The Carotenogenesis Pathway via the Isoprenoid-β-carotene Interference Approach in a New Strain of Dunaliella salina Isolated from Baja California Mexico

Abstract
:1. Introduction

2. Materials and Methods
2.1 Dunaliella salina strain and Carotenogenesis
2.2 Inhibitors of Carotenogenesis
2.3 Growth and pigments
2.4. Protein measurements
2.5 Cellular RNA
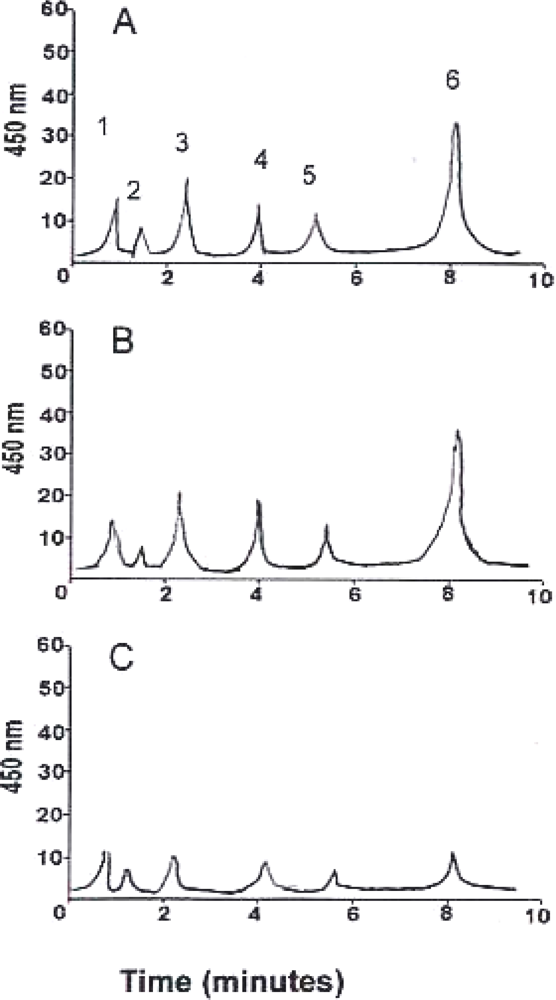
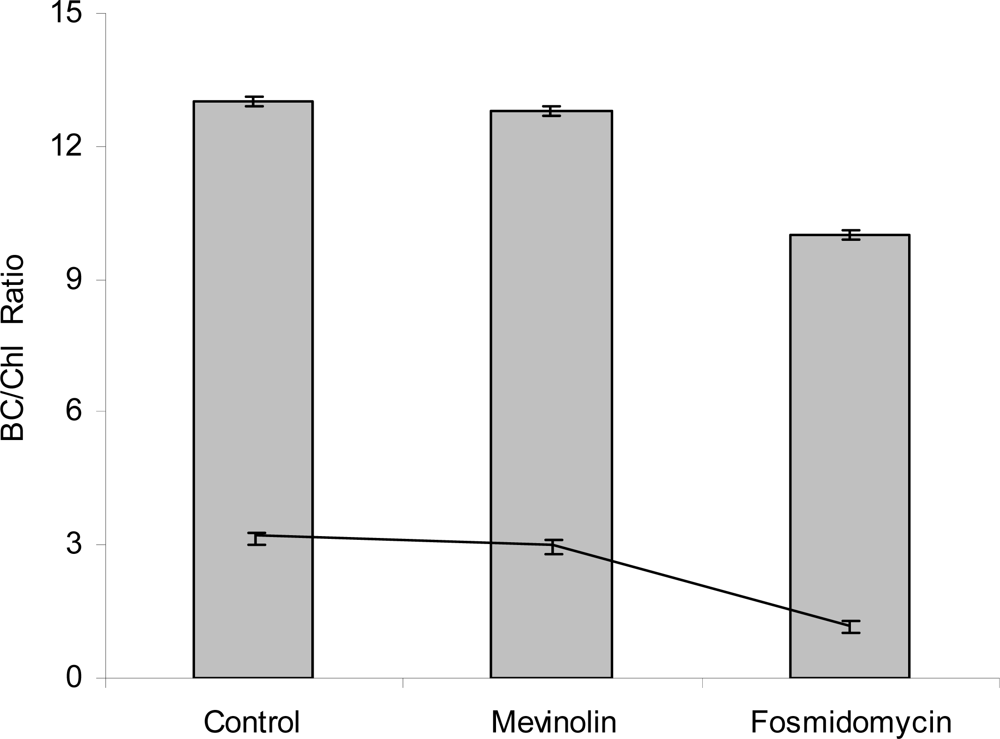
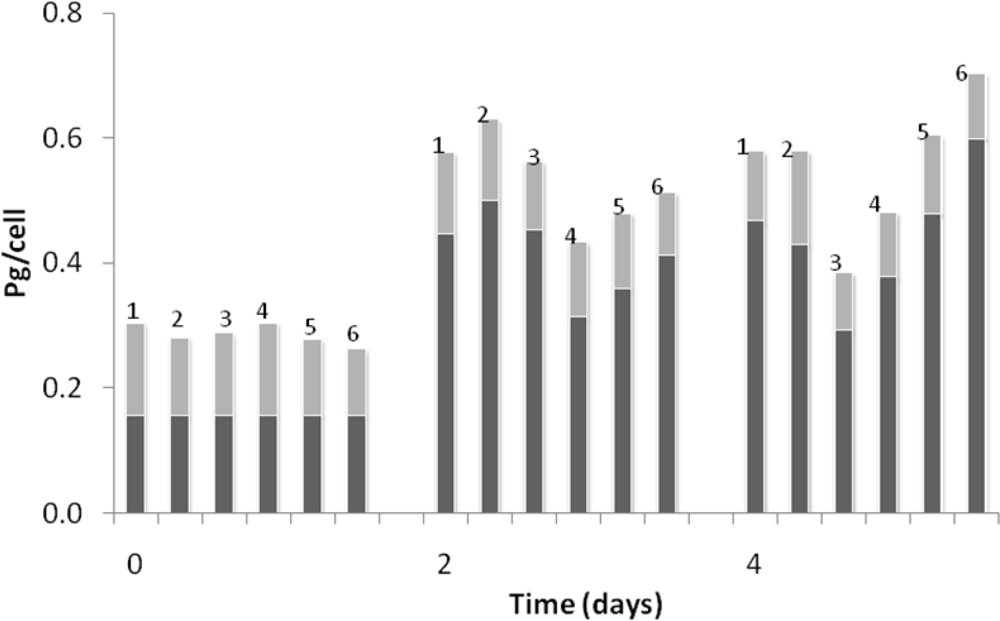
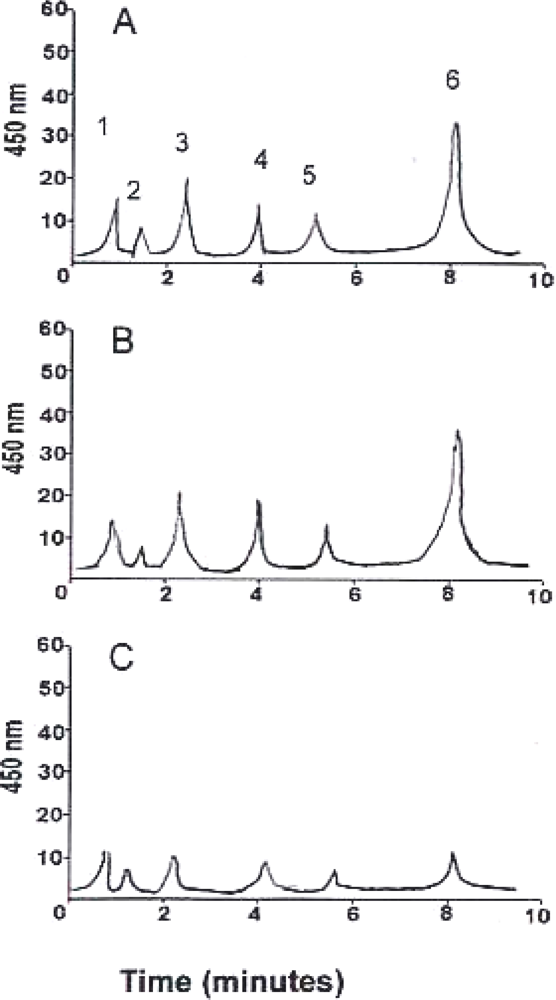
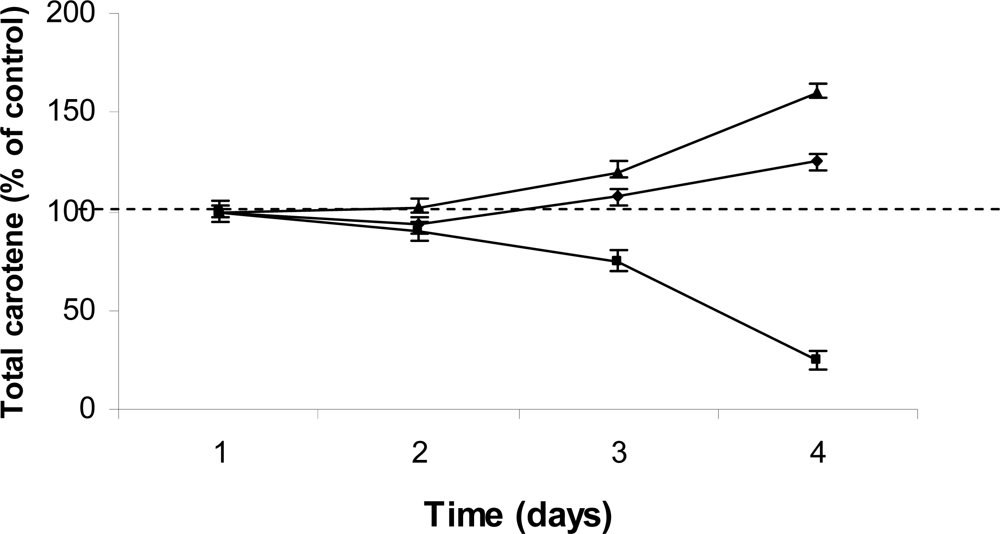
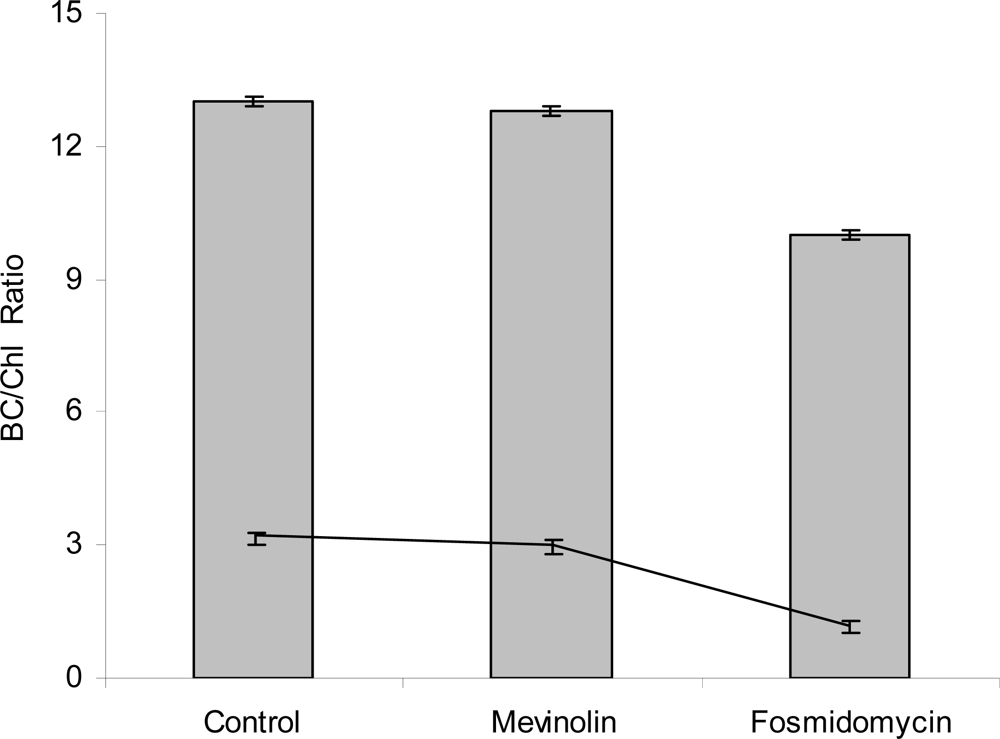
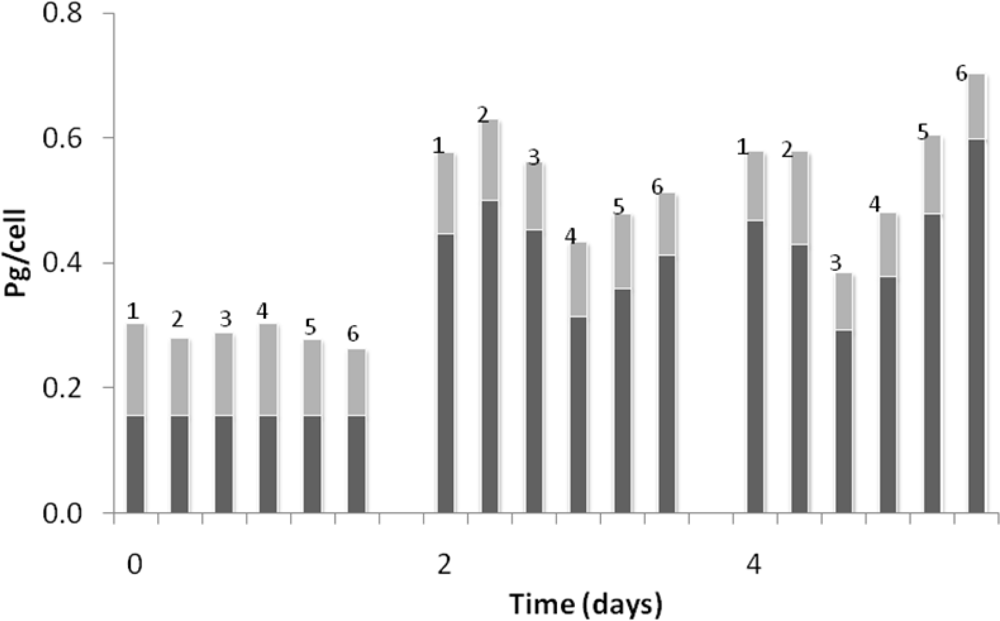
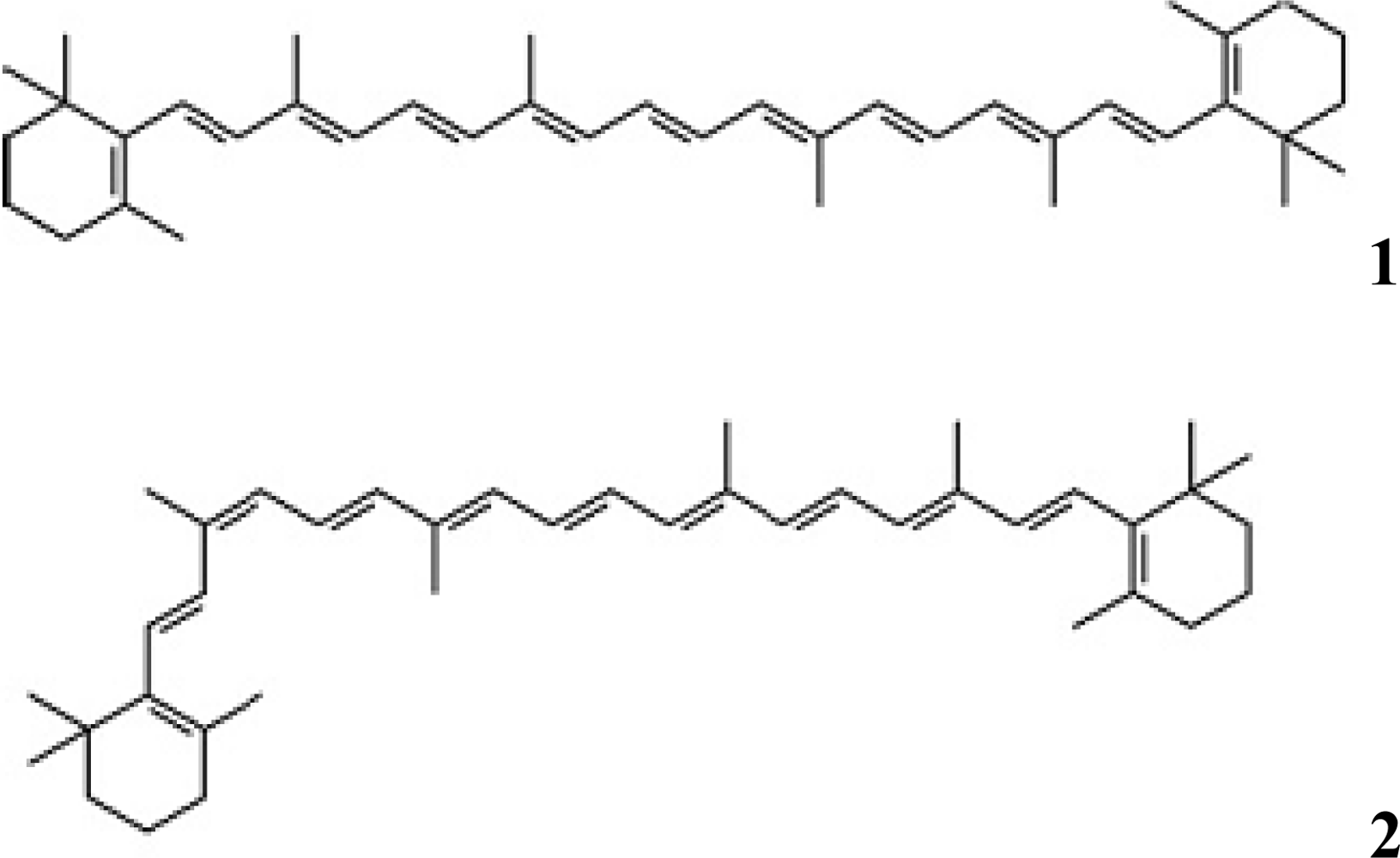
3. Results and Discussion
Acknowledgments
References and Notes
- Ben-Amotz, A. Effect of irradiance and nutrient deficiency on the chemical composition of Dunaliella bardawil. J. Plant. Physiol 1987, 131, 479–487. [Google Scholar] [CrossRef]
- Borowitzka, M. Microalgae as source of pharmaceuticals and other biologically active compounds. J. Appl. Phycol 1995, 7, 3–15. [Google Scholar] [CrossRef]
- Orset, SCH; Young, AJ. Exposure of low irradiance favors the synthesis of 9-cis β, β-carotene in Dunaliella salina (Teod.). Plant Physiol 2000, 122, 609–617. [Google Scholar] [CrossRef] [PubMed]
