Mycotoxins’ Activity at Toxic and Sub-Toxic Concentrations: Differential Cytotoxic and Genotoxic Effects of Single and Combined Administration of Sterigmatocystin, Ochratoxin A and Citrinin on the Hepatocellular Cancer Cell Line Hep3B


 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Cell Culture
2.3. MTT Assay
2.4. SCE Assay
2.5. Statistical Analysis
2.6. Coefficient of Drug Interaction (CDI) Assessment
3. Results
3.1. Effect of CTN, OTA and STER on MTT Reduction in Hep3B Cells
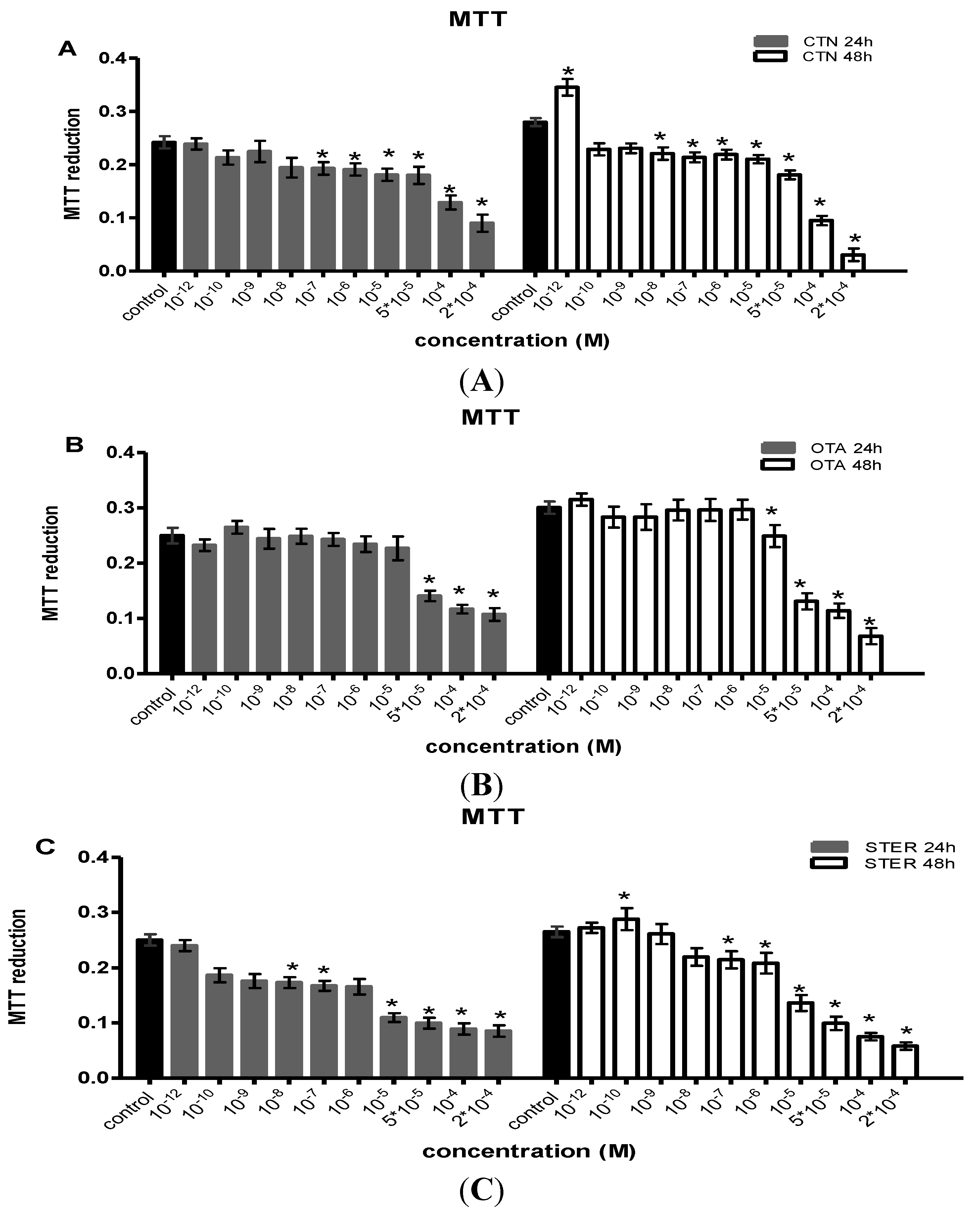

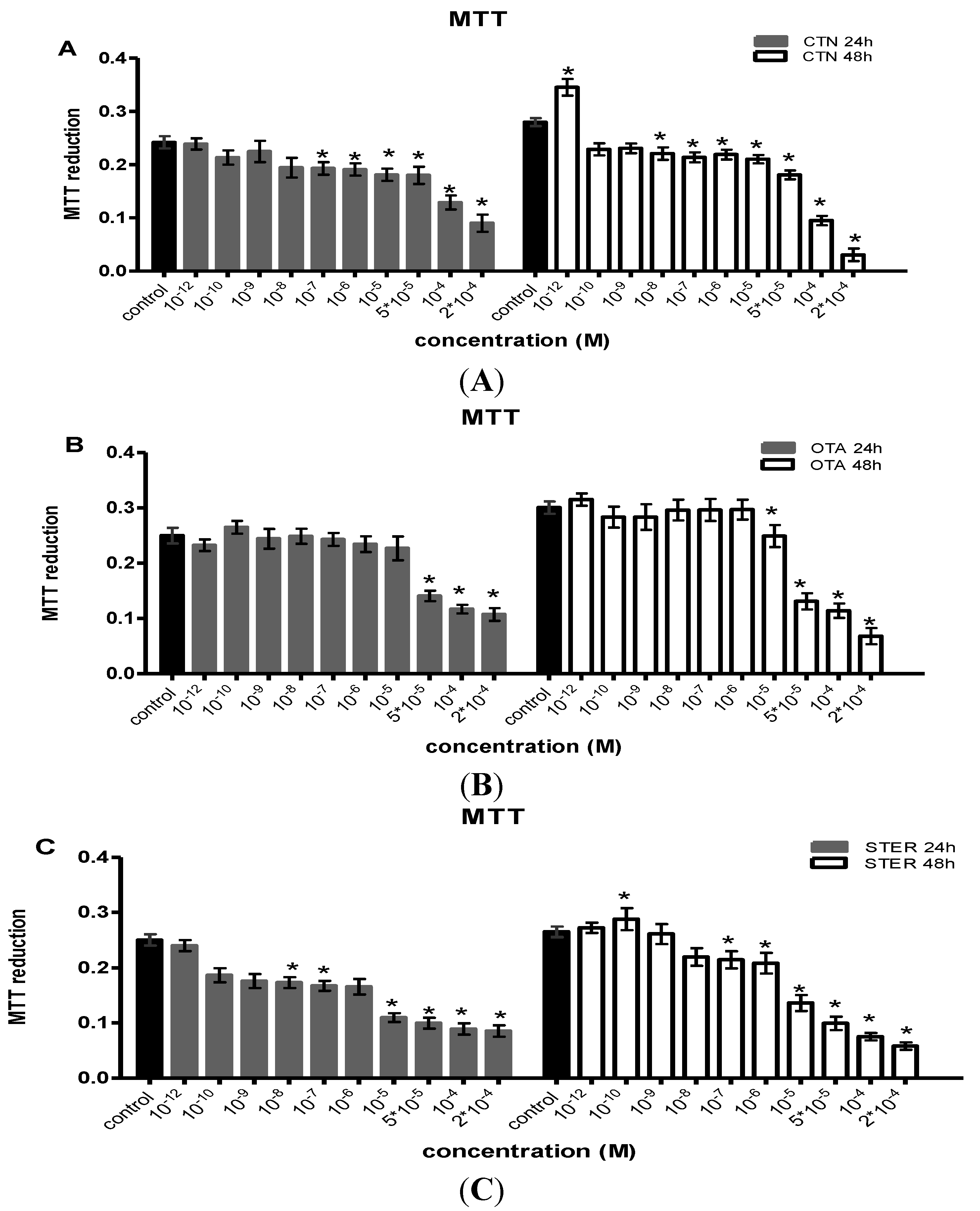{kind=link}
{kind=link}
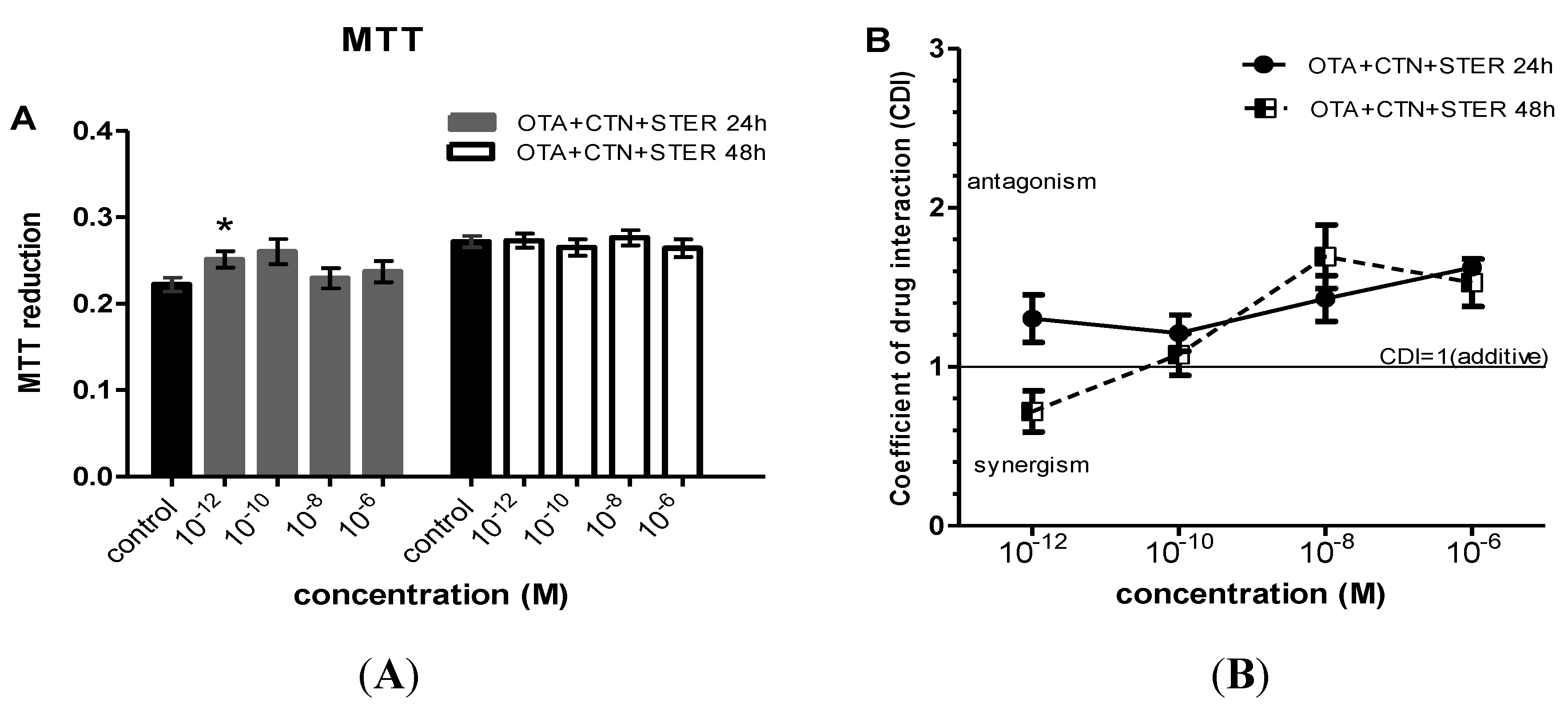{kind=link}
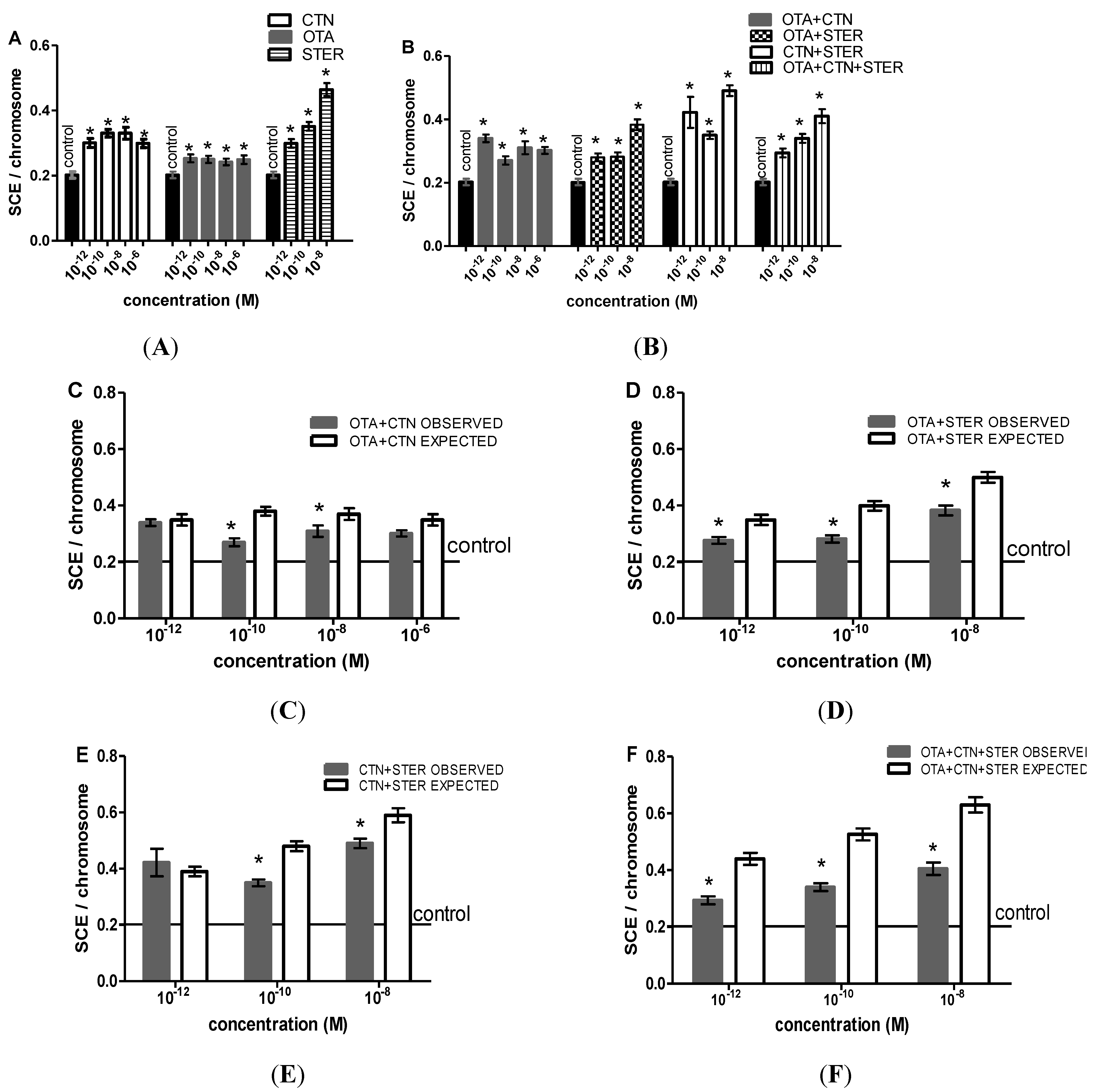{kind=link}
{kind=link}
| 24 h | 48 h | |
|---|---|---|
| IC50 (μM) ± SD | IC50 (μM) ± SD | |
| CTN | 124 ± 4.4 | 77 ± 2.3 |
| OTA | 104 ± 3.1 | 45 ± 1.4 |
| STER | 58 ± 3.1 | 22 ± 0.7 |
3.2. Effect of CTN, OTA and STER Co-Administration on MTT Reduction in Hep3B Cells
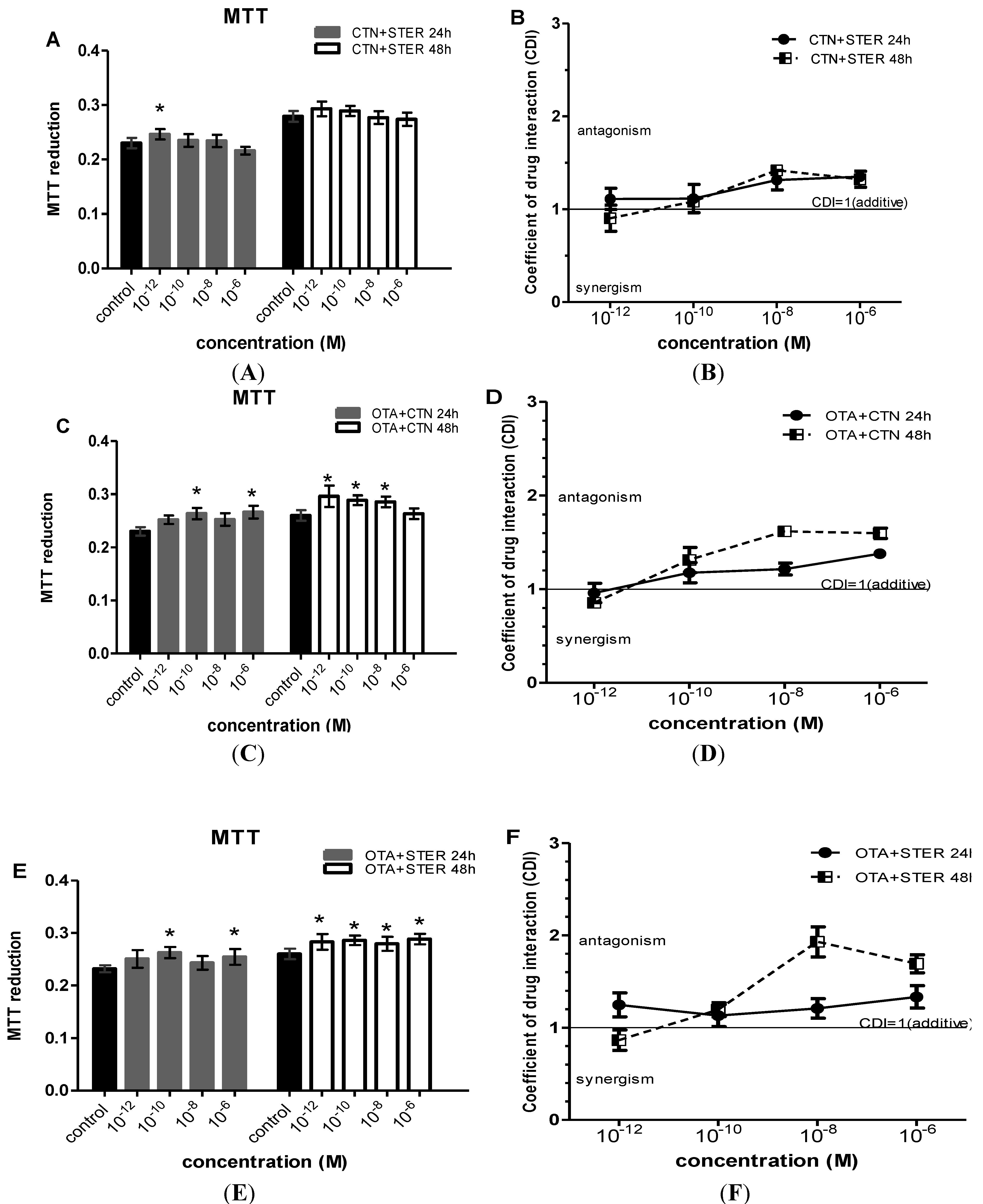

