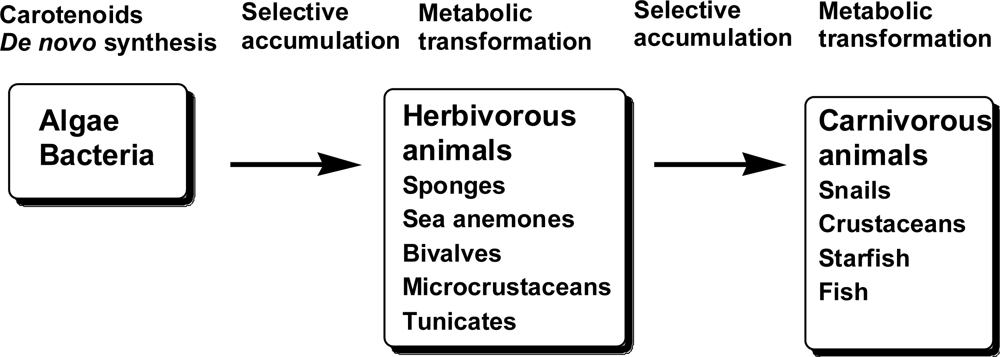
Carotenoids in Marine Animals

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Porifera (Marine Sponges)
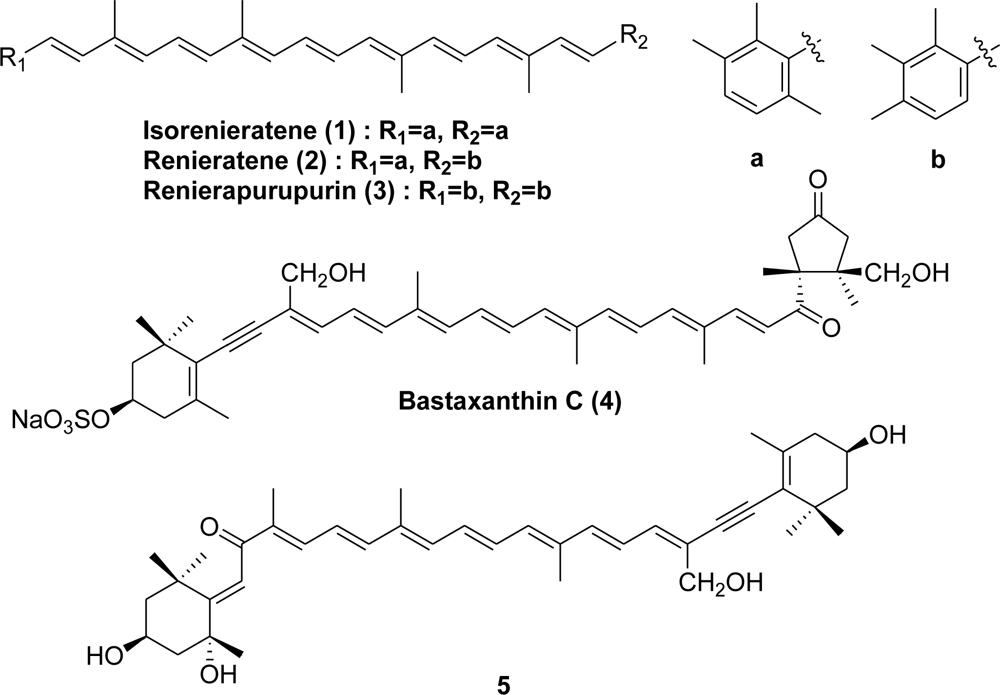
3. Coelenterata (Sea Anemones)
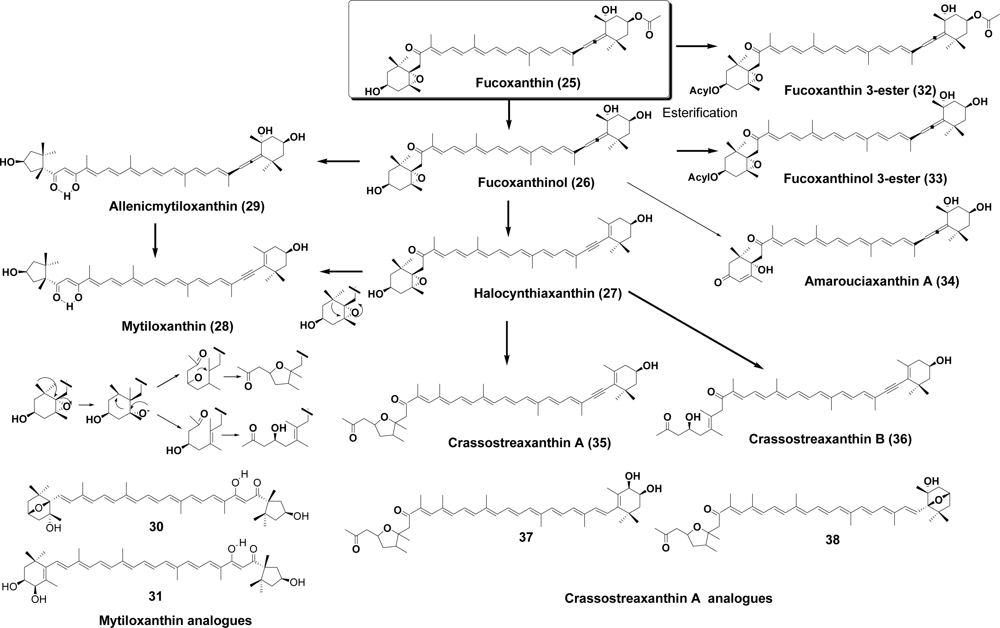
4. Mollusca (Mollusks)
5. Arthropoda (Crustaceans)
6. Echinodermata (Echinoderms)
7. Protochordata (Tunicates)
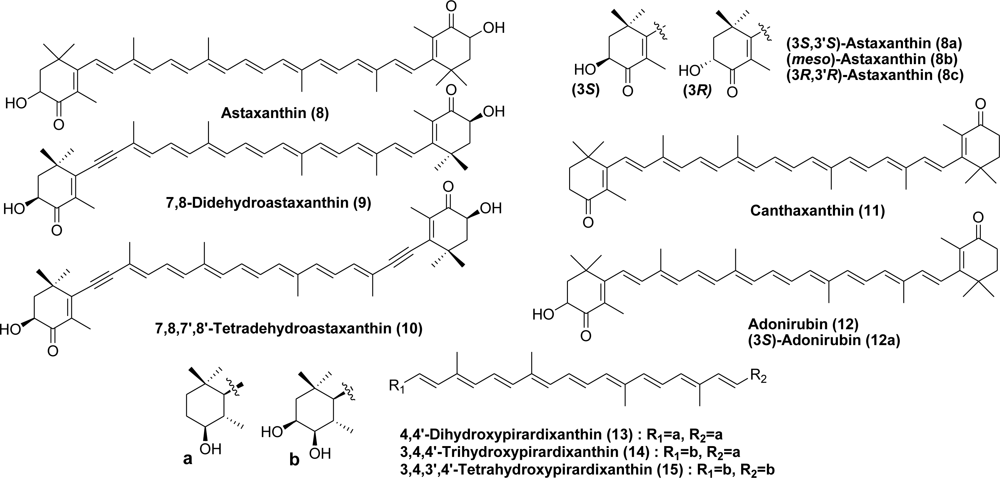
8. Pisces (Fish)
9. Mammalia (Mammals)
10. Role of Carotenoids in Marine Animals and Utilization of Carotenoids for Aquaculture
11. Conclusions
References and Notes
- Britton, G; Liaaen-Jensen, S; Pfander, H. Carotenoids Hand Book; Birkhäuser: Basel, Switzerland, 2004. [Google Scholar]
- Maoka, T; Akimoto, N. Natural product chemistry in carotenoid, some experimental techniques for structural elucidation and analysis of natural carotenoids. Carotenoid Sci 2008, 13, 10–17. [Google Scholar]
- Maoka, T. Recent progress in structural studies of carotenoids in animals and plants. Arch Biochem Biophys 2009, 483, 191–195. [Google Scholar]
- Maoka, T. Structural studies of natural carotenoids by our research group during the three decade. Carotenoid Sci 2009, 14, 26–36. [Google Scholar]
- Liaaen-Jensen, S. Marine carotenoids—Selected topics. New J Chem 1990, 14, 747–759. [Google Scholar]
- Liaaen-Jensen, S. Carotenoids in Food Chain. In Carotenoids: Biosynthesis and Metabolism; Britton, G, Liaaen-Jensen, S, Pfander, H, Eds.; Birkhäuser: Basel, Switzerland, 1998; Volume 3; pp. 359–371. [Google Scholar]