- Ben-Amotz, A; Shaish, A. β-carotene biosynthesis. In Dunaliella: Physiology, Biochemistry and Biotechnology; Avron, M, Ben-Amotz, A, Eds.; CRC Press, Boca Raton: Florida, USA, 1992; pp. 205–216. [Google Scholar]
- Borowitzka, M; Borowitzka, L; Kessly, D. Effects of salinity increase on carotenoid accumulation in the green alga Dunaliella salina. J. Appl. Phycol 1990, 2, 111–119. [Google Scholar] [CrossRef]
- Paniagua-Michel, J; Dujardin, E; Sironval, C. Growth of Dunaliella bardawil under carotenogenic conditions. J. Mar. Biotech 1995, 2, 101–104. [Google Scholar]
- Ben-Amotz, A; Avron, M. On the factors which determine massive beta-carotene accumulation in the halotolerant alga Dunaliella bardawil. Plant Physiol 1983, 72, 593–597. [Google Scholar] [CrossRef] [PubMed]
- Rohmer, M. The discovery of a mevalonate-independent pathway for isoprenoid biosynthesis in bacteria, algae and higher plants. Nat. Prod. Rep 1999, 16, 565–574. [Google Scholar] [CrossRef] [PubMed]
- Lange, BM; Rujan, T; Martin, W; Croteau, R. Isoprenoid biosynthesis: the evolution of two ancient and distinct pathways across genomes. Proc. Nat. Acad. Sci. USA 2000, 97, 13172–13177. [Google Scholar] [CrossRef] [PubMed]