3.3. Cytogenetic Effects of CTN, OTA and STER in Hep3B Cells
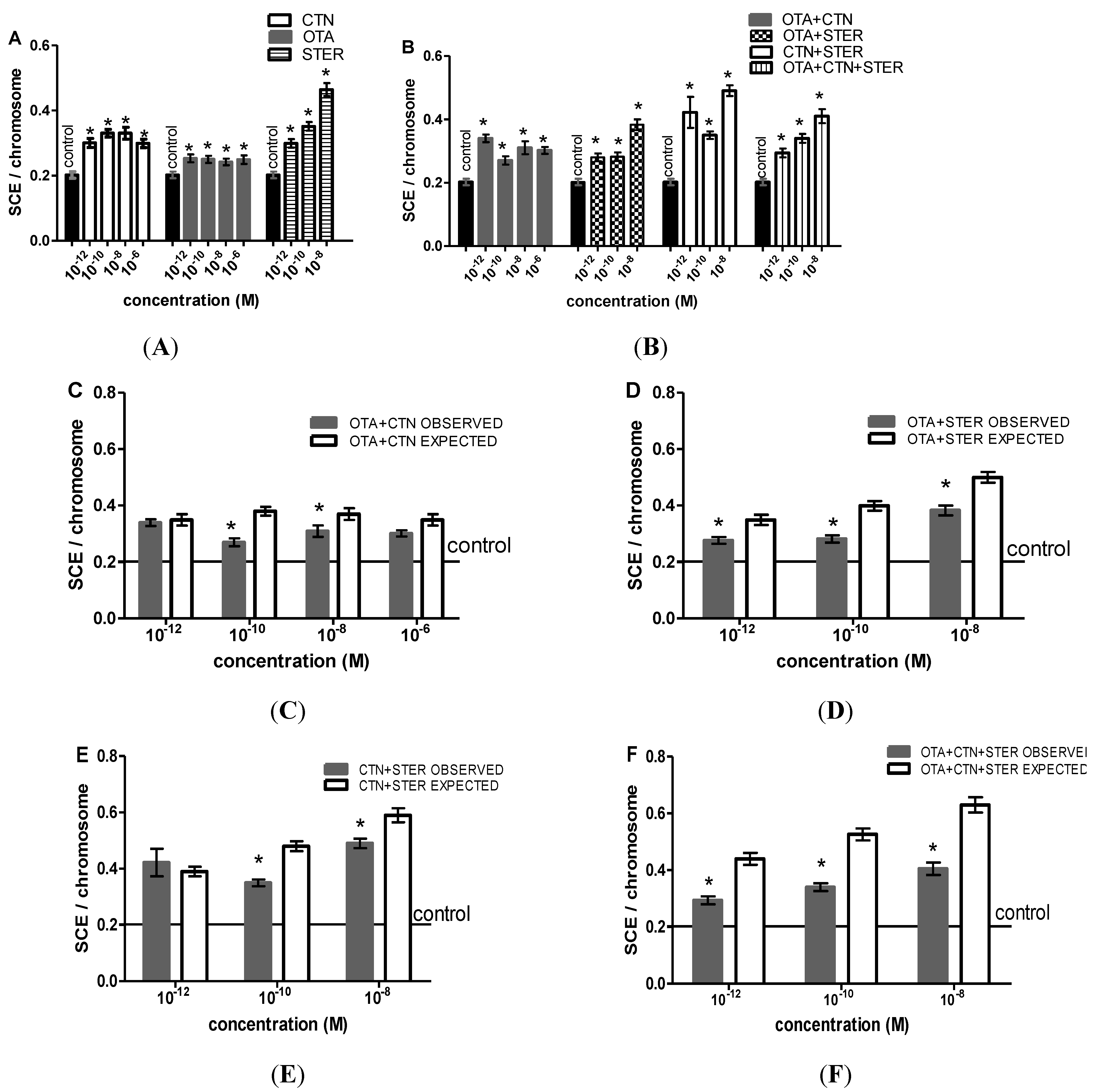

4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Peraica, M.; Radic, B.; Lucic, A.; Pavlovic, M. Toxic effects of mycotoxins in humans. Bull. World Health Organ. 1999, 77, 754–766. [Google Scholar]
- Mycotoxins, Risks in Plant, Animal and Human Systems. Available online: http://www.trilogylab.com/uploads/Mycotoxin_CAST_Report.pdf (accessed on 18 November 2013).
- Chu, F.S. Mycotoxins: Food contamination, mechanism, carcinogenic potential and preventive measures. Mutat. Res. 1991, 259, 291–306. [Google Scholar] [CrossRef]
- Bryden, W.L. Mycotoxins in the food chain: Human health implications. Asia Pac. J. Clin. Nutr. 2007, 16, 95–101. [Google Scholar]
- Shephard, G.S. Impact of mycotoxins on human health in developing countries. Food Addit. Contam. Part A Chem. Anal. Control. Expo. Risk Assess. 2008, 25, 146–151. [Google Scholar] [CrossRef]
- Wild, C.P.; Gong, Y.Y. Mycotoxins and human disease: A largely ignored global health issue. Carcinogenesis 2010, 31, 71–82. [Google Scholar] [CrossRef]