- Matsuno, T. Aquatic animal carotenoids. Fish Sci 2001, 67, 771–789. [Google Scholar]
- Matsuno, T. Animal carotenoids. In Carotenoids Chemistry and Biology; Krinsky, NI, Mathews-Roth, MM, Taylor, RF, Eds.; Plenum Press: New York, NY, USA, 1989; pp. 59–74. [Google Scholar]
- Matsuno, T; Hirao, S. Marine carotenoids. In Marine Biogenic Lipids, Fats, and Oils; Ackman, RG, Ed.; CRC Press: Boca Raton, FL, USA, 1989; Volume 1; pp. 251–388. [Google Scholar]
- Rogers, EW; Molinski, TF. A cytotoxic carotenoid from the marine sponge Prianos osiro. J Nat Prod 2005, 68, 450–452, The IUPAC name (3R,3′R,5S)-3,3′,5,19′-tetrahydroxy-7′,8′-didehydro-γ,ɛ-carotene-8-one was given for compound 5 in this literature. However, this IUPAC name is un-correct. Correct IUPAC name for 5 is (3R,3′R,5S)-3,3′,5,19′-tetrahydroxy-7′,8′-didehydro-5,8-dihydro-β,β-caroten-8-one.. [Google Scholar]
- Maoka, T; Akimoto, N; Tsushima, M; Komemushi, S; Harada, R; Sameshima, N; Iwase, F; Sakagami, Y. Carotenoids in corals and sea snails. Carotenoid Sci 2010. submitted for publication. [Google Scholar]
- Tsushima, M; Maoka, T; Matsuno, T. Comparative biochemical studies of carotenoids in marine invertebrates. The first positive identification of ɛ,ɛ-carotene derivatives and isolation of two new carotenoids from chitons. Comp Biochem Physiol 1989, 93B, 665–671. [Google Scholar]
