The Halogenated Metabolism of Brown Algae (Phaeophyta), Its Biological Importance and Its Environmental Significance

Abstract
:1. Introduction
2. Halogenated Metabolites of Brown Algae
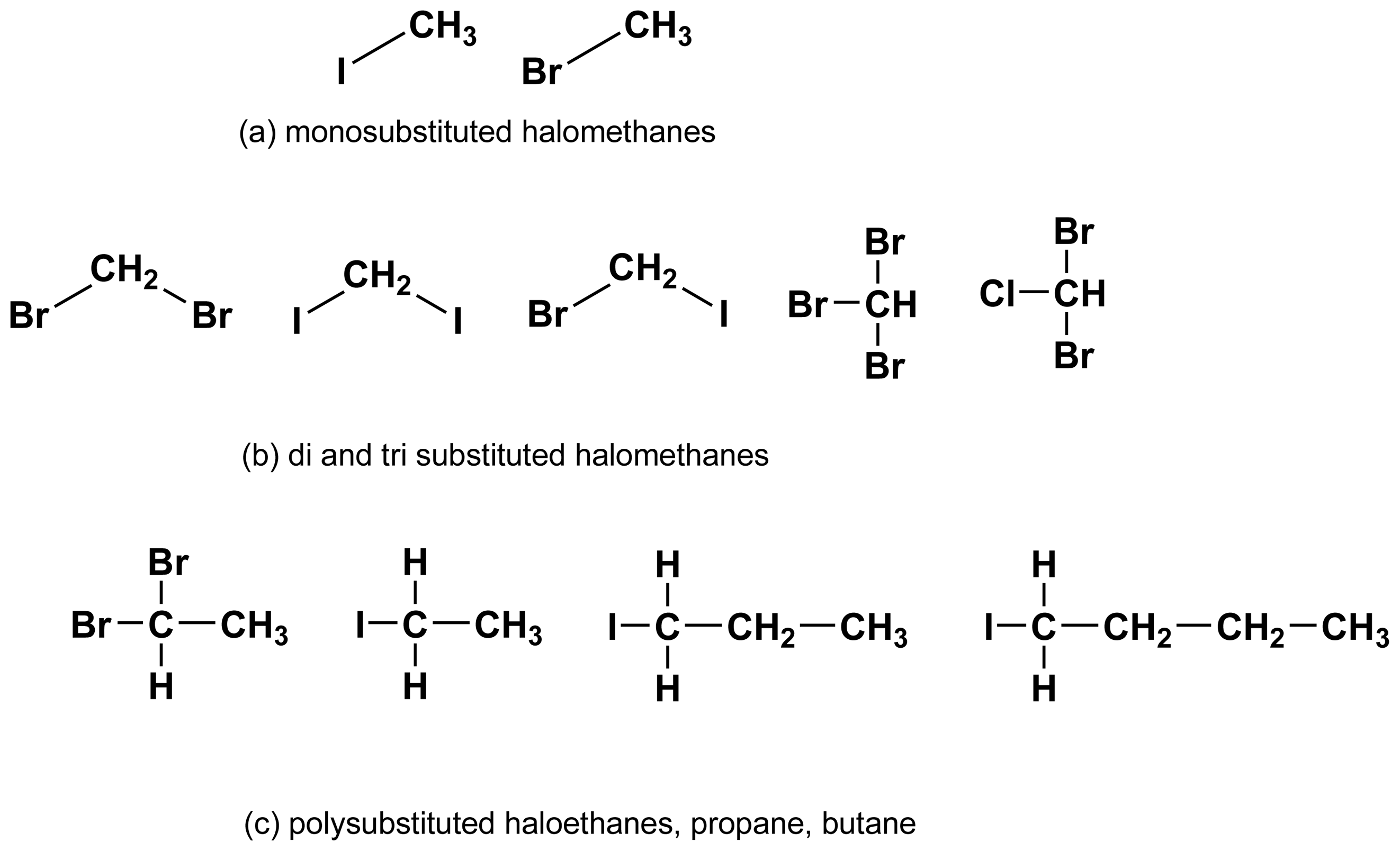
2.1. Halogenated alkanes
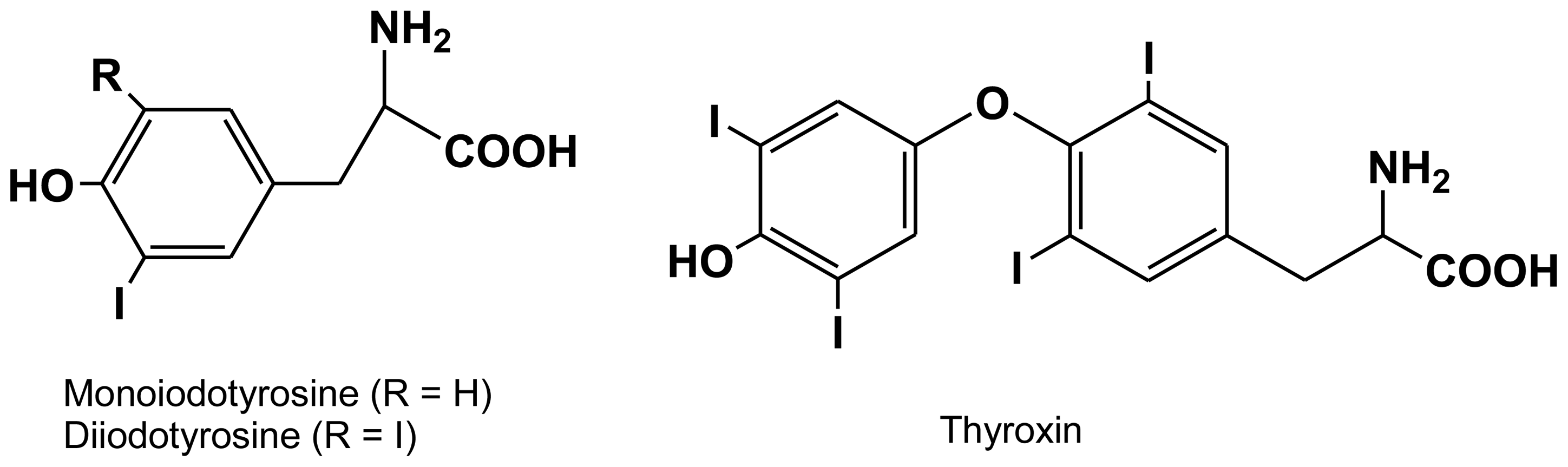
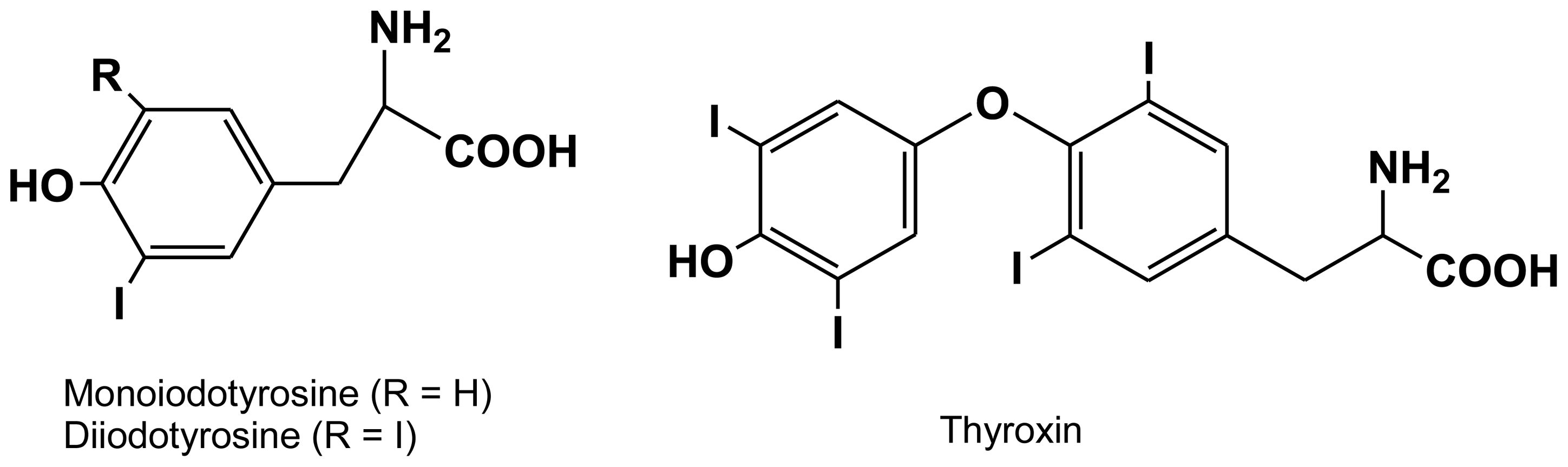
2.2. Tyrosine-bound halogens
2.3. Halogenated phenolics
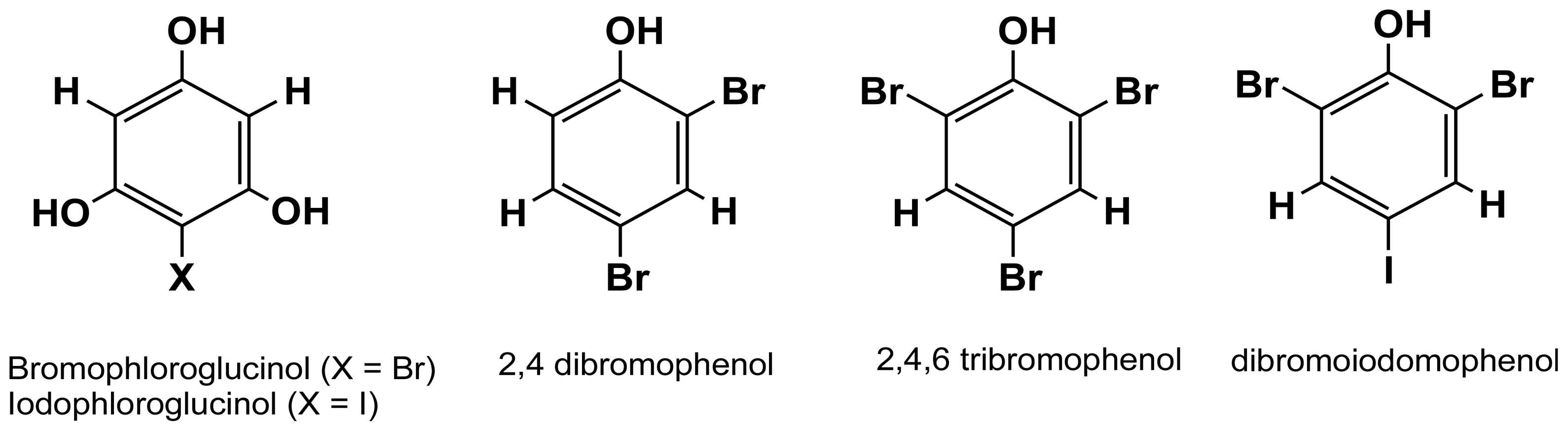
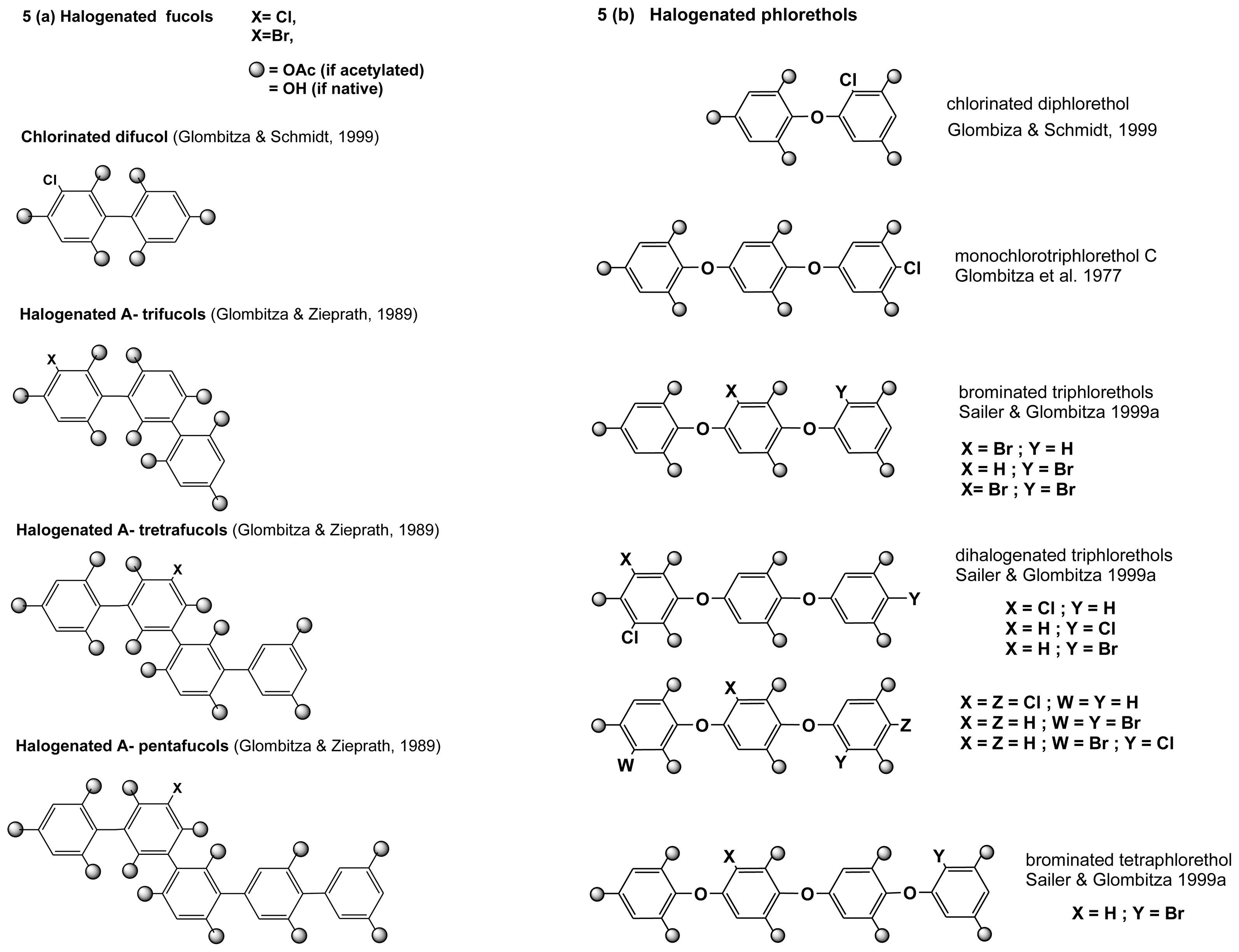
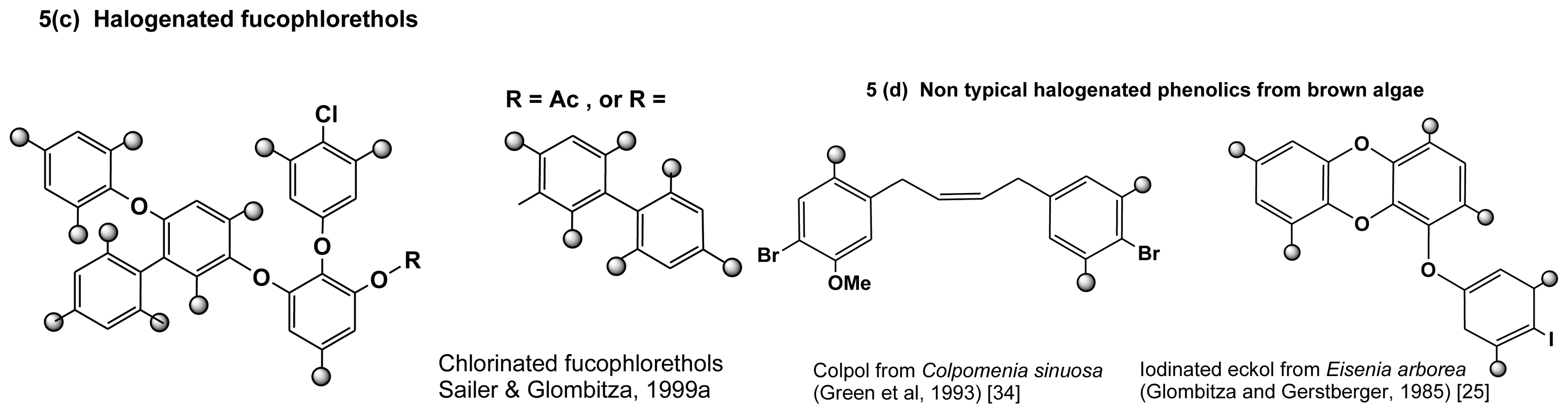
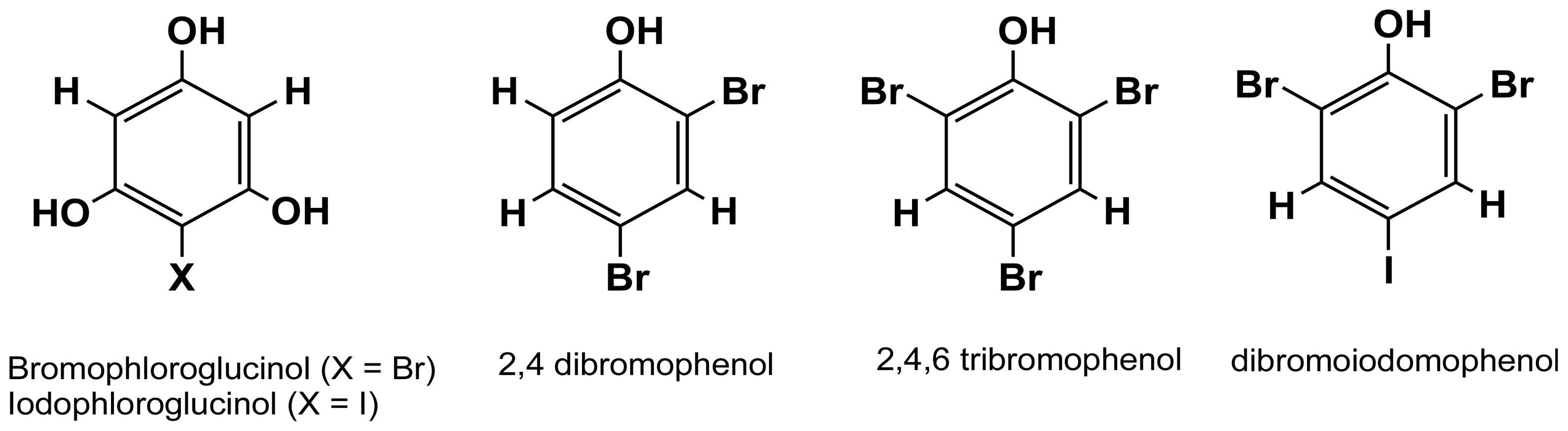