- Berthiller, F.; Schuhmacher, R.; Adam, G.; Krska, R. Formation, determination and significance of masked and other conjugated mycotoxins. Anal. Bioanal. Chem. 2009, 395, 1243–1252. [Google Scholar] [CrossRef]
- Versilovskis, A.; de Saeger, S. Sterigmatocystin: Occurrence in foodstuffs and analytical methods—An overview. Mol. Nutr. Food Res. 2010, 54, 136–147. [Google Scholar] [CrossRef]
- Wang, J.S.; Groopman, J.D. DNA damage by mycotoxins. Mutat. Res. 1999, 424, 167–181. [Google Scholar] [CrossRef]
- Scientific Opinion on the Risk for Public and Animal Health Related to the Presence of Sterigmatocystin in Food and Feed. Available online: http://www.efsa.europa.eu/en/efsajournal/pub/3254.htm (accessed on 18 November 2013).
- Yabe, K.; Nakajima, H. Enzyme reactions and genes in aflatoxin biosynthesis. Appl. Microbiol. Biotechnol. 2004, 64, 745–755. [Google Scholar] [CrossRef]
- International Agency for Research on Cancer (IARC). Some Naturally Occurring Substances. In Monographs on the Evaluation of Carcinogenic Risks to Humans; IARC: Lyon, France, 1976; Volume 10, p. 245. [Google Scholar]
- International Agency for Research on Cancer (IARC). Overall Evaluations of Carcinogenicity: An Updating of IARC Monographs Volumes 1 to 42. In Monographs on the Evaluation of Carcinogenic Risks to Humans; IARC: Lyon, France, 1987; Supplementary 7, p. 449. [Google Scholar]
- Wang, J.; Huang, S.; Xing, L.; Shen, H.; Yan, X.; Wang, J.; Zhang, X. Role of hMLH1 in sterigmatocystin-induced G2 phase arrest in human esophageal epithelial Het-1A cells in vitro. Toxicol. Lett. 2013, 217, 226–234. [Google Scholar] [CrossRef]
- International Agency for Research on Cancer (IARC). Some Naturally Occurring Substances: Food Items and Constituents, Heterocyclic Aromatic Amines and Mycotoxins. In Monographs on the Evaluation of Carcinogenic Risks to Humans; IARC: Lyon, France, 1993; Volume 56. [Google Scholar]