- Tsushima, M; Maoka, T; Matsuno, T. Structure of carotenoids with 5,6-dihydro-β-end group from the spindle shell Fusinus perplexus. J Nat Prod 2001, 64, 1139–1142. [Google Scholar]
- Yamashita, E; Matsuno, T. A new apocarotenoid from the sea hare Aplysia kurodai. Comp Biochem Physiol 1990, 96B, 465–470. [Google Scholar]
- Maoka, T. A new apocarotenoid from marine shellfish. J Nat Prod 1997, 60, 616–617. [Google Scholar]
- Maoka, T; Hashimoto, K; Akimoto, N; Fujiwara, Y. Structures of five new carotenoids from the oyster Crassostrea gigas. J Nat Prod 2001, 64, 578–581. [Google Scholar]
- Maoka, T; Fujiwara, Y; Hashimoto, K; Akimoto, N. Structures of new carotenoids with a 3,4-dihydroxy-β-end group from the oyster Crassostrea gigas. Chem Pharm Bull 2005, 53, 1207–1209. [Google Scholar]
- Maoka, T; Fujiwara, Y; Hashimoto, K; Akimoto, N. Characterizatoin of fucoxanthin and fucoxanthinol esters in the chinese surf clam Mactra chinensis. J Agric Food Chem 2007, 55, 1563–1567. [Google Scholar]
- Maoka, T; Akimoto, N; Murakoshi, M; Sugiyama, K; Nishino, H. Carotenoids in clams, Ruditapes philippinarum and Meretrix petechialis. J Agric Food Chem 2010, 58, 5784–5788. [Google Scholar]
- Maoka, T; Akimoto, N; Yim, M-J; Hosokawa, M; Miyashita, K. A new C37-skeletal carotenoid from the clam Paphia amabillis. J Agric Food Chem 2008, 56, 12069–12072. [Google Scholar]