2.3.1. Halogenated phloroglucinols and phenols
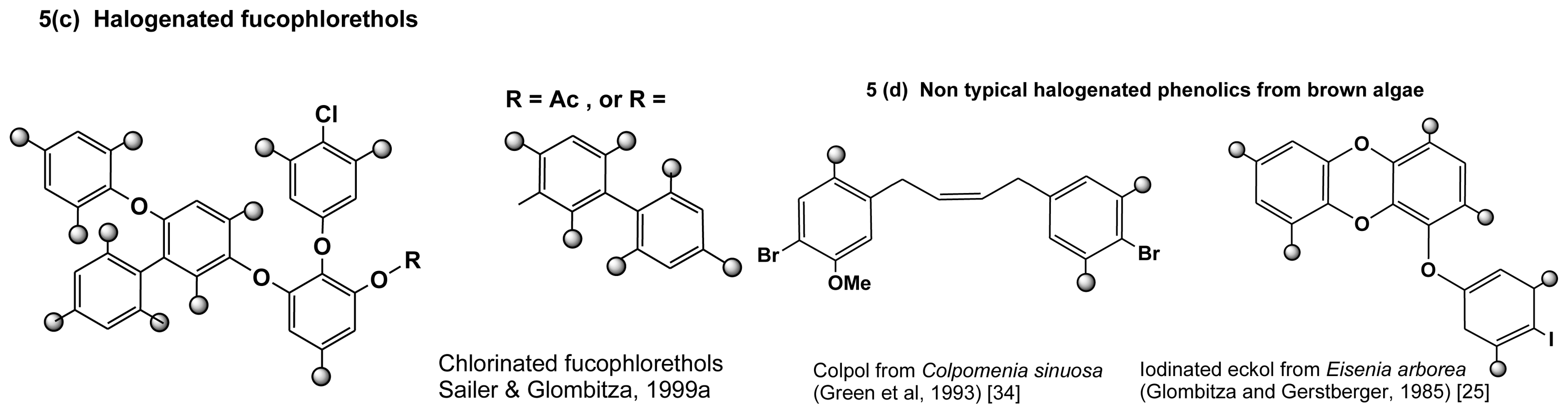
2.3.2. Halogenated phlorotannins
2.3.3. Biosynthesis of halogenated phlorotannins
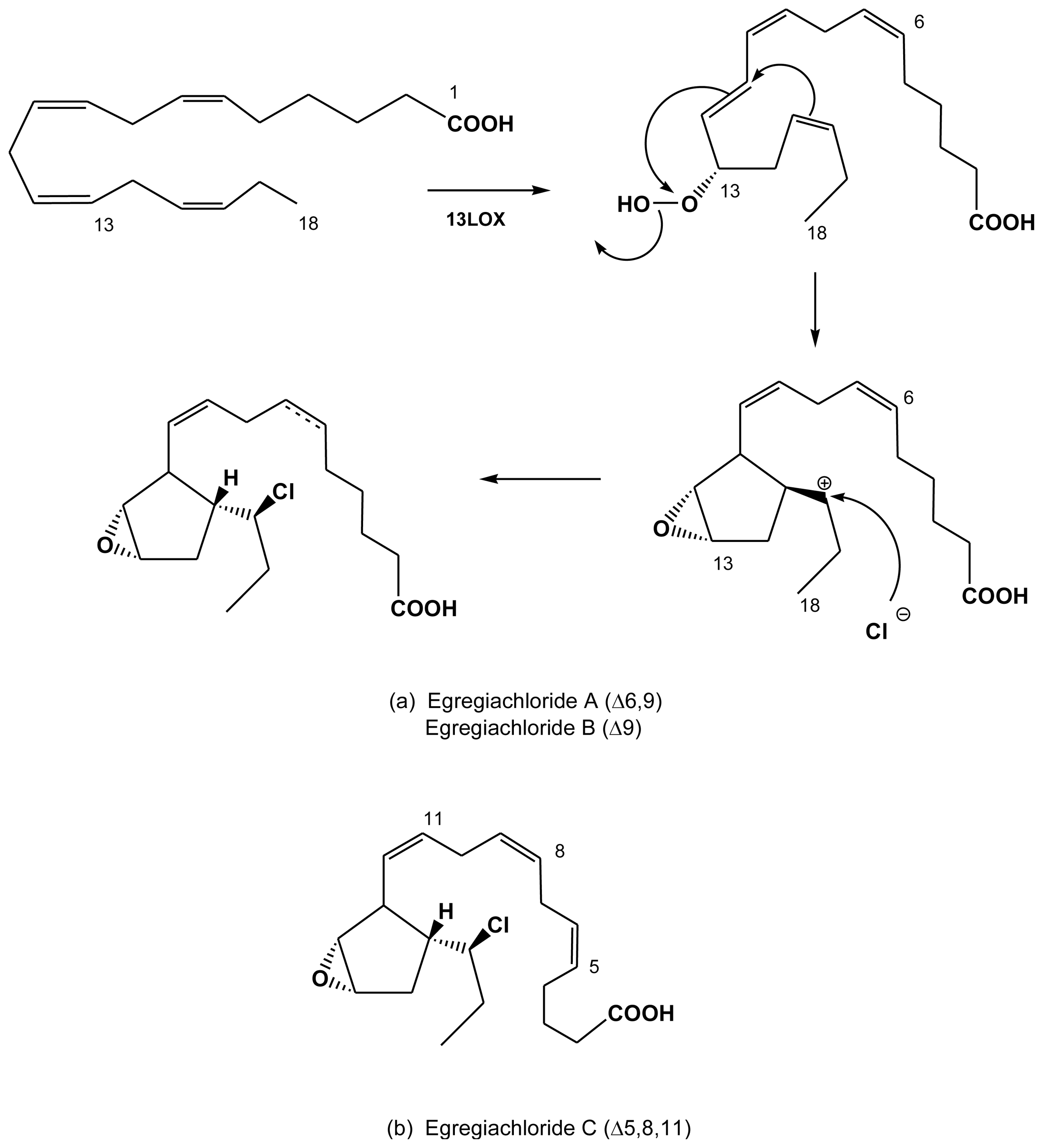
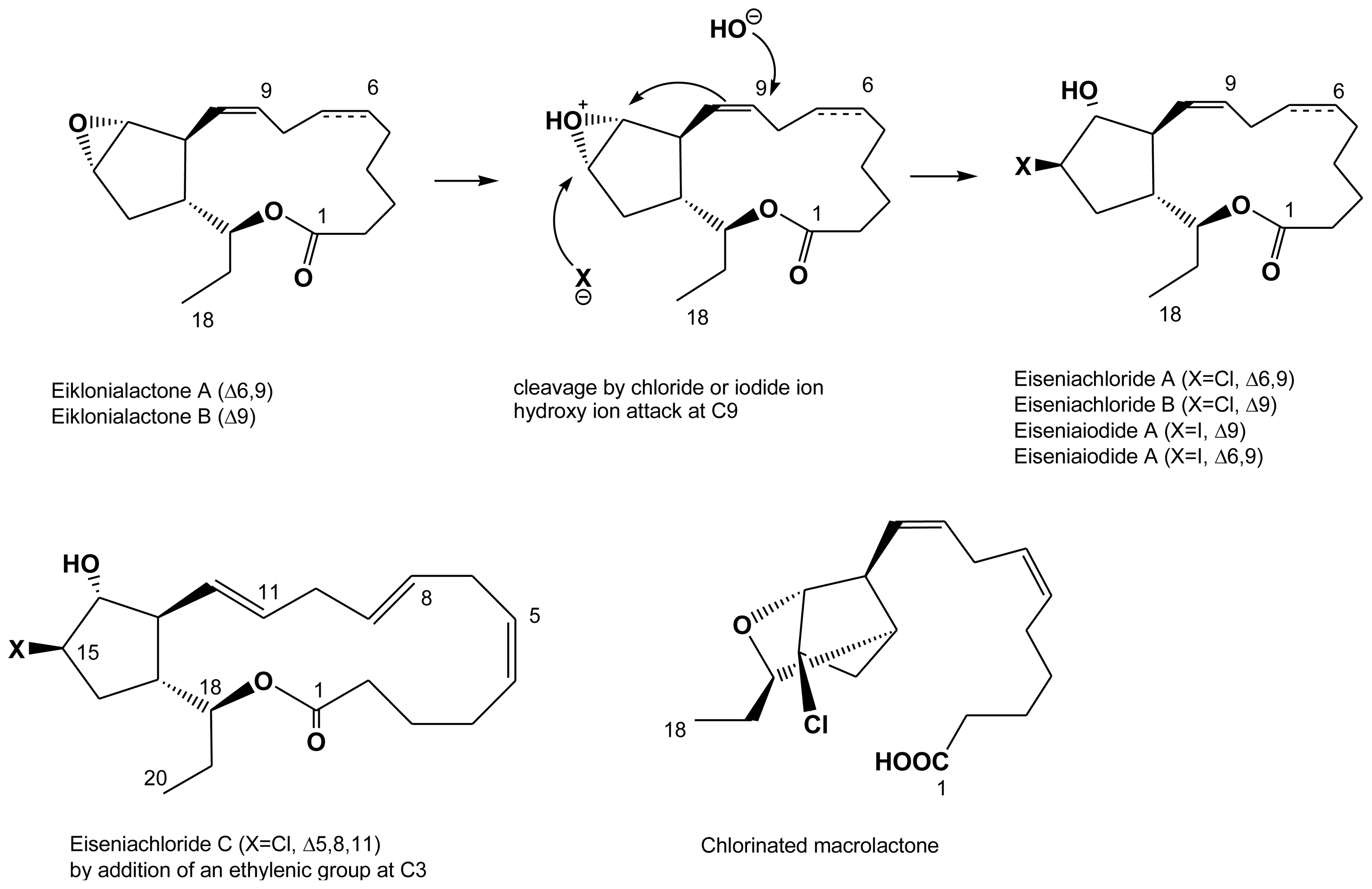
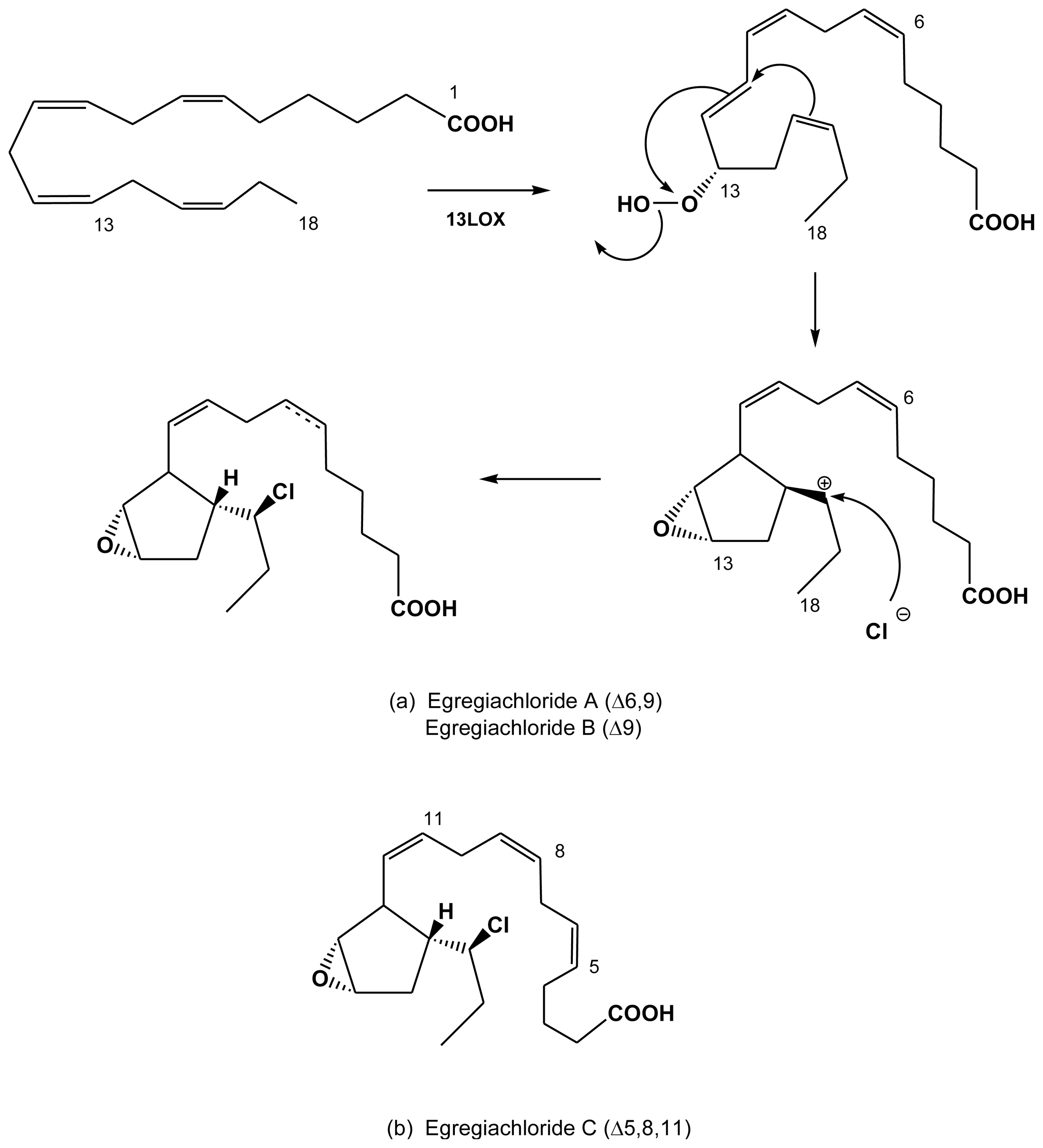
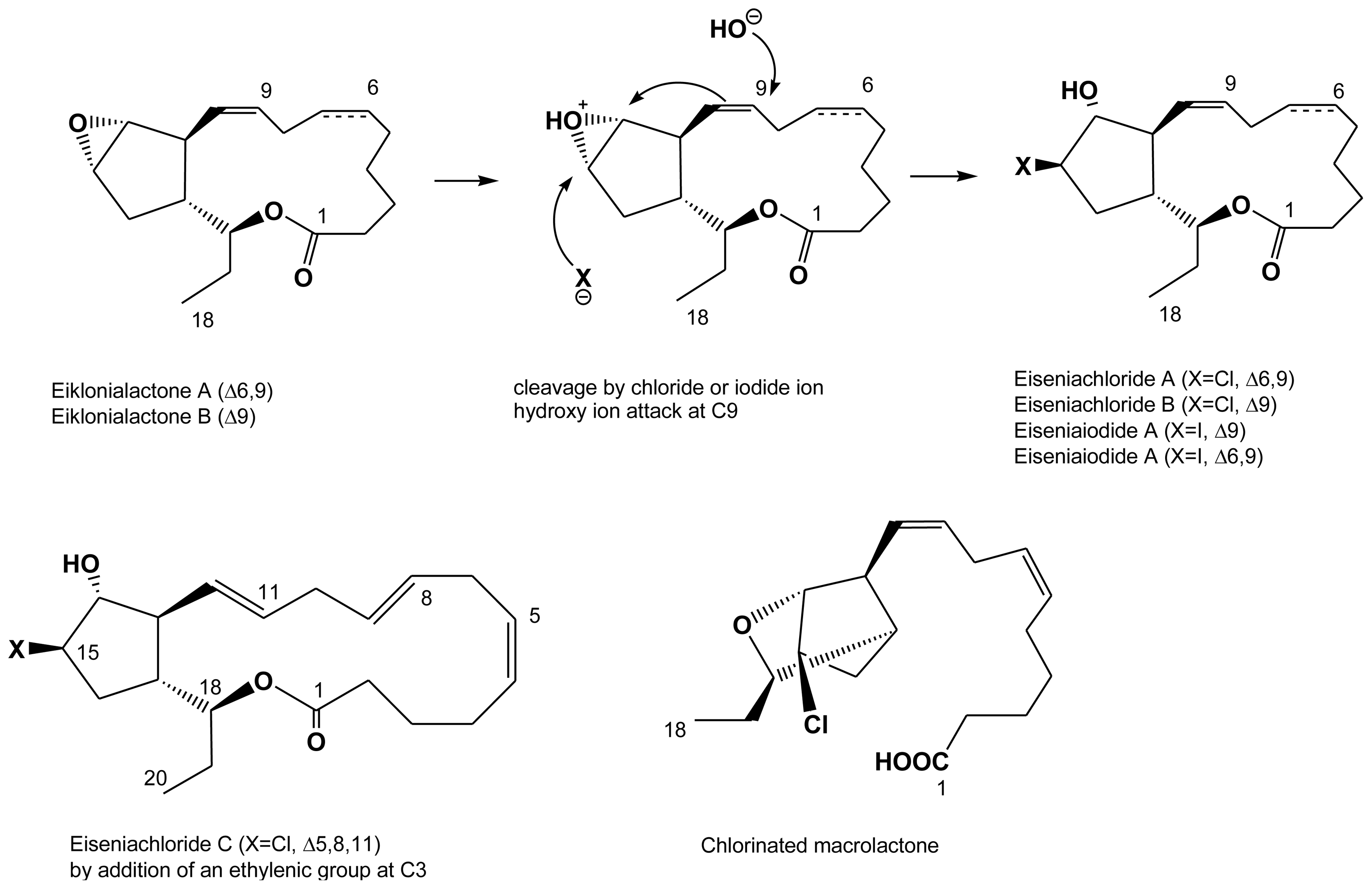
2.4. Halogenated fatty acids
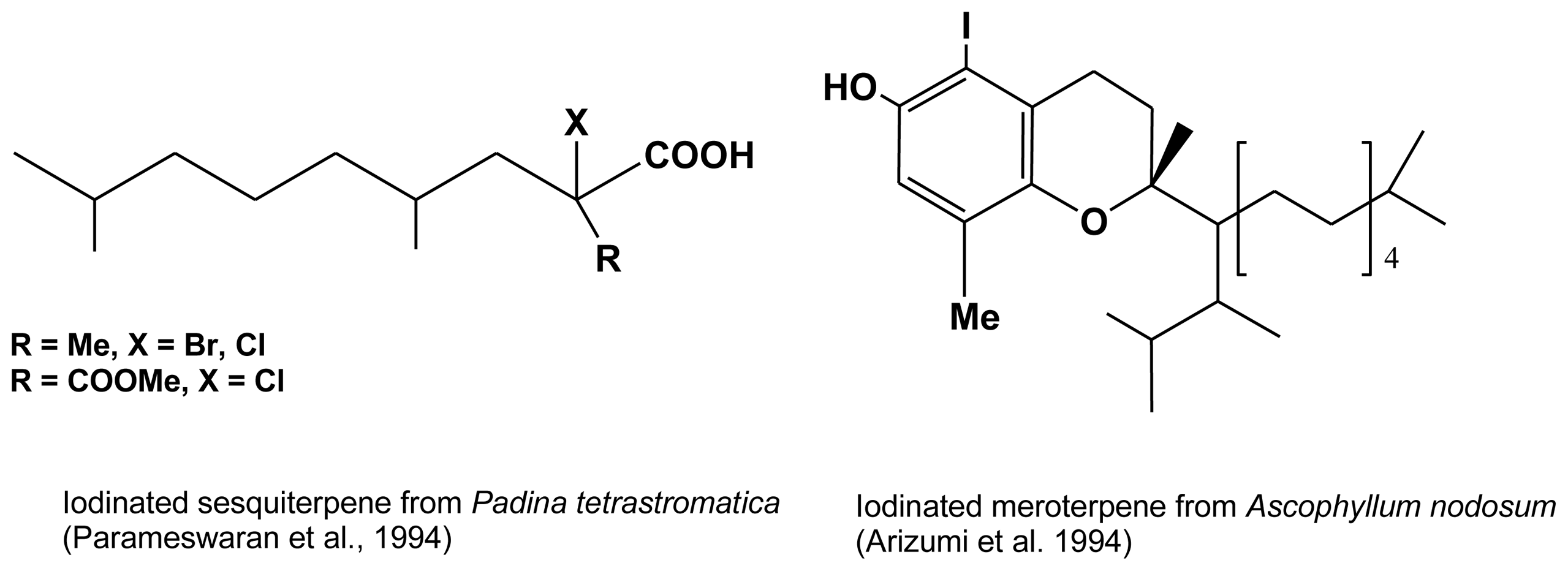
2.5. Halogenated terpenes
2.6. Halogenated polysaccharides
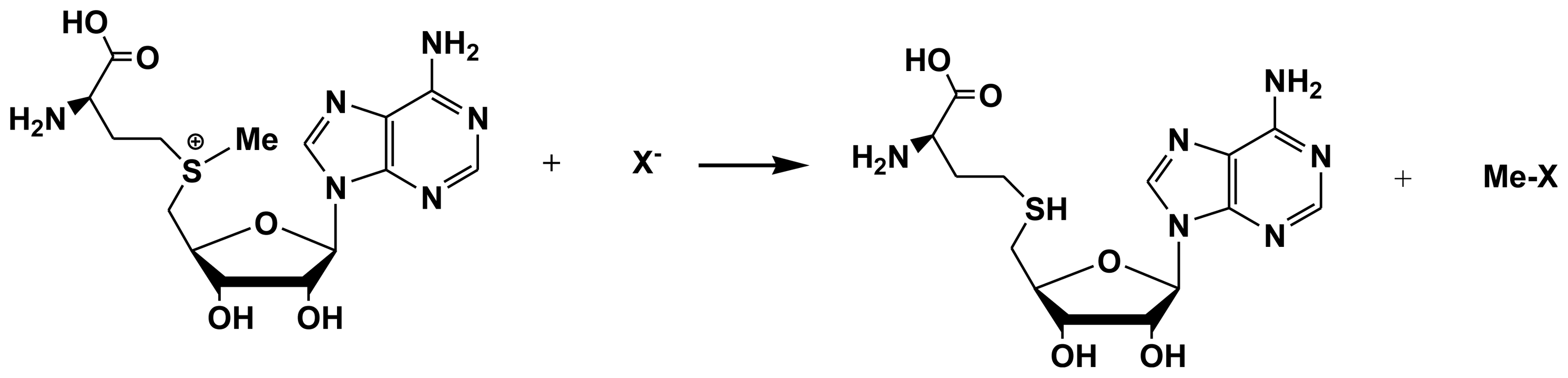