- Miliccevicc, D.; Jovanovicc, M.; Matekalosverak, V.; Radiccevicc, T.; Petrovicc, M.M.; Lilicc, S. A survey of spontaneous occurrence of ochratoxin a residues in chicken tissues and with histopathological changes in liver and kidneys. J. Environ. Sci. Health. C Environ. Carcinog. Ecotoxicol. Rev. 2011, 29, 159–175. [Google Scholar] [CrossRef]
- Gupta, R.S. Veterinary Toxicology; Academic Press: New York, NY, USA, 2007; pp. 939–1018. [Google Scholar]
- Marquardt, R.R.; Frohlich, A.A. A Review of recent advances in understanding ochratoxicosis. J. Anim. Sci. 1992, 70, 3968–3988. [Google Scholar]
- International Agency for Research on Cancer (IARC). Some Naturally Occurring and Synthetic Food Components, Coumarins and Ultraviolet Radiation. In Monographs on the Evaluation of Carcinogenic Risks to Humans; IARC: Lyon, France, 1998; Volume 40. [Google Scholar]
- Scientific Opinion on the Risks for Public Animal and Human Health Related to the Presence of Citrinin in Food and Feed. Available online: http://www.efsa.europa.eu/en/efsajournal/doc/2605.pdf (accessed on 18 November 2013).
