Climate Change and Public Health Policy: Translating the Science


Abstract
:1. Introduction

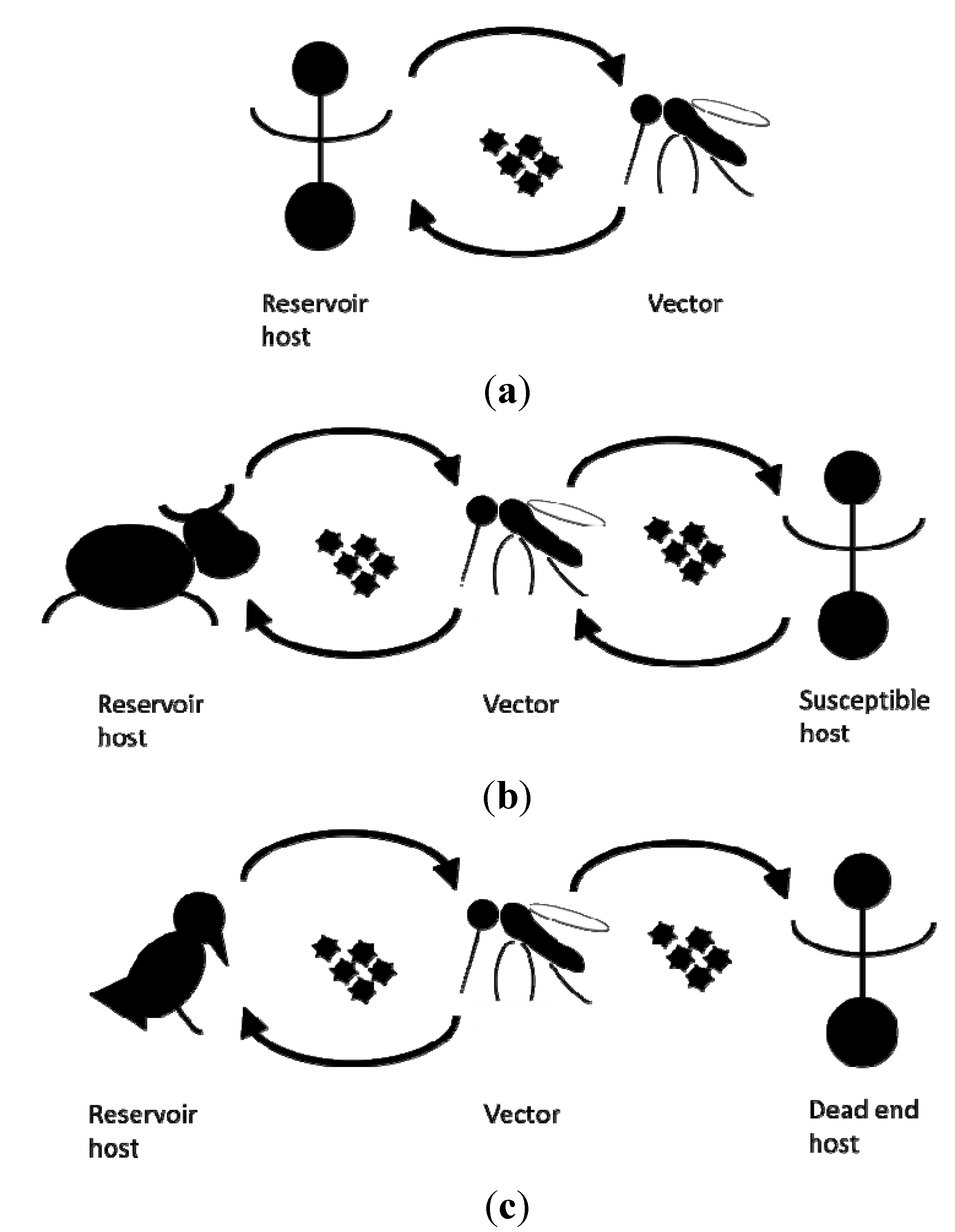{kind=link}
| Disease | Annual Global Cases 1 | Pathogen | Vector Genus | Infectious Period | Prophylaxis | Vaccine | Curative Medicine |
|---|---|---|---|---|---|---|---|
| Malaria | 451 million 2 | Plasmodium | Anopheles | Up to year 7 | √ | - | √ |
| Dengue | 96 million 3 | Flavivirus | Aedes | 3–5 days | - | - | - |
| Yellow fever | 200,000 4 | Flavivirus | Aedes | 3–5 days | - | √ | - |
| Japanese Encephalitis | 67,900 5 | Flavivirus | Culex | dead end host | - | √ | - |
| West Nile fever | 20, 000 6 | Flavivirus | Culex | dead end host | - | - 9 | - |
| Chikungunya | Epidemic | Alphavirus | Aedes | 6–7 days | - | - | - |
| Rift Valley fever | Epidemic | Phlebovirus | Culex/Aedes | short 8 | - | - | - |

2. Pitfalls and Lessons
2.1. Global Warming versus Global Change
2.2. Models versus Real World
2.3. Retrospective versus Prospective Studies
2.4. Generalized versus Contextual Approach
3. The Way Forward
| Context | Locally Acquired Human Case | Pathogen | Vector | Mosquito Borne Diseases in Western Europe |
|---|---|---|---|---|
| 1a | √ (every year) | √ | √ | No examples |
| 1b | √ (not every year) | √ | √ | No examples |
| 2 | - | √ | √ | Heartworm [57], Usutu [58], Ockelbo [59] |
| 3 | - | - | √ | West Nile virus [60], Malaria [61], Rift Valley Fever [62] |
| 4 | - | √ | - | Chikungunya; Dengue [63] |
| 5 | - | - | - | Japanese encephalitis * |
4. Conclusions
Acknowledgments
Conflicts of Interest
References
- Hay, S.I.; Okiro, E.A.; Gething, P.W.; Patil, A.P.; Tatem, A.J.; Guerra, C.A.; Snow, R.W. Estimating the global clinical burden of Plasmodium falciparum malaria in 2007. PLoS Med. 2010, 7. [Google Scholar] [CrossRef]