- Schwender, J; Gemunden, C; Lichtenthaler, H. Chlorophyta exclusively use the 1-deoxyxylulose 5-phosphate/2-C-methylerythritol 4-phosphate pathway for the biosynthesis of isoprenoids. Planta 2001, 212, 416–423. [Google Scholar] [CrossRef] [PubMed]
- Gutierrez-Millan, LE. Desoxyribonucleic acid, protein and pigments during induced carotenogenesis in a new strain of Dunaliella salina (Chlorophyta isolated from a hipersaline coastal lagoon of Baja California. MSc Thesis, CICESE Mexico, 1996; 1–81. [Google Scholar]
- Olmos-Soto, J; Paniagua-Michel, J; Contreras-Flores, R; Trujillo-Valle, ML. Molecular identification of β-carotene hyper-producer strains of Dunaliella from saline environments using species-specific oligonucleotides. Biotechol. Letters 2002, 24, 365–369. [Google Scholar] [CrossRef]
- Ben-Amotz, A; Avron, M. On the Factors which Determine Massive β-Carotene Accumulation in the Halotolerant Alga Dunaliella bardawil. Plant Physiol 1983, 72, 593–597. [Google Scholar] [CrossRef] [PubMed]
- Hagen, C; Grünewald, K. Fosmidomycin as an inhibitor of the non-mevalonate terpenoid pathway depresses synthesis of secondary carotenoids in flagellates of the green alga Haematococcus pluvialis. J Appl. Bot 2000, 74, 37–140. [Google Scholar]