- Fujiwara, Y; Maoka, T; Ookubo, M; Matsuno, T. Crassostreaxanthins A and B: novel marine carotenoids from the oyster Crassostrea gigas. Tetrahedron Lett 1992, 33, 4941–4944. [Google Scholar]
- Tode, C; Yamano, Y; Ito, M. First total synthesis of crassostreaxanthin B. J. Chem. Soc., Perkin Trans 1999, 1, 1625–1626. [Google Scholar] [CrossRef]
- Tode, C; Yamano, Y; Ito, M. Carotenoids and related polyenes. Part 7. Total synthesis of crassostreaxanthin B Applying the stereoselective rearrangement of tetrasubstituted epoxides. J Chem Soc, Perkin Trans 2001, 1, 3338–3345. [Google Scholar] [CrossRef]
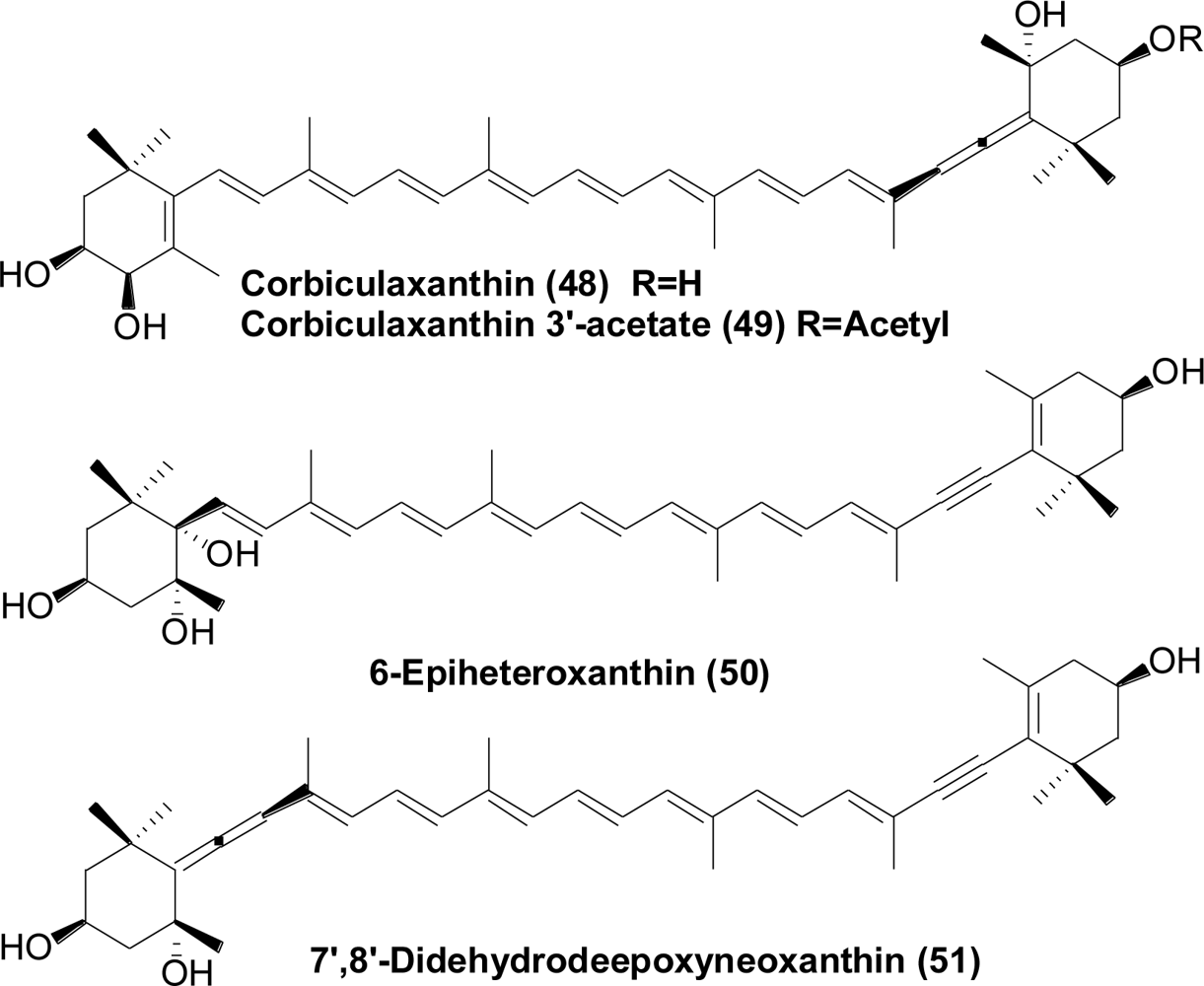
- Maoka, T; Fujiwara, Y; Hashimoto, K; Akimoto, N. Carotenoids in three species of corbicula clams, Corbicula japonica, Corbicula sandai, and Corbicula sp. (Chinese freshwater corbicula clam). J Agric Food Chem 2005, 53, 8357–8364. [Google Scholar]
- Maoka, T; Fujiwara, Y; Hashimoto, K; Akimoto, N. Structure of new carotenoids from the corbicula clam Corbicula japonic. J Nat Prod 2005, 68, 1341–1344. [Google Scholar]
- Maoka, T; Yokoi, S; Matsuno, T. Comparative biochemical stidies of carotenoids in nine species of Cephalopoda. Comp Biochem Physiol 1989, 92B, 247–250. [Google Scholar]
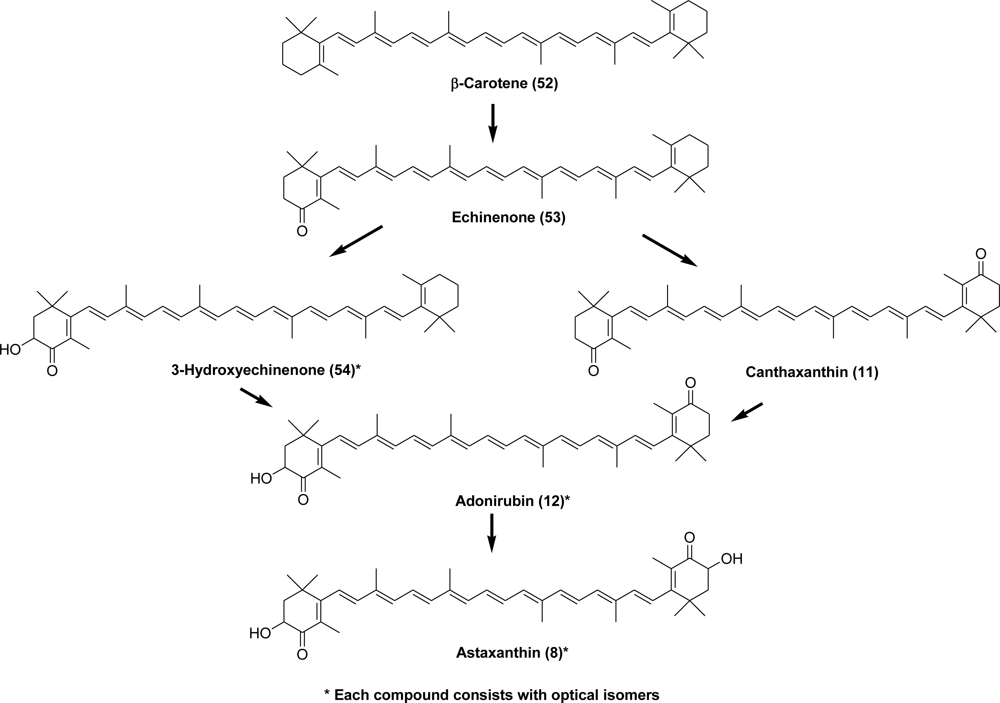
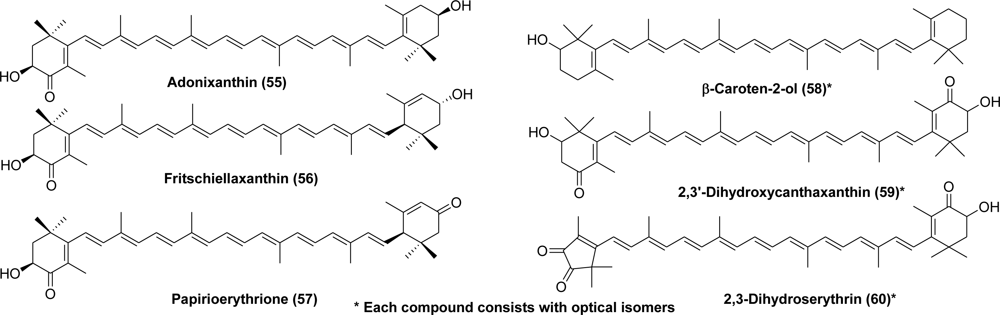
- Maoka, T; Akimoto, N. 2,3′-Dihydroxycanthaxanthin, a new carotenoid with a 2-hydroxy-4-oxo-β-end group from the hermit Crab Paralithodes brevipes. Chem Pharm Bull 2006, 54, 1462–1464. [Google Scholar]
