Immunomodulatory Effects of Domoic Acid Differ Between In vivo and In vitro Exposure in Mice

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
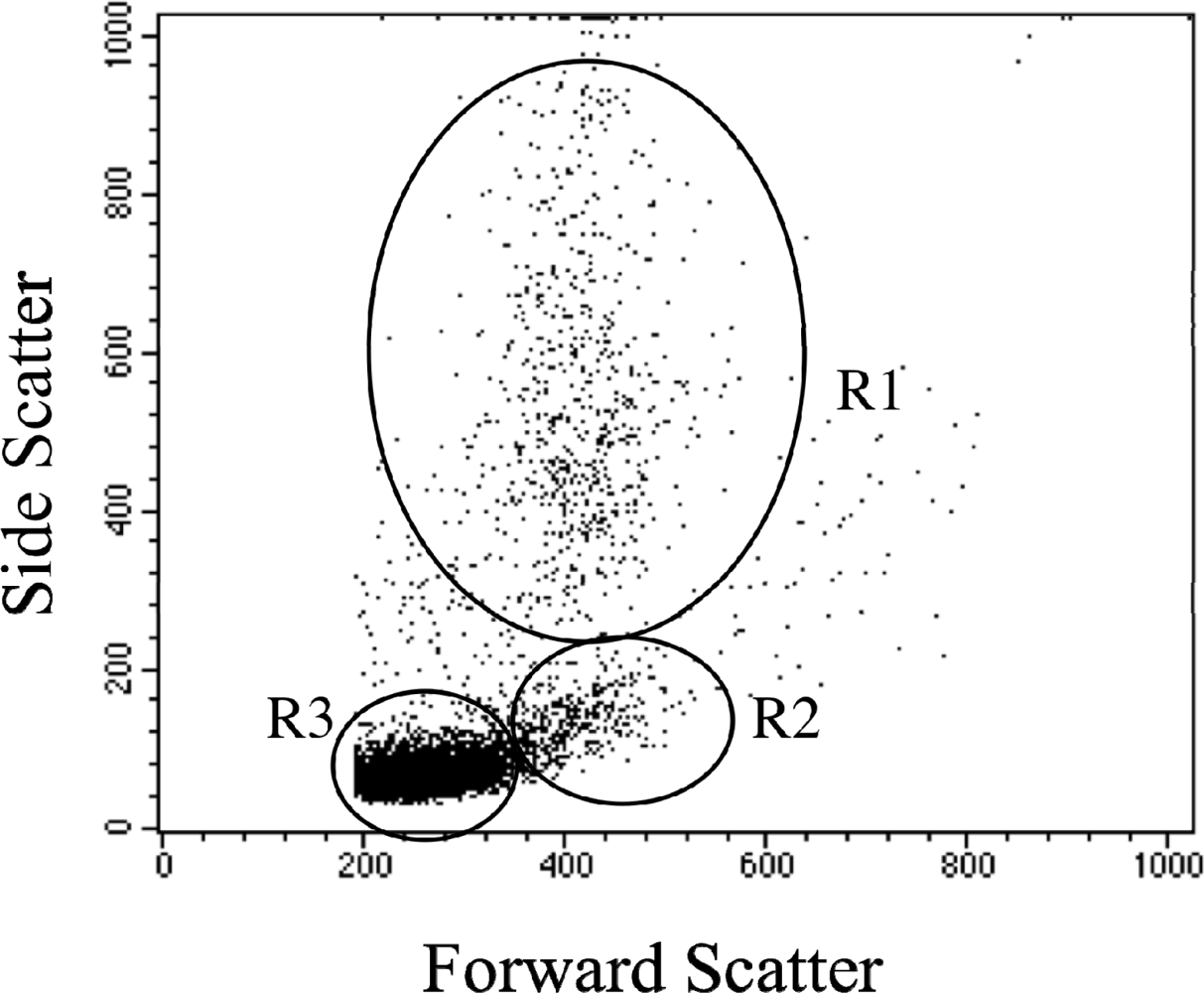
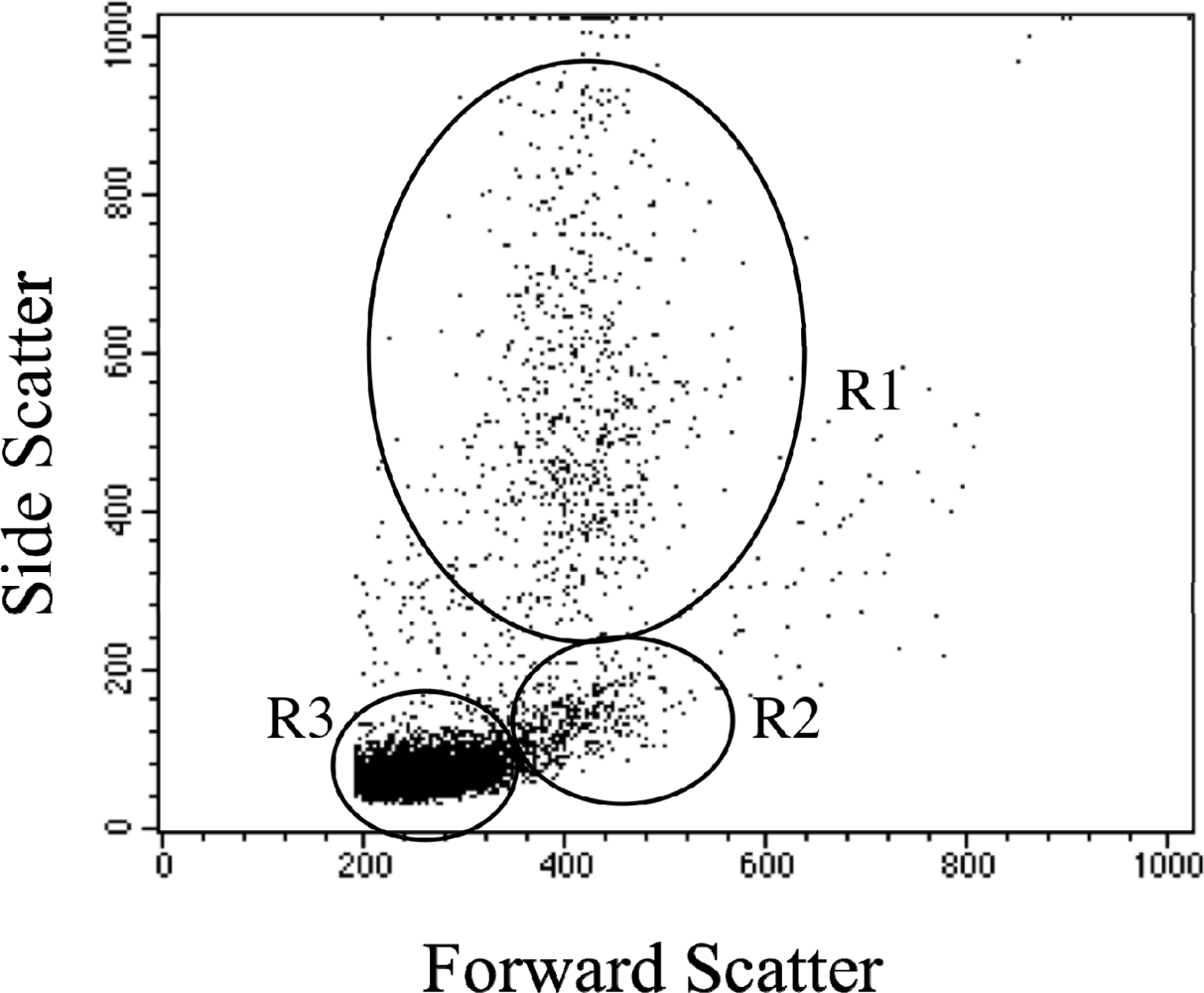
2.1 Leukocyte flow cytometry profile
2.2 In vivo clinical signs