- Gething, P.W.; Patil, A.P.; Smith, D.L.; Guerra, C.A.; Elyazar, I.R.; Johnston, G.L.; Tatem, A.J.; Hay, S.I. A new world malaria map: Plasmodium falciparum endemicity in 2010. Malar. J. 2011, 10, 378. [Google Scholar] [CrossRef]
- Bhatt, S.; Gething, P.W.; Brady, O.J.; Messina, J.P.; Farlow, A.W.; Moyes, C.L.; Drake, J.M.; Brownstein, J.S.; Hoen, A.G.; Sankoh, O.; et al. The global distribution and burden of dengue. Nature 2013, 496, 504–507. [Google Scholar] [CrossRef]
- Campbell, G.L.; Hills, S.L.; Fischer, M.; Jacobson, J.A.; Hoke, C.H.; Hombach, J.M.; Marfin, A.A.; Solomon, T.; Tsai, T.F.; Tsu, V.D.; et al. Estimated global incidence of Japanese encephalitis: A systematic review. Bull. WHO 2011, 89, 766–774. [Google Scholar]
- Adalja, A.A. The Increasing Burden of West Nile Virus 2012. Available online: http://www.upmc-cbn.org/report_archive/2012/cbnreport_08242012.html (accessed 30 October 2013).
- Kasari, T.R.; Carr, D.A.; Lynn, T.V.; Weaver, J.T. Evaluation of pathways for release of Rift Valley fever virus into domestic ruminant livestock, ruminant wildlife, and human populations in the continental United States. Javma-J. Am. Vet. Med. Assoc. 2008, 232, 514–529. [Google Scholar] [CrossRef]
- Schaffner, F.; Bellini, R.; Petric, D.; Scholte, E.J.; Zeller, H.; Rakotoarivony, L.M. Development of guidelines for the surveillance of invasive mosquitoes in Europe. Parasit. Vectors 2013, 6, 209. [Google Scholar] [CrossRef]
- Scholte, E.; Den Hartog, W.; Dik, M.; Schoelitsz, B.; Brooks, M.; Schaffner, F.; Foussadier, R.; Braks, M.; Beeuwkes, J. Introduction and control of three invasive mosquito species in the Netherlands, July–October 2010. Euro Surveill. 2010, 15. Available online: http://www.eurosurveillance.org/ViewArticle.aspx?ArticleId=19710 (accessed on 11 December 2013).