- Maoka, T; Ando, S. Isolation of purple nor-carotenoid, 2,3-dihydroserythrin, from crawfish Procambarus clarkii. Fish Sci 2007, 73, 967–968. [Google Scholar]
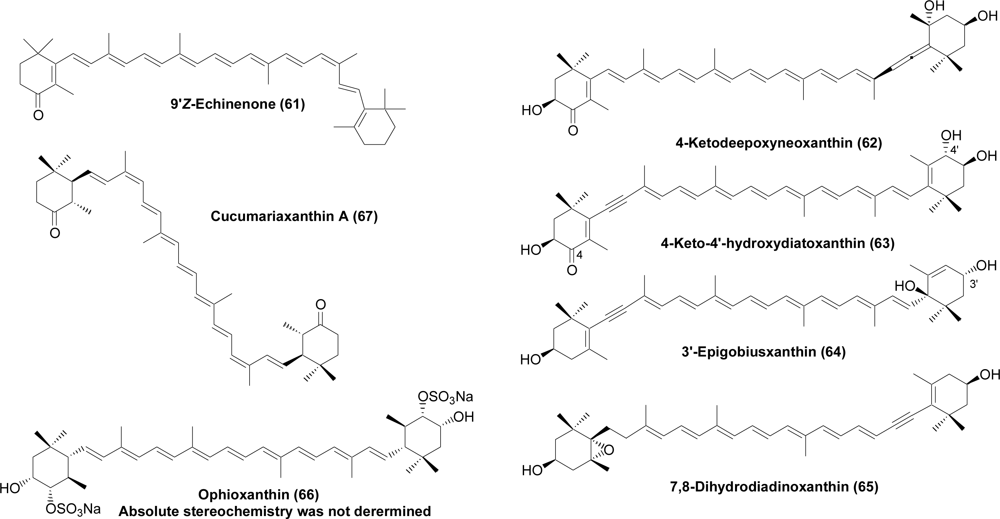
- Tsushima, M; Matsuno, T. Occurrence of 9′Z-β-echinenone of the sea urchin Pseudocentrotus depressus. Comp Biochem Physiol 1997, 118B, 921–925. [Google Scholar]
- Maoka, T; Akimoto, N; Terada, Y; Komemushi, S; Harada, R; Sameshima, N; Sakagami, Y. Structure of minor carotenoids from crown-of-thorns starfish Acanthaster planci. J Nat Prod 2010, 73, 675–678. [Google Scholar]
- D’auria, MV; Riccio, R; Minale, L. Ophioxanthin, a new marine carotenoid sulphate from the ophiuroid Ophioderma longicaudum. Tetrahedron Lett 1985, 26, 1871–1872. [Google Scholar]
- Tsushima, M; Fujiwara, Y; Matsuno, T. Novel marine di-Z-carotenoids, cucumariaxanthins A, B and C from the sea cucumber Cucumaria japonica. J Nat Prod 1996, 59, 30–34. [Google Scholar]
- Ferreres, F; Pereira, DM; Gil-Izquierdo, A; Valentão, P; Botelho, J; Mouga, T; Andrade, PB. HPLC-PAD-atmospheric pressure chemical ionization-MS metabolite profiling of cytotoxic carotenoids from the echinoderm Marthasterias glacialis (spiny sea-star). J Sep Sci 2010, 33, 2250–2257. [Google Scholar]
- Matsuno, T; Ookubo, M; Nishizawa, T; Shimizu, I. Carotenoids of sea suirts. I. New marine carotenids, halocynthiaxanthin and mytiloxanthinone from Halocynthia roretzi. Chem Pharm Bull 1984, 32, 4309–4315. [Google Scholar]
