Inducible ASABF-Type Antimicrobial Peptide from the Sponge Suberites domuncula: Microbicidal and Hemolytic Activity in Vitro and Toxic Effect on Molluscs in Vivo †

 ,
,
Abstract
:1. Introduction
2. Results and Discussion
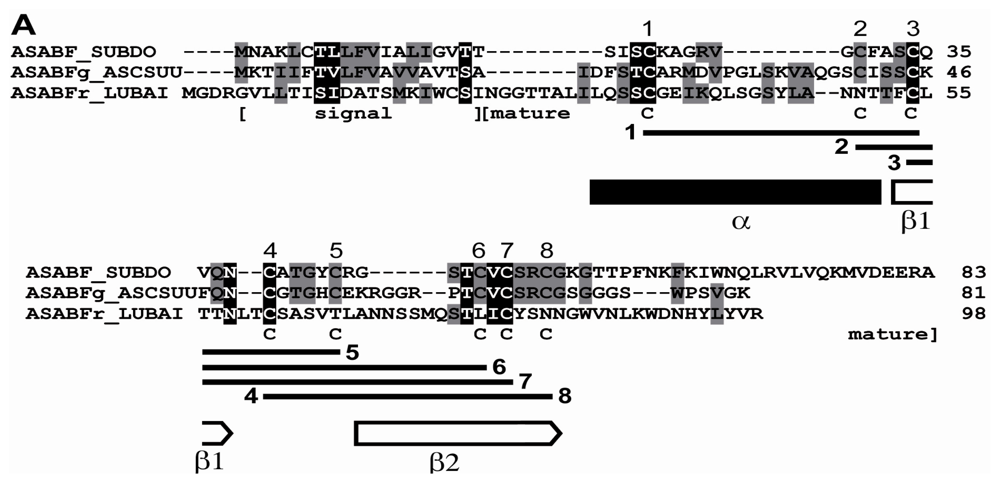
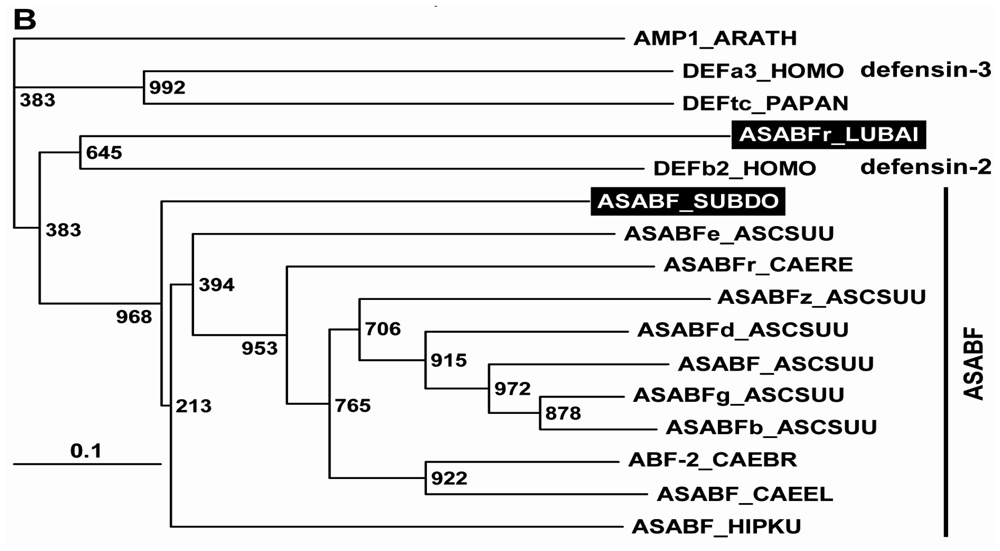
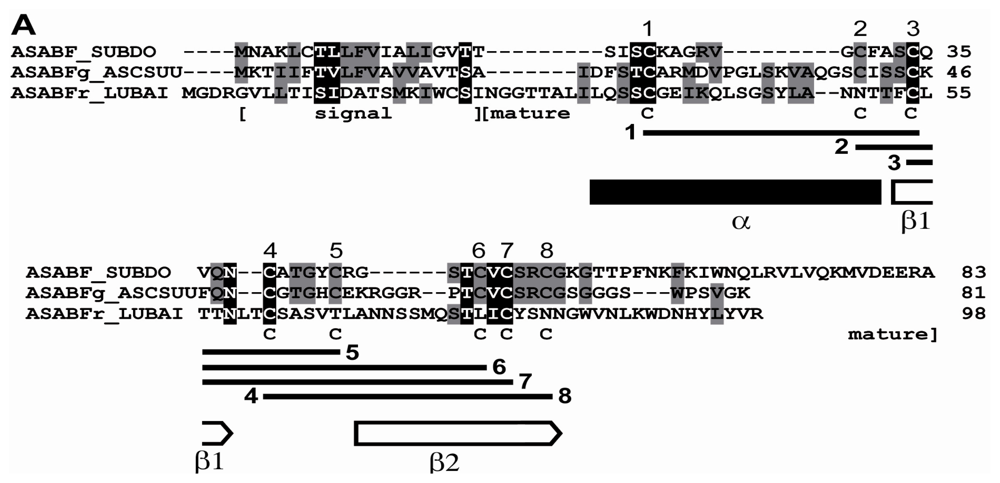
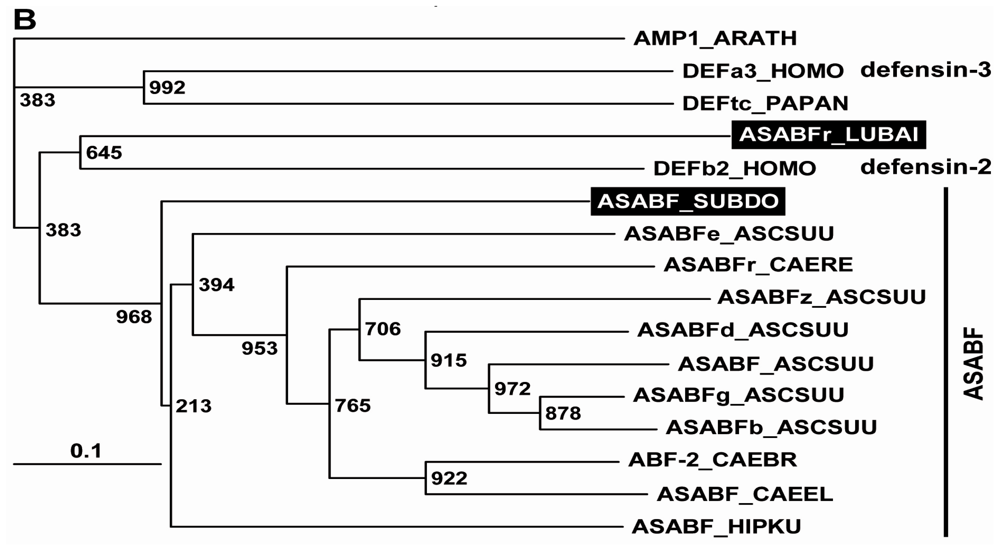
