 2–3. [Google Scholar] [CrossRef]
- Ozuolmez, D.; Na, H.; Lever, M.A.; Kjeldsen, K.U.; Jørgensen, B.B.; Plugge, C.M. Methanogenic archaea and sulfate reducing bacteria co-cultured on acetate: Teamwork or coexistence? Front. Microbiol. 2015, 6, 1–12. [Google Scholar] [CrossRef]
- Cord-Ruwisch, R.; Seitz, H.-J.; Conrad, R. The capacity of hydrogenotrophic anaerobic bacteria to compete for traces of hydrogen depends on the redox potential of the terminal electron acceptor. Arch. Microbiol. 1988, 149, 350–357. [Google Scholar] [CrossRef]
- Madigan, M.T.; Martinko, J.M.; Bender, K.S.; Buckley, D.H.; Stahl, D.A. Brock Biology of Microorganisms, Global Edition: UEL, 15th ed.; Pearson Education Limited: Harlow, UK, 2015; ISBN 1292068310. [Google Scholar]
- Buan, N.R. Methanogens: Pushing the boundaries of biology. Emerg. Top. Life Sci. 2018, 2, 629–646. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Condition Name | Humin (1 g) | Humin State | Medium | Headspace Composition | Composition of Inoculum | Additional Treatment | |
|---|---|---|---|---|---|---|---|
| 10% (v/v) HMBC | 5% (v/v) HC | ||||||
| M-CO | + | Intact | medium Z | H2/CO2 | + | + | − |
| CO | − | − | medium Z | H2/CO2 | + | + | − |
| HC-PCP | − | − | medium Z | H2/CO2 | + | − | − |
| Basal | + | Intact | medium Z | H2/CO2 | − | − | − |
| M-CO-B | + | Intact | medium ZN | H2/CO2 | + | + | − |
| MCO-Na2S | + | Intact | medium ZN | H2/CO2 | + | + | 1.21 mM Na2S |
| oxd-H2 | + | Oxidized | medium ZN | H2/CO2 | + | + | − |
| oxd-N2 | + | Oxidized | medium ZN | N2/CO2 | + | + | − |
| PCP-Sulfate | + | Intact | medium PCP | N2/CO2 | + | − | 0.5 g/L NA2SO4 |
| PCP-Sal | + | Intact | medium PCP | N2/CO2 | + | − | 2.25 g/L NaCl |
| Culture Name | Hydrogen Used | Electron Sink | Electron Balance ** | |||
|---|---|---|---|---|---|---|
| Acetate Produced | Methane Produced | Sulfate Reduction | Dechlorination | |||
| M-CO | 3.973 | 0.062 | 3.366 | 0.484 | 0.003 | 0.061 |
| M-CO-B | 3.082 | 0.067 | 2.576 | - | 0.010 | 0.429 |
| oxd-H2 | 4.942 | 0.455 | 5.096 | - | 0.010 | −0.619 |
| oxd-N2 | - | - | 0.784 | - | 0.008 | −0.776 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Laskar, M.; Awata, T.; Kasai, T.; Katayama, A. Anaerobic Dechlorination by a Humin-Dependent Pentachlorophenol-Dechlorinating Consortium under Autotrophic Conditions Induced by Homoacetogenesis. Int. J. Environ. Res. Public Health 2019, 16, 2873. https://doi.org/10.3390/ijerph16162873
Laskar M, Awata T, Kasai T, Katayama A. Anaerobic Dechlorination by a Humin-Dependent Pentachlorophenol-Dechlorinating Consortium under Autotrophic Conditions Induced by Homoacetogenesis. International Journal of Environmental Research and Public Health. 2019; 16(16):2873. https://doi.org/10.3390/ijerph16162873
Chicago/Turabian StyleLaskar, Mahasweta, Takanori Awata, Takuya Kasai, and Arata Katayama. 2019. "Anaerobic Dechlorination by a Humin-Dependent Pentachlorophenol-Dechlorinating Consortium under Autotrophic Conditions Induced by Homoacetogenesis" International Journal of Environmental Research and Public Health 16, no. 16: 2873. https://doi.org/10.3390/ijerph16162873