- Vrabcheva, T.; Usleber, E.; Dietrich, R.; Martlbauer, E. Co-occurrence of ochratoxin a and citrinin in cereals from bulgarian villages with a history of balkan endemic nephropathy. J. Agric. Food Chem. 2000, 48, 2483–2488. [Google Scholar] [CrossRef]
- Leszkowicz, A.P.; Tozlovanu, M.; Manderville, R.; Peraica, C.M.; Stefanovic, V. New molecular and field evidences for the implication of mycotoxins but not aristolochic acid in human nephropathy and urinary tract tumor. Mol. Nutr. Food Res. 2007, 51, 1131–1146. [Google Scholar] [CrossRef]
- Fan, L.L.; Sun, G.P.; Wei, W.; Wang, Z.G.; Ge, L.; Fu, W.Z.; Wang, H. Melatonin and doxorubicin synergistically induse cell apoptosis in human hepatoma cell lines. World J. Gastroenterol. 2010, 16, 1473–1481. [Google Scholar]
- Hutanasu, C.; Sfarti, C.; Trifan, A.; Cojocariu, C.; Singeap, A.M.; Spac, A.; Stanciu, C. High levels of sterigmatocystinin patients with chronic liver diseases. Rev. Med. Chir. Soc. Med. Nat. Iasi. 2011, 115, 33–37. [Google Scholar]
- Blaszkewicz, M.; Munoz, K.; Degen, G.H. Methods for analysis of citrinin in human blood and urine. Arch. Toxicol. 2013, 87, 1087–1094. [Google Scholar] [CrossRef]
- Ozcelik, N.; Kosar, A.; Soysal, D. Ochratoxin A in human serum samples collected in Isparta-Turkey from healthy individuals and individuals suffering from different urinary disorders. Toxicol. Lett. 2001, 121, 9–13. [Google Scholar] [CrossRef]
- Breitholtz-Emanuelsson, A.; Minervini, F.; Hult, K.; Visconti, A. Ochratoxin A in human serum samples collected in southern Italy from healthy individuals and individuals suffering from different kidney disorders. Nat. Toxins 1994, 2, 366–370. [Google Scholar]
- Skaug, M.A. Levels of ochratoxin A and IgG against conidia of Penicillium verrucosum in blood samples from healthy farm workers. Ann. Agric. Environ. Med. 2003, 10, 73–77. [Google Scholar]
- Prange, A.; Modrow, H.; Hormes, J.; Kramer, J.; Kohler, P. Influence of Mycotoxin producing fungi (Fusarium, Aspergillus, Penicillium) on gluten proteins during suboptimal storage of wheat after harvest and competitive interactions between field and storage fungi. J. Agric. Food Chem. 2005, 53, 6930–6938. [Google Scholar] [CrossRef]
- Guo, L.; Dial, S.; Shi, L.; Branham, W.; Liu, J.; Fang, J.L.; Green, B.; Deng, H.; Kaput, J.; Ning, B. Similarities and differences in the expression of drug-metabolizing enzymes between human hepatic cell lines and primary human hepatocytes. Drug Metab. Despos. 2011, 39, 528–538. [Google Scholar] [CrossRef]
- Mayer, B.J.; Sundermann, V.M.; Darroudi, F.; Laky, B.; Wit, K.; Knasmüller, S. Genotoxic effects of dietary and lifestyle related carcinogens in human derived hepatoma (HepG2, Hep3B) cells. Mutat. Res. 2004, 551, 153–166. [Google Scholar] [CrossRef]
- Volders, M.K.; Decordier, I.; Elhajouji, A.; Plas, G.; Aardema, M.J.; Fenech, M. In vitro genotoxicity testing using the micronucleus assay in cell lines, human lymphocytes and 3D human skin models. Review. Mutagenesis 2011, 26, 177–184. [Google Scholar] [CrossRef]
- Perry, P.; Wolf, S. New Giemsa method for the differential staining of sister chromatids. Nature 1974, 251, 156–158. [Google Scholar] [CrossRef]
- Xu, S.P.; Sun, G.P.; Shen, Y.X.; Peng, W.R.; Wang, H.; Wei, W. Synergistic effect of combining paeonol and cisplatin on apoptotic induction of human hepatoma cell lines. Acta. Pharmacol. Sin. 2007, 28, 869–878. [Google Scholar] [CrossRef]
- Follmann, W.; Hillebrand, I.E.; Creppy, E.E.; Bolt, H.M. Sister chromatid exchange frequency in cultured isolated porcine urinary bladder epithelial cells (PUBEC) treated with ochratoxin A and alpha. Arch. Toxicol. 1995, 69, 280–286. [Google Scholar] [CrossRef]
- Rensburg, S.J.V.; Mozaffari, P.C.; Schalkwyk, D.J.V.; Watt, J.J.V.D.; Vincent, T.J.; Purchase, I.F. Hepatocellular carcinoma and dietary aflatoxin in Mozambique and Transkei. Br. J. Cancer 1985, 51, 713–726. [Google Scholar] [CrossRef]
- Wogan, G.S.; Hecht, S.S.; Felton, J.S.; Conney, A.H.; Loeb, L.A. Environmental and chemical carcinogenesis. Semin. Cancer Biol. 2004, 14, 473–486. [Google Scholar] [CrossRef]