3. The Enzymology of Halogenation in Brown Algae
4. The Function of Bromination in Brown Algal Adhesion
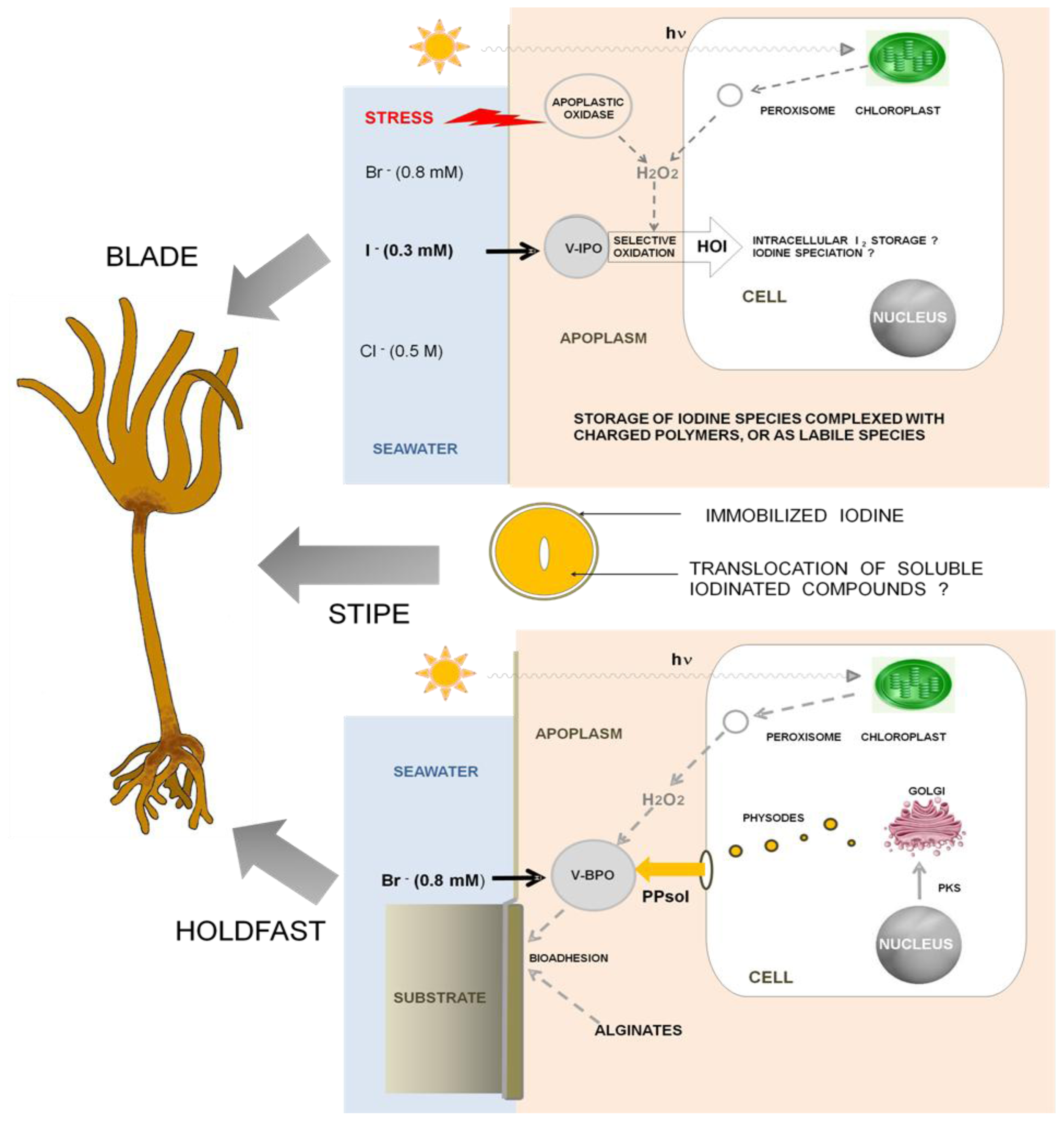
5. The Role of Iodine in Brown Algae
5.1. The emergence of specificity for iodine oxidation in brown algae
5.2. The function of the iodine metabolism
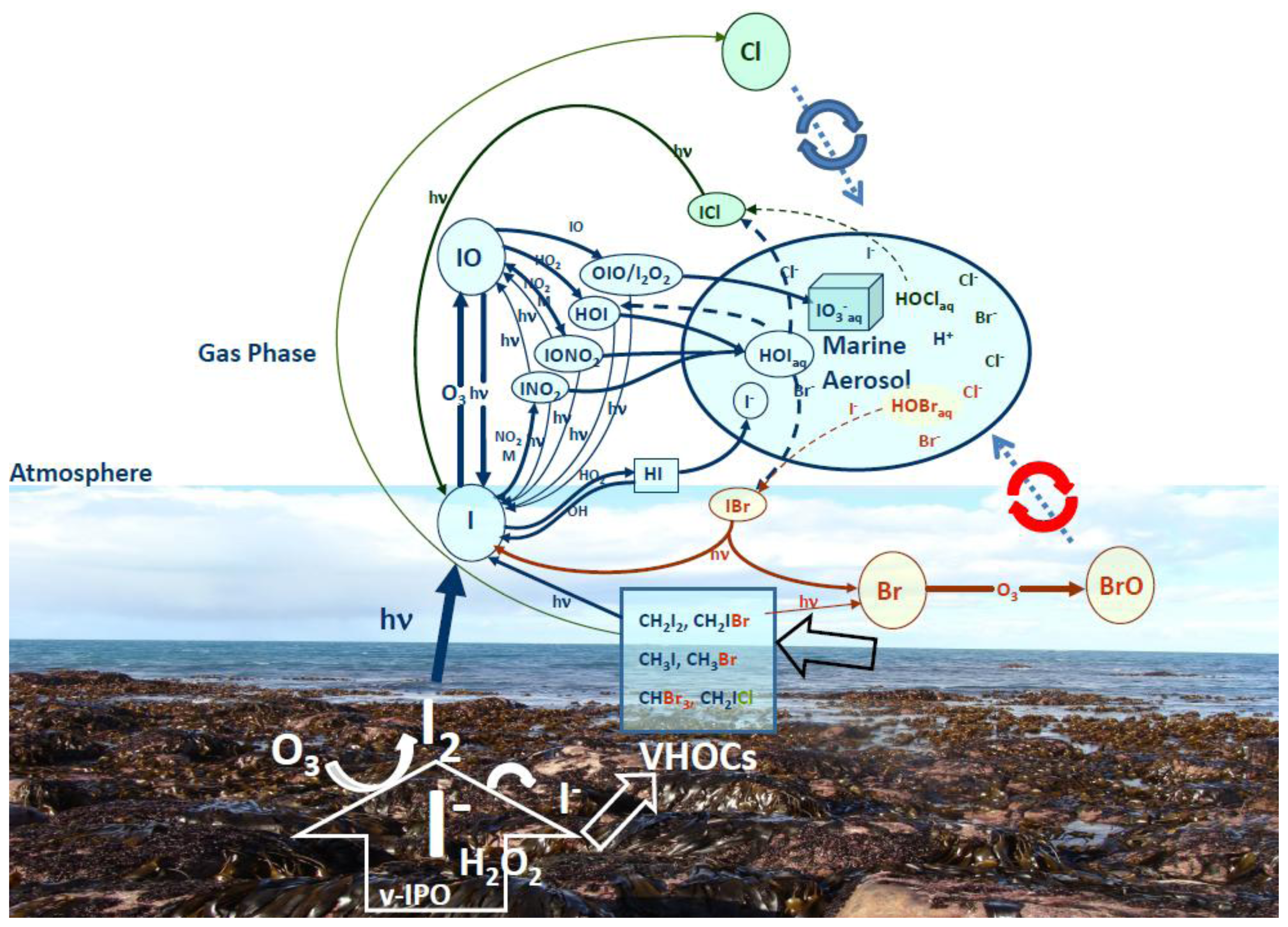
5.3. Environmental consequences of the iodine metabolism
6. New Approaches, New Answers?
Ascophyllum nodosum
References
- Gribble, G. The diversity of naturally produced organohalogens. Chemosphere 2003, 52, 289–297. [Google Scholar]