- Khachik, F; Beecher, GR; Goli, MB. Separation, identification, and quantification of carotenoids in fruits, vegetables and human plasma by high performance liquid chromatography. Pure Appl. Chem 1991, 63, 71–80. [Google Scholar] [CrossRef]
- Bradford, MM. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Analyt. Biochem 1976, 72, 248–294. [Google Scholar] [CrossRef] [PubMed]
- Berges, J; Fisher, A; Harrison, P. A comparison of Lowry, Bradford and Smith protein assays using different protein standards and protein isolated from the marine diatom Thalassiosira pseudonana. Mar. Biol 1993, 115, 187–193. [Google Scholar] [CrossRef]
- Sambrook, J; Fritsh, EF; Maniatis, T. Molecular cloning: a laboratory manual; Cold Spring Harbor laboratory Press: New York; NY, USA, 1989. [Google Scholar]
- Bajguz, A; Asami, T. Effects of brassinazole, an inhibitor of brassinosteroid biosynthesis, on light and dark grown Chlorella vulgaris. Planta 2004, 218, 869–877. [Google Scholar] [CrossRef] [PubMed]
- Lichtenthaler, HK. The 1-deoxy-D-xylulose-5-phosphate pathway of isoprenoid biosynthesis in plants. Annu Rev Plant Physiol Plant Mol Biol 1999, 50, 47–65. [Google Scholar] [CrossRef] [PubMed]