2.1. Sponge (S. domuncula, L. baicalensis) ASABF Peptide: Sequence Analysis and Phylogenetic Relationships
2.2. Preparation of Recombinant ASABF
2.3. Microbicidal Assay
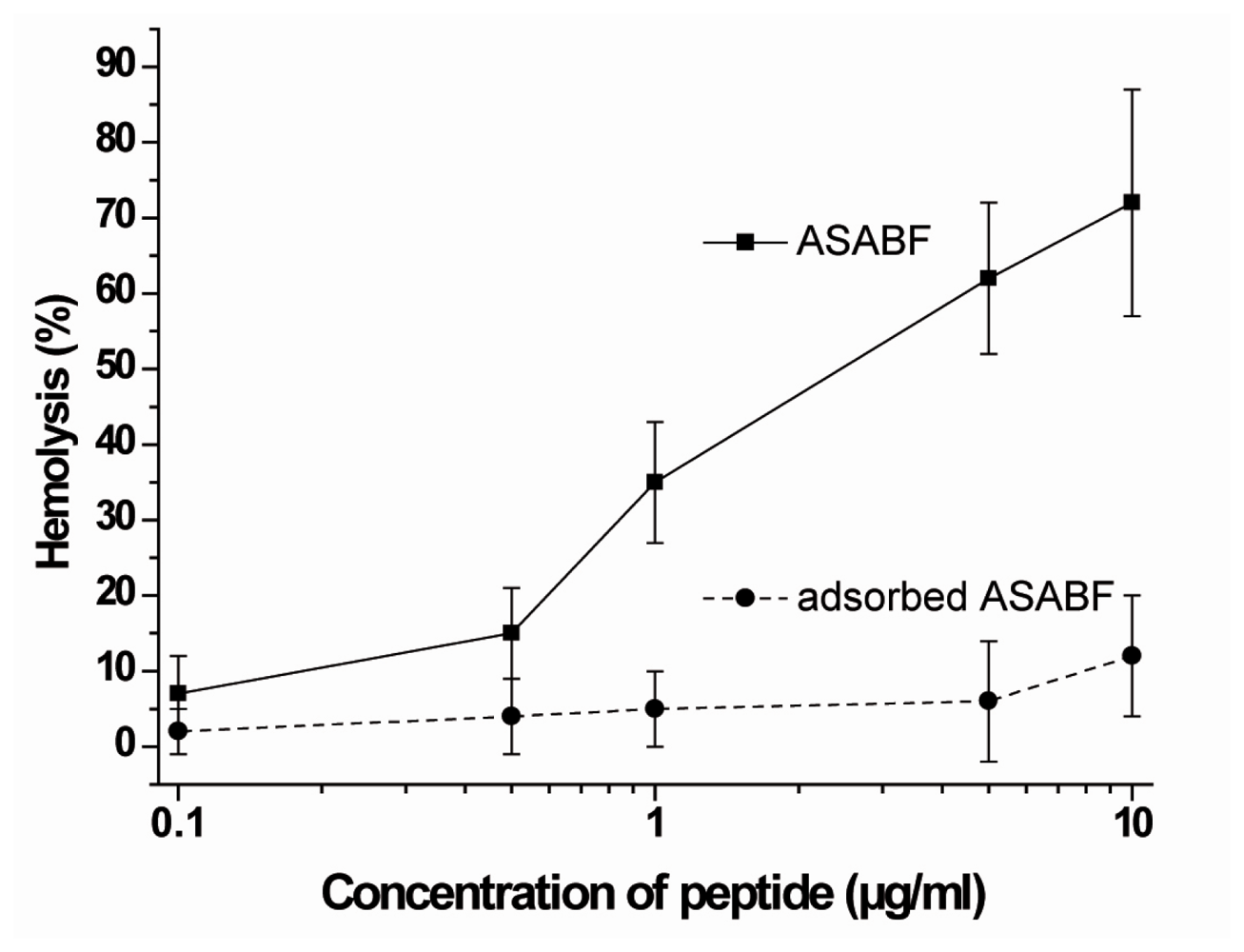
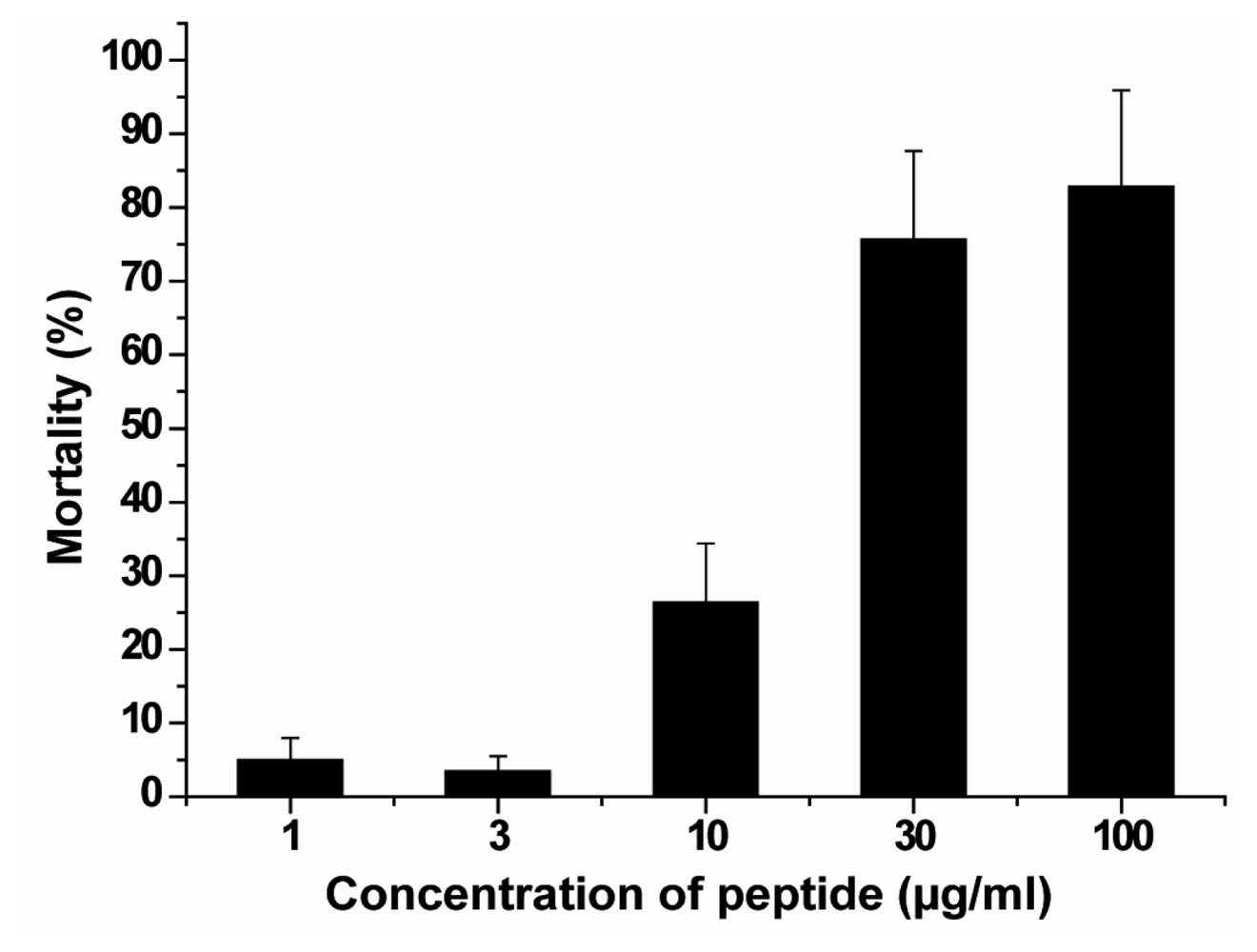
2.4. Hemolytic Activity
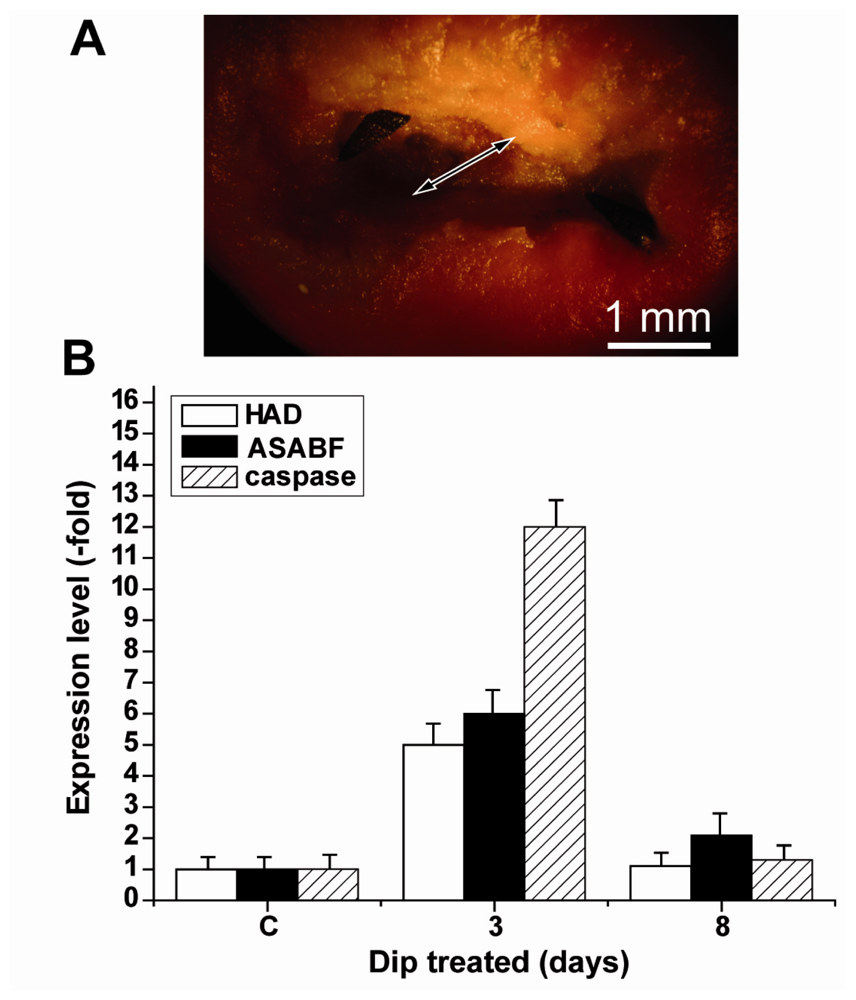
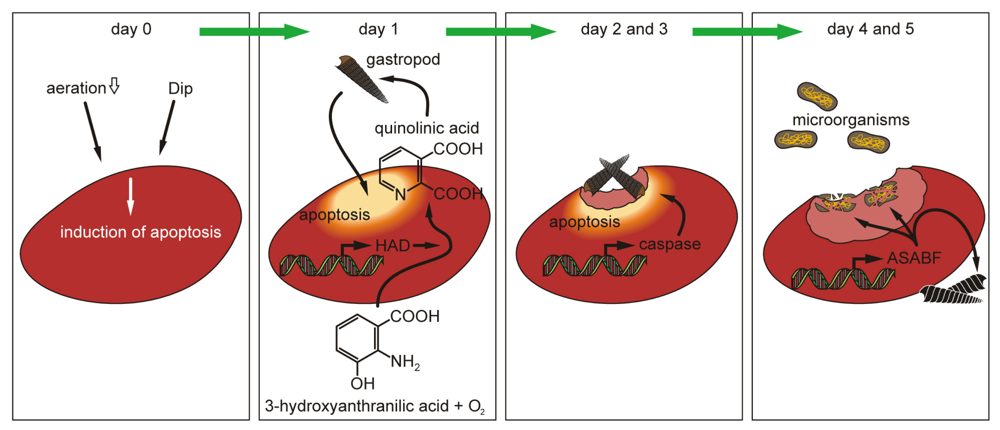
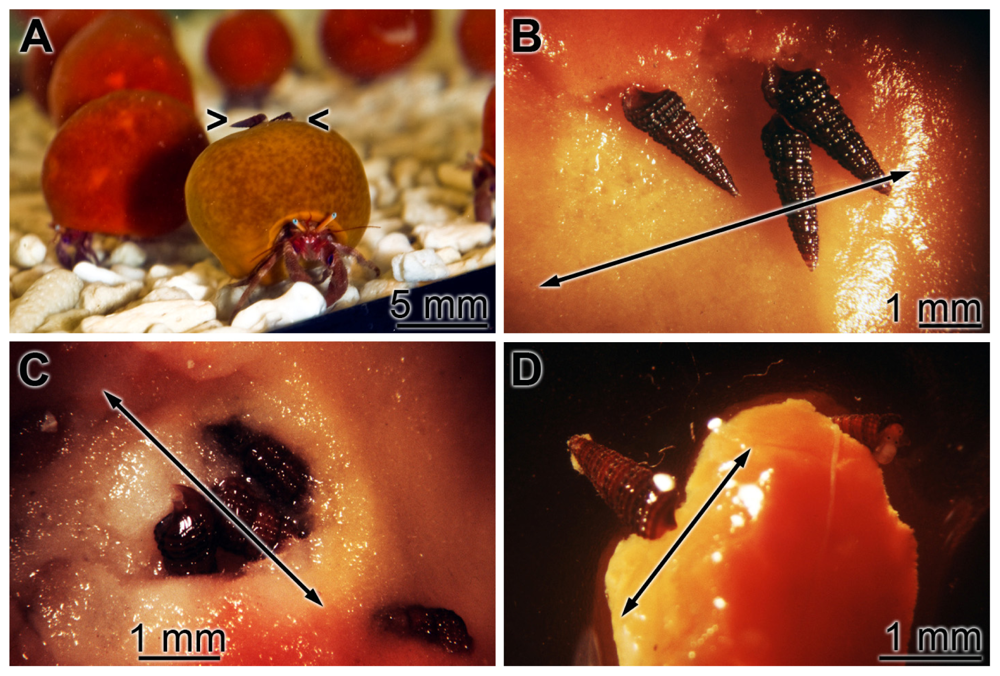
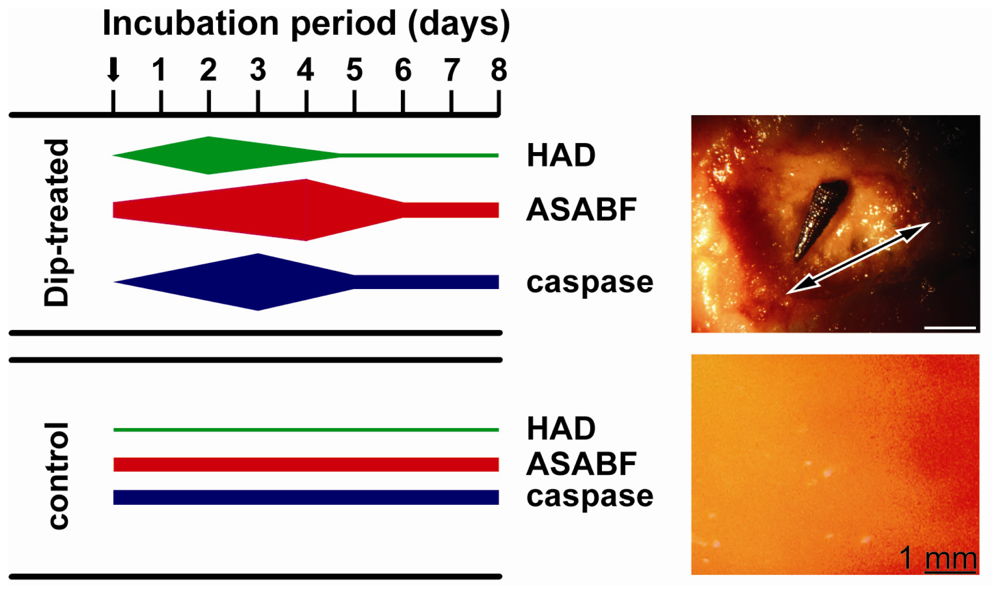
