Nature and Lability of Northern Adriatic Macroaggregates

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
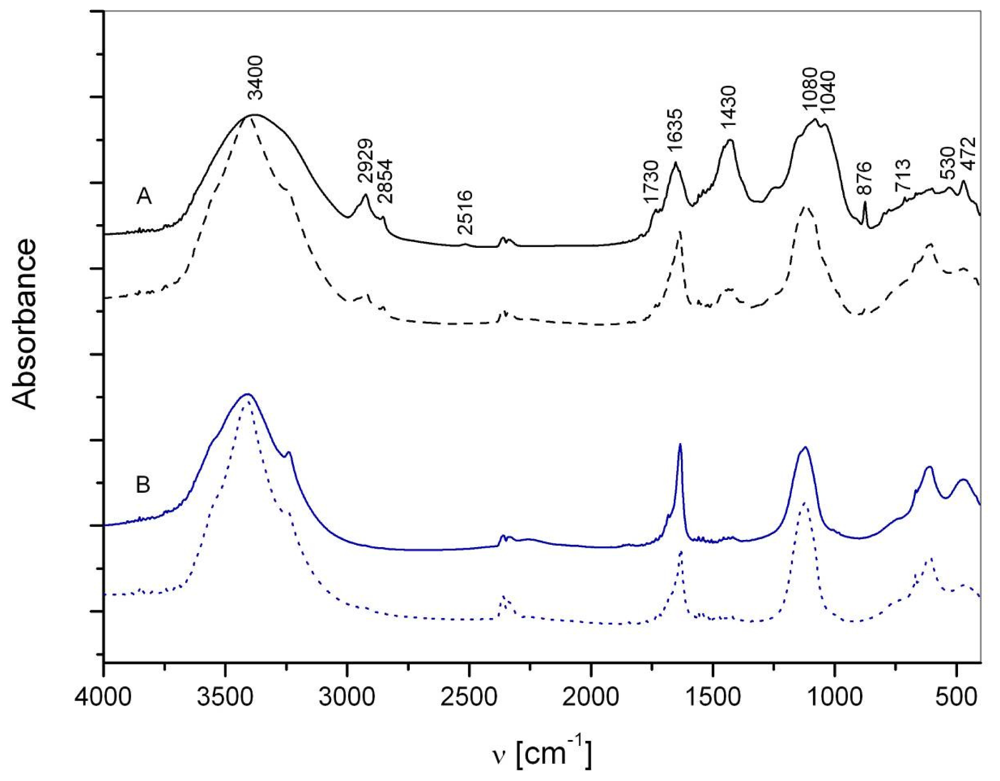
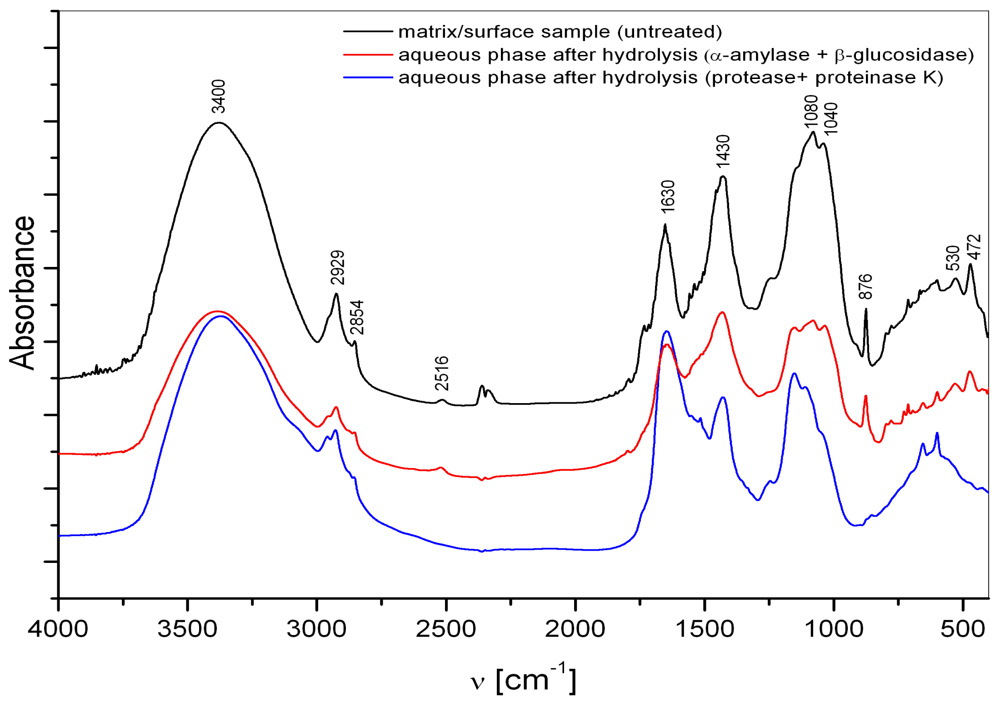
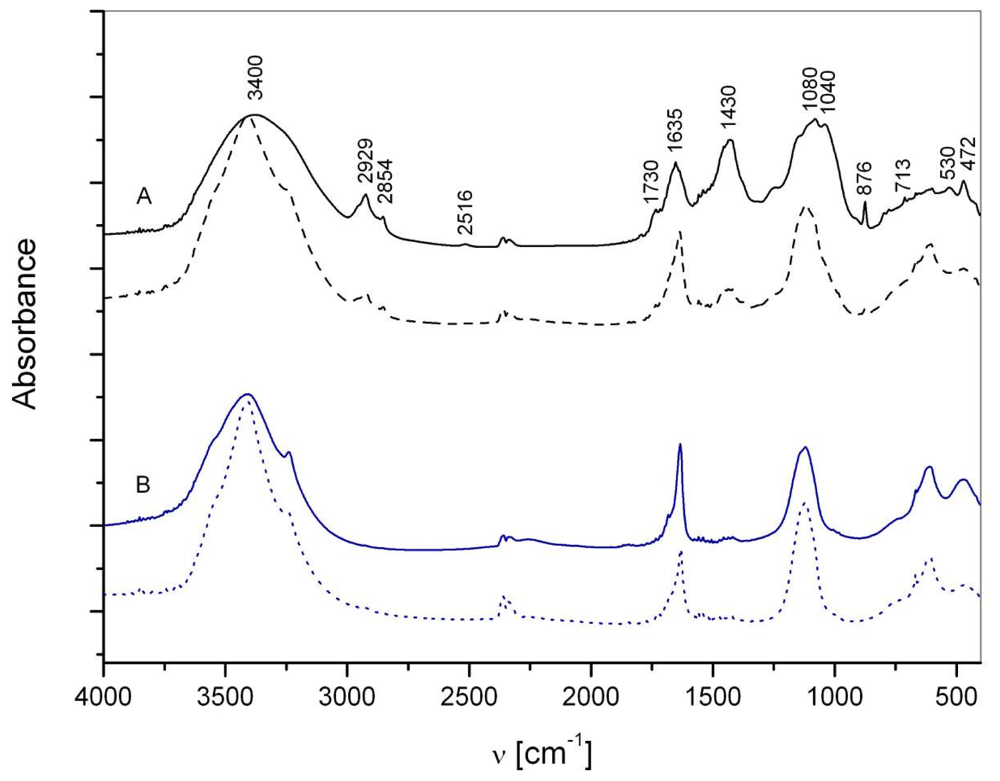
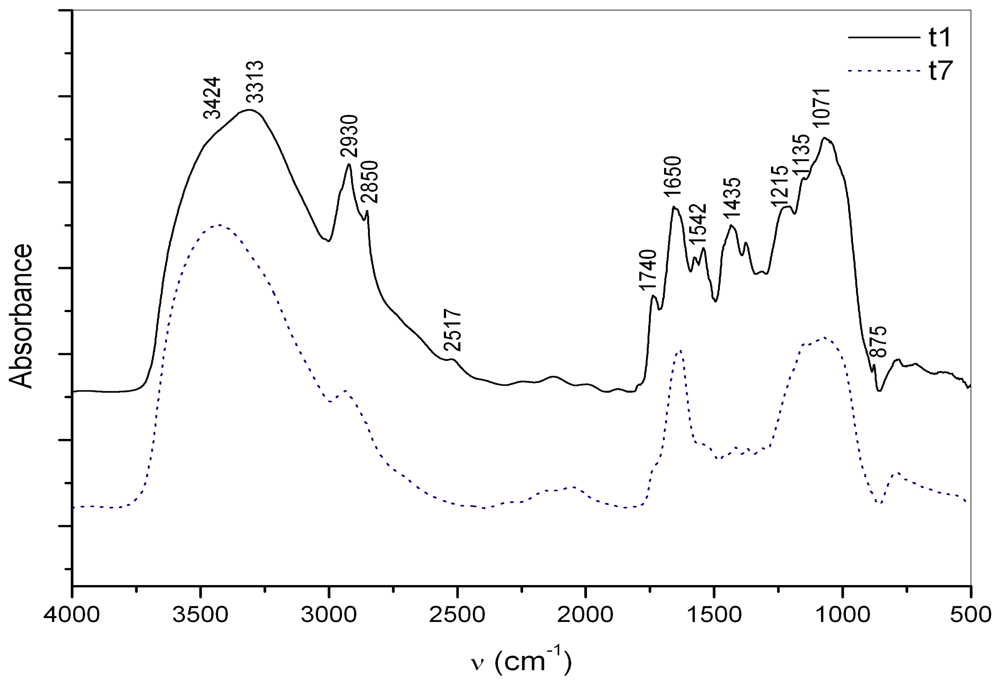
2.1. Macroaggregate composition: matrix
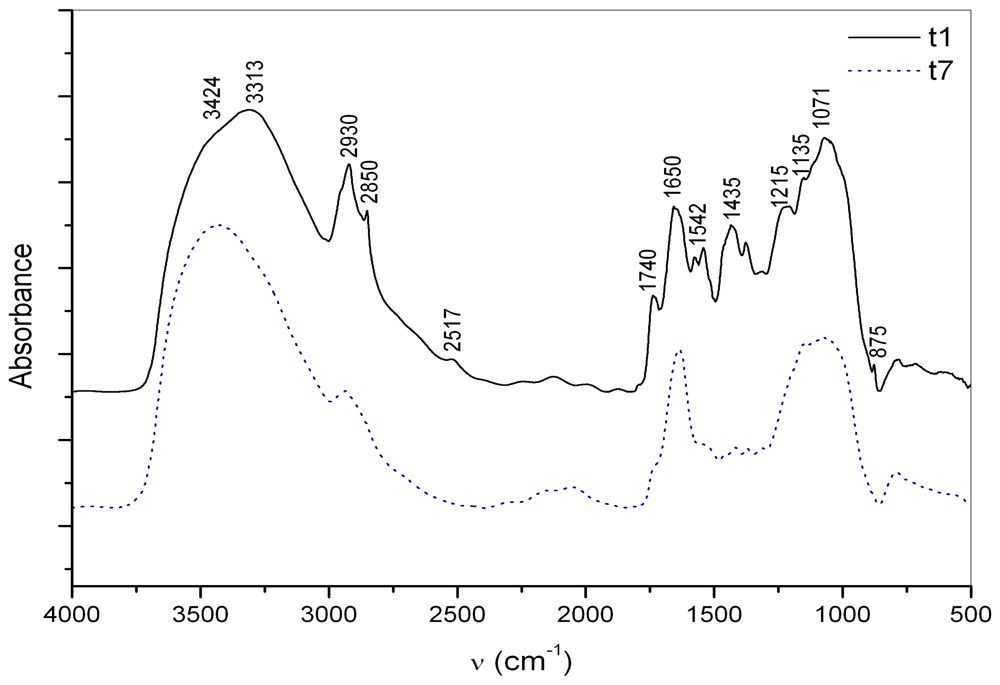
2.2. Macroaggregate composition: interstitial water colloids
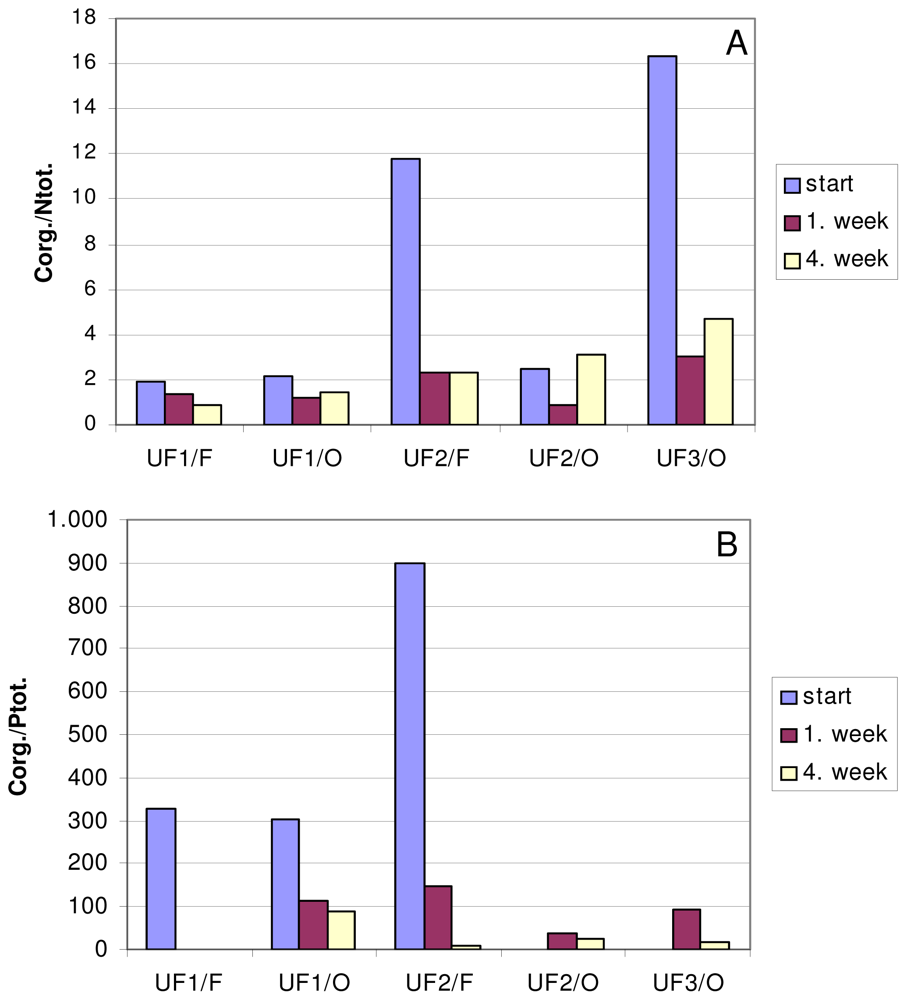
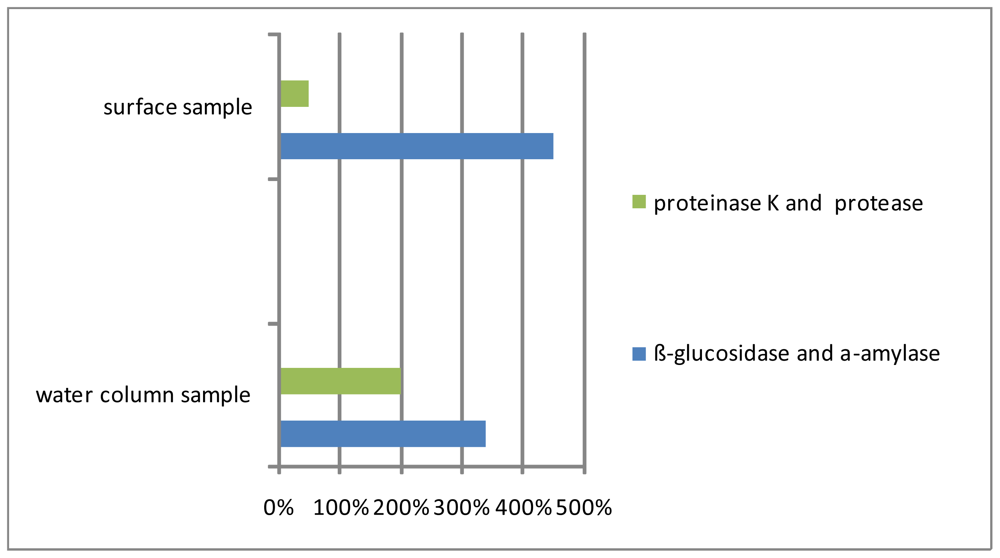
3. Macroaggregate Lability
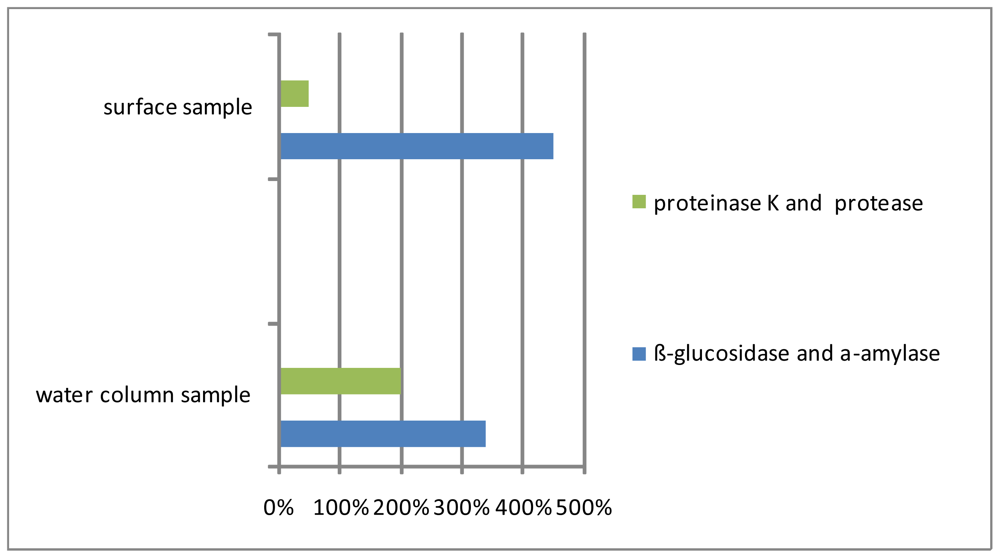
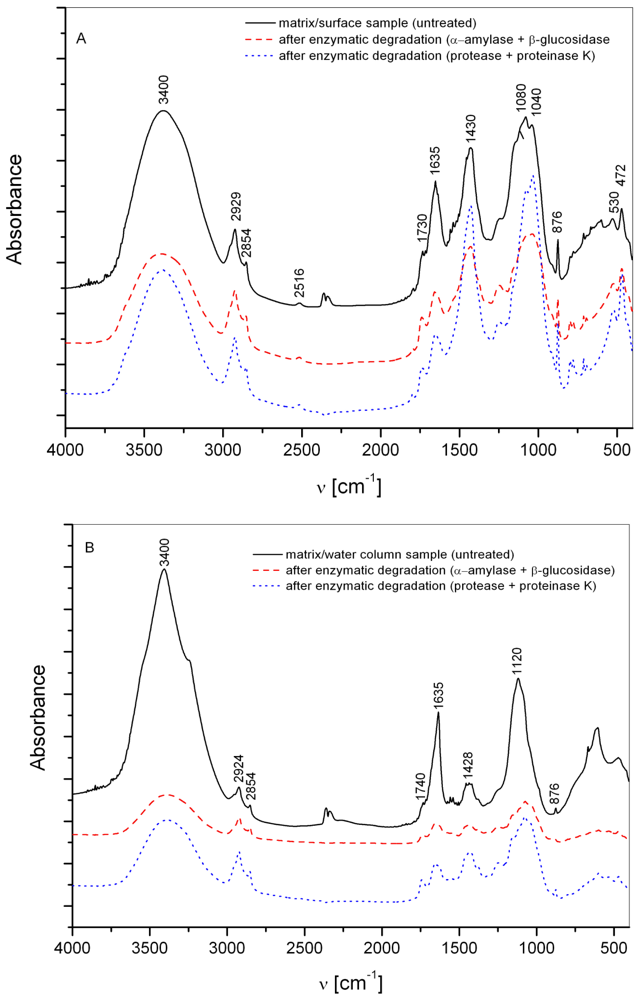
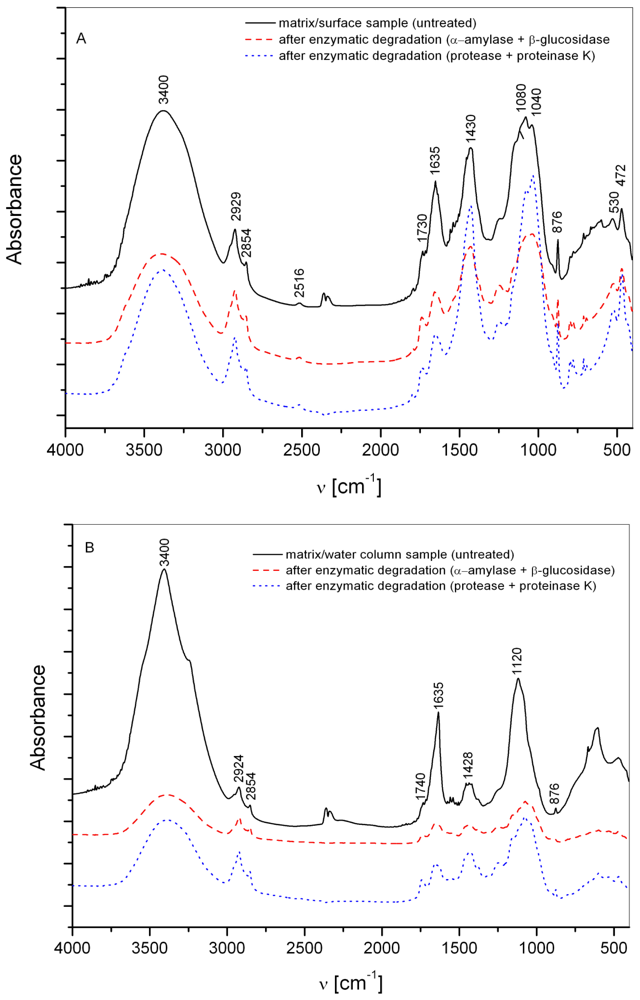
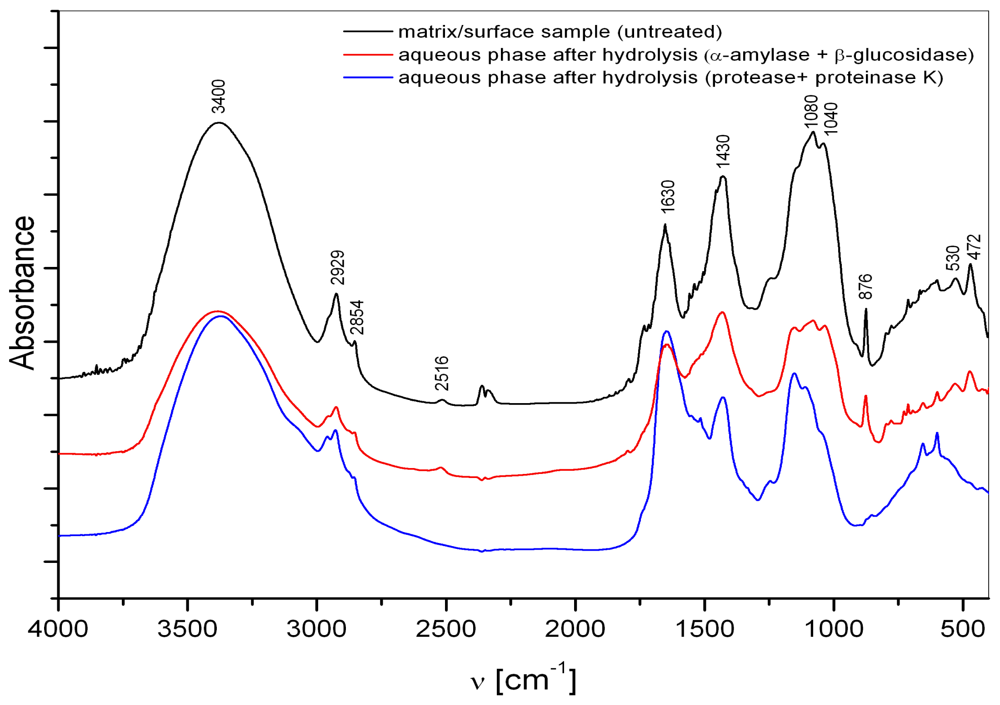
3.1. Matrix
3.2. Interstitial water colloids
4. Experimental Section
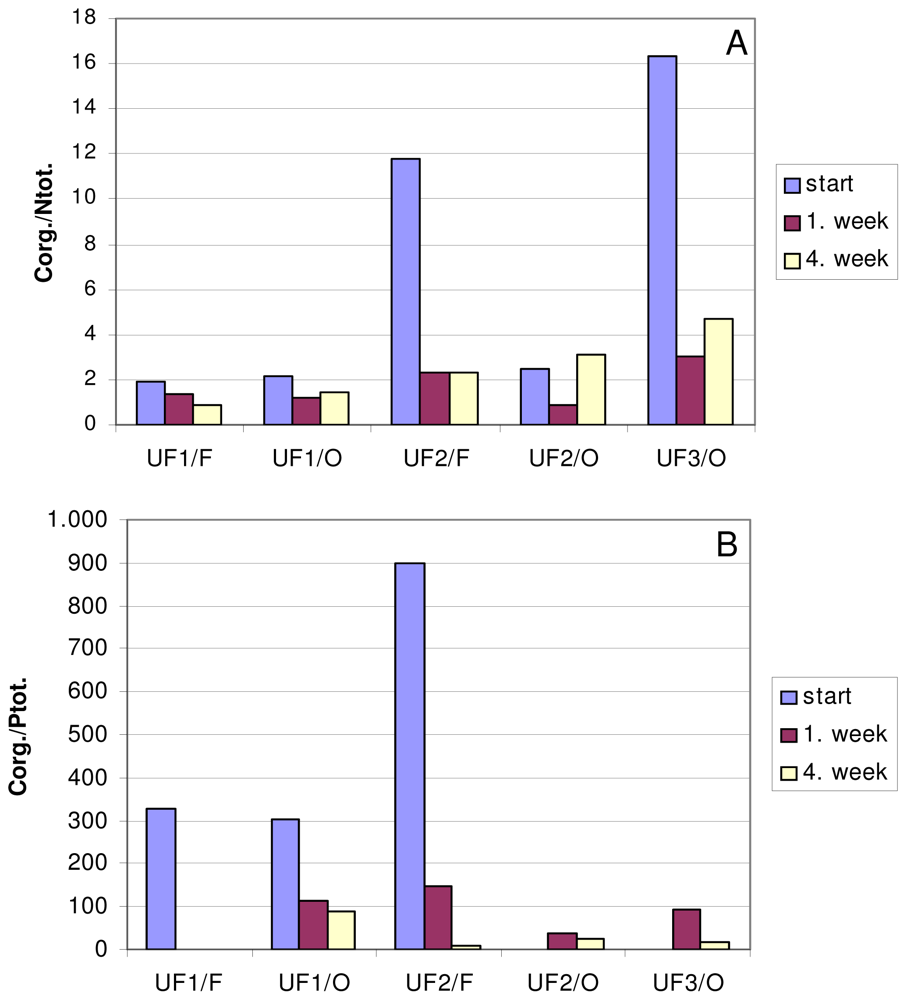
4.1. Samples
4.2. Degradation experiments and separations
4.2.1. Matrix
4.2.2. Interstitial water colloids
4.3. Analyses
5. Conclusions
- Samples Availability: Available from the authors.
References
- Cozzi, S; Ivančić, I; Catalano, G; Djakovac, T; Degobbis, D. Dynamics of the oceanographic properties during mucilage appearance in the Northern Adriatic Sea: Analysis of the 1997 event in comparison to earlier events. J. Mar. Syst 2004, 50, 223–241. [Google Scholar]
- Penna, N; Kovac, N; Ricci, F; Penna, A; Capellacci, S; Faganeli, J. The role of dissolved carbohydrates in the northern Adriatic macroaggregate formation. Acta Chim. Slov 2009, 56, 305–314. [Google Scholar]