- Bernroitner, M; Zamocky, M; Furtmüller, PG; Peschek, GA; Obinger, C. Occurrence, phylogeny, structure, and function of catalases and peroxidases in cyanobacteria. J Exp Bot 2009, 60, 423–440. [Google Scholar]
- Palmer, JD. The symbiotic birth and spread of plastids: how many times and whodunit. J Phycol 2003, 39, 4–11. [Google Scholar]
- Phillips, N; Calhoun, S; Moustafa, A; Bhattacharia, D; Braun, EL. Genomic insights into evolutionary relationships among heterokont lineages emphasizing the Phaeophyceae. J Phycol 2008, 44, 15–18. [Google Scholar]
- Harper, MK; Bugni, TS; Copp, BR; James, RD; Lindsay, BS; Richardson, AD; Schnabel, PC; Tasdemir, D; VanWagoner, RM; Verbitzki, SM; Ireland, CM. McClintock, JB, Baker, BJ, Eds.; Introduction to the chemical ecology of marine natural products. In Marine Chemical Ecology; CRC: Boca Raton, FL, USA, 2001; pp. 3–71. [Google Scholar]
- MarinLit database, Department of Chemistry, University of Canterbury. http://www.chem.canterbury.ac.nz/marinlit/marinlit.shtml (accessed January 2010).