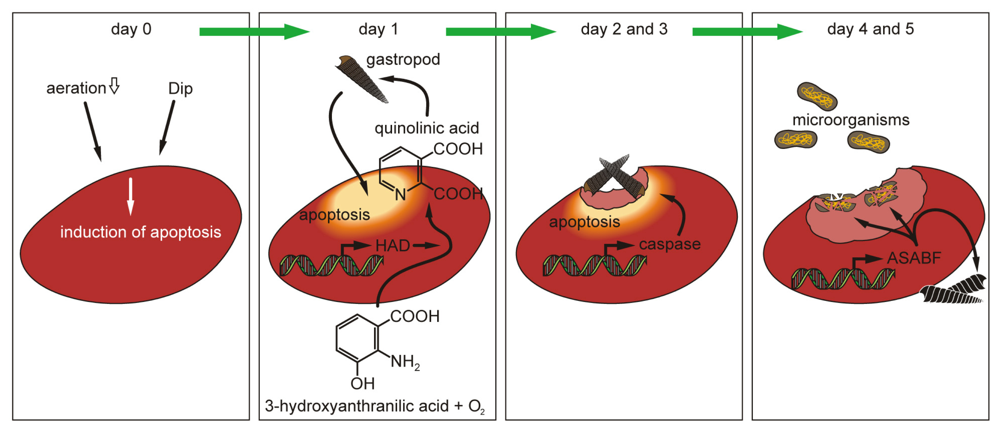
2.5. Induction of Apoptosis in S. domuncula after Exposure of the Animals to Dip
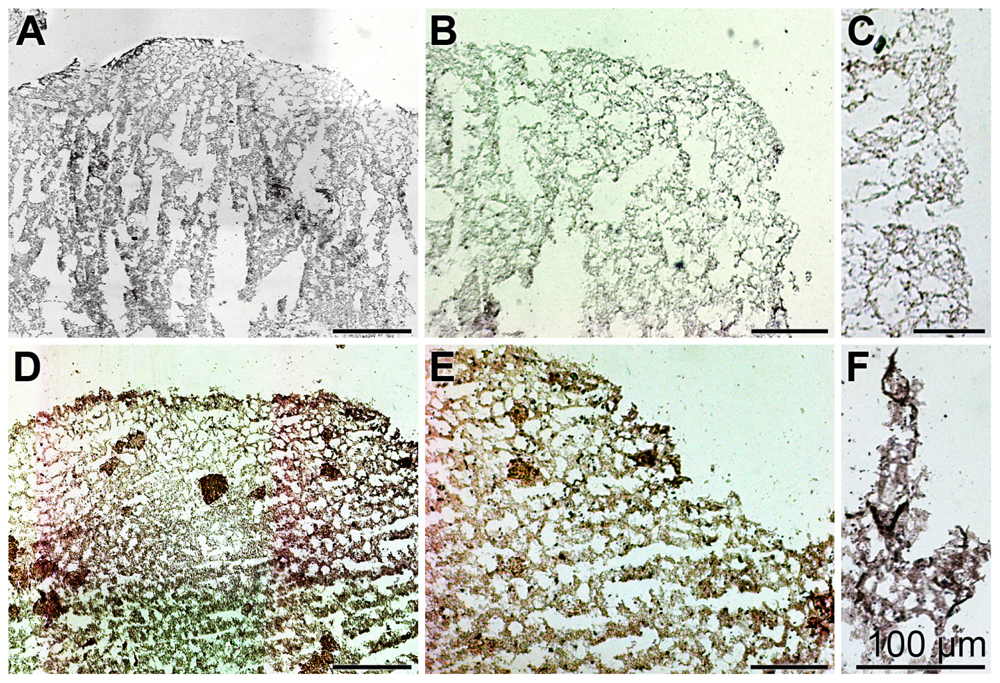
2.6. Expression of the ASABF Gene in S. domuncula Specimens: in Situ Hybridization
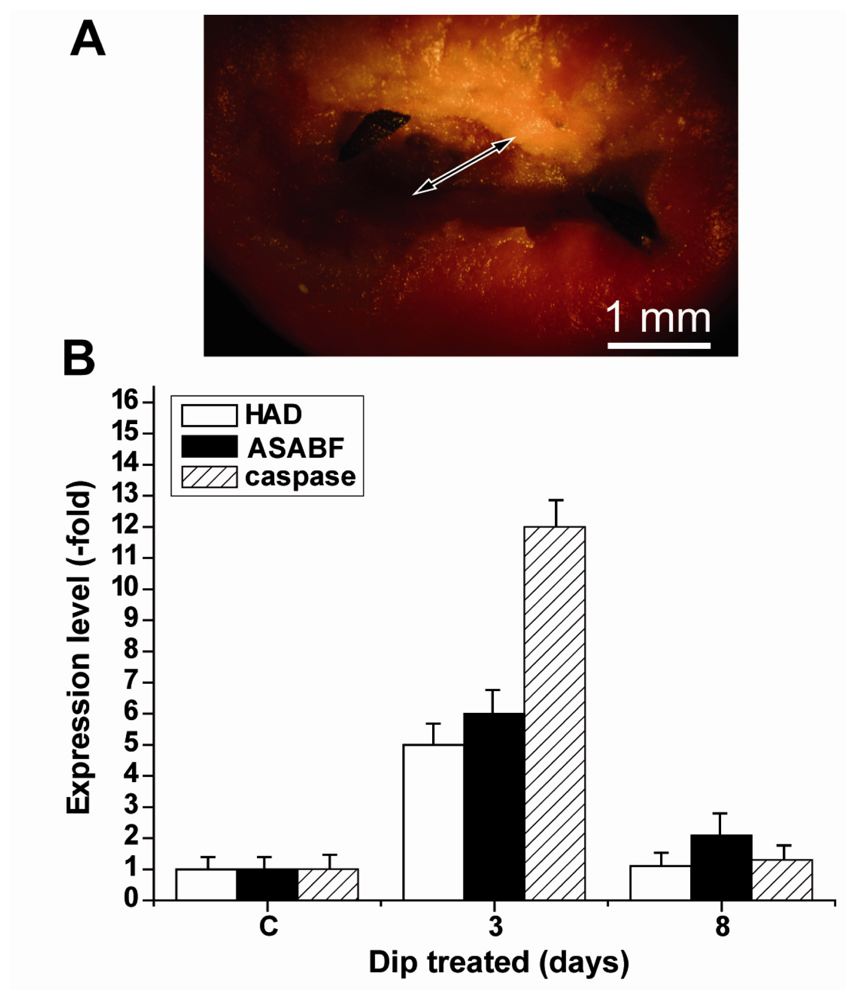
2.7. Expressions of the HAD, the ASABF as well as the Caspase Genes: Determination by qRT-PCR
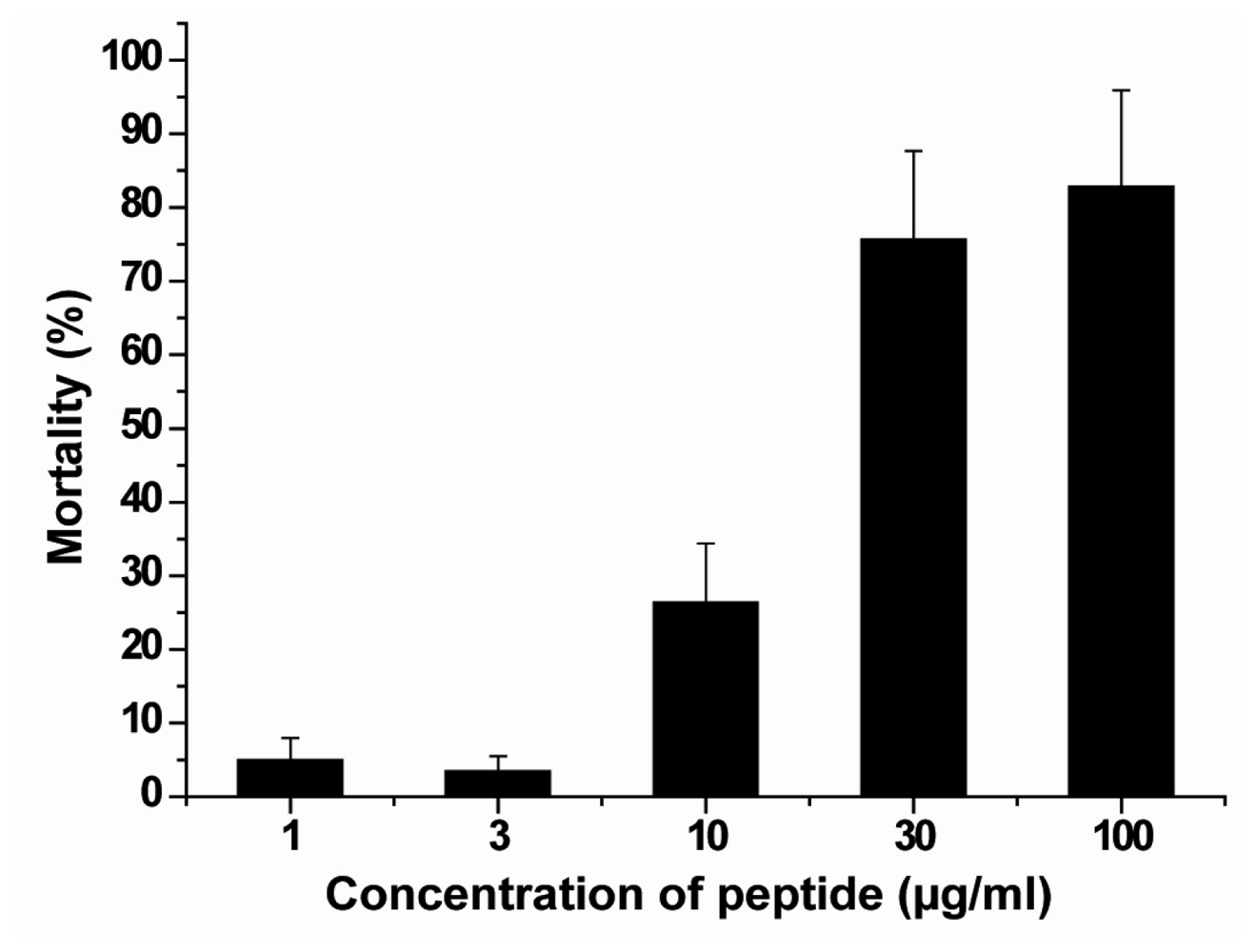
2.8. Acute/Subchronic Toxicity Testing of the Gastropod
3. Discussion
4. Experimental Section
4.1. Chemicals, Materials and Enzymes
4.2. Sponges and Gastropod/Mollusc Colonization
4.3. Maintenance of the Gastropod Bittium sp
4.4. Identification and Cloning of the Poriferan ASABF Peptides
4.5. Sequence Analyses
4.6. Preparation of Recombinant S. domuncula ASABF Peptide
4.7. Preparation of Antibodies
4.8. SDS-PAGE and Immunodetection Analysis
4.9. Quantitative Real-Time RT-PCR (qRT-PCR)
4.10. In Situ Hybridization
4.11. Microbicidal Assay
4.12. Hemolytic Assay
4.13. Acute/Subchronic Toxicity Testing
4.14. Further Methods
5. Conclusion
Acknowledgements
- †This contribution is dedicated to Dr. Peter Murphy (Townsville, Australia) in memory of his comprehensive studies in worldwide marine biodiversity.