- Myklestad, SM. Production of carbohydrates by marine planktonic diatoms. Influence of N/P ratio in the growth medium on the assimilation ratios, growth rate and production of cellular and extracellular carbohydrates by Chaetoceros affinis var, willei (Gran) Husted and Skeletonema costatum (Grev) Cleve. J. Exp. Mar. Biol. Ecol 1977, 29, 161–179. [Google Scholar]
- Penna, N; Rinaldi, A; Montanari, G; Di Paolo, A; Penna, A. Mucilaginous masses in the Adriatic Sea in the summer of 1989. Water Res 1993, 27, 1767–1771. [Google Scholar]
- Maestrini, SY; Breret, M; Bechim, C; Berland, BR; Poletti, R; Rinaldi, A. Nutrients limiting the algal growth potential (AGP) in the Po River Plume and an adjacent area, northwest Adriatic Sea: Enrichment bioassays with the test algae Nitzschia closterium and Thalassiosira pseudonana. Estuaries 1997, 20, 416–429. [Google Scholar]
- Myklestad, SM. Release of extracellular products by phytoplankton with special emphasis on polysaccharides. Sci. Total Environ 1995, 165, 155–164. [Google Scholar]
- Baldi, F; Minacci, A; Saliot, A; Mejanelle, L; Mozetic, P; Turk, V; Malej, A. Cell lysis and release of particulate polysaccharides in extensive marine mucilage assessed by lipid biomarkers and molecular probes. Mar. Ecol. Prog. Ser 1997, 153, 45–58. [Google Scholar]
