Halogenated Compounds from Marine Algae

1
IPIMAR, Av. de Brasília, 1449-006 Lisboa, Portugal
2
Centro de Química e Bioquímica/Departamento de Química e Bioquímica da Faculdade de Ciências da Universidade de Lisboa, Ed C8, Piso 5, Campo Grande, 1749-016 Lisboa, Portugal
*
Author to whom correspondence should be addressed.
Mar. Drugs 2010, 8(8), 2301-2317; https://doi.org/10.3390/md8082301
Submission received: 30 June 2010
/
Revised: 23 July 2010
/
Accepted: 2 August 2010
/
Published: 9 August 2010
(This article belongs to the Special Issue Bioactive Halogenated Metabolites of Marine Origin)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Marine algae produce a cocktail of halogenated metabolites with potential commercial value. Structures exhibited by these compounds go from acyclic entities with a linear chain to complex polycyclic molecules. Their medical and pharmaceutical application has been investigated for a few decades, however other properties, such as antifouling, are not to be discarded. Many compounds were discovered in the last years, although the need for new drugs keeps this field open as many algal species are poorly screened. The ecological role of marine algal halogenated metabolites has somehow been overlooked. This new research field will provide valuable and novel insight into the marine ecosystem dynamics as well as a new approach to comprehending biodiversity. Furthermore, understanding interactions between halogenated compound production by algae and the environment, including anthropogenic or global climate changes, is a challenging target for the coming years. Research of halogenated metabolites has been more focused on macroalgae than on phytoplankton. However, phytoplankton could be a very promising material since it is the base of the marine food chain with quick adaptation to environmental changes, which undoubtedly has consequences on secondary metabolism. This paper reviews recent progress on this field and presents trends on the role of marine algae as producers of halogenated compounds.
1. Introduction
Marine algae produce a wide variety of remarkable natural compounds, usually referred to as secondary metabolites because they are not involved in the basic machinery of life [1]. Although these molecules often contribute to only a very small fraction of the organism total biomass [2], the contribution of these compounds to survival may sometimes be comparable to metabolites resulting from the primary metabolism [3]. In that sense, the use of the term “secondary metabolite” seems less appropriate since these compounds also contribute to growth, reproduction and defense and thus play a primary role for the organism integrity.
Many of these secondary metabolites are halogenated, reflecting the availability of chloride and bromide ions in seawater. Interestingly, bromide is more frequently used by algae for organohalogen production, although chlorine occurs in higher concentrations than bromine in seawater. Marine halogenated compounds comprise a varied assembly of compounds, ranging from peptides, polyketides, indoles, terpenes, acetogenins and phenols to volatile halogenated hydrocarbons [4]. The prevalence of halogens is not similar in marine algae: chlorine and bromine appear to be the main halogens used to increase biological activity of secondary metabolites, whereas iodine and fluorine remain quite unusual within the chemical structures [5]. However, some orders of brown algae such as Laminariales accumulate and use iodine for halogenation processes. For example, the kelp Laminaria digitata accumulates iodine to more than 30,000-times the concentration found in seawater, representing an average content of 1% of dry weight [6]. In fact, iodination is more frequent in brown algae than in red and green algae metabolites [6]. As a result, only less than 1% of secondary metabolites from of brown algae contain bromine or chlorine in contrast with as much as 90 and 7% of red and green algal compounds, respectively [7].
Halogenation often provides these compounds with interesting key features and marine algae hold diverse and unique biosynthetic pathways for the production of halogenated metabolites. The halogenated sesterterpenes, neomangicols A–C, isolated from the marine fungus Fusarium [8], offer a striking example of the halogenation effect. Neomangicol A and B display in vitro cytotoxic effect toward the HCT-116 human colon tumor cell line, while their nonhalogenated analog neomangicol C was shown to be inactive [8].
Biological properties of halogenated compounds have been researched for the past decades, with results showing antibacterial, antifungal, antiviral, anti-inflammatory, antiproliferative, antifouling, antifeedant, cytotoxic, ichthyotoxic, and insecticidal activity [9]. Research major focal points indubitably have been the discovery and characterization of new halogenated compounds, along with a remarkable effort toward the evaluation of their possible biomedical and biotechnological applications. Also, chemical profiles have been used to differentiate cryptic species but the chemotaxonomic value of natural halogenated compounds has been questioned due to geographical and seasonal variations in the chemical composition of algal species [10]. The ecological role of marine halogenated metabolites, alongside with other natural compounds, has been disregarded, although a few halogenated metabolites have been shown to have important and critical roles on the community structure of marine ecosystems in previous years [11–17]. In fact, most of the publications on marine ecological issues rarely include a chemical approach of compounds responsible for the ecological interactions found. Excellent reviews on marine natural compounds have been recently published [9,18,19], with no particular emphasis on halogenated metabolites. These showed a variety of structure types that were isolated from a wide range of marine organisms, including microorganisms and phytoplankton, macroalgae, sponges, cnidarians, bryozoans, molluscs, tunicates, echinoderms and true mangrove plants. During 2009, new naturally occurring algal halogenated compounds were found, most of them with little or no reported biological activity. Reports on the ecological role of algal natural compounds, presenting both chemical and ecological approaches were still few. The present paper focuses on halogenated compounds only, and is restricted to micro and macroalgae as natural sources. It attempts to provide an overview of new marine halogenated compounds reported in 2009 and to examine progress and present trends on the role of marine algae as producers of halogenated compounds.
2. Macroalgae
Interest in the actual marine natural compounds responsible for such a wide range of properties is far more recent and new compounds are still being discovered and isolated, chemical structures elucidated and properties being sorted out, with the aim of finding new molecular entities with industrial application. From time to time, a new compound emerges as particularly promising in a specific area and additional testing starts to allow further pharmacological evaluation.
Many compounds have been found in marine macroalgae in recent years, mainly in red and brown algae, and fewer in green algae [9]. Among all marine macroalgae, red algae are the main producers of halogenated compounds. Laurencia (family Rhodomalaceae, order Ceramiales, class Rhodophyceae, phylum Rhodophyta) is considered one of the most prolific genera [20,21], being mainly found in tropical, subtropical, and temperate coastal waters. It has been intensively screened over the last fifty years, although a variety of new halogenated molecules are still being reported. Characteristically, the halogenation degree found in compounds from Laurencia is relatively high [19]. Diterpenes sesquiterpenes, triterpenes, and C15-acetogenins are the main secondary compounds of this genus [22,23] with which antimicrobial [24], antifeedant [25], antihelmintic [26,27] and cytotoxic [28,29] properties are generally associated.
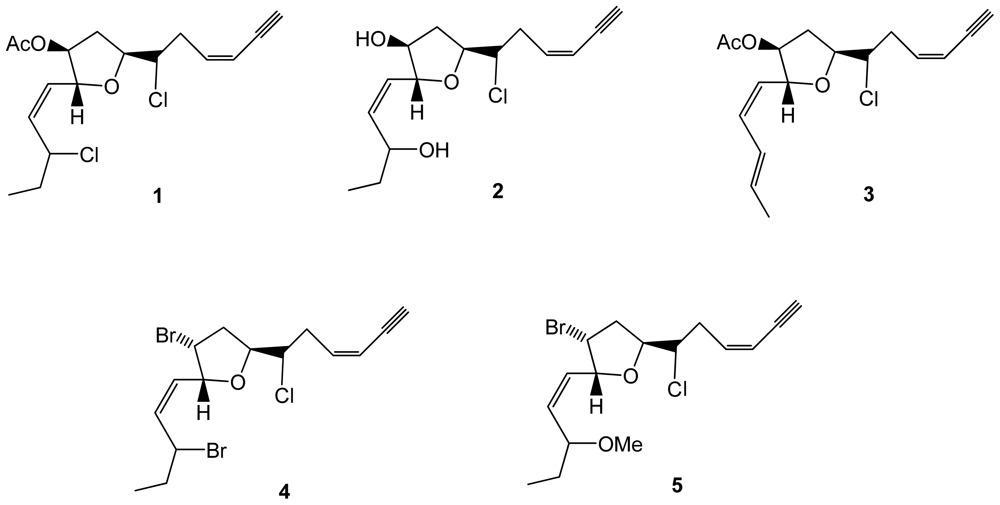
Five new chlorinated compounds, C15 acetogenin en-ynes (1–5), were obtained from Laurencia glandulifera collected from the island of Crete (Figure 1). Four of them were evaluated for their cytotoxicity toward HT-29 (colorectal adenocarcinoma), MCF-7 (mammary adenocarcinoma), PC-3 (prostate adenocarcinoma), HeLa (cervical adenocarcinoma), and A431 (epidermoid carcinoma) human tumor cell lines, but no significant activity was found [30]. The characteristic terminal cis ene-yne moiety is also present in C15 eight-membered cyclic ethers previously isolated from L. glandulifera collected on the Crete Island, which exhibited antistaphylococcal activity with minimum inhibitory concentrations (MICs) in the range of 8–256 mg/mL [31].