- Sousa, C.A.; Clairouin, M.; Seixas, G.; Viveiros, B.; Novo, M.T.; Silva, A.C.; Escoval, M.T.; Economopoulou, A. Ongoing outbreak of dengue type 1 in the autonomous region of Madeira, Portugal: Preliminary report. Euro. Surveill. 2012, 17. Available online: http://www.eurosurveillance.org/ViewArticle.aspx?ArticleId=20333 (accessed on 11 December 2013).
- Weaver, S.C. Host range, amplification and arboviral disease emergence. Arch. Virol. Suppl. 2005, 19, 33–44. [Google Scholar]
- Garrett-Jones, C. Prognosis for interruption of malaria transmission through assessment of the mosquito's vectorial capacity. Nature 1964, 204, 1173–1175. [Google Scholar] [CrossRef]
- Reisen, W.K. Landscape epidemiology of vector-borne diseases. Annu. Rev. Entomol. 2010, 55, 461–483. [Google Scholar] [CrossRef]
- Renn, O.; Klinke, A.; van Asselt, M. Coping with complexity, uncertainty and ambiguity in risk governance: A synthesis. Ambio 2011, 40, 231–246. [Google Scholar] [CrossRef]
- Brownson, R.C.; Fielding, J.E.; Maylahn, C.M. Evidence-based public health: A fundamental concept for public health practice. Annu. Rev. Public Health 2009, 30, 175–201. [Google Scholar] [CrossRef]
- Kilpatrick, A.M.; Meola, M.A.; Moudy, R.M.; Kramer, L.D. Temperature, viral genetics, and the transmission of West Nile virus by Culex pipiens mosquitoes. PLoS Pathog. 2008, 4. [Google Scholar] [CrossRef]
- Veronesi, R.; Gentile, G.; Carrieri, M.; Maccagnani, B.; Stermieri, L.; Bellini, R. Seasonal pattern of daily activity of Aedes caspius, Aedes detritus, Culex modestus, and Culex pipiens in the Po Delta of northern Italy and significance for vector-borne disease risk assessment. J. Vector Ecol. 2012, 37, 49–61. [Google Scholar] [CrossRef]
- Bancroft, B.A.; Baker, N.J.; Blaustein, A.R. Effects of UVB radiation on marine and freshwater organisms: A synthesis through meta-analysis. Ecol. Lett. 2007, 10, 332–345. [Google Scholar] [CrossRef]
- Robinet, C.; Roques, A. Direct impacts of recent climate warming on insect populations. Integr. Zool. 2010, 5, 132–142. [Google Scholar] [CrossRef]
- Kingsolver, J.G.; Woods, H.A.; Buckley, L.B.; Potter, K.A.; MacLean, H.J.; Higgins, J.K. Complex life cycles and the responses of insects to climate change. Integr. Comp. Biol. 2011, 51, 719–732. [Google Scholar] [CrossRef]
- Stanke, C.; Kerac, M.; Prudhomme, C.; Medlock, J.; Murray, V. Health effects of drought: A systematic review of the evidence. PLoS Curr. 2013, 5. [Google Scholar] [CrossRef]
- Boggs, C.L.; Inouye, D.W. A single climate driver has direct and indirect effects on insect population dynamics. Ecol. Lett. 2012, 15, 502–508. [Google Scholar] [CrossRef]
- Rogers, D.J.; Suk, J.E.; Semenza, J.C. Using global maps to predict the risk of dengue in Europe. Acta Trop 2013. [Google Scholar] [CrossRef]
- Lounibos, L.P. Invasions by insect vectors of human disease. Annu. Rev. Entomol. 2002, 47, 233–266. [Google Scholar] [CrossRef]