- Fanuko, N; Rode, J; Drašlar, K. Microflora from the Adriatic mucous aggregations. Biol. Vestn 1989, 37, 27–34. [Google Scholar]
- Stachowitsch, M; Funuko, N; Richter, M. Mucus aggregates in the Adriatic Sea: An overview of stages and occurrences. Mar. Ecol 1990, 11, 327–350. [Google Scholar]
- Degobbis, D; Fonda-Umani, S; Franco, P; Malej, A; Precali, R; Smodlaka, N. Changes in the northern Adriatic ecosystem and the hypertrophic appearance of gelatinous aggregates. Sci. Total Environ 1995, 165, 43–58. [Google Scholar]
- Kovač, N; Mozetič, P; Trichet, J; Défarge, C. Phytoplankton composition and organic matter organization of mucous aggregates by means of light and cryo-scanning electron microscopy. Mar. Biol 2005, 147, 261–271. [Google Scholar] [Green Version]
- Degobbis, D; Precali, R; Ferrari, CR; Djakovac, T; Rinaldi, A; Ivančić, I; Gismondi, M; Smodlaka, N. Change in nutrient concentrations and ratios during mucilage event in the period 1999–2002. Sci. Total Environ 2005, 353, 103–114. [Google Scholar]
- Supić, N; Orlić, M. Seasonal and interannual variability of the northern Adriatic surface fluxes. J. Mar. Syst 2000, 20, 205–229. [Google Scholar]
- Chin, WC; Orellana, MV; Verdugo, P. Spontaneous assembly of marine dissolved organic matter in polymer gels. Nature 1998, 391, 568–572. [Google Scholar]