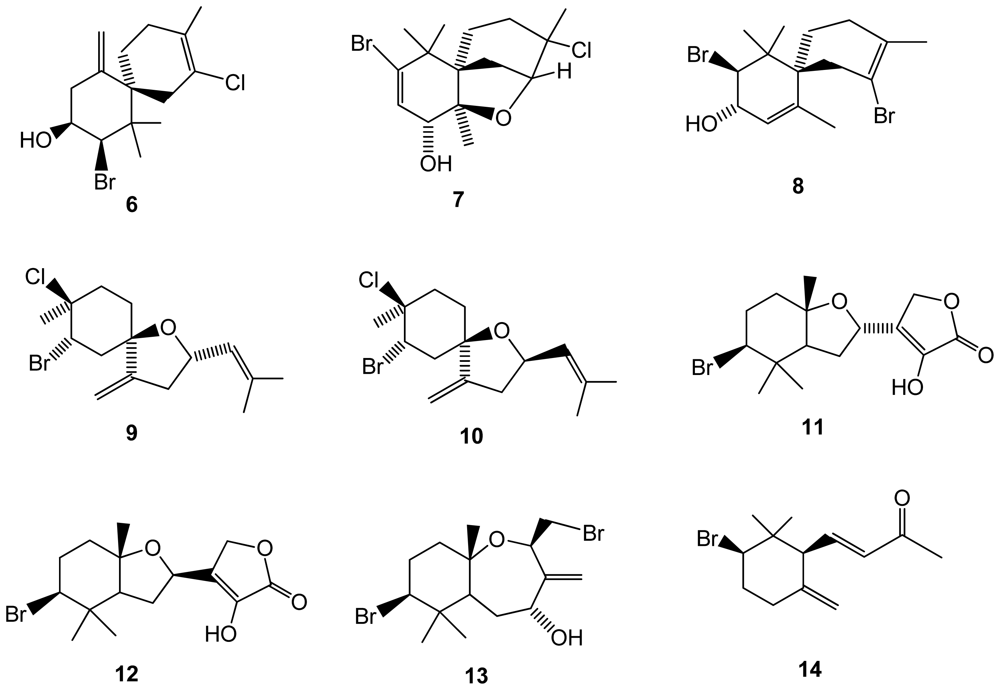
Elatol (6)(Figure 2), a halogenated sesquiterpene alcohol, commonly found in many species of Laurencia, and known for its potent antibacterial activity, was isolated for the first time in Laurencia microcladia, collected in the Southern Brazilian coast [32]. Previous analysis of anti-herbivory proprieties of metabolites from Laurencia species (including elatol) have been conducted but no investigation had been reported for Laurencia microcladia. Elatol anti-herbivory properties were investigated using the black sea urchin Echinometra lucunter, but it was found that this species can tolerate elatol, at least in the concentrations tested [32]. However, this compound was able to deter feeding of Thalassia by reef fishes with a reduced loss of Thalassia by 60% and was shown to be a very efficient deterrent against the sea urchin Diadema antillarum, reducing grazing by 86% [11]. However, anti-herbivory proprieties could not be detected by testing the compound on the black sea urchin Echinometra lucunter.
Laurencia saitoi, with exceptionally few reported halogenated compounds [25] in the past, was found to produce four novel halogenated sesquiterpenes: 10-bromo-3-chloro-2,7-epoxychamigr-9-en- 8-ol (7), 2,10-dibromochamigra-2,7-dien-9-ol (8), (9S)-2-bromo-3-chloro-6,9-epoxybisabola-7(14),10-diene (9), and (9R)-2-bromo-3-chloro-6,9-epoxybisabola-7(14),10-diene (10)[33](Figure 2). In addition, other brominated compounds were already reported for different macroalgal species, namely aplysistatin, a well known antileukemic agent [34,35], 5-acetoxypalisadin B, palisadin A, palisadin B belonging to a series of antimicrobial compounds, and 2,3,5,6-tetrabromoindole [36]. Cytotoxicity of the isolated compounds was evaluated by the MTT method, all the tested compounds being found inactive [36]. New brominated sesquiterpenes and a norsesquiterpene were also reported from the same species [37], namely 2-hydroxyluzofuranone (11), 2-hydroxyluzofuranone B (12), 4-hydroxypalisadin C (13), and 2-bromo-γ-ionone (14)(Figure 2).
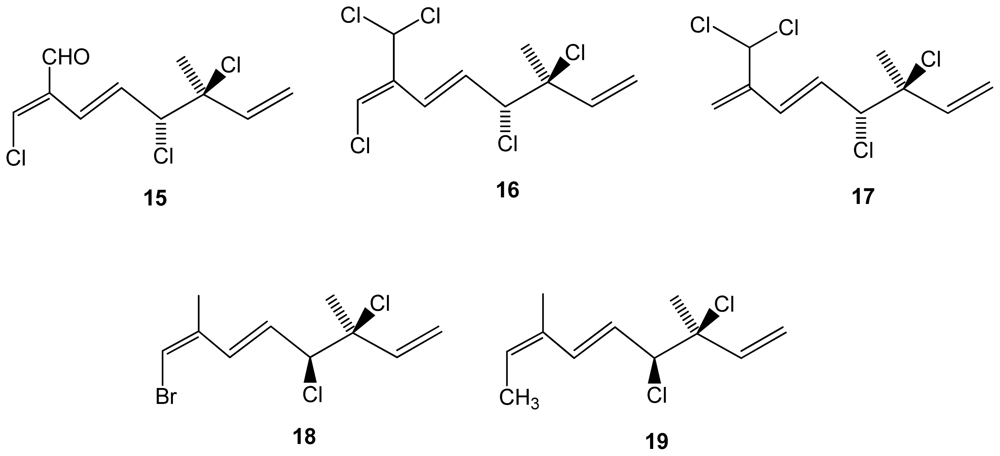
Other red algal genera have also been screened and interesting findings were recently reported. From the chemical-rich genus Plocamium, a South African red algae Plocamium cornutum yielded five halogenated monoterpenes (15–19) from which two are new (compounds 18–19)(Figure 3). The antiplasmodial activity against the chloroquine sensitive strain of the most frequent and deadly human malaria parasite Plasmodium falciparum was tested. Although the compounds were significantly less active than standard chloroquine, compounds 16 and 17 containing 7-dichloromethyl moiety were the most active ones (IC50 = 16 and 17 μM, respectively), followed by compound 15 (IC50 = 27 Mm), which contains an aldehyde functional group at this position, while the new compounds were essentially inactive [38].