- Matsuno, T; Ookubo, M; Komori, T. Carotenoids of tunicates. III: The structural elucidation of two new marine carotenoids, amarouciaxanthin A and B. J Nat Prod 1985, 48, 606–613. [Google Scholar]
- Schiedt, K. Absorption and metabolism of carotenoids in birds, fish and crustaceans. In Carotenoids Biosynthesis and Metabolism; Britton, G, Liaaen-Jensen, S, Pfander, H, Eds.; Birkhäuser: Basel, Switzerland, 1998; Volume 3; pp. 285–358. [Google Scholar]
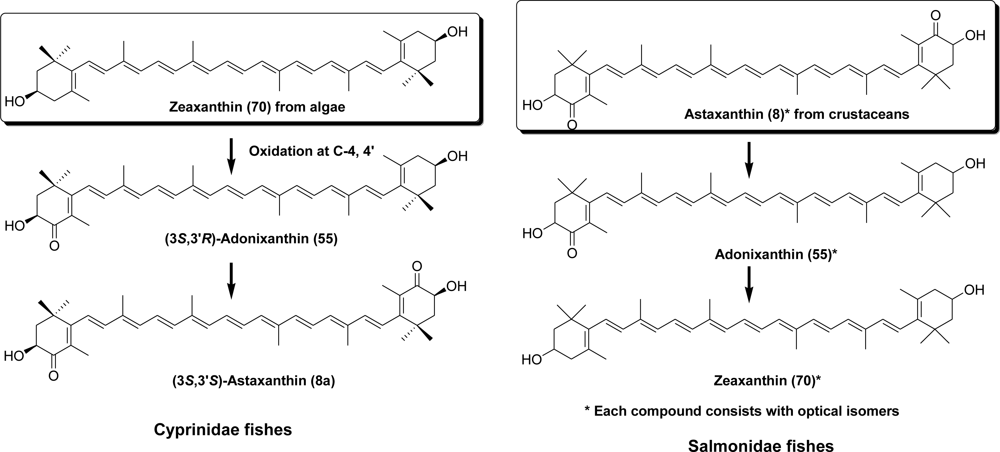
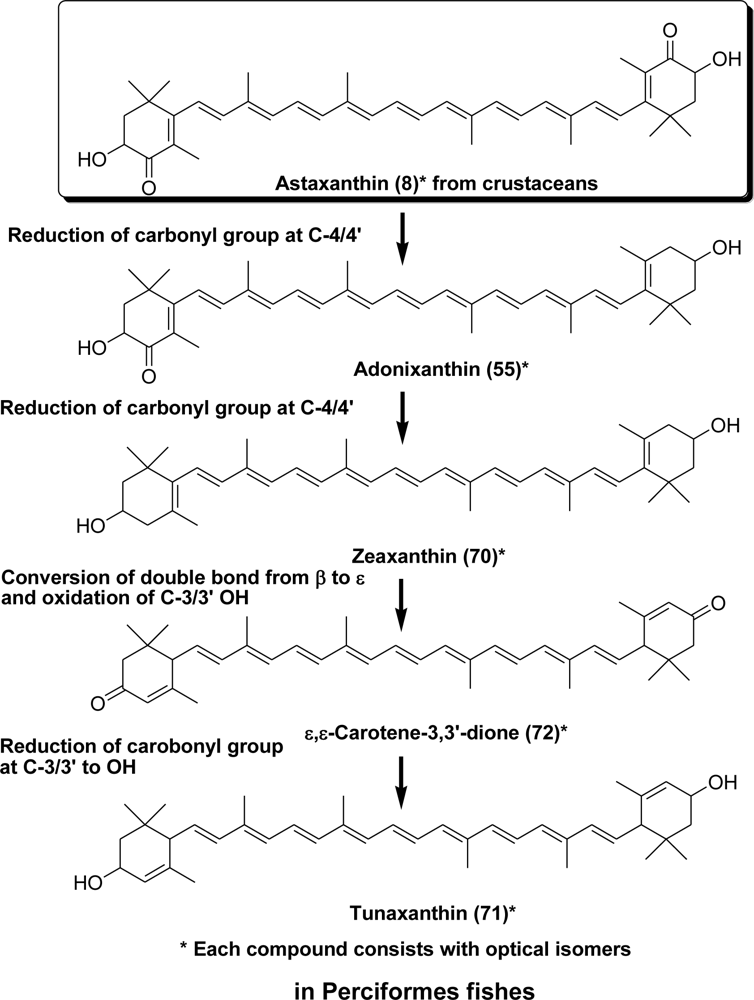
- Matsuno, T; Katsuyama, M; Maoka, T; Hirono, T; Komori, T. Reductive metabolic pathways of carotenoids in fish (3S,3′S)-astaxanthin to tunaxanthin A, B and C. Comp Biochem Physiol 1985, 80B, 779–789. [Google Scholar]
- Maoka, T; Arai, A; Shimizu, M; Matsuno, T. The first isolation of enantiomeric and meso-zeaxanthin in nature. Comp Biochem Physiol 1986, 83B, 121–124. [Google Scholar]
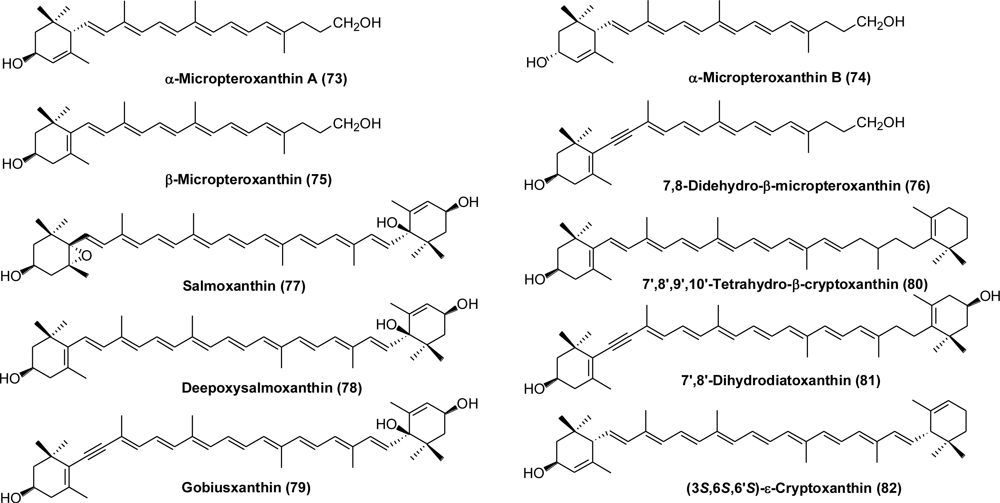
- Yamashita, E; Arai, S; Matsuno, T. Metabolism of xanthophylls to vitamin A and new apocarotenoids in liver and skin of black bass Micropterus salmoides. Comp Biochem Physiol 1996, 113B, 485–489. [Google Scholar]
- Matsuno, T; Tsushima, M; Maoka, T. Salmoxanthin, deepoxy-salmoxanthin and 7,8-didehydrodeepoxy-salmoxanthin from the salmon Oncorhynchus keta. J Nat Prod 2001, 64, 507–510. [Google Scholar]