- Maschek, JA; Baker, BJ. Amsler, CD, Ed.; The chemistry of algal secondary metabolism. In Algal Chemical Ecology; Springer-Verlag Berlin: Heidelberg, Germany, 2008; pp. 1–24. [Google Scholar]
- Blunt, JW; Copp, BR; Hu, WP; Munro, MHG; Northcote, PT; Prinsep, MR. Marine natural products. Nat Prod Rep 2009, 26, 170–244. [Google Scholar]
- Amachi, S; Kamagata, Y; Kanagawa, T; Muramatsu, Y. Bacteria mediate methylation of iodine in marine and terrestrial environments. Appl Environ Microbiol 2006, 67, 2718–2722. [Google Scholar]
- Neilson, AH. Neilson, AH, Ed.; Biological effects and biosynthesis of brominated metabolites. In The Handbook of Environmental Chemistry, 1st ed; Springer: Berlin, Germany, 2003; Volume 3, pp. 75–204. [Google Scholar]
- Milkova, T; Talev, G; Christov, R; Dimitrikova-Konaklieva, S; Popov, S. Sterols and volatiles in Cystoseira barbata and Cystoseira crinita from the Black Sea. Phytochemistry 1997, 45, 93–95. [Google Scholar]
- Nightingale, PD; Malin, G; Liss, PS. Production of chloroform and other low-molecular-weight halocarbons by some species of macroalgae. Limnol Oceanogr 1995, 40, 680–689. [Google Scholar]
- Borchardt, SA; Allain, EJ; Michels, JJ; Stearns, GW; Kelly, RF; McCoy, WF. Reaction of acylated homoserine lactone bacterial signaling molecules with oxidized halogen antimicrobials. Appl Environ Microbiol 2001, 67, 3174–3179. [Google Scholar]
- Wagner, C; El Omari, M; König, GM. Biohalogenation: nature’s way to synthesize halogenated metabolites. J Nat Prod 2009, 72, 540–553. [Google Scholar]
- Verhaeghe, EF; Fraysse, A; Guerquin-Kern, JL; Wu, TD; Devès, G; Mioskowski, C; Leblanc, C; Ortega, R; Ambroise, Y; Potin, P. Microchemical imaging of iodine distribution in the brown alga Laminaria digitata suggests a new mechanism for its accumulation. J Biol Inorg Chem 2008, 13, 257–269. [Google Scholar]
- Tong, W; Chaikoff, IL. Metabolism of 131I by the marine alga Nereocystis luetkeana. J Biol Chem 1955, 215, 473–484. [Google Scholar]
- Roche, J; Yagi, Y. Sur la fixation de l’iode radioactif par les algues et sur les constituants iodés des Laminaires. C R Soc Biol Paris 1952, 146, 642–645. [Google Scholar]
- Hou, X; Chai, C; Qian, Q; Yan, X; Fan, X. Determination of chemical species of iodine in some seaweeds (I). Sci Total Environ 1997, 204, 215–221. [Google Scholar]
- Shah, M; Wuilloud, RG; Kannamkumaratha, SS; Caruso, JA. Iodine speciation studies in commercially available seaweed by coupling different chromatographic techniques with UV and ICP-MS detection. J Anal At Spectrom 2005, 20, 176–182. [Google Scholar]
- Crockford, SJ. Evolutionary roots of iodine and thyroid hormones in cell cell signaling. Integr Comp Biol 2009, 49, 155–166. [Google Scholar]
- Heyland, A; Moroz, LL. Cross-kingdom hormonal signaling: an insight from thyroid hormone functions in marine larvae. J Exp Biol 2005, 208, 4355–4361. [Google Scholar]
- Morrison, M; Schonbaum, GR. Snell, EE, Ed.; Peroxide-catalysed halogenations. In Annual Review of Biochemistry; Annual Reviews Inc: Palo Alto, CA, USA, 1976; Volume 45, pp. 861–888. [Google Scholar]
- Küpper, FC; Müller, D; Peters, A; Kloareg, B; Potin, P. Oligoalginate recognition and oxidative burst play a key role in natural and induced resistance of the sporophytes of Laminariales. J Chem Ecol 2002, 28, 2037–2061. [Google Scholar]
- Singh, IP; Bharate, SB. Phloroglucinol compounds of natural origin. Nat Prod Rep 2006, 23, 558–591. [Google Scholar]
- Glombitza, K-W; Gerstberger, G. Phlorotannins with dibenzodioxin structural elements from the brown alga Eisenia arborea. Phytochemistry 1985, 24, 543–551. [Google Scholar]
- Shibata, T; Hama, Y; Miyasaki, T; Ito, M; Nakamura, T. Extracellular secretion of phenolic substances from living brown algae. J Appl Phycol 2006, 18, 787–794. [Google Scholar]
- Whitfield, FB; Helidoniotis, F; Shaw, KJ; Svoronos, D. Distribution of bromophenols in species of marine algae from Eastern Australia. J Agric Food Chem 1999, 47, 2367–2373. [Google Scholar]
- Koch, ML; Gregson, RP. Brominated phlorethols and non-halogenated phlorotannins from the brown alga Cystophora congesta. Phytochemistry 1984, 13, 2633–2637. [Google Scholar]
- Sailer, B; Glombitza, K-W. Halogenated phlorethols and fucophlorethols from the brown alga Cystophora retroflexa. Nat. Toxins 1999a, 7, 57–62. [Google Scholar]
- Sailer, B; Glombitza, K-W. Phlorethols and fucophlorethols from the brown alga Cystophora retroflexa. Phytochemistry 1999b, 50, 869–881. [Google Scholar]
- Glombitza, K-W; Koch, M; Eckhardt, G. Chlorierte phlorethole aus Laminaria ochroleuca. Phytochemistry 1977, 16, 796–798. [Google Scholar]
- Glombitza, K-W; Schmidt, A. Nonhalogenated and halogenated phlorotannins from the brown alga Carpophyllum angustifolium. J Nat Prod 1999, 62, 1238–1240. [Google Scholar]
- Glombitza, K-W; Zieprath, G. Phlorotannins of the brown alga Analipus japonicus. Planta Med 1989, 55(2), 171–175. [Google Scholar]
- Green, D; Kashman, Y; Miroz, A. Colpol, a new cytotoxic C6-C4-C6 metabolite from the alga Colpomenia sinuosa. J Nat Prod 1993, 56, 1201–1202. [Google Scholar]
- Waterman, PG; Mole, S. Analysis of phenolic plant metabolites; Blackwell Scientific Publications: Oxford, UK, 1994. [Google Scholar]
- Arnold, TM; Targett, N. To grow and defend: lack of tradeoffs for brown algal phlototannins. Oikos 2003, 102, 406–408. [Google Scholar]
- Amsler, CD; Fairhead, VA. Defensive and sensory chemical ecology of brown algae. Adv Bot Res 2006, 43, 1–73. [Google Scholar]
- Berglin, M; Delage, L; Potin, P; Vilter, H; Elwing, H. Enzymatic cross-linking of a phenolic polymer extracted from the marine alga Fucus serratus. Biomacromolecules 2004, 5, 2376–2383. [Google Scholar]
- Bitton, R; Berglin, M; Elwing, H; Colin, C; Delage, L; Potin, P; Bianco-Peled, H. The influence of halide-mediated oxidation on algae-born adhesives. Macromol Biosci 2007, 7, 1280–1289. [Google Scholar]
- Ragan, MA; Jensen, A. Widespread distribution of sulfated polyphenols in brown algae. Phytochemistry 1979, 18, 261–262. [Google Scholar]
- Todd, JS; Proteau, PJ; Gerwick, WH. Egregiachlorides A-C: new chlorinated oxylipins from the brown marine alga Egregia menziesii. Tetrahedron Lett 1993, 34, 7689–7692. [Google Scholar]
- Kousaka, K; Ogi, N; Akazawa, Y; Fujieda, M; Yamamoto, Y; Takada, Y; Kimura, J. Novel oxylipin metabolites from the brown alga Eisenia bicyclis. J Nat Prod 2003, 66, 1318–1323. [Google Scholar]
- Andreou, A; Brodhun, F; Feussner, I. Biosynthesis of oxylipins in non-mammals. Prog Lipid Res 2009, 48, 148–170. [Google Scholar]
- Mercer, EI; Davies, CL. Description of chlorosulfolipids in algae. Phytochemistry 1979, 18, 457–462. [Google Scholar]
- Parameswaran, PS; Bhat, KL; Das, B; Kamat, SY; Harnos, S. Halogenated terpenoids from the brown alga Padina tetrastromatica (Hauck). Indian J Chem Sect B 1994, 33, 1006–1008. [Google Scholar]