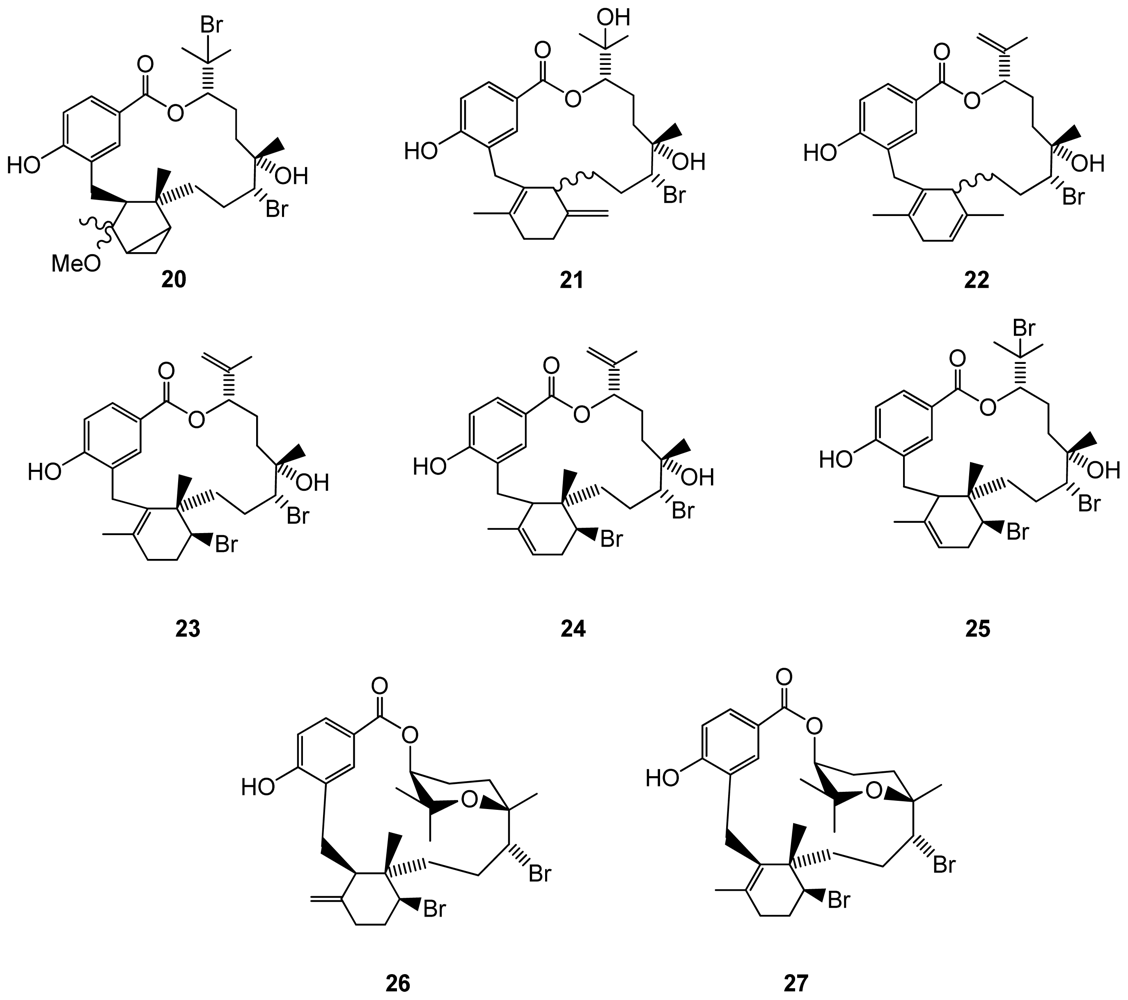
Another red algal species, the Fijian Callophycus serratus, was found to produce eight bromophycolides J–Q (20–27)(Figure 4), which exhibited IC50 values against Plasmodium falciparum in the low micromolar range (44 for compound 22, 0.5 for 23, 1.4 for 24, 25 and 27)[39]. Antimalarian activity appeared to be associated with the presence of a macrolide motif in the chemical structure of these molecules. Antibacterial assays were performed using methicillin-resistant Staphylococcus aureus and vancoymcin-resistant Enterococcus faecium as test pathogens. The macrolides were also evaluated against a panel of 12 tumor cell lines including breast, colon, lung, prostate, and ovarian cancer cells. Of the eight tested compounds, bromophycolides 26 and 27 exhibited the most potent antibacterial activity against S. aureus and E. faecium, suggesting that conformational rigidity and/or hydrophobicity conferred by the tetrahydropyran system contributes to antibacterial activity. While all tested bromophycolides exhibited moderate antineoplastic activity, only 24 displayed some cell line selectivity, with an IC50 of 1.5 μM against the breast tumor cell line. Interestingly, while 24 demonstrated cancer cell line selectivity, its regioisomer 23 was quite active against all cancer cell lines tested (IC50’s 2.1–7.2 μM). Bromophycolide 27 was the most potent C. serratus natural product evaluated but showed little cell line selectivity.
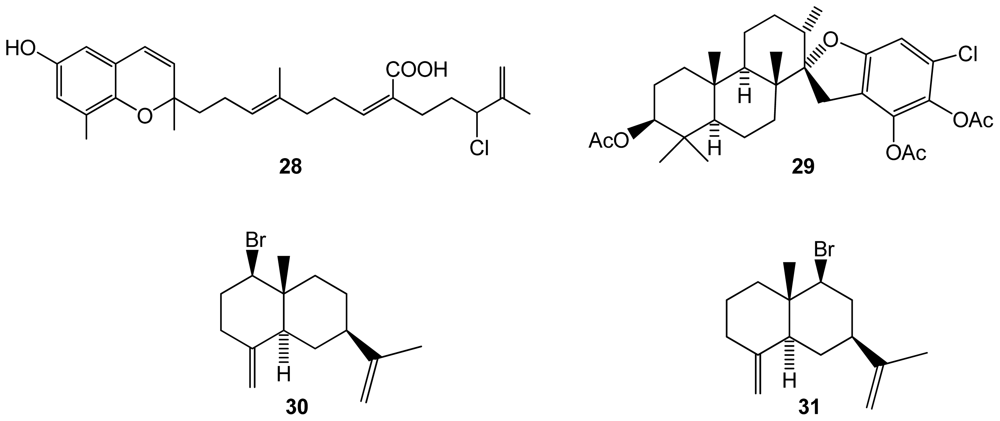
Halogenated metabolites from brown algae, reported as unusual for this algal group [40], have been found to be mainly terpenes. However, some new molecules were reported for brown algae in 2009. A new halogenated meroditerpenoid, fallachromenoic acid (28)(Figure 5), was isolated from the southern Australian brown algae Sargassum fallax, and displayed antitumor activities against a P388 Murine Leukaemia cell line [41]. Although there were no previous reports of halogenated compounds in the Stypopodium genus, Areche et al. [40] found an unusual 40-chlorostypotriol triacetate (29)(Figure 5) in Stypopodium flabelliforme, from which biological properties are still unknown. Another brown algal species, Dictyopteris divaricata, was found to produce two new brominated selinane sesquiterpenes, 1-bromoselin-4(14),11-diene (30) and 9-bromoselin-4(14),11-diene (31)[42](Figure 5).
No reports associated with green algae were found in the selected bibliography from 2009.
3. Cyanobacteria
Marine phytoplankton has been much less investigated than macroalgae although marine microorganisms are, in general, increasingly considered successful sources of natural products. Earlier reviews and reports on marine natural products including phytoplankton refer mainly to cyanobacteria and dinoflagellates [9,18,43,44] as sources of natural compounds.
Selected examples of recently discovered halogenated molecules produced by marine phytoplankton, from 2009 literature, exhibited the same trend. Among phytoplankton major groups, cyanobacteria have come forward as one of the most promising groups of microorganisms for the isolation of interesting new natural halogenated compounds [45,46]. A wide range of compounds with different properties is produced by these blue-green algae [44]. For example, many of these compounds were found to be anticancer agents or powerful neurotoxins performing either as blockers or activators of the eukaryotic voltage-gated sodium (Nav) channels [44].
Cyanobacterial toxins have received increased research effort in recent years [47–49] because these molecules are among the most found hazardous substances in surface waters and occurrence of harmful cyanobacterial bloom events is rising worldwide [50].
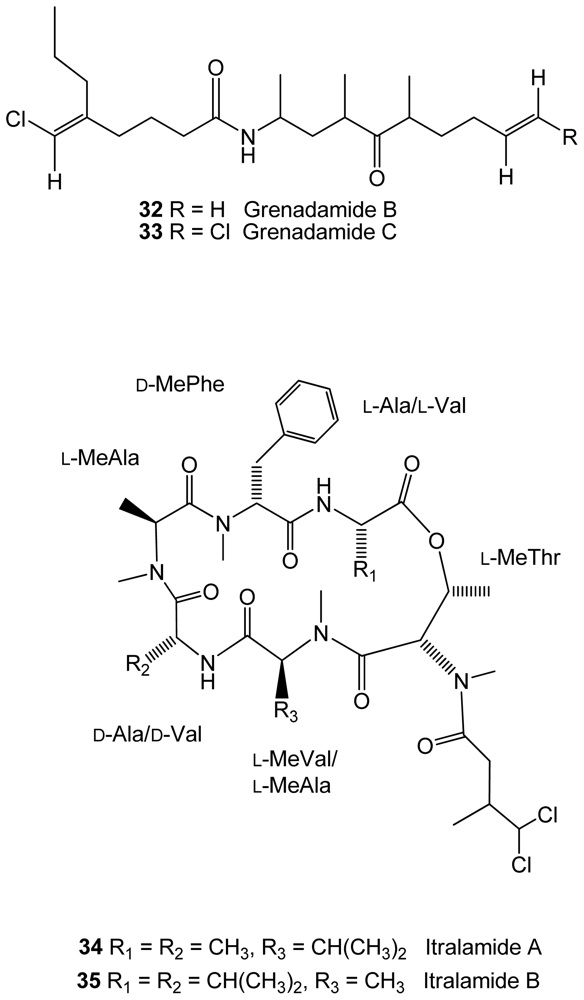
The cyanobacteria Lyngbya majuscula, from an Eastern Caribbean collection, was found to produce two new halogenated fatty acid amides, grenadamides B and C (32–33), and two new depsipeptides, itralamides A and B (34–35)[51](Figure 6). The first two compounds displayed marginal activity against the beet armyworm (Spodoptera exigua).