References
- Sponges (Porifera) Marine Molecular Biotechnology; Müller, WEG (Ed.) Springer-Press: Berlin, Germany, 2003.
- Osinga, R; Belarbi, EH; Grima, EM; Tramper, J; Wijffels, RH. Progress towards a controlled culture of the marine sponge Pseudosuberites andrewsi in a bioreactor. J. Biotechnol 2003, 100, 141–144. [Google Scholar]
- Fattorusso, E; Gerwick, WH; Taglialatela-Scafati, O. Handbook of Marine Natural Products; Springer-Press: Berlin, Germany, 2011; in press. [Google Scholar]
- Müller, WEG; Klemt, M; Thakur, NL; Schröder, HC; Aiello, A; D’Esposito, M; Menna, M; Fattorusso, E. Molecular/chemical ecology in sponges: Evidence for an adaptive antibacterial response in Suberites domuncula. Mar. Biol 2003, 144, 19–29. [Google Scholar]
- Thakur, NL; Müller, WEG. Biotechnological potential of marine sponges. Curr. Sci 2004, 86, 1506–1512. [Google Scholar]
- Müller, WEG; Blumbach, B; Müller, IM. Evolution of the innate and adaptive immune systems: Relationships between potential immune molecules in the lowest metazoan phylum [Porifera] and those in vertebrates. Transplantation 1999, 68, 1215–1227. [Google Scholar]
- Blumbach, B; Diehl-Seifert, B; Seack, J; Steffen, R; Müller, IM; Müller, WEG. Cloning and expression of new receptors belonging to the immunoglobulin superfamily from the marine sponge Geodia cydonium. Immunogenetics 1999, 49, 751–763. [Google Scholar]
- Wiens, M; Korzhev, M; Perovic-Ottstadt, S; Luthringer, B; Brandt, D; Klein, S; Müller, WEG. Toll-like receptors are part of the innate immune defense system of sponges (Demospongiae: Porifera). Mol. Biol. Evol 2007, 24, 792–804. [Google Scholar]
- Perović-Ottstadt, S; Adell, T; Proksch, P; Wiens, M; Korzhev, M; Gamulin, V; Müller, IM; Müller, WEG. A (1→3)-β-d-glucan recognition protein from the sponge Suberites domuncula: Mediated activation of fibrinogen-like protein and epidermal growth factor gene expression. Eur. J. Biochem 2004, 271, 1924–1937. [Google Scholar]
- Thakur, NL; Perović-Ottstadt, S; Batel, R; Korzhev, M; Diehl-Seifert, B; Müller, IM; Müller, WEG. Innate immune defense of the sponge Suberites domuncula against gram-positive bacteria: Induction of lysozyme and AdaPTin. Mar. Biol 2005, 146, 271–282. [Google Scholar]
- Wiens, M; Korzhev, M; Krasko, A; Thakur, NL; Perović-Ottstadt, S; Breter, HJ; Ushijima, H; Diehl-Seifert, B; Müller, IM; Müller, WEG. Innate immune defense of the sponge Suberites domuncula against bacteria involves a MyD88-dependent signaling pathway: Induction of a perforin-like molecule. J. Biol. Chem 2005, 280, 27949–27959. [Google Scholar]
- Schröder, HC; Ushijima, H; Krasko, A; Gamulin, V; Schütze, J; Müller, IM; Müller, WEG. Emergence and disappearance of an immune molecule, an antimicrobial lectin, in basal Metazoa: The tachylectin family. J. Biol. Chem 2003, 278, 32810–32817. [Google Scholar]
- Müller, WEG; Wiens, M; Adell, T; Gamulin, V; Schröder, HC; Müller, IM. Bauplan of Urmetazoa: Basis for genetic complexity of Metazoa. Int. Rev. Cytol 2004, 235, 53–92. [Google Scholar]
- Zhua, S; Gao, B; Tytgat, J. Phylogenetic distribution, functional epitopes and evolution of the CSαβ superfamily. Cell. Mol. Life Sci 2005, 62, 2257–2269. [Google Scholar]
- Ueno, S; Masaomi, M; Yuji, N; Misako, T; Yasushi, T; Toshimasa, Y; Kato, Y. Generation of novel cationic antimicrobial peptides from natural non-antimicrobial sequences by acid-amide substitution. Ann. Clin. Microbiol. Antimicrobials 2011, 10, 1–7. [Google Scholar]
- Nijnik, A; Hancock, REW. Host defence peptides: Antimicrobial and immunomodulatory activity and potential applications for tackling antibiotic-resistant infections. Emerg Health Threats J 2009, 2, e1. [Google Scholar] [CrossRef]
- Poriferan EST database. Available online: https://octavia.vk.medizin.uni-mainz.de/login.cgi accessed on 15 January 2011.
- Kato, Y; Komatsu, S. ASABF, a novel cysteine-rich antibacterial peptide isolated from the nematode Ascaris suum: Purification, primary structure, and molecular cloning of cDNA. J. Biol. Chem 1996, 271, 30493–30498. [Google Scholar]
- Kato, Y; Aizawa, T; Hoshino, H; Kawano, K; Nitta, K; Zhang, H. Abf-1 and abf-2, ASABF-type antimicrobial peptide genes in Caenorhabditis elegans. Biochem. J 2002, 361, 221–230. [Google Scholar]
- Selsted, ME; Ouellette, AJ. Mammalian defensins in the antimicrobial immune response. Nature Immunol 2005, 6, 551–557. [Google Scholar]
- Minaba, M; Ueno, S; Pillai, A; Kato, Y. Evolution of ASABF (Ascaris suum antibacterial factor)-type antimicrobial peptides in nematodes: Putative rearrangement of disulfide bonding patterns. Develop. Comp. Immunol 2009, 33, 1147–1150. [Google Scholar]
- Kato, Y. Nematode antimicrobial peptides. Curr. Topics Pept. Prot. Res 2007, 8, 1–10. [Google Scholar]
- Pillai, A; Ueno, S; Zhang, H; Kato, Y. Induction of ASABF (Ascaris suum antibacterial factor)-type antimicrobial peptides by bacterial injection: Novel members of ASABF in the nematode Ascaris suum. Biochem. J 2003, 371, 663–668. [Google Scholar]
- Schröder, HC; Sudek, S; de Caro, S; de Rosa, S; Perović, S; Steffen, R; Müller, IM; Müller, WEG. Synthesis of the neurotoxin quinolinic acid in apoptotic tissue from Suberites domuncula: Cell biological, molecular biological and chemical analyses. Mar. Biotechnol 2002, 4, 546–558. [Google Scholar]
- Wiens, M; Krasko, A; Perović, S; Müller, WEG. Caspase-mediated apoptosis in sponges: Cloning and function of the phylogenetic oldest apoptotic proteases from Metazoa. Biochim. Biophys. Acta 2003, 1593, 179–189. [Google Scholar]
- Cole, C; Barber, JD; Barton, GJ. The Jpred 3 secondary structure prediction server. Nucl. Acids Res 2008, 35, W197–W201. [Google Scholar]
- Chou, PY; Fasman, GD. Prediction of the secondary structure of proteins from their amino acid sequence. Adv. Enzymol. Relat. Areas Mol. Biol 1978, 47, 45–148. [Google Scholar]
- InterProScan Sequence Search. Available online: http://www.ebi.ac.uk/Tools/pfa/iprscan/ accessed on 27 January 2011.