- Lange, M; Ketchum, EB; Croteau, RB. Isoprenoid biosynthesis. Plant Physiol 2001, 127, 305–314. [Google Scholar] [CrossRef] [PubMed]
- Cvejic, J; Rohmer, M. CO2 as main carbon source for isoprenoid biosynthesis via the mevalonate-independent methylerythritol 4-phosphate route in the marine diatoms Phaeodactylum tricornutum and Nitzschia ovalis. Phytochem 2000, 53, 21–28. [Google Scholar] [CrossRef]
- Sirev, R; Levine, RP. Transcription and translation of carotenoid synthesis in Chlamydomonas reinhardtii. Planta 1973, 111, 73–84. [Google Scholar] [CrossRef]
- Disch, A; Schwender, J; Müller, C; Lichtenthaler, H; Rohmer, M. Distribution of the mevalonate and glyceraldehyde phosphate/pyruvate pathways for isoprenoid biosynthesis in unicellular algae and the cyanobacterium Synechocystis PCC 6714. Biochem J 1998, 333, 381–388. [Google Scholar] [PubMed]
- Laule, O; Fürholz, A; Chang, H; Zhu, T; Wang, X; Heifetz, P; Gruissem, W; Lange, B. Crosstalk between cytosolic and plastidial pathways of isoprenoid biosynthesis in. Arabidopsis thaliana Proc Nat Acad Sci USA 2003, 100, 6866–6871. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cellular growth (N × 106 cells. ml−1) Days | |||||