- Tsushima, M; Mune, E; Maoka, T; Matsuno, T. Isolation of stereoisomeric epoxy carotenoids and new acetylenic carotenoid from the common freshwater goby Rhinogobius brunneus. J Nat Prod 2000, 63, 960–964. [Google Scholar]
- Tsushima, M; Ikuno, Y; Nagata, S; Kodama, K; Matsuno, T. Comparative biochemical studies of carotenoids in catfishes. Comp Biochem Physiol 2002, 133B, 331–336. [Google Scholar]
- Maoka, T; Akiomoto, N. Structures of minor carotenoids from the Japanese common catfish Silurus asotus. Chem Phram Bull 2011, 59, 140–145. [Google Scholar]
- Slifka, KA; Bowen, PE; Stacewicz-Sapuntzakis, M; Susan, D; Crissey, SD. A survey of serum and dietary carotenoids in captive wild animals. J Nutr 1999, 129, 380–390. [Google Scholar]
- Asai, A; Sugawara, H; Ono, H; Nagao, A. Biotransformation of fucoxanthinol into amarouciaxanthin A in mice and HepG2 cells: formation and cytotoxicity of fucoxanthin metabolites. Drug Metab Dispos 2004, 32, 205–211. [Google Scholar]
- Hashimoto, T; Ozaki, Y; Taminato, M; Das, SK; Mizuno, M; Yoshimura, K; Maoka, T; Kanazawa, K. The distribution and accumulation of fucoxanthin and its metabolites after oral administration in mice. Br J Nutr 2009, 102, 242–248. [Google Scholar]
- Matsuno, T. Xanthophylls as precursor of retinoids. Pure Appl. Chem 1991, 63, 81–88. [Google Scholar]
- Nakano, T; Tosa, M; Takeuchi, M. Improvement of biochemical features in fish health by red yeast and synthetic astaxanthin. J Agric Food Chem 1995, 43, 1570–1573. [Google Scholar]