- Verdugo, P; Alldredge, AL; Azam, F; Kirchman, DL; Passow, U; Santschi, PH. The oceanic gel phase: A bridge in the DOM-POM continuum. Mar. Chem 2004, 92, 67–85. [Google Scholar]
- Svetličić, V; Žutić, V; Zimmermann, AH. Biophysical scenario of giant gel formation in the Northern Adriatic Sea. Ann. N. Y. Acad. Sci 2005, 1048, 524–527. [Google Scholar]
- Kovač, N; Faganeli, J; Bajt, O. Stefansson, O, Ed.; Mucous macroaggregates in the Northern Adriatic. In Geochemistry Research Advances; Nova Science: New York, NY, USA, 2008; pp. 119–141. [Google Scholar]
- Kovac, N; Faganeli, J; Bajt, O; Sket, B; Orel, B; Penna, N. Chemical composition of macroaggregates in the northern Adriatic Sea. Org. Geochem 2004, 35, 1095–1104. [Google Scholar]
- Herndl, GJ. Marine snow in the Northern Adriatic Sea: Possible couses and consequences for a shallow ecosystem. Mar. Microb. Food Webs 1992, 6, 149–172. [Google Scholar]
- Rinaldi, A; Montanari, G; Ferrari, CR; Ghetti, A; Vollenweider, AR. Evoluzione dello stato trofico nelle acque costiere Emiliano-Romagnole nel periodo 1982–1994. Proceedings of Evoluzione dello stato trofico in Adriatico: Analisi degli interventi attuati e future linee di intervento, Bologna, Italy, 28–29 September 1995; pp. 33–49.
- Mingazzini, M; Thake, B. Summary and conclusions of the workshop on marine mucilages in the Adriatic Sea and elsewhere. Sci. Total Environ 1995, 165, 9–14. [Google Scholar]