- Tomlinson, C.J.; Chapman, L.; Thornes, J.E.; Baker, C.J. Including the urban heat island in spatial heat health risk assessment strategies: A case study for birmingham, UK. Int. J. Health Geogr. 2011, 10, 42. [Google Scholar] [CrossRef] [Green Version]
- Beebe, N.W.; Cooper, R.D.; Mottram, P.; Sweeney, A.W. Australia’s dengue risk driven by human adaptation to climate change. PLoS Negl. Trop. Dis. 2009, 3. [Google Scholar] [CrossRef]
- Reiter, P.; Lathrop, S.; Bunning, M.; Biggerstaff, B.; Singer, D.; Tiwari, T.; Baber, L.; Amador, M.; Thirion, J.; Hayes, J.; et al. Texas lifestyle limits transmission of dengue virus. Emerg. Infect. Dis. 2003, 9, 86–89. [Google Scholar] [CrossRef]
- Tabachnick, W.J. Challenges in predicting climate and environmental effects on vector-borne disease episystems in a changing world. J. Exp. Biol. 2010, 213, 946–954. [Google Scholar] [CrossRef]
- Gould, E.A.; Higgs, S. Impact of climate change and other factors on emerging arbovirus diseases. Trans. R. Soc. Trop. Med. Hyg. 2009, 103, 109–121. [Google Scholar] [CrossRef]
- Medlock, J.; Jameson, L. Ecological approaches to informing public health policy and risk assessments on emerging vector-borne zoonoses. Emerg. Health Threats J. 2010, 3. [Google Scholar] [CrossRef]
- Rogers, D.J.; Randolph, S.E. Climate change and vector-borne diseases. Adv. Parasitol. 2006, 62, 345–381. [Google Scholar] [CrossRef]
- Semenza, J.C.; Menne, B. Climate change and infectious diseases in Europe. Lancet Infect. Dis. 2009, 9, 365–375. [Google Scholar] [CrossRef]
- Vora, N. Impact of anthropogenic environmental alterations on vector-borne diseases. Medscape J. Med. 2008, 10. Available online: http://www.ncbi.nlm.nih.gov/pmc/articles/PMC2605134/?report=classic (accessed on 11 december 2013).
- Chaves, L.F.; Koenraadt, C.J.M. Climate change and highland malaria: Fresh air for a hot debate. Q. Rev. Biol. 2010, 85, 27–55. [Google Scholar] [CrossRef]
- Béguin, A.; Hales, S.; Rocklöv, J.; Åström, C.; Louis, V.; Sauerborn, R. The opposing effects of climate change and socio-economic development on the global distribution of malaria. Global Environ. Change 2011, 1209–1214. [Google Scholar]
- Hunter, P.R. Climate change and waterborne and vector-borne disease. J. Appl. Microbiol. 2003, 94, S37–S46. [Google Scholar] [CrossRef]
- Chaves, L.F.; Keogh, C.L.; Nguyen, A.M.; Decker, G.M.; Vazquez-Prokopec, G.M.; U.D., K. Combined sewage overflow accelerates immature development and increases body size in the urban mosquito Culex quinquefasciatus. J. Appl. Entomol. 2011, 135, 611–620. [Google Scholar] [CrossRef]
- Couret, J. Meta-analysis of factors affecting ontogenetic development rate in the Culex pipiens (diptera: Culicidae) complex. Environ. Entomol. 2013, 42, 614–626. [Google Scholar] [CrossRef]
- Reiner, R.C., Jr.; Perkins, T.A.; Barker, C.M.; Niu, T.; Chaves, L.F.; Ellis, A.M.; George, D.B.; Le Menach, A.; Pulliam, J.R.; Bisanzio, D.; et al. A systematic review of mathematical models of mosquito-borne pathogen transmission: 1970–2010. J. R. Soc. Interface 2013, 10. [Google Scholar] [CrossRef] [Green Version]