- Arizumi, H; Hata, K; Shimizu, N. Isolation and determination of structure of antioxidant chromanol derivative from Ascophyllum nodosum. Jpn Kokkai Tokyo Koho 1994, 122, 160312. [Google Scholar]
- Kladi, M; Vagias, C; Roussis, V. Volatile halogenated metabolites from marine red algae. Phytochem Rev 2004, 3, 337–366. [Google Scholar]
- Kloareg, B; Quatrano, RS. Structure of the cell walls of marine algae and ecophysiological functions of the matrix polysaccharides. Oceanogr Mar Biol Ann Rev 1988, 26, 259–315. [Google Scholar]
- Vaillancourt, FH; Yeh, E; Vosburg, DA; Garneau-Tsodikova, S; Walsh, CT. Nature’s Inventory of Halogenation Catalysts: Oxidative Strategies Predominate. Chem Rev 2006, 106, 3364–3378. [Google Scholar]
- Butler, A; Sandy, M. Mechanistic considerations of halogenating enzymes. Nature 2009, 460, 848–854. [Google Scholar]
- Natalio, F; Humanes, M; Wever, R. Vanadium Haloperoxidases Versatile enzymes with multiple applications. Mar Drugs 2010. submitted. [Google Scholar]
- Winter, JM; Moore, BS. Exploring the chemistry and biology of vanadium-dependant haloperoxidases. J Biol Chem 2009, 284, 18577–18580. [Google Scholar]
- Vilter, H. Peroxidases from Phaeophyceae: a vanadium (V)-dependent peroxidase from Ascophyllum nodosum. Phytochemistry 1984, 23, 1387–1390. [Google Scholar]
- Weyand, MH; Hecht, HJ; Kiess, M; Liaud, MF; Vilter, H; Schomburg, D. X-ray structure determination of a vanadium-dependant haloperoxidase from Ascophyllum nodosum at 2.0 A resolution. J Biol Mol 1999, 293, 595–611. [Google Scholar]
- Feiters, MC; Küpper, FC; Meyer-Klaucke, W. X-ray absorption spectroscopic studies on model compounds for biological iodine and bromine. J Synchrotron Radiat 2005, 12, 85–93. [Google Scholar]
- Leblanc, C; Colin, C; Cosse, A; Delage, L; La Barre, S; Morin, P; Fiévet, B; Voiseux, C; Ambroise, Y; Verhaeghe, E; et al. Iodine transfers in the coastal marine environment: the key role of brown algae and of their vanadium-dependent haloperoxidases. Biochimie 2006, 88, 1773–1785. [Google Scholar]
- Moore, RM; Groszko, W; Niven, SJ. Ocean-atmosphere exchange of methyl chloride: Results from NW Atlantic and Pacific Ocean studies. J Geophys Res 1996, 101, 28529–28538. [Google Scholar]
- Bowler, C; Allen, AE; Badger, JH; Grimwood, J; Jabbari, K; Kuo, A; Maheswari, U; Martens, C; Maumus, F; Otillar, RP; et al. The Phaeodactylum genome reveals the evolutionary history of diatom genomes. Nature 2008, 456, 239–244. [Google Scholar]
- Collén, J; Roeder, V; Rousvoal, S; Collin, O; Kloareg, B; Boyen, C. An expressed sequence tag analysis of thallus and regenerating protoplasts of Chondrus crispus (Gigartinales, Rhodophyceae). J Phycol 2006, 42, 104–112. [Google Scholar]
- Colin, C; Leblanc, C; Michel, G; Wagner, E; Leize-Wagner, E; van Dorsselaer, A; Potin, P. Vanadium-dependent iodoperoxidases in Laminaria digitata, a novel biochemical function diverging from brown algal bromoperoxidases. J Biol Inorg Chem 2005, 10, 156–166. [Google Scholar]
- Cosse, A; Potin, P; Leblanc, C. Patterns of gene expression induced by oligoguluronates reveal conserved and environment-specific molecular defense responses in the brown alga Laminaria digitata. New Phytol 2009, 182, 239–250. [Google Scholar]
- Yotsukura, N; Nagai, K; Kimura, H; Morimoto, K. Seasonal changes in proteomic profiles of Japanese kelp Saccharina japonica (Laminariales, Phaeophyceae). J Appl Phycol 2010. [Google Scholar] [CrossRef]
- Vreeland, V; Laetsch, WM. Adair, WS, Mecham, RP, Eds.; A gelling carbohydrate in algal cell wall formation. In Organization and Assembly of Plant and Animal Extracellular Matrix; Academic Press: San Diego, CA, USA, 1990; pp. 137–171. [Google Scholar]
- Vreeland, V; Epstein, L. Linskins, HF, Jackson, JF, Eds.; Analysis of plant-substratum adhesives. In Plant Cell Wall Analysis: Modern Methods of Plant Analysis; Springer-Verlag: Berlin, Germany, 1996; Volume 17, pp. 95–116. [Google Scholar]
- Vreeland, V; Waite, JH; Epstein, L. Polyphenols and oxydases in substratum adhesion by marine algae and mussels. J Phycol 1998, 34, 1–8. [Google Scholar]
- Ragan, MA; Glombitza, K-W. Phlorotannins, brown algal polyphenols. Progr Phycol Res 1986, 4, 129–241. [Google Scholar]
- Bitton, R; Ben-Yehuda, M; Davidovich, M; Balazs, Y; Potin, P; Delage, L; Colin, C; Bianco-Peled, H. Structure of Algal-Born Phenolic Polymeric Adhesives. Macromol Biosci 2006, 7, 1280–1289. [Google Scholar]
- Salgado, LT; Cinelli, LP; Viana, NB; Tomazetto de Carvalho, R; Mourão, P; Teixeira, VL; Farina, M; Amado Filho, GM. Vanadium bromoperoxidase catalyzes the formation of high-molecular-weight complexes between brown algal phenolic substances and alginates. J Phycol 2009, 45, 193–202. [Google Scholar]
- Potin, P; Leblanc, C. Smith, AM, Callow, JA, Eds.; Phenolic-based adhesives of marine brown algae. In Biological Adhesives; Springer: Berlin, Germany, 2006; Volume Chapter 6, pp. 105–119. [Google Scholar]
- Küpper, FC; Schweigert, N; Ar Gall, E; Legendre, J-M; Vilter, H; Kloareg, B. Iodine uptake in Laminariales involves extracellular, haloperoxidase-mediated oxidation of iodide. Planta 1998, 207, 163–167. [Google Scholar]
- Küpper, FC; Carpenter, L; McFiggans, GB; Palmer, CJ; Waite, TJ; Boneberg, EM; Woitsch, S; Weiller, M; Abela, R; Grolimund, D; et al. Iodide accumulation provides kelp with an inorganic antioxidant impacting atmospheric chemistry. Proc Natl Acad Sci USA 2008, 105, 6954–6958. [Google Scholar]
- Colin, C; Leblanc, C; Wagner, E; Delage, L; Leize-Wagner, E; van Dorsselaer, A; Kloareg, B; Potin, P. The brown algal kelp Laminaria digitata features distinct bromoperoxidase and iodoperoxidase activities. J Biol Chem 2003, 278, 23545–23552. [Google Scholar]
- Connan, S; Delisle, F; Deslandes, E; Ar Gall, E. Intra-thallus phlorotannin content and antioxidant activity in Phaeophyceae of temperate waters. Bot Mar 2006, 49, 39–46. [Google Scholar]
- Ar Gall, E; Küpper, FC; Kloareg, B. A survey of iodine contents in Laminaria digitata. Bot Mar 2004, 47, 30–37. [Google Scholar]
- Lobban, CS; Harrison, PJ. Lobban, CS, Harrison, PJ, Eds.; Light and photosynthesis. In Seaweed Ecology and Physiology, 1st ed; Cambridge University Press: Cambridge, UK, 1994; pp. 151–153. [Google Scholar]
- Lobban, C. Translocation of 14C in Macrocystis pyrifera (Giant Kelp). Plant Physiol 1978, 61, 585–589. [Google Scholar]
- Manley, SL. Composition of sieve-tube sap from Macrocystis pyrifera (Phaeophyta) with emphasis on the inorganic constituents. J Phycol 1993, 19, 118–121. [Google Scholar]
- Amat, MA; Srivastava, LM. Translocation of iodine in Laminaria saccharina (Phaeophyta). J Phycol 1985, 21, 330–333. [Google Scholar]
- Palmer, CJ; Anders, TL; Carpenter, LJ; Küpper, FC; McFiggans, G. Iodine and halocarbon response of Laminaria digitata to oxidative stress and links to atmospheric new particle production. Environ Chem 2005, 2, 282–290. [Google Scholar]