- Nakano, T; Kanmuri, T; Sato, M; Takeuchi, M. Effect of astaxanthin rich red yeast (Phaffia rhodozyma) on oxidative stress in rainbow trout. Biochim Biophys Acta 1999, 1426, 119–125. [Google Scholar]
- Torrissen, OJ; Christiansen, R. Requirements for carotenoids in fish diets. J Appl Ichthyol 1995, 11, 225–230. [Google Scholar]
- Tsushima, M; Kawakami, T; Mine, M; Matsuno, T. The role of carotenoids in the development of the sea urchin Pseudocentrotus depressus. Invert Reprod Develop 1997, 32, 149–153. [Google Scholar]
- Thompson, I; Choubert, GD; Houlihan, F; Secombes, CJ. The effect of dietary Vitamin A and Astaxanthin on the immunocompetence of rainbow trout. Aquaculture 1995, 133, 91–102. [Google Scholar]
- Kawakami, T; Tsushima, M; Katabami, Y; Mine, M; Ishida, A; Matsuno, T. Effect of β,β-carotene, β-echinenone, astaxanthin, fucoxanthin, vitamin A and vitamin E on the biological defense of the sea urchin Pseudocentrotus depressus. J Exp Mar Biol Ecol 1998, 226, 165–174. [Google Scholar]



© 2011 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Maoka, T. Carotenoids in Marine Animals. Mar. Drugs 2011, 9, 278-293. https://doi.org/10.3390/md9020278
Maoka T. Carotenoids in Marine Animals. Marine Drugs. 2011; 9(2):278-293. https://doi.org/10.3390/md9020278
Chicago/Turabian StyleMaoka, Takashi. 2011. "Carotenoids in Marine Animals" Marine Drugs 9, no. 2: 278-293. https://doi.org/10.3390/md9020278