- Randolph, S.E.; Rogers, D.J. The arrival, establishment and spread of exotic diseases: Patterns and predictions. Nat. Rev. Microbiol. 2010, 8, 361–371. [Google Scholar] [CrossRef]
- Anyamba, A.; Chretien, J.P.; Small, J.; Tucker, C.J.; Formenty, P.B.; Richardson, J.H.; Britch, S.C.; Schnabel, D.C.; Erickson, R.L.; Linthicum, K.J. Prediction of a Rift Valley fever outbreak. Proc. Natl. Acad. Sci. USA 2009, 106, 955–959. [Google Scholar] [CrossRef]
- Poletti, P.; Messeri, G.; Ajelli, M.; Vallorani, R.; Rizzo, C.; Merler, S. Transmission potential of chikungunya virus and control measures: The case of Italy. PLoS One 2011, 6. [Google Scholar] [CrossRef]
- Hartemink, N. Vector-Borne Diseases: The Basic Reproduction Number R0 and Risk Maps. In Faculty of Veterinary Medicine; Utrecht University: Utrecht, The Netherland, 2009. [Google Scholar]
- Lindsay, S.W.; Hole, D.G.; Hutchinson, R.A.; Richards, S.A.; Willis, S.G. Assessing the future threat from vivax malaria in the United Kingdom using two markedly different modelling approaches. Malar. J. 2010, 9, 70. [Google Scholar] [CrossRef] [Green Version]
- Wonham, M.J.; Lewis, M.A.; Renclawowicz, J.; van den Driessche, P. Transmission assumptions generate conflicting predictions in host-vector disease models: A case study in West Nile virus. Ecol. Lett. 2006, 9, 706–725. [Google Scholar] [CrossRef]
- Taleb, N.N. The Black Swan: The Impact of the Highly Improbable, 1st ed.; Penguin Ltd.: London, England, 2007. [Google Scholar]
- Rogers, D.J.; Randolph, S.E. Studying the global distribution of infectious diseases using GIS and RS. Nat. Rev. Microbiol. 2003, 1, 231–237. [Google Scholar] [CrossRef]
- Ramasamy, R.; Surendran, S.N. Possible impact of rising sea levels on vector-borne infectious diseases. BMC Infect. Dis. 2011, 11, 18. [Google Scholar] [CrossRef]
- Dobson, M.J. Malaria in England: A geographical and historical perspective. Parassitologia 1994, 36, 35–60. [Google Scholar]
- Semenza, J.C.; Suk, J.E.; Estevez, V.; Ebi, K.L.; Lindgren, E. Mapping climate change vulnerabilities to infectious diseases in Europe. Environ. Health Perspect. 2012, 120, 385–392. [Google Scholar]
- Paz, S.; Semenza, J.C. Environmental drivers of West Nile fever epidemiology in Europe and Western Asia—A review. Int. J. Environ. Res. Public Health 2013, 10, 3543–3562. [Google Scholar] [CrossRef]
- Boxall, A.B.; Hardy, A.; Beulke, S.; Boucard, T.; Burgin, L.; Falloon, P.D.; Haygarth, P.M.; Hutchinson, T.; Kovats, R.S.; Leonardi, G.; et al. Impacts of climate change on indirect human exposure to pathogens and chemicals from agriculture. Environ. Health Perspect. 2009, 117, 508–514. [Google Scholar]
- Gubler, D.J.; Reiter, P.; Ebi, K.L.; Yap, W.; Nasci, R.; Patz, J.A. Climate variability and change in the United States: Potential impacts on vector- and rodent-borne diseases. Environ. Health Perspect. 2001, 109, S223–S233. [Google Scholar] [CrossRef]