- Weber, F.; Freudinger, R.; Schwerdt, G.; Gekle, M. A rapid screening method to test apoptotic synergisms of ochratoxin A with other nephrotoxic substances. Toxicol. In Vitro 2005, 19, 135–143. [Google Scholar] [CrossRef]
- Bouslimi, A.; Bouaziz, C.; Boussema, I.A.; Hassen, W.; Bacha, H. Individual and combined effects of ochratoxin A and citrinin on viability and DNA fragmentation in cultured Vero cells and on chromosome aberrations in mice bone marrow cells. Toxicology 2008, 251, 1–7. [Google Scholar] [CrossRef]
- Heussner, A.H.; Dietrich, D.R.; Brien, E.O. In vitro investigation of individual and combined cytotoxic effects of ochratoxin A and other selected mycotoxins on renal cells. Toxicol. In Vitro 2006, 20, 332–341. [Google Scholar] [CrossRef]
- Bouaziz, C.; Dein, O.S.; Golli, E.; Essefi, S.A.; Brenner, C.; Lemaire, C.; Bach, H. Different apoptotic pathways induced by zearalenone, T-2 toxin and ochratoxin A in human hepatoma cells. Toxicology 2008, 254, 19–28. [Google Scholar] [CrossRef]
- Bunger, J.; Westphal, G.; Monnich, A.; Hinnendahl, B.; Hallier, E.; Muller, M. Cytotoxicity of occupationally and environmentally relevant mycotoxins. Toxicology 2004, 202, 199–211. [Google Scholar] [CrossRef]
- Jaksic, D.; Puel, O.; Canlet, C.; Kopjar, N.; Kosalec, I.; Klaric, M.S. Cytotoxicity and genotoxicity of versicolorins and 5-methoxysterigmatocystin in A549 cells. Arch. Toxicol. 2012, 86, 1583–1591. [Google Scholar] [CrossRef]
- Chang, C.H.; Yu, F.Y.; Wu, T.S.; Wang, L.T.; Liu, B.H. Mycotoxin Citrinin induced cell cycle G2/m arrest and numerical chromosomal aberration associated with disruption of microtubule formation in human cells. Toxicol. Sci. 2011, 119, 84–92. [Google Scholar] [CrossRef]
- Lioi, M.B.; Santoro, A.; Barbieri, R.; Salzano, S.; Ursini, M.V. Ochratoxin A and zearalenone: A comparative study on genotoxic effects and cell death induced in bovine lymphocytes. Mutat. Res. 2004, 557, 19–27. [Google Scholar] [CrossRef]
- Bernhard, D.; Schwaiger, W.; Crazzolara, R.; Tinhofer, I.; Kofler, R.; Csordas, A. Enhanced MTT-reducing activity under growth inhibition by resveratrol in CEM-C7H2 lymphocytic leukemia cells. Cancer Lett. 2003, 195, 193–199. [Google Scholar] [CrossRef]
- Vistica, D.T.; Skehan, P.; Scudiero, D.; Monks, A.; Pittman, A.; Boyd, M.R. Tetrazolium-based assays for cellular viability: A critical examination of selected parameters affecting formazan production. Cancer Res. 1991, 51, 2515–2520. [Google Scholar]
- Follmann, W.; Lebrun, S.; Kullik, B.; Koch, M.; Romer, H.C.; Golka, K. Cytotoxicity of ochratoxin A and citrinin in different cell types in vitro. Mycotoxin. Res. 2000, 16, 123–126. [Google Scholar] [CrossRef]
- Klaric, M.S.; Zeljezic, D.; Rumora, L.; Peraica, M.; Pepeljnjak, S.; Domijan, A.M. A potential role of calcium in apoptosis and aberrant chromatin forms in porcine kidney PK15 cells induced by individual and combined ochratoxin A and citrinin. Arch. Toxicol. 2012, 86, 97–107. [Google Scholar] [CrossRef]
- Speijers, G.J.A.; Speijers, M.H.M. Combined toxic effects of mycotoxins. Toxicol. Lett. 2004, 153, 91–98. [Google Scholar] [CrossRef]
- Bradley, M.O.; Hsu, I.C.; Harris, C.C. Relationship between sister chromatid exchange and mutagenicity, toxicity and DNA damage. Nature 1979, 282, 318–320. [Google Scholar] [CrossRef]
- Curry, P.T.; Reed, R.N.; Martino, R.M.; Kitchin, R.M. Induction of sister-chromatid exchanges in vivo in mice by the mycotoxins sterigmatocystin and griseofulvin. Mutat. Res. 1984, 137, 111–115. [Google Scholar] [CrossRef]
- Ueda, N.; Fujie, K.; Mimura, K.G.; Chattopadhyay, S.C.; Sugiyama, T. Acute cytogenetic effect of sterigmatocystin on rat bone-marrow cells in vivo. Mutat. Res. 1984, 139, 203–206. [Google Scholar] [CrossRef]
- Hennig, A.; Fink-Gremmels, J.; Leistner, L. Mutagenicity and effects of ochratoxin A on the frequency of sister chromatid exchange after metabolic activation. IARC Sci Publ. 1991, 115, 255–260. [Google Scholar]
- Bendele, A.M.; Neal, S.B.; Oberly, T.J.; Thompson, C.Z.; Bewsey, B.J.; Hill, L.E.; Rexroat, M.A.; Carlton, W.W.; Probst, G.S. Evaluation of ochratoxin A for mutagenicity in a battery of bacterial and mammalian cell assays. Food Chem. Toxicol. 1985, 23, 911–918. [Google Scholar] [CrossRef]