- Müller-Niklas, S; Schuster, S; Kaltenböck, E; Herndl, GJ. Organic content and bacterial metabolism in amorphous aggregations of the Northern Adriatic Sea. Limnol. Oceanogr 1994, 39, 58–68. [Google Scholar]
- Grilli, F; Marini, M; Degobbis, D; Ferrari, CR; Fornasiero, P; Russo, A; Gismondi, M; Djakovac, T; Precali, R; Simonetti, R. Circulation and horizontal fluxes in the northern Adriatic Sea in the period June 1999–July 2002. Part II: Nutrients transport. Sci. Total Environ 2005, 353, 115–125. [Google Scholar]
- Nishikida, K; Hannah, RW. Selected Applications of Modern FT-IR Techniques; CRC: Boca Raton, FL, USA, 1996; p. 279. [Google Scholar]
- Legras, JI; Chuzel, G; Arnaud, A; Galzy, P. Natural nitriles and their metabolism. World J. Microbiol. Biotechnol 1990, 6, 83–108. [Google Scholar]
- Leskovšek, H; Perko, S; Žigon, D; Faganeli, J. Analysis of carbohydrates in marine particulates by gas chromatography and tandem mass spectrometry. Analyst 1994, 119, 1125–1128. [Google Scholar]
- Binkley, ER; Binkley, RW. Carbohydrate Photochemistry; American Chemical Society: Washington DC, USA, 1999; p. 448. [Google Scholar]
- Chin, Y-P; Aiken, GR; O’Loughlin, E. Molecular weight, polydispersity, and spectroscopic properties of aquatic humic substances. Environ. Sci. Technol 1994, 28, 1853–1858. [Google Scholar]