- BLAST Assembled RefSeq Genomes. Available online: http://blast.ncbi.nlm.nih.gov/Blast.cgi accessed on 31 January 2011.
- Wiens, M; Luckas, B; Brümmer, F; Ammar, MSA; Steffen, R; Batel, R; Diehl-Seifert, B; Schröder, HC; Müller, WEG. Okadaic acid: A potential defense molecule for the sponge Suberites domuncula. Mar. Biol 2003, 142, 213–223. [Google Scholar]
- Yeaman, MR; Yount, NY. Mechanisms of antimicrobial peptide action and resistance. Pharmacol. Rev 2003, 55, 27–55. [Google Scholar]
- Toke, O. Antimicrobial peptides: New candidates in the fight against bacterial infections. Biopolymers 2005, 80, 717–735. [Google Scholar]
- Brogden, KA; Ackermann, M; McCray, PB, Jr; Tack, BF. Antimicrobial peptides in animals and their role in host defences. Int. J. Antimicrob Agents 2003, 22, 465–478. [Google Scholar]
- Yonezawa, A; Kuwahara, J; Fujii, N; Sugiura, Y. Binding of tachyplesin I to DNA revealed by footprinting analysis: Significant contribution of secondary structure to DNA binding and implication for biological action. Biochemistry 1992, 31, 2998–3004. [Google Scholar]
- Beisswenger, C; Bals, R. Functions of antimicrobial peptides in host defense and immunity. Curr. Protein Pept. Sci 2005, 6, 255–264. [Google Scholar]
- Hochmuth, T; Piel, J. Polyketide synthases of bacterial symbionts in sponges—Evolution-based applications in natural products research. Phytochemistry 2009, 70, 1841–1849. [Google Scholar]
- Soruri, A; Grigat, J; Forssmann, U; Riggert, J; Zwirner, J. β-Defensins chemoattract macrophages and mast cells but not lymphocytes and dendritic cells: CCR6 is not involved. Eur. J. Immunol 2007, 37, 2474–2486. [Google Scholar]
- Klotman, ME; Chang, TL. Defensins in innate antiviral immunity. Nat. Rev. Immunol 2006, 6, 447–456. [Google Scholar]
- Zhang, H; Yoshida, S; Aizawa, T; Murakami, R; Suzuki, M; Koganezawa, N; Matsuura, A; Miyazawa, M; Kawano, K; Nitta, K; et al. In vitro antimicrobial properties of recombinant ASABF, an antimicrobial peptide isolated from the nematode Ascaris suum. Antimicrob. Agents Chemother 2000, 44, 2701–2705. [Google Scholar]
- Roche Applied Science, Apoptosis, Cell Death and Cell Proliferation; Roche Diagnostics: Mannheim, Germany, 2011.
- Pucci, L; Perozzi, S; Cimadamore, F; Orsomando, G; Raffaelli, N. Tissue expression and biochemical characterization of human 2-amino 3-carboxymuconate 6-semialdehyde decarboxylase, a key enzyme in tryptophan catabolism. FEBS J 2007, 274, 827–840. [Google Scholar]
- Guillemin, GJ; Wang, L; Brew, BJ. Quinolinic acid selectively induces apoptosis of human astrocytes: Potential role in AIDS dementia complex. J Neuroinflammation 2005, 2. [Google Scholar] [CrossRef] [Green Version]
- Müller, WEG; Wang, XH; Wiens, M; Schloßmacher, U; Jochum, KP; Schröder, HC. Hardening of bio-silica in sponge spicules involves an aging process after its enzymatic polycondensation: Evidence for an aquaporin-mediated water absorption. Biochim. Biophys. Acta 2011, 1810, 713–726. [Google Scholar]
- Actis, LA; Tolmasky, ME; Crosa, LM; Crosa, JH. Effect of iron-limiting conditions on growth of clinical isolates of Acinetobacter baumannii. J. Clin. Microbiol 1993, 31, 2812–2815. [Google Scholar]
- Wright, RA; Crowder, LB; Martin, TH. Selective predation by blue crabs on the gastropod, Bittium varium: Confirmation from opercula found in the sediments. Estuaries 1996, 19, 75–81. [Google Scholar]
- LePennec, G; Perović, S; Ammar, MSA; Grebenjuk, VA; Steffen, R; Müller, WEG. Cultivation of primmorphs from the marine sponge Suberites domuncula: Morphogenetic potential of silicon and iron. J. Biotechnol 2003, 100, 93–108. [Google Scholar]
- Müller, WEG; Krasko, A; Skorokhod, A; Bünz, C; Grebenjuk, VA; Steffen, R; Batel, R; Schröder, HC. Histocompatibility reaction in tissue and cells of the marine sponge Suberites domuncula in vitro and in vivo: Central role of the allograft inflammatory factor 1. Immunogenetics 2002, 54, 48–58. [Google Scholar]
- Thompson, JD; Higgins, DG; Gibson, TJ. CLUSTAL W: Improving the sensitivity of progressive multiple sequence alignment through sequence weighting, positions-specific gap penalties and weight matrix choice. Nucl. Acids Res 1994, 22, 4673–4680. [Google Scholar]
- Dayhoff, MO; Schwartz, RM; Orcutt, BC. A Model of Evolutionary Change in Protein. In Atlas of Protein Sequence and Structure; Dayhoff, MO, Ed.; Natural Biomedical Research Foundation: Washington, DC, USA, 1978; pp. 345–352. [Google Scholar]
- Saitou, N; Nei, M. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol 1987, 4, 406–425. [Google Scholar]
- Felsenstein, J. PHYLIP, Version 35; University of Washington: Seattle, WA, USA, 1993. [Google Scholar]
- Treeview. Available online: http://taxonomy.zoology.gla.ac.uk/rod/treeview.html accessed on 17 February 2011.