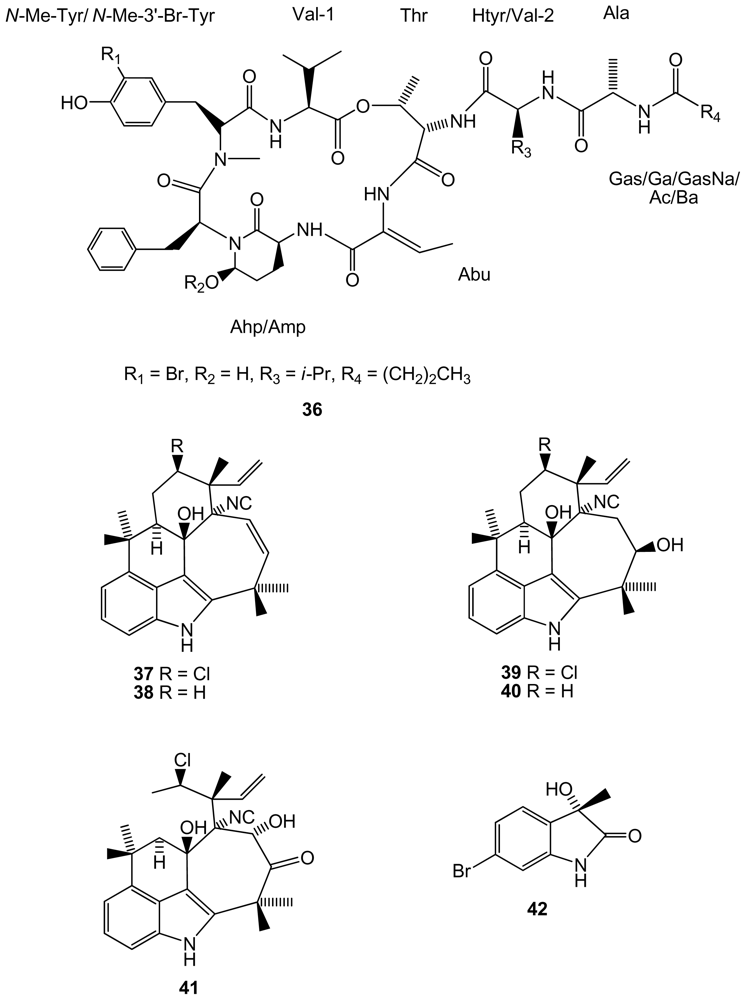
From the same genus, a new brominated compound, lyngbyastatin 10 (36)(Figure 7), closely related to lyngbyastatins, was found in Lyngbya semiplena, collected in Tumon Bay (Guam, Pacific ocean), inhibiting porcine pancreatic elastase [52]. Another cyanobacteria, Fischerella ambigua, obtained from a culture, produces five newly found antibacterial chlorinated molecules, ambiguine K–O isonitriles (37–41)[53](Figure 7). Ambiguine K and M isonitriles showed the most potent activity against Mycobacterium tuberculosis, the causal agent of most cases of tuberculosis. Ambiguine A isonitrile was the most active one against Bacillus anthracis, the bacterium that causes anthrax. A new brominated indole alkaloid, named as bromoanaindolone, (IUPAC name: 6-bromo-3-hydroxy-3-methyl-indol-2-one)(42)(Figure 7), yielded by the cyanobacteria Anabaena constricta [54], showed antibacterial activity against the Gram-positive species Bacillus cereus, the bacterium that causes foodborne disease commonly known as food poisoning. Bromoanaindolone also displayed anticyanobacterial activity against the filamentous species Arthrospira laxissima and Nostoc carneum, and also against the unicellular species Chroococcus minutus, Synechocystis aquatilis and Synechococcus sp., which indicates the allelopathic potential of this compound.
4. Limitations and Possible Solutions to Detection of Bioactivity
The fact that some of the newly found halogenated compounds show minor or no activity at all against a specific target does not exclude the possibility of other hidden unidentified active biological effects. Additionally, the supposedly absence of bioresponse may be related to limitations in the analytical chemistry methods employed. Bioassay-guided fractionation is the iterative approach traditionally applied to disclose bioactive natural compounds [55,56]. Difficulties in perception of bioactivity may be linked to compound instability, caused by fractionation and isolation from the natural cellular environment or simply because compounds may act synergistically or more than one compound may be associated to a specific property. This is particularly true for macroalgae and microalgae which are complex compound matrices [57]. Metabolite-oriented approaches, namely metabolite profiling and metabolomics, may supplement bioassay-guided fractionation [58]. These two approaches are focused on the chemical properties of the metabolome, which includes the complete set of metabolites of an organism (metabolomics), or of a limited group of pre-defined metabolites (metabolite profiling)[59]. In the latter approach, pre-defined metabolites are selected either according to the class they belong to (organic phosphates, carbohydrates, etc.) or to an associated specific pathway. Both metabolite profiling and metabolomics allow all compounds to be simultaneously measured, so that detection of unstable compounds is facilitated and synergistic effects detected [59]. Also, the influence of a particular condition on the organism physiology can be revealed through qualitative and quantitative differences in the metabolic profiles obtained [60,61]. This could be a valuable tool for the assessment of effects of environmental changes on the community structure and organism physiology. However, useful metabolomics may be used for a more thorough and complete detection of metabolites, limitations subsist when applied to chemical ecology, due to the difficulty in establishing a link between one or more specific metabolites and an ecological role [58].
Other techniques have been applied for the study of halogenated compounds in marine algae. Desorption electrospray ionization mass spectrometry (DESI-MS) and some applications, such as imaging, have very recently started to be used for natural products detection on intact algal tissue surfaces [62,63]. DESI-MS is an ambient ionization technique coupled with mass spectrometry, allowing probing the surface of solids, liquids, frozen solutions, and adsorbed gases [64]. In ambient DESI-MS, samples are kept under atmospheric pressure in open air, with minimum or no preparation at all [64]. Tissue surfaces are subject to a focused spray of charged particles of a polar solvent, which leads to the release of intact gaseous ions from surfaces into the mass spectrometer [65]. Low detection limits and high analysis speed are achieved, and innate chemical information is preserved to a maximum [64]. DESI-MS has been shown to be a sensitive and effective approach to detect algal diterpene-benzoate macrolide natural products, namely bromophycolides, directly on the surface and interior of a marine red alga, Callophycus serratus [62]. Reactive DESI approaches were investigated to improve ionization efficiency and detection limits. Different polar solvent compositions, tested directly on the algal surfaces, were demonstrated to enhance sensitivity for discovery of various bromophycolides, which would have otherwise been undetected. Imaging, one of DESI-MS possible applications, has been shown to provide an exceptional capability to map secondary metabolites to distinct algal surface sites [63]. Bromophycolides were shown to be non homogenously distributed across surfaces but instead associated with distinct patches, again in the red algae Callophycus serratus. DESI-MS imaging appears to be a powerful tool for the investigation of the function of surface-associated natural products in ecological interactions, as spatially-resolved measurements with lateral resolutions in the hundreds of micrometers are made possible. These studies [62,63] illustrate the potential of DESI-MS and its applications, such as imaging, in understanding chemically-mediated biological processes in macroalgae.
5. Ecological Role
Micro and macroalgal survival and success in the marine environment reflect to a great extent the many strategies these organisms have had to employ throughout their evolutionary history to adapt to a hostile and high resource-demanding milieu. Extreme competition for space, light and nutrients has shaped the organisms’ physiology, resulting in a complex array of biologically active metabolites characteristic of each species. Many of these secondary metabolites have been shown to support the defense mechanisms developed by marine algae [66–69]. Secondary metabolites are now suggested to be responsible for marine biodiversity on the genetic, species, and ecosystem levels [70,71].
Chemical ecology, which tackles the role of natural compounds in the interactions between organisms and also between organisms and their environment, has already a few decades of investigation dedicated to marine systems [46]. Historically, interest in natural products from marine algae was initially driven to meet pressing biomedical requirements for new drugs against fungal [72], parasitic [73], bacterial [74], and viral [75] diseases, which were either used in their natural form or as templates for synthesis and further chemical structure modification [46]. For this purpose, macroalgae from tropical areas were heavily screened and many new halogenated compounds were found in the last decade [18]. Apart from potent and original therapeutic agents for infectious diseases and cancer, among many other disorders, natural products isolated from marine algae were also found to clearly control interactions between organisms [11–13], and therefore influence population structure, communities organization, and ecosystem function [14–17]. Biodiversity in marine systems may be a result of chemical diversity [68]. Presently, specific ecological aspects such as chemical sensing of the environment by algae, intraspecific signaling, allelopathy, predator-prey and host-parasite interactions, and bioaccumulation and transfer of toxins within food webs have been addressed on recent research on the ecology of natural compounds from marine algae. Excellent reviews and essays on marine chemical ecology covering years prior to 2009 have been published (see [3,17,68,76–78]), however, with no particular focus on halogenated compounds. Interactions between algae and herbivorous invertebrates or describing allelopathic effects have received most of the attention. These ecological interactions continue to be predominantly documented from an ecological perspective [79,80]; the chemical identification of compounds responsible for the found ecological relationships was overlooked during 2009. A multidisciplinary approach including organic chemistry, biology, and ecology components is required for the in depth elucidation of the ecological relationships in the near future. So far, the ecological and the chemical approaches rarely coincide in the same study; the cellular and molecular bases for these interactions remain disregarded. In fact, to the best of our knowledge, an ecological study on marine macroalgae, including elucidation of chemical structure of molecules responsible for ecological interactions, was found during 2009. A direct fine-scale evaluation of bromophycolides and callophycoic acids was conducted on red macroalga Callophycus serratus surfaces in order to evaluate whether these natural compounds had a role in surface-mediated defense against pathogenic microbes [63]. These novel compounds had been reported for the first time in C. serratus [39], represent the largest group of algal antifungal compounds described to date, and were shown to inhibit growth of Lindra thalassiae, a marine pathogenic fungus [63]. DESI-MS imaging revealed bromophycolides among heterogeneous patches on algal surfaces and within internal algal tissues. The bromophycolide concentrations found on these surface patches were sufficient suppress growth of L. thalassia, indicating that probably these compounds are kept internally and released at sparsely distributed surface sites. The above cited study [63] provides the first direct evidence for localization of chemical signals with spatial resolution <200 μm on biological surfaces in concentrations sufficient for targeted antimicrobial defense, highlighting the potential of DESI-MS imaging for the understanding of small-scale ecological interactions.