|---|---|---|---|---|---|
| 1 | 2 | 7 | |||
| Control (No Inhibitor) | 1.1 ± 0.02 | 1.4 ± 0.01 | 1.6 ± 0.03 | ||
| Mev (μM) | 0.5 | 1.1 ± 0.01 | 1.2 ± 0.02 | 1.6 ± 0.01 | |
| 1 | 1.2 ± 0.01 | 1.3 ± 0.02 | 1.4 ± 0.02 | ||
| 2 | 1.1 ± 0.04 | 1.3 ± 0.01 | 1.5 ± 0.02 | ||
| Fos (μM) | 50 | 1.1 ± 0.02 | 1.1 ± 0.01 | 0.5 ± 0.03 | |
| 100 | 1.2 ± 0.02 | 1.0 ± 0.03 | 0.4 ± 0.01 | ||
| 150 | 1.1 ± 0.03 | 1.0 ± 0.02 | 0.4 ± 0.02 | ||
| 200 | 0.8 ± 0.01 | 0.8 ± 0.02 | 0.3 ± 0.01 | ||
| Chlorophyll (pg. cell−1) | |||||
| Control (No Inhibitor) | 0.4 ± 0.01 | 0.5 ± 0.02 | 0.6 ± 0.02 | ||
| Mev (μM) | 0.5 | 0.4 ± 0.02 | 0.4 ± 0.03 | 0.5 ± 0.02 | |
| 1 | 0.4 ± 0.02 | 0.5 ± 0.01 | 0.5 ± 0.02 | ||
| 2 | 0.5 ± 0.02 | 0.5 ± 0.01 | 0.5 ± 0.03 | ||
| Fos (μM) | 50 | 0.4 ± 0.01 | 0.4 ± 0.02 | 0.3 ± 0.01 | |
| 100 | 0.4 ± 0.02 | 0.4 ± 0.01 | 0.1 ± 0.02 | ||
| 150 | 0.3 ± 0.02 | 0.3 ± 0.01 | 0.1 ± 0.01 | ||
| 200 | 0.3 ± 0.01 | 0.3 ± 0.02 | 0.1 ± 0.01 | ||
| Total carotene (pg. cell−1) | |||||
| Control (No Inhibitor) | 1.2 ± 0.03 | 1.5 ± 0.02 | 1.7 ± 0.01 | ||
| Mev (μM) | 0.5 | 1.3 ± 0.02 | 1.4 ± 0.01 | 1.7 ± 0.02 | |
| 1 | 1.3 ± 0.04 | 1.5 ± 0.02 | 1.8 ± 0.01 | ||