- Sannigrahi, P; Ingall, ED; Benner, R. Nature and dynamics of phosphorous-containing components of marine dissolved and particulate organic matter. Geochim. Cosmochim. Acta 2006, 70, 5868–5882. [Google Scholar]
- Faganeli, J; Ogrinc, N; Kovac, N; Kukovec, K; Falnoga, I; Mozetic, P; Bajt, O. Carbon and nitrogen isotope composition of particulate organic matter in relation to mucilage formation in the northern Adriatic Sea. Mar. Chem 2009, 114, 102–109. [Google Scholar]
- Smucker, RA; Dawson, R. Products and photosynthesis by marine phytoplankton: Chitin in TCA “protein” precipitates. J. Exp. Mar. Biol. Ecol 1986, 104, 143–152. [Google Scholar]
- Kovac, N; Bajt, O; Faganeli, J; Sket, B; Orel, B. Study of macroaggregate composition using FT-IR and 1H-NMR spectroscopy. Mar. Chem 2002, 78, 205–215. [Google Scholar]
- Herndl, GJ; Arrieta, JM; Stoderegger, K. Interaction between specific hydrological and microbial activity leading to extensive mucilage formation in the northern Adriatic Sea. Ann. Ist. Super. Sanità 1999, 35, 405–409. [Google Scholar]
- Zaccone, R; Carus, G; Cal, C. Heterotrophic bacteria in the northern Adriatic Sea: Seasonal changes and ectoenzyme profile. Mar. Environ. Res 2002, 54, 1–19. [Google Scholar]
- Grossart, HP; Ploug, H. Microbial degradation of organic carbon and nitrogen on diatom aggregates. Limnol. Oceanogr 2001, 46, 267–277. [Google Scholar]