6. Present Trends
A lot still remains to be done concerning research on the role of marine natural halogenated compounds: (i) isolation and characterization of new halogenated compounds (ii) screening untested micro and macroalgal species (iii) improving assessment in the detection of bioactivities, fully exploring potential of metabolite-oriented approaches, (iv) testing biomedical and ecological relevance of novel and known chemical natural compounds, (v) ensuring reliable taxonomic identification of investigated species, (vi) biogeographical assessing of micro- and macroalgae with relevant compounds, and (vii) developing appropriate strategies for data management.
From a purely ecological point of view, success toward the in-depth understanding of marine ecological interactions is very much dependent on a multidisciplinary approach, relying on adequate bioactivity detection methodologies. Chemical metabolite-oriented approaches may prove to be reliable tools helping to elucidate compound’s biological properties, which may not be detectable in any other way. Ecological studies may just benefit from an interactive and close combination of chemical methodologies with ecological experiments.
Acknowledgements
This study was funded by “Fundação para a Ciência e Tecnologia” grant SFRH/BPD/50348/2009 to M.T. Cabrita.
References
- Cimino, G; Ghiselin, MT. McClintock, JB, Baker, BJ, Eds.; Marine natural products chemistry as an evolutionary narrative. In Marine Chemical Ecology; CRC Press: Boca Raton, FL, USA, 2001; pp. 115–154. [Google Scholar]
- Cannell, RJP. Cannell, RPJ, Ed.; How to approach the isolation of a natural product. In Natural Products Isolation, Methods in Biotechnology; Humana Press: Totowa, NJ, USA, 1998; Volume 4, pp. 1–51. [Google Scholar]
- Ianora, A; Boersma, M; Casotti, R; Fontana, A; Harder, J; Hoffmann, F; Pavia, H; Potin, P; Poulet, SA; Toth, G. New trends in marine chemical ecology. Estuaries Coasts 2006, 29, 531–551. [Google Scholar]
- Butler, A; Sandy, M. Mechanistic considerations of halogenating enzymes. Nature 2009, 460, 848–854. [Google Scholar]
- Neumann, CS; Fujimori, DG; Walsh, CT. Halogenation strategies in natural product biosynthesis. Chem. Biol 2008, 15, 99–109. [Google Scholar]
- Küpper, FC; Schweigert, N; Ar Gall, E; Legendre, J-M; Vilter, H; Kloareg, B. Iodine uptake in Laminariales involves extracellular, haloperoxidase-mediated oxidation of iodide. Planta 1998, 207, 163–167. [Google Scholar]
- Harper, MK; Bugni, TS; Copp, BR; James, RD; Lindsay, BS; Richardson, AD; Schnabel, PC; Tasdemir, D; Van Wagoner, RM; Verbitzki, SM; Ireland, CM. McClintock, JB, Baker, BJ, Eds.; Introduction to the chemical ecology of marine natural products. In Marine Chemical Ecology; CRC: Boca Raton, FL, USA, 2001; pp. 3–71. [Google Scholar]
- Renner, MK; Jensen, PR; Fenical, W. Neomangicols: structures and absolute stereochemistries of unprecedented halogenated sesterterpenes from a marine fungus of the genus Fusarium. J. Org. Chem 1998, 63, 8346–8354. [Google Scholar]
- Blunt, JW; Copp, BR; Hu, WP; Munro, MHG; Northcote, PT; Prinsep, MR. Marine natural products. Nat. Prod. Rep 2009, 26, 170–244. [Google Scholar]
- Fenical, W; Norris, JN. Chemotaxonomy in marine algae: chemical separation of some Laurencia species (Rhodophyta) from the Gulf of California). J. Phycol 1975, 11, 104–108. [Google Scholar]
- Hay, ME; Fenical, W; Gustafson, K. Chemical defense against diverse coral-reef herbivores. Ecology 1987, 68, 1581–1591. [Google Scholar]
- Vairappan, CS. Potent antibacterial activity of halogenated metabolites from Malaysian red algae, Laurencia majuscula (Rhodomelaceae, Ceramiales). Biomol. Eng 2003, 20, 255–259. [Google Scholar]
- Vairappan, CS; Ang, MY; Ong, CY; Phang, SM. Biologically active polybrominated indoles in the red alga Laurencia similis from the coastal waters of Sabah (Rhodomelaceae, Ceramiales). Malaysian J. Sci 2004, 23, 119–126. [Google Scholar]
- Paul, NA; de Nys, R; Steinberg, PD. Seaweed–herbivore interactions at a small scale: direct tests of feeding deterrence by filamentous algae. Mar. Ecol. Prog. Ser 2006, 323, 1–9. [Google Scholar]
- Harlin, MM; Rice, EL. Allelochemistry in marine macroalgae. Crit. Rev. Plant Sci 1987, 3, 237–249. [Google Scholar]
- Bolser, RC; Hay, ME. Are tropical plants better defended? Palatability and defenses of temperate vs. tropical seaweeds. Ecology 1996, 77, 2269–2286. [Google Scholar]
- Hay, ME. Marine chemical ecology: Chemical signals and cues structure marine populations, communities, and ecosystems. Annu. Rev. Mar. Sci 2009, 1, 193–212. [Google Scholar]
- Blunt, JW; Copp, BR; Hu, W-P; Munro, MHG; Northcote, PT; Prinsep, MR. Marine natural products. Nat. Prod. Rep 2007, 24, 31–86. [Google Scholar]
- Hill, RA. Marine natural products. Annu. Rep. Prog. Chem., Sect. B: Org. Chem 2007, 103, 125–139. [Google Scholar]
- Faulkner, DJ. Marine natural products. Nat. Prod. Rep 2001, 18, 1–49. [Google Scholar]
- Wright, AD; Goclik, E; König, GM. Three new sesquiterpenes from the red alga Laurencia perforate. J. Nat. Prod 2003, 66, 435–437. [Google Scholar]
- Erickson, KL. Scheuer, PJ, Ed.; Constituents of Laurencia. In Marine Natural Products: Chemical and Biological Perspectives; Academic Press: New York, NY, USA, 1983; Volume 5, pp. 131–257. [Google Scholar]
- Faulkner, DJ. Marine natural products. Nat. Prod. Rep 1995, 12, 223–269. [Google Scholar]
- König, GM; Wright, AD. Laurencia rigida: Chemical investigations of its antifouling dichloromethane extract. J. Nat. Prod 1997, 60, 967–970. [Google Scholar]