- Nicholas, KB; Nicholas, HB, Jr. GeneDoc: A Tool for Editing and Annotating Multiple Sequence Alignments, Version 1.1.004; 1997. [Google Scholar]
- Finn, RD; Mistry, J; Schuster-Böckler, B; Griffiths-Jones, S; Hollich, V; Lassmann, T; Moxon, S; Marshall, M; Khanna, A; Durbin, R; et al. Pfam: Clans, web tools and services. Nucl. Acids Res 2006, 34, D247–D251. [Google Scholar]
- Letunic, I; Copley, RR; Pils, B; Pinkert, S; Schultz, J; Bork, P. SMART 5: Domains in the context of genomes and networks. Nucl. Acids Res 2006, 34, D257–D260. [Google Scholar]
- Romanos, MA; Scorer, CA; Clare, JJ. Foreign gene expression in yeast: A review. Yeast 1992, 8, 423–488. [Google Scholar]
- Müller, WEG; Rothenberger, M; Boreiko, A; Tremel, W; Reiber, A; Schröder, HC. Formation of siliceous spicules in the marine demosponge Suberites domuncula. Cell Tissue Res 2005, 321, 285–297. [Google Scholar]
- Laemmli, UK. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar]
- Wiens, M; Wang, XH; Unger, A; Schröder, HC; Grebenjuk, VA; Pisignano, D; Jochum, KP; Müller, WEG. Flashing light signaling circuit in sponges: Endogenous light generation after tissue ablation in Suberites domuncula. J. Cell Biochem 2010, 111, 1377–1389. [Google Scholar]
- Müller, WEG; Wang, XH; Schröder, HC; Korzhev, M; Grebenjuk, VA; Markl, JS; Jochum, KP; Pisignano, D; Wiens, M. A cryptochrome-based photosensory system in the siliceous sponge Suberites domuncula (Demospongiae). FEBS J 2010, 277, 1182–1201. [Google Scholar]
- Livak, KJ; Schmittgen, TD. Analysis of relative gene expression data using real-time quantitative PCR and the 2 D DCT method. Methods 2001, 25, 402–408. [Google Scholar]
- Schröder, HC; Perović-Ottstadt, S; Rothenberger, M; Wiens, M; Schwertner, H; Batel, R; Korzhev, M; Müller, IM; Müller, WEG. Silica transport in the demosponge Suberites domuncula: Fluorescence emission analysis using the PDMPO probe and cloning of a potential transporter. Biochem. J 2004, 381, 665–673. [Google Scholar]
- Seibert, G; Raether, W; Dogovic, N; Gasic, MJ; Zahn, RK; Müller, WEG. Antibacterial and antifungal activity of avarone and avarol. Zentralbl. Bakteriol. Mikrobiol. Hyg. A 1985, 260, 379–386. [Google Scholar]
- Thakur, NL; Hentschel, U; Krasko, A; Pabel, CT; Anil, AC; Müller, WEG. Antibacterial activity of the sponge Suberites domuncula and its primmorphs: Potential basis for the defense. Aquat. Microbiol. Ecol 2003, 31, 77–83. [Google Scholar]
- Wei, GX; Bobek, LA. In vitro synergic antifungal effect of MUC7 12-mer with histatin-5 12-mer or miconazole. J. Antimicr. Chemother 2004, 53, 750–758. [Google Scholar]
- Sachs, L. Angewandte Statistik; Springer: Berlin, Germany, 1984; p. 242. [Google Scholar]
- Compton, S; Jones, CG. Mechanism of dye response and interference in the Bradford protein assay. Anal. Biochem 1985, 151, 369–374. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Microorganism | Antimicrobial activity MIC (μg/mL) | ||
|---|---|---|---|
| Non-Adsorbed ASABF | Adsorbed ASABF | ||
| Gram-positive bacteria: | Staphylococcus aureus | 1.7 ± 0.5 | >20 |
| Bacillus subtilis | 4.8 ± 0.6 | >20 | |
| Micrococcus luteus | 3.6 ± 1.0 | >20 | |
| Gram-negative bacteria: | Pseudomonas aeruginosa | 12.4 ± 5.5 | >20 |
| Escherichia coli | 17.4 ± 6.0 | >20 | |
| Fungi: | Candida albicans | 8.5 ± 4.0 | >20 |
| Aspergillus niger | 12.9 ± 4.0 | >20 | |
| Time point (days) | Gene expression: (mRNAx ± SEM)/mRNAtubulin | |||||
|---|---|---|---|---|---|---|
| HAD | ASABF | Caspase | ||||
| Control (×10−3) | Dip-Treated (×10−3) | Control (×10−2) | Dip-Treated (×10−2) | Control (×10−4) | Dip-Treated (×10−4) | |
| 0 | 2.1 ± 0.3 | 2.2 ± 0.3 | 1.3 ± 0.2 | 1.3 ± 0.2 | 1.3 ± 0.1 | 1.5 ± 0.2 |
| 1 | 2.3 ± 0.3 | 5.3 ± 0.6 | 0.9 ± 0.1 | 2.8 ± 0.3 | 1.6 ± 0.2 | 6.7 ± 0.7 |
| 2 | 2.4 ± 0.4 | 27.1 ± 3.8 | 1.2 ± 0.2 | 4.0 ± 0.5 | 1.9 ± 0.2 | 9.3 ± 1.0 |
| 3 | 1.9 ± 0.2 | 19.9 ± 1.7 | 1.7 ± 0.2 | 6.3 ± 0.8 | 1.8 ± 0.1 | 12.4 ± 1.7 |
| 4 | 1.9 ± 0.2 | 7.2 ± 0.9 | 1.6 ± 0.2 | 9.8 ± 0.9 | 1.4 ± 0.3 | 6.9 ± 0.8 |
| 5 | 2.2 ± 0.5 | 4.2 ± 0.6 | 2.1 ± 0.3 | 6.9 ± 0.7 | 1.8 ± 0.2 | 1.7 ± 0.1 |
| 7 | 2.3 ± 0.3 | 3.4 ± 0.4 | 1.9 ± 0.4 | 3.7 ± 0.5 | 1.8 ± 0.2 | 1.5 ± 0.2 |
| 9 | 2.5 ± 0.4 | 3.1 ± 0.3 | 1.8 ± 0.2 | 4.4 ± 0.5 | 2.1 ± 0.3 | 1.9 ± 0.3 |
© 2011 by the authors; licensee MDPI, Basel, Switzerland This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Wiens, M.; Schröder, H.C.; Korzhev, M.; Wang, X.-H.; Batel, R.; Müller, W.E.G. Inducible ASABF-Type Antimicrobial Peptide from the Sponge Suberites domuncula: Microbicidal and Hemolytic Activity in Vitro and Toxic Effect on Molluscs in Vivo. Mar. Drugs 2011, 9, 1969-1994. https://doi.org/10.3390/md9101969
Wiens M, Schröder HC, Korzhev M, Wang X-H, Batel R, Müller WEG. Inducible ASABF-Type Antimicrobial Peptide from the Sponge Suberites domuncula: Microbicidal and Hemolytic Activity in Vitro and Toxic Effect on Molluscs in Vivo. Marine Drugs. 2011; 9(10):1969-1994. https://doi.org/10.3390/md9101969
Chicago/Turabian StyleWiens, Matthias, Heinz C. Schröder, Michael Korzhev, Xiao-Hong Wang, Renato Batel, and Werner E. G. Müller. 2011. "Inducible ASABF-Type Antimicrobial Peptide from the Sponge Suberites domuncula: Microbicidal and Hemolytic Activity in Vitro and Toxic Effect on Molluscs in Vivo" Marine Drugs 9, no. 10: 1969-1994. https://doi.org/10.3390/md9101969