- Liu, B.H.; Yu, F.Y.; Wu, T.S.; Li, S.Y.; Su, M.C.; Wang, M.C.; Shiha, S.M. Evaluation of genotoxic risk and oxidative DNA damage in mammalian cells exposed to mycotoxins, patulin and citrinin. Toxicol. Appl. Pharmacol. 2003, 191, 255–263. [Google Scholar] [CrossRef]
- Cavin, C.; Delatour, T.; Marin-Kuan, M.; Fenaille, F.; Holzhäuser, D.; Guignard, G.; Bezencon, C.; Piguet, D.; Parisod, V.; Richoz-Payot, J.; et al. Ochratoxin A-mediated DNA and protein damage: Roles of nitrosative and oxidative stresses. Toxicol. Sci. 2009, 110, 84–94. [Google Scholar] [CrossRef]
- Liu, J.; Wang, Y.; Cui, J.; Xing, L.; Shen, H.; Wu, S.; Lian, H.; Wang, J.; Yan, X.; Zhang, X. Ochratoxin A induces oxidative DNA damage and G1 phase arrest in human peripheral blood mononuclear cells in vitro. Toxicol. Lett. 2012, 211, 164–171. [Google Scholar]
- Chen, C.C.; Chan, W.H. Inhibition of citrinin-induced apoptotic biochemical signaling in human hepatoma G2 cells by resveratrol. Int. J. Mol. Sci. 2009, 10, 3338–3357. [Google Scholar] [CrossRef]
- Cooke, M.S.; Evans, M.D.; Dizdaroglu, M.; Lunec, J. Oxidative DNA damage: Mechanisms, mutation, and disease. FASEB J. 2003, 17, 1195–1214. [Google Scholar] [CrossRef]
- Nicotera, T.M.; Notaro, J.; Notaro, S.; Schumer, J.; Sandberg, A.A. Elevated superoxide dismutase in bloom’s syndrome: A genetic condition of oxidative stress. Cancer Res. 1989, 49, 5239–5243. [Google Scholar]
- Schaaf, G.J.; Nijmeijer, S.M.; Maas, R.F.; Roestenberg, P.; de Groene, E.M.; Fink-Gremmels, J. The role of oxidative stress in the ochratoxin A-mediated toxicity in proximal tubular cells. Biochim. Biophys. Acta 1588, 149–158. [Google Scholar]
- McConnell, I.R.; Garner, R.C. DNA adducts of aflatoxins, sterigmatocystin and other mycotoxins. IARC Sci. Publ. 1994, 125, 49–55. [Google Scholar]
- Pfohl-Leszkowicz, A.; Grosse, Y.; Kane, A.; Creppy, E.E.; Dirheimer, G. Differential DNA adduct formation and disappearance in three mouse tissues after treatment with the mycotoxin ochratoxin A. Mutat. Res. 1993, 289, 265–273. [Google Scholar] [CrossRef]
- Heflich, R.H.; Morris, S.M.; Beranek, D.T.; McGarrity, L.J.; Chen, J.J.; Beland, F.A. Relationships between the DNA adducts and the mutations and sister chromatid exchanges produced in Chinese hamster ovary cells by N-hydroxy-2-aminofluorene, N-hydroxy-N’-acetylbenzidine and 1-nitrosopyrene. Mutagenesis 1986, 1, 201–206. [Google Scholar]
- Heflich, R.H.; Beranek, D.T.; Kodell, R.L.; Morris, S.M. Induction of mutations and sister-chromatid exchanges in Chinese hamster ovary cells by ethylating agents. Mutat. Res. 1982, 106, 147–161. [Google Scholar] [CrossRef]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Αnninou, N.; Chatzaki, E.; Papachristou, F.; Pitiakoudis, Μ.; Simopoulos, C. Mycotoxins’ Activity at Toxic and Sub-Toxic Concentrations: Differential Cytotoxic and Genotoxic Effects of Single and Combined Administration of Sterigmatocystin, Ochratoxin A and Citrinin on the Hepatocellular Cancer Cell Line Hep3B. Int. J. Environ. Res. Public Health 2014, 11, 1855-1872. https://doi.org/10.3390/ijerph110201855
Αnninou N, Chatzaki E, Papachristou F, Pitiakoudis Μ, Simopoulos C. Mycotoxins’ Activity at Toxic and Sub-Toxic Concentrations: Differential Cytotoxic and Genotoxic Effects of Single and Combined Administration of Sterigmatocystin, Ochratoxin A and Citrinin on the Hepatocellular Cancer Cell Line Hep3B. International Journal of Environmental Research and Public Health. 2014; 11(2):1855-1872. https://doi.org/10.3390/ijerph110201855
Chicago/Turabian StyleΑnninou, Nikolia, Ekaterini Chatzaki, Fotini Papachristou, Μichail Pitiakoudis, and Constantinos Simopoulos. 2014. "Mycotoxins’ Activity at Toxic and Sub-Toxic Concentrations: Differential Cytotoxic and Genotoxic Effects of Single and Combined Administration of Sterigmatocystin, Ochratoxin A and Citrinin on the Hepatocellular Cancer Cell Line Hep3B" International Journal of Environmental Research and Public Health 11, no. 2: 1855-1872. https://doi.org/10.3390/ijerph110201855