- Mills, J.N.; Gage, K.L.; Khan, A.S. Potential influence of climate change on vector-borne and zoonotic diseases: A review and proposed research plan. Environ. Health Perspect. 2010, 118, 1507–1514. [Google Scholar] [CrossRef]
- Rosenthal, J. Climate change and the geographic distribution of infectious diseases. Ecohealth 2009, 6, 489–495. [Google Scholar] [CrossRef]
- Braks, M.; van der Giessen, J.; Kretzschmar, M.; van Pelt, W.; Scholte, E.J.; Reusken, C.; Zeller, H.; van Bortel, W.; Sprong, H. Towards an integrated approach in surveillance of vector-borne diseases in Europe. Parasit. Vectors 2011, 4, 192. [Google Scholar] [CrossRef]
- Hubalek, Z. Mosquito-borne viruses in Europe. Parasitol. Res. 2008, 103, S29–S43. [Google Scholar] [CrossRef]
- Morchon, R.; Carreton, E.; Gonzalez-Miguel, J.; Mellado-Hernandez, I. Heartworm disease (Dirofilaria immitis) and their vectors in Europe—New distribution trends. Front Physiol. 2012, 3, 196. [Google Scholar] [CrossRef]
- Becker, N.; Jost, H.; Ziegler, U.; Eiden, M.; Hoper, D.; Emmerich, P.; Fichet-Calvet, E.; Ehichioya, D.U.; Czajka, C.; Gabriel, M.; et al. Epizootic emergence of usutu virus in wild and captive birds in Germany. PLoS One 2012, 7. [Google Scholar] [CrossRef]
- Jost, H.; Bialonski, A.; Storch, V.; Gunther, S.; Becker, N.; Schmidt-Chanasit, J. Isolation and phylogenetic analysis of sindbis viruses from mosquitoes in Germany. J. Clin. Microbiol. 2010, 48, 1900–1903. [Google Scholar] [CrossRef]
- Lohmus, M.; Lindstrom, A.; Bjorklund, M. How often do they meet? Genetic similarity between European populations of a potential disease vector Culex pipiens. Infect. Ecol. Epidemiol. 2012, 2. [Google Scholar] [CrossRef]
- Dekoninck, W.; Hendrickx, F.; Vasn Bortel, W.; Versteirt, V.; Coosemans, M.; Damiens, D.; Hance, T.; de Clercq, E.M.; Hendrickx, G.; Schaffner, F.; et al. Human-induced expanded distribution of Anopheles plumbeus, experimental vector of West Nile virus and a potential vector of human malaria in Belgium. J. Med. Entomol. 2011, 48, 924–928. [Google Scholar] [CrossRef]
- Chevalier, V.; Pepin, M.; Plee, L.; Lancelot, R. Rift valley fever—A threat for Europe? Euro Surveill. 2010, 15. Available online: http://www.eurosurveillance.org/ViewArticle.aspx?ArticleId=19506 (accessed on 11 December 2013).
- Wieten, R.W.; Vlietstra, W.; Goorhuis, A.; van Vugt, M.; Hodiamont, C.J.; Leenstra, T.; de Vries, P.J.; Janssen, S.; van Thiel, P.P.; Stijnis, K.; et al. Dengue in travellers: Applicability of the 1975–1997 and the 2009 WHO classification system of dengue fever. Trop. Med. Int. Health 2012, 17, 1023–1030. [Google Scholar] [CrossRef]
- Ravanini, P.; Huhtamo, E.; Ilaria, V.; Crobu, M.; Nicosia, A.; Servino, L.; Rivasi, F.; Allegrini, S.; Miglio, U.; Magri, A.; et al. Japanese encephalitis virus RNA detected in Culex pipiens mosquitoes in Italy. Euro Surveill. 2012, 17. Available online: http://www.eurosurveillance.org/ViewArticle.aspx?ArticleId=20221 (accessed on 11 December 2013).