| 2 | 1.5 ± 0.02 | 1.6 ± 0.03 | 1.8 ± 0.02 | ||
| Fos (μM) | 50 | 1.1 ± 0.01 | 1.1 ± 0.03 | 0.8 ± 0.01 | |
| 100 | 0.9 ± 0.02 | 0.9 ± 0.02 | 0.4 ± 0.01 | ||
| 150 | 0.7 ± 0.02 | 0.6 ± 0.01 | 0.3 ± 0.03 | ||
| 200 | 0.4 ± 0.01 | 0.4 ± 0.01 | 0.2 ± 0.01 | ||
© 2009 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Paniagua-Michel, J.; Capa-Robles, W.; Olmos-Soto, J.; Gutierrez-Millan, L.E. The Carotenogenesis Pathway via the Isoprenoid-β-carotene Interference Approach in a New Strain of Dunaliella salina Isolated from Baja California Mexico. Mar. Drugs 2009, 7, 45-56. https://doi.org/10.3390/md7010045
Paniagua-Michel J, Capa-Robles W, Olmos-Soto J, Gutierrez-Millan LE. The Carotenogenesis Pathway via the Isoprenoid-β-carotene Interference Approach in a New Strain of Dunaliella salina Isolated from Baja California Mexico. Marine Drugs. 2009; 7(1):45-56. https://doi.org/10.3390/md7010045
Chicago/Turabian StylePaniagua-Michel, J., Willian Capa-Robles, Jorge Olmos-Soto, and Luis Enrique Gutierrez-Millan. 2009. "The Carotenogenesis Pathway via the Isoprenoid-β-carotene Interference Approach in a New Strain of Dunaliella salina Isolated from Baja California Mexico" Marine Drugs 7, no. 1: 45-56. https://doi.org/10.3390/md7010045