- Kurata, K; Taniguchi, K; Agatsuma, Y; Suzuki, M. Diterpenoid feeding-deterrents from Laurencia saitoi. Phytochemistry 1998, 47, 363–369. [Google Scholar]
- Davyt, D; Fernandez, R; Suescun, L; Mombrú, AW; Saldaña, J; Domínguez, L; Coll, J; Fujii, MT; Do Manta, E. New sesquiterpene derivatives from the red alga Laurencia scoparia. Isolation, structure determination, and anthelmintic activity. J. Nat. Prod 2001, 64, 1552–1555. [Google Scholar]
- Topcu, G; Aydogmus, Z; Imre, S; Gören, AC; Pezzuto, JM; Clement, JA; Kingston, DGI. Brominated sesquiterpenes from the red alga Laurencia obtusa. J. Nat. Prod 2003, 66, 1505–1508. [Google Scholar]
- Juagdan, EG; Kalidindi, R; Scheuer, P. Two new chamigranes from an hawaiian red alga, Laurencia cartilaginea. Tetrahedron 1997, 53, 521–528. [Google Scholar]
- Sun, J; Shi, D; Ma, M; Li, S; Wang, S; Han, L; Yang, Y; Fan, X; Shi, J; He, L. Sesquiterpenes from the red alga Laurencia tristicha. J. Nat. Prod 2005, 68, 915–919. [Google Scholar]
- Kladi, M; Vagias, C; Papazafiri, P; Brogi, S; Tafi, A; Roussis, V. Tetrahydrofuran acetogenins from Laurencia glandulifera. J. Nat. Prod 2009, 72, 190–193. [Google Scholar]
- Kladi, M; Vagias, C; Stavri, M; Rahman, M; Gibbons, S; Roussis, V. C15 acetogenins with antistaphylococcal activity from the red alga Laurencia glandulifera. Phytochem. Lett 2008, 1, 31–36. [Google Scholar]
- Lhullier, C; Donnangelo, A; Caro, M; Palermo, JA; Horta, PA; Falkenberg, M; Schenkel, EP. Isolation of elatol from Laurencia microcladia and its palatability to the sea urchin Echinometra lucunter. Biochem. Syst. Ecol 2009, 37, 254–259. [Google Scholar]
- Ji, N-Y; Li, X-M; Li, K; Wang, B-G. Halogenated sesquiterpenes from the marine red alga Laurencia saitoi (Rhodomelaceae). Helv. Chim. Acta 2009, 92, 1873–1879. [Google Scholar]
- Pettit, GR; Herald, CL; Allen, MS; Von Dreele, RB; Vanell, LD; Kao, JPY; Blake, W. The isolation and structure of aplysistatin. J. Am. Chem. Soc 1977, 99, 262–263. [Google Scholar]
- Pandey, PK. Endangered medicinal species of the Indian Ocean: radical need for conservation. Chem. Biodivers 2009, 6, 990–1001. [Google Scholar]
- Su, H; Yuan, Z; Li, J; Guo, S; Han, L; Zhu, X; Shi, D. Studies on chemical constituents of Laurencia saitoi. Zhongguo Zhong Yao Za Zhi 2009, 34, 871–874. [Google Scholar]
- Su, H; Yuan, Z-H; Li, J; Guo, S-J; Deng, L-P; Han, L-J; Zhu, X-B; Shi, D-Y. Sesquiterpenes from the marine red alga Laurencia saitoi. Helv. Chim. Acta 2009, 92, 1291–1297. [Google Scholar]
- Afolayan, AF; Mann, MGA; Lategan, CA; Smith, PJ; Bolton, JJ; Denzil, R; Beukes, DR. Antiplasmodial halogenated monoterpenes from the marine red alga Plocamium cornutum. Phytochemistry 2009, 70, 597–600. [Google Scholar]
- Lane, AL; Stout, EP; Lin, A-S; Prudhomme, J; Le Roch, K; Fairchild, CR; Franzblau, SG; Hay, ME; Aalbersberg, W; Kubanek, J. Antimalarial bromophycolides J-Q from the fijian red alga Callophycus serratus. J. Org. Chem 2009, 74, 2736–2742. [Google Scholar]
- Areche, C; San-Martín, A; Rovirosa, J; Soto-Delgado, J; Contreras, R. An unusual halogenated meroditerpenoid from Stypopodium flabelliforme: Studies by NMR spectroscopic and computational methods. Phytochemistry 2009, 70, 1315–1320. [Google Scholar]
- Reddy, P; Urban, S. Meroditerpenoids from the southern Australian marine brown alga Sargassum fallax. Phytochemistry 2009, 70, 250–255. [Google Scholar]
- Ji, N-Y; Wen, W; Li, X-M; Xue, Q-Z; Xiao, H-L; Wang, B-G. Brominated selinane sesquiterpenes from the Marine Brown Alga Dictyopteris divaricata. Mar. Drugs 2009, 7, 355–360. [Google Scholar]
- Cardozo, KHM; Guaratini, T; Barros, MP; Falcão, VR; Tonon, AP; Lopes, NP; Campos, S; Torres, MA; Anderson, O; Souza, AO; Colepicolo, P; Pinto, E. Metabolites from algae with economical impact. Comp. Biochem. Physiol. C: Toxicol. Pharmacol 2007, 146, 60–78. [Google Scholar]
- Tan, LT. Bioactive natural products from marine cyanobacteria for drug discovery. Phytochemistry 2007, 68, 954–979. [Google Scholar]
- Borowitzka, MA. Microalgae as sources of pharmaceuticals and other biologically active compounds. J. Appl. Phycol 1995, 7, 3–15. [Google Scholar]
- Burja, AM; Banaigs, B; Abou-Mansour, E; Burgess, JG; Wright, PC. Marine cyanobacteria-a prolific source of natural products. Tetrahedron 2001, 57, 9347–9377. [Google Scholar]
- Kwan, JC; Rocca, JR; Abboud, KA; Paul, VJ; Luesch, H. Total structure determination of grassypeptolide, a new marine cyanobacterial cytotoxin. Org. Lett 2008, 10, 789–792. [Google Scholar]
- Luesch, H; Yoshida, WY; Moore, RE; Paul, VJ; Corbett, TH. Total structure determination of apratoxin A, a potent novel cytotoxin from the marine cyanobacterium Lyngbya majuscula. J. Am. Chem. Soc 2001, 123, 5418–5423. [Google Scholar]
- Taori, K; Paul, VJ; Luesch, H. Structure and activity of largazole, a potent antiproliferative agent from the Floridian marine cyanobacterium Symploca sp. J. Am. Chem. Soc 2008, 130, 1806–1807. [Google Scholar]
- Paerl, HW; Huisman, J. Climate change: a catalyst for global expansion of harmful cyanobacterial blooms. Environ. Microbiol. Rep 2009, 1, 27–37. [Google Scholar]
- Jiménez, JI; Vansach, T; Yoshida, WY; Sakamoto, B; Pörzgen, P; Horgen, FD. Halogenated fatty acid amides and cyclic depsipeptides from an eastern caribbean collection of the cyanobacterium Lyngbya majuscule. J. Nat. Prod 2009, 72, 1573–1578. [Google Scholar]
- Kwan, JC; Taori, K; Paul, VJ; Luesch, H. Lyngbyastatins 8–10, elastase inhibitors with cyclic depsipeptide scaffolds isolated from the marine cyanobacterium Lyngbya semiplena. Mar. Drugs 2009, 7, 528–538. [Google Scholar]