- Kovac, N; Faganeli, J; Bajt, O; Sket, B; Surca Vuk, A; Orel, B; Mozetič, P. Degradation and preservation of organic matter in marine macroaggregates. Acta Chim. Slov 2006, 53, 81–87. [Google Scholar]
- Hoppe, HG; Ullrich, S. Profile of ectoenzymes in the Indian Ocean: Phenomena of phosphatase activity in the mesopelagic zone. Aquat. Microb. Ecol 1999, 19, 139–148. [Google Scholar]
- Chrost, RJ; Gajewski, AJ. Microbial utilization of lipids in lake water. FEMS Microbiol. Ecol 1995, 18, 45–50. [Google Scholar]
- Zoppini, A; Puddu, A; Fazi, S; Rosati, M; Sist, P. Extracellular enzyme activity and dynamics of bacterial community in mucilaginous aggregates of the northern Adriatic Sea. Sci. Total Environ 2005, 353, 270–286. [Google Scholar]
- Aldrkamp, A-C; Buma, AGJ; Rijssel, M. The carbohydrates of Phaeocystis and their degradation in the microbial food web. Biogeochemistry 2007, 83, 99–118. [Google Scholar]
- Koron, N; Faganeli, J; Falnoga, I; Kovac, N. Interaction and recativity of macroaggregates and Hg in coastal waters (Gulf of Trieste, northern Adriatic Sea). Geomicrobiol. J 2010. submitted. [Google Scholar]
- Ogawa, H; Amagai, Y; Koike, I; Kaiser, K; Benner, R. Production and refractory dissolved organic matter by bacteria. Science 2001, 292, 917–920. [Google Scholar]
- Nagata, T; Meon, B; Kirchman, DL. Microbial degradation of peptidoglycan in seawater. Limnol. Oceanogr 2003, 48, 745–754. [Google Scholar]