- Goulitquer, S; Ritter, A; Thomas, F; Ferec, C; Salaun, JP; Potin, P. Release of volatile aldehydes by the brown algal kelp Laminaria digitata in response to both biotic and abiotic stress. ChemBioChem 2009, 6, 977–982. [Google Scholar]
- Carpenter, LJ; Sturges, WT; Liss, PS; Penkett, SA; Alicke, B; Hebestreit, K; Platt, U. Short lived alkyl-iodides and bromides at Mace Head: Links to macroalgal emission and halogen oxide formation. J Geophys Res 1999, 104, 1679. [Google Scholar]
- Carpenter, LJ; Malin, G; Liss, PS; Küpper, FC. Novel biogenic iodine-containing trihalomethanes and other short-lived halocarbons in the coastal East Atlantic. Global Biogeochem Cycles 2000, 14, 1191–1204. [Google Scholar]
- Hughes, C; Malin, G; Nightingale, PD; Liss, PS. The effect of light stress on the release of volatile iodocarbons by three species of marine microalgae. Limnol Oceanogr 2006, 51, 2849–2854. [Google Scholar]
- Hughes, C; Malin, G; Turley, C; Keely, B; Nightingale, P; Liss, P. The production of volatile iodocarbons by biogenic marine aggregates. Limnol Oceanog 2008, 53, 867–872. [Google Scholar]
- McFiggans, G; Coe, H; Burgess, R; Allan, J; Cubison, MR; Alfarra, MR; Saunders, R; Saiz-Lopez, A; Plane, JMC; Wevill, D; et al. Direct evidence for coastal iodine particles from Laminaria macroalgae - linkage to emissions of molecular iodine. Atm Chem Phys 2004, 4, 701–713. [Google Scholar]
- O’Dowd, C; McFiggans, G; Creasey, DJ; Pirjola, L; Hoell, C; Smith, MH; Allan, BJ; Plane, JMC; Heard, DE; Lee, JD; et al. On the photochemical production of new particles in the coastal boundary layer. Geophys Res Lett 1999, 26, 1707–1710. [Google Scholar]
- O’Dowd, CD; Jimenez, JL; Bahreini, R; Flagan, R; Seinfeld, JH; Hämeri, K; Pirjola, L; Kulmala, M; Jennings, SG; Hoffman, T. Marine aerosol formation from biogenic iodine emissions. Nature 2002, 417, 632–636. [Google Scholar]
- Lee, JD; McFiggans, G; Allan, JD; Baker, AR; Ball, SM; Benton, AK; Carpenter, LJ; Commane, R; Finley, BD; Evans, M; et al. Iodine-mediated coastal particle formation: an overview of the Reactive Halogens in the Marine Boundary Layer (RHaMBLe) Roscoff coastal study. Atmos Chem Phys Discuss 2009, 9, 26421–26489. [Google Scholar]
- Mahajan, AS; Oetjen, H; Saiz-Lopez, A; Lee, JD; McFiggans, G; Plane, JMC. Reactive iodine species in a semi-polluted environment. Geophys Res Lett 2009, 36, L16803. [Google Scholar] [CrossRef]
- Whalley, LK; Furneaux, KL; Gravestock, T; Atkinson, HM; Bale, CSE; Ingham, T; Bloss, WJ; Heard, DE. Detection of iodine monoxide radicals in the Marine Boundary Layer using laser induced fluorescence spectroscopy. J Atmos Chem 2007, 58, 19–39. [Google Scholar]
- Ball, SM; Hollingsworth, AM; Humbles, J; Leblanc, C; Potin, P; Langridge, JM; LeCrane, JP; McFiggans, G. Spectroscopic studies of molecular iodine emitted into the gas phase by seaweed. Atmos Chem Phys Discuss 2009, 9, 26329–26376. [Google Scholar]
- Leigh, RJ; Ball, SM; Whitehead, J; Leblanc, C; Shillings, AJL; Mahajan, AS; Oetjen, H; Dorsey, JR; Gallagher, M; Jones, RL; et al. Measurements and modelling of molecular iodine emissions, transport and photodestruction in the coastal region around Roscoff. Atmos Chem Phys Discuss 2009, 9, 21165–21198. [Google Scholar]
- Cainey, JM; Keywood, M; Bigg, EK; Grose, MR; Gillett, RW; Meyer, M. Flux chamber study of particle formation from Durvillaea potatorum. Environ Chem 2007, 4, 151–154. [Google Scholar]
- Almeida, M; Filipe, S; Humanes, M; Maia, MF; Melo, R; Severino, N; da Silva, JAL; Fraùsto da Silva, JJR; Wever, R. Vanadium haloperoxidases from brown algae of the Laminariaceae family. Phytochemistry 2001, 57, 633–642. [Google Scholar]
- Krenn, BE; Tromp, MG; Wever, R. The brown alga Ascophyllum nodosum contains two different vanadium bromoperoxidases. J Biol Chem 1989, 264, 19287–92. [Google Scholar]
- Carpenter, LJ. Iodine in the Marine Boundary Layer. Chem Rev 2003, 103, 4953–4962. [Google Scholar]
- Kornprobst, J-M. Kornprobst, J-M, Ed.; Les ressources documentaires. In Substances Naturelles d’Origine Marine, 1st ed; Lavoisier Tech & Doc: Paris, France, 2005; Volume 1, pp. 89–101. [Google Scholar]
- Carter-Franklin, J; Butler, A. Vanadium bromoperoxidase-catalyzed biosynthesis of halogenated marine natural products. J Am Chem Soc 2004, 126, 15060–15066. [Google Scholar]
- Ten Brink, HB; Schoemaker, HE; Wever, R. Sulfoxidation mechanism of vanadium bromoperoxidase from Ascophyllum nodosum-evidence for direct oxygen transfer catalysis. Eur J Biochem 2001, 268, 132–138. [Google Scholar]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species (Order) | Halogenated metabolite | Reference |
|---|---|---|
| Laminariales (La) | MIT, DIT | no specific reference |
| Laminaria digitata | volatile alkanes (Cl, Br, I) | Nightingale et al. (1995) [12] |
| Laminaria ochroleuca | monochlorotriPL C | Glombitza et al. (1977) [31] |
| Laminaria ochroleuca | monochlorotriphenol | Glombitza et al. (1977) (31] |
| Egregia menziesii | egregiachlorides A, B and C | Todd et al. (1993) [41] |
| Eisenia bicyclis | eiseniachlorides A, B and C | Kousaka et al. (2003) [42] |
| Eisenia bicyclis | eiseniaiodides A, B | Kousaka et al. (2003) [42] |
| Eisenia arborea | PG (I), EK (I) | Glombitza & Gerstberger (1985) [25] |
| Fucales (Fu) | ||
| Cystoseira barbata | halogenated alkanes | Milkova et al. (1997) [11] |
| Cystophora congesta | bromotriPL A2 | Koch & Gregson. (1984) (28] |
| Cystophora reflexa | mono, di, tri, tetraPL (Cl, Br, both), FP (Cl) | Sailer & Glombitza (1999)a [29] |
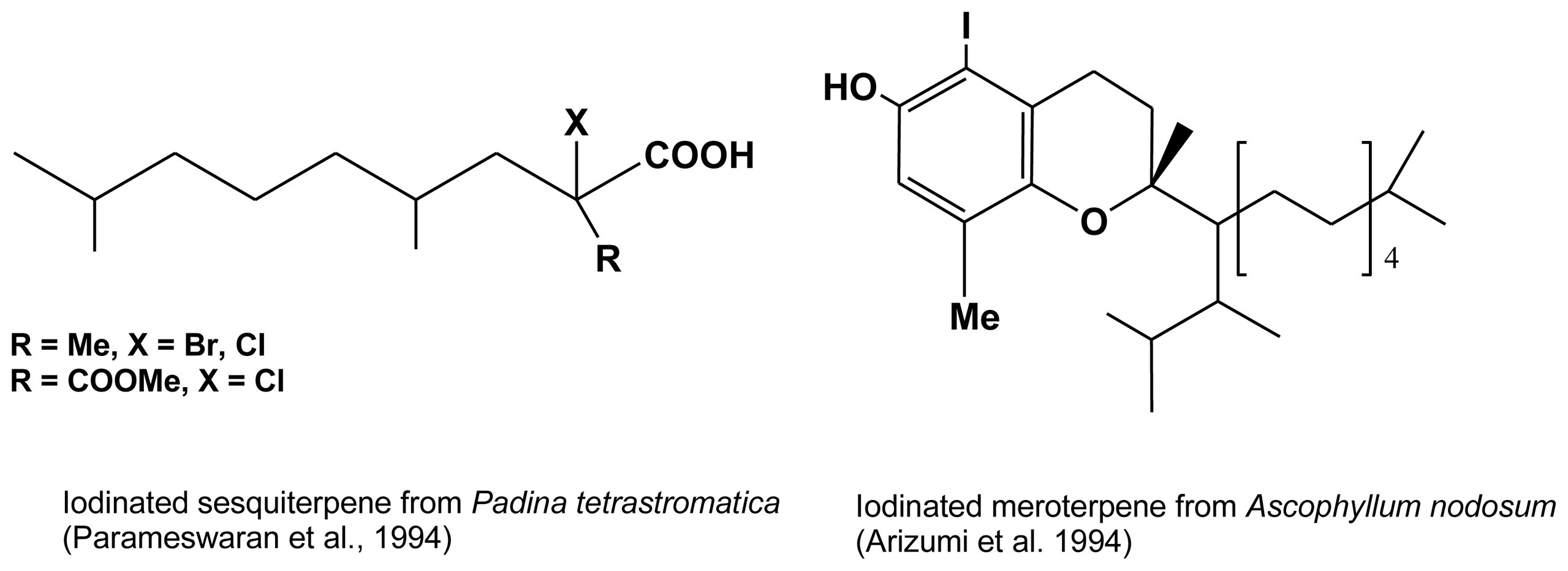
| Ascophyllum nodosum | iodinated meroterpene | Arizumi et al. (1994) (46] |
| Carpophyllum angustifolium | diFC (Cl), PG (Cl), diPL (I), biFH (Cl) | Glombitza & Schmidt (1999) [32] |
| Dictyotales (Di) | ||
| Padina tetrastromatica | Cl and Br sesquiterpenes | Parameswaran et al. (1994) [45] |
| Colpomenia sinuosa | colpol (1,4 diphenylbutene 2) | Green et al. (1993) [34] |
| Ectocarpales (Ec) | ||
| Analipus japonicus | tri, tetra and pentaFC | Glombitza & Zieprath (1989) [33] |
© 2010 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
La Barre, S.; Potin, P.; Leblanc, C.; Delage, L. The Halogenated Metabolism of Brown Algae (Phaeophyta), Its Biological Importance and Its Environmental Significance. Mar. Drugs 2010, 8, 988-1010. https://doi.org/10.3390/md8040988
La Barre S, Potin P, Leblanc C, Delage L. The Halogenated Metabolism of Brown Algae (Phaeophyta), Its Biological Importance and Its Environmental Significance. Marine Drugs. 2010; 8(4):988-1010. https://doi.org/10.3390/md8040988
Chicago/Turabian StyleLa Barre, Stéphane, Philippe Potin, Catherine Leblanc, and Ludovic Delage. 2010. "The Halogenated Metabolism of Brown Algae (Phaeophyta), Its Biological Importance and Its Environmental Significance" Marine Drugs 8, no. 4: 988-1010. https://doi.org/10.3390/md8040988