- Mo, S; Krunic, A; Chlipala, G; Orjala, J. Antimicrobial ambiguine isonitriles from the cyanobacterium Fischerella ambigua. J. Nat. Prod 2009, 72, 894–899. [Google Scholar]
- Volk, R-B; Girreser, U; Al-Refai, M; Laatsch, H. Bromoanaindolone, a novel antimicrobial exometabolite from the cyanobacterium Anabaena constricta. Nat. Prod. Res 2009, 23, 607–612. [Google Scholar]
- Hayes, KF; Millar, JG. Methods in Chemical Ecology: Bioassay Methods; Kluwer Academic: Norwell, MA, USA, 1998. [Google Scholar]
- Millar, JG; Hayes, KF. Methods in Chemical Ecology: Chemical Methods; Kluwer Academic: Norwell, MA, USA, 1998. [Google Scholar]
- Plaza, M; Santoyo, S; Jaime, L; García-Blairsy Reina, G; Herrero, M; Señoráns, FJ; Ibáñez, E. Screening for bioactive compounds from algae. J. Pharm. Biomed. Anal 2010, 51, 450–455. [Google Scholar]
- Prince, EK; Pohnert, G. Searching for signals in the noise: metabolomics in chemical ecology. Anal. Bioanal. Chem 2010, 396, 193–197. [Google Scholar]
- Fiehn, O. Combining genomics, metabolome analysis, and biochemical modeling to understand metabolic networks. Compar. Funct. Genom 2001, 2, 155–168. [Google Scholar]
- Bundy, JG; Davey, MP; Viant, MR. Environmental metabolomics: a critical review and future perspectives. Metabolomics 2009, 5, 3–21. [Google Scholar]
- Barofsky, A; Vidoudez, C; Pohnert, G. Metabolic profiling reveals growth stage variability in diatom exudates. Limnol. Oceanogr. Meth 2009, 7, 382–390. [Google Scholar]
- Nyadong, L; Hohenstein, EG; Galhena, AS; Lane, AL; Kubanek, J; Sherrill, CD; Fernandez, FM. Reactive desorption electrospray ionization mass spectrometry (DESI-MS) of natural products of a marine alga. Anal. Bioanal. Chem 2009, 394, 245–254. [Google Scholar]
- Lane, AL; Nyadong, L; Galhena, AS; Shearerb, TL; Stout, EP; Parry, RM; Kwasnik, M; Wang, MD; Hay, ME; Fernandez, FM; Kubanek, J. Desorption electrospray ionization mass spectrometry reveals surface-mediated antifungal chemical defense of a tropical seaweed. Proc. Natl. Acad. Sci. USA 2009, 106, 7314–7319. [Google Scholar]
- Wiseman, JM; Laughlin, BC. Desorption electrospray ionization (DESI) mass spectrometry: a brief introduction and overview. Curr. Sep. Drug Dev 2007, 22, 11–14. [Google Scholar]
- Cooks, RG; Ouyang, Z; Takats, Z; Wiseman, JM. Ambient mass spectrometry. Science 2006, 311, 1566–1570. [Google Scholar]
- Lobban, CS; Harrison, PJ. Seaweed Ecology and Physiology; Cambridge University Press: New York, NY, USA, 1994. [Google Scholar]
- Amsler, CD; Iken, K; McClintock, JB; Baker, BJ. Defenses of polar macroalgae against herbivores and biofoulers. Bot. Mar 2009, 52, 535–545. [Google Scholar]
- Hay, ME; Fenical, W. Chemical ecology and marine biodiversity: insights and products from the sea. Oceanography 1996, 9, 10–19. [Google Scholar]
- Lane, AL; Nyadong, L; Galhena, AS; Shearer, TL; Stout, EP; Parry, RM; Kwasnik, M; Wang, MD; Hay, ME; Fernandez, FM; Kubanek, J. Desorption electrospray ionization mass spectrometry reveals surface-mediated antifungal chemical defense of a tropical seaweed. Proc. Natl. Acad. Sci. USA 2009, 106, 7314–7319. [Google Scholar]
- Norse, EA. Global Marine Biological Diversity: A Strategy for Building Decision into Decision Making; Island Press: Washington DC, USA, 1993. [Google Scholar]
- Paul, VJ; Arthur, KE; Ritson-Williams, R; Ross, C; Sharp, K. Chemical defenses: from compounds to communities. Biol. Bull 2007, 213, 226–251. [Google Scholar]
- Nagai, H; Murata, M; Torigoe, K; Satake, M; Yasumoto, T. Gambieric acids: new potent antifungal substances with unprecedented polyether structures from a marine dinoflagellate Gambierdiscus toxicus. J. Org. Chem 1992, 57, 5448–5453. [Google Scholar]
- Capon, RJ; Barrow, RA; Rochfort, S; Jobling, M; Skene, C. Marine nematodes: tetrahydrofurans from a Southern Australian brown alga Notheia anomala. Tetrahedron 1998, 54, 2227–2242. [Google Scholar]
- Etahiri, S; Bultel-Ponce, V; Caux, C; Guyot, M. New bromoditerpenes from the red alga Sphaerococcus coronopifolius. J. Nat. Prod 2001, 64, 1024–1027. [Google Scholar]
- Sakemi, S; Higa, T; Jefford, CW; Bernardinelli, G. Venustatriol, a new anti-viral triterpenes tetracyclic ether from Laurencia venusta. Tetrahedron Lett 1986, 27, 4287–4290. [Google Scholar]
- Pohnert, G; Steinke, M; Tollrian, R. Chemical cues, defense metabolites and the shaping of pelagic interspecific interactions. Trends Ecol. Evol 2007, 22, 198–204. [Google Scholar]
- Amsler, CD. Amsler, CD, Ed.; Algal sensory chemical ecology. In Algal Chemical Ecology; Springer: Berlin, Heidelberg, Germany, 2008; pp. 297–309. [Google Scholar]
- Poulson, KL; Sieg, RD; Kubanek, J. Chemical ecology of the marine plankton. Nat. Prod. Rep 2009, 26, 729–745. [Google Scholar]
- Prince, EK; Myers, TL; Naar, J; Kubanek, J. Competing phytoplankton undermines allelopathy of a bloom-forming dinoflagellate. Proc. R. Soc. B 2008, 275, 2733–2741. [Google Scholar]
- Tameishi, M; Yamasaki, Y; Nagasoe, S; Shimasaki, Y; Oshima, Y; Honjo, T. Allelopathic effects of the dinophyte Prorocentrum minimum on the growth of the bacillariophyte Skeletonema costatum. Harmful Algae 2009, 8, 421–429. [Google Scholar]
Figure 1.
Structures of compounds 1–5 isolated from Laurencia glandulifera.
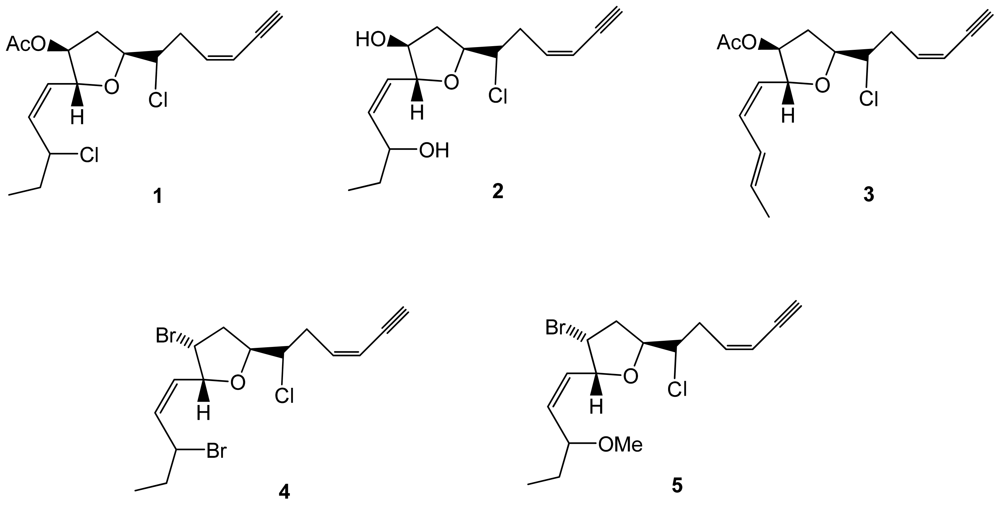
Figure 2.
Structures of halogenated compounds 6–14 isolated from Laurencia saitoi.
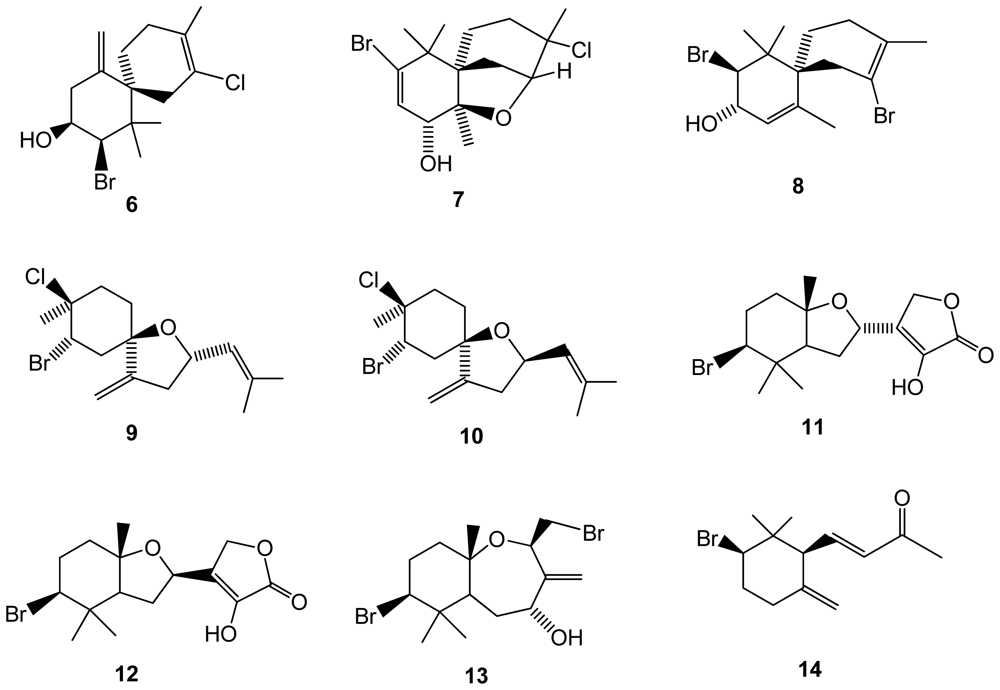
Figure 3.
Structures of monoterpenes 15–19 isolated from Plocamium cornutum.