- Murphy, J; Riley, JP. A modified single solution method for the determination of phosphate in natural waters. Anal. Chim. Acta 1962, 27, 31–36. [Google Scholar]
- Setchell, FW. Particulate protein measurements in oceanographic samples by dye binding. Mar. Chem 1981, 10, 301–313. [Google Scholar]
- Dubois, M; Gilles, KA; Hamilton, JK; Rebers, PA; Smith, F. Colorimetric method for determination of sugars and related substances. Anal. Chem 1956, 28, 350–356. [Google Scholar]
- Folch, J; Lees, M; Sloane-Stanley, GH. A simple method for the isolation and purification of total lipids from animal tissues. J. Biol. Chem 1957, 226, 497–502. [Google Scholar]
© 2010 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Faganeli, J.; Mohar, B.; Kofol, R.; Pavlica, V.; Marinšek, T.; Rozman, A.; Kovač, N.; Vuk, A.Š. Nature and Lability of Northern Adriatic Macroaggregates. Mar. Drugs 2010, 8, 2480-2492. https://doi.org/10.3390/md8092480
Faganeli J, Mohar B, Kofol R, Pavlica V, Marinšek T, Rozman A, Kovač N, Vuk AŠ. Nature and Lability of Northern Adriatic Macroaggregates. Marine Drugs. 2010; 8(9):2480-2492. https://doi.org/10.3390/md8092480
Chicago/Turabian StyleFaganeli, Jadran, Bojana Mohar, Romina Kofol, Vesna Pavlica, Tjaša Marinšek, Ajda Rozman, Nives Kovač, and Angela Šurca Vuk. 2010. "Nature and Lability of Northern Adriatic Macroaggregates" Marine Drugs 8, no. 9: 2480-2492. https://doi.org/10.3390/md8092480