- Medlock, J.M.; Hansford, K.M.; Schaffner, F.; Versteirt, V.; Hendrickx, G.; Zeller, H.; Bortel, W.V. A review of the invasive mosquitoes in Europe: Ecology, public health risks, and control options. Vector-Borne Zoonotic Dis. 2012. [Google Scholar] [CrossRef] [Green Version]
- Reiter, P. Climate change and mosquito-borne disease: Knowing the horse before hitching the cart. Rev. Sci. Tech. 2008, 27, 383–398. [Google Scholar]
- Randolph, S.E. To what extent has climate change contributed to the recent epidemiology of tick-borne diseases? Vet. Parasitol. 2010, 167, 92–94. [Google Scholar] [CrossRef]
- Morse, S.S. Factors in the emergence of infectious diseases. Emerg. Infect. Dis. 1995, 1, 7–15. [Google Scholar] [CrossRef]
- Morens, D.M.; Folkers, G.K.; Fauci, A.S. The challenge of emerging and re-emerging infectious diseases. Nature 2004, 430, 242–249. [Google Scholar] [CrossRef]
- Jones, K.E.; Patel, N.G.; Levy, M.A.; Storeygard, A.; Balk, D.; Gittleman, J.L.; Daszak, P. Global trends in emerging infectious diseases. Nature 2008, 451, 990–993. [Google Scholar] [CrossRef]
- Pfeffer, M.; Dobler, G. Emergence of zoonotic arboviruses by animal trade and migration. Parasit. Vectors 2010, 3, 35. [Google Scholar] [CrossRef]
- Weiss, R.A.; McMichael, A.J. Social and environmental risk factors in the emergence of infectious diseases. Nat. Med. 2004, 10, S70–S76. [Google Scholar] [CrossRef]
- Calistri, P.; Giovannini, A.; Hubalek, Z.; Ionescu, A.; Monaco, F.; Savini, G.; Lelli, R. Epidemiology of West Nile in Europe and in the Mediterranean basin. Open Virol. J. 2010, 4, 29–37. [Google Scholar]
- Gratz, N. Vector- and Rodent-borne Diseases in Europe and North America: Distribution, Public Health Burden, and Control; Gratz, N., Ed.; Cambridge University: Cambridge, UK, 2006; p. 410. [Google Scholar]
- Rezza, G.; Nicoletti, L.; Angelini, R.; Romi, R.; Finarelli, A.C.; Panning, M.; Cordioli, P.; Fortuna, C.; Boros, S.; Magurano, F.; et al. Infection with chikungunya virus in Italy: An outbreak in a temperate region. Lancet 2007, 370, 1840–1846. [Google Scholar] [CrossRef]
- Danis, K.; Baka, A.; Lenglet, A.; Van Bortel, W.; Terzaki, I.; Tseroni, M.; Detsis, M.; Papanikolaou, E.; Balaska, A.; Gewehr, S.; et al. Autochthonous Plasmodium vivax malaria in Greece, 2011. Euro Surveill. 2011, 16. Available online: http://www.eurosurveillance.org/ViewArticle.aspx?ArticleId=19993 (accessed on 11 December 2013).
- Vazquez, A.; Jimenez-Clavero, M.; Franco, L.; Donoso-Mantke, O.; Sambri, V.; Niedrig, M.; Zeller, H.; Tenorio, A. Usutu virus: Potential risk of human disease in Europe. Euro Surveill. 2011, 16. Available online: http://www.eurosurveillance.org/ViewArticle.aspx?ArticleId=19935 (accessed on 11 December 2013).
- Tsetsarkin, K.A.; Vanlandingham, D.L.; McGee, C.E.; Higgs, S. A single mutation in chikungunya virus affects vector specificity and epidemic potential. PLoS Pathog. 2007, 3. [Google Scholar] [CrossRef]
- Cliff, A.; Haggett, P. Time, travel and infection. Br. Med. Bull. 2004, 69, 87–99. [Google Scholar] [CrossRef]
- Poncon, N.; Tran, A.; Toty, C.; Luty, A.J.; Fontenille, D. A quantitative risk assessment approach for mosquito-borne diseases: Malaria re-emergence in southern France. Malar. J. 2008, 7, 147. [Google Scholar] [CrossRef]
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Braks, M.; Van Ginkel, R.; Wint, W.; Sedda, L.; Sprong, H. Climate Change and Public Health Policy: Translating the Science. Int. J. Environ. Res. Public Health 2014, 11, 13-29. https://doi.org/10.3390/ijerph110100013
Braks M, Van Ginkel R, Wint W, Sedda L, Sprong H. Climate Change and Public Health Policy: Translating the Science. International Journal of Environmental Research and Public Health. 2014; 11(1):13-29. https://doi.org/10.3390/ijerph110100013
Chicago/Turabian StyleBraks, Marieta, Rijk Van Ginkel, William Wint, Luigi Sedda, and Hein Sprong. 2014. "Climate Change and Public Health Policy: Translating the Science" International Journal of Environmental Research and Public Health 11, no. 1: 13-29. https://doi.org/10.3390/ijerph110100013