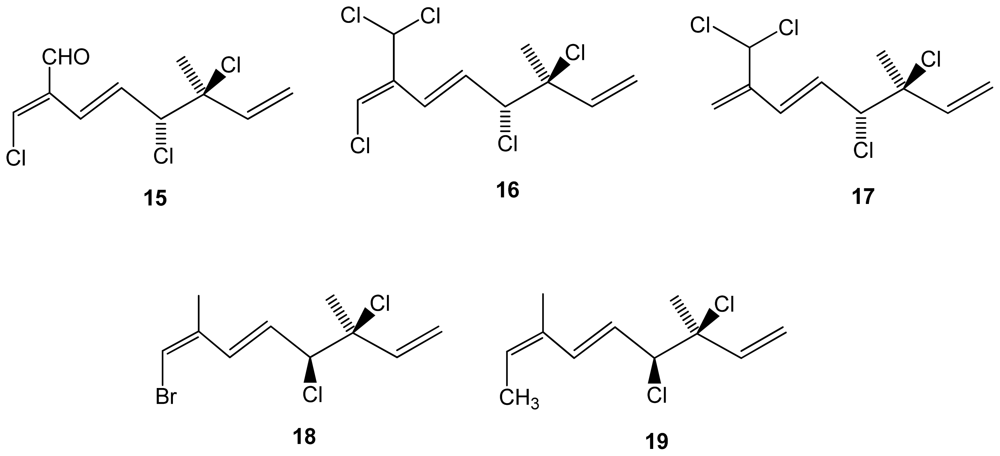
Figure 4.
Structures of bromophycolides 20–27 from Callophycus serratus.
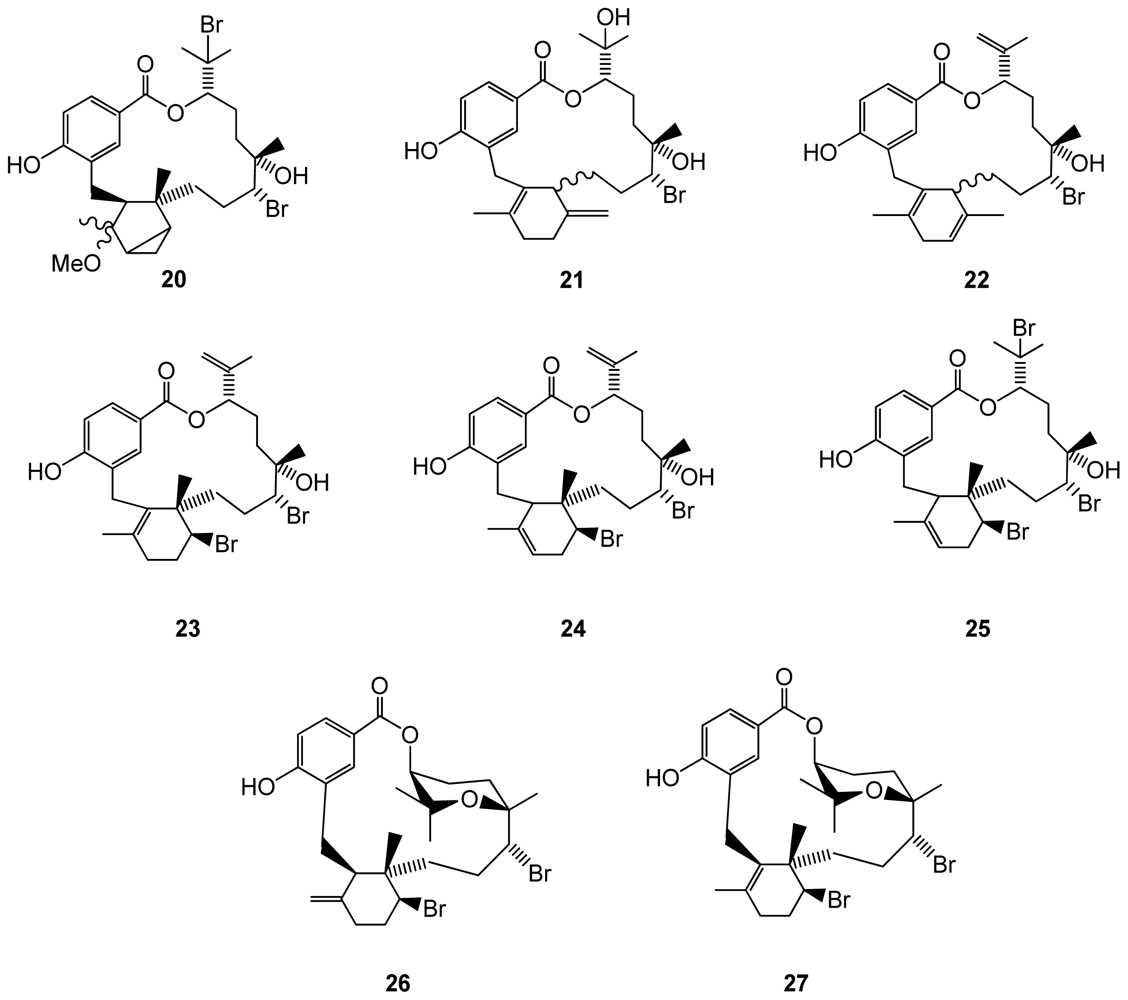
Figure 5.
Structures of compounds 28–31 isolated from brown algae.
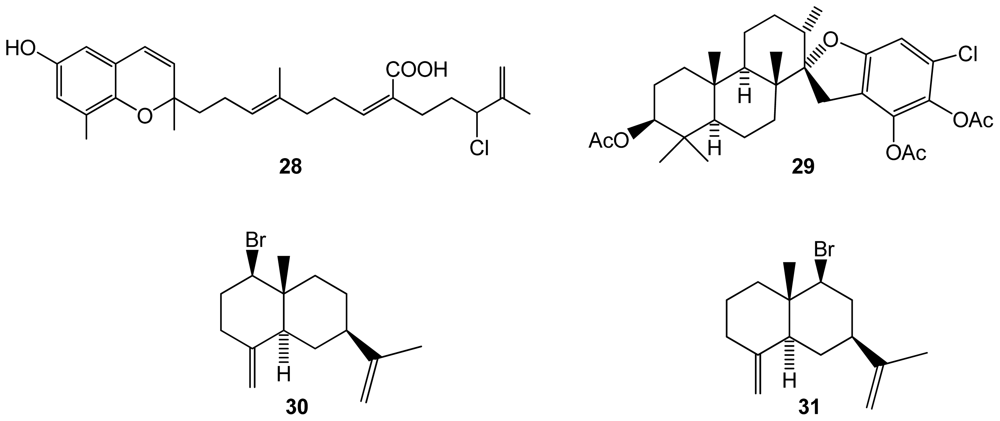
Figure 6.
Structures of compounds 32–35 isolated from the cyanobacteria Lyngbya majuscula.
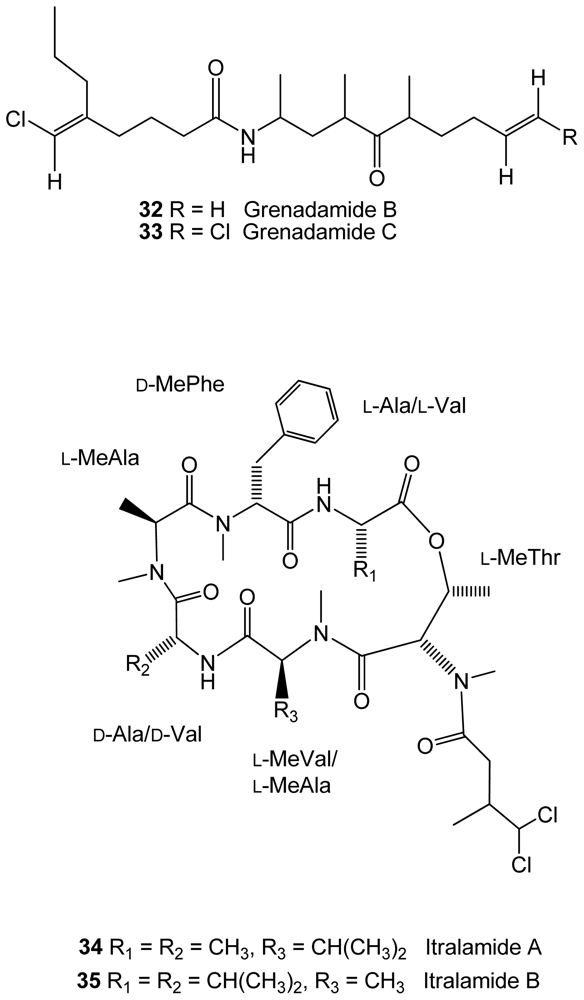
Figure 7.
Structures of compounds 36–42 isolated from cyanobacteria.
© 2010 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
MDPI and ACS Style
Cabrita, M.T.; Vale, C.; Rauter, A.P. Halogenated Compounds from Marine Algae. Mar. Drugs 2010, 8, 2301-2317. https://doi.org/10.3390/md8082301
AMA Style
Cabrita MT, Vale C, Rauter AP. Halogenated Compounds from Marine Algae. Marine Drugs. 2010; 8(8):2301-2317. https://doi.org/10.3390/md8082301
Chicago/Turabian StyleCabrita, Maria Teresa, Carlos Vale, and Amélia Pilar Rauter. 2010. "Halogenated Compounds from Marine Algae" Marine Drugs 8, no. 8: 2301-2317. https://doi.org/10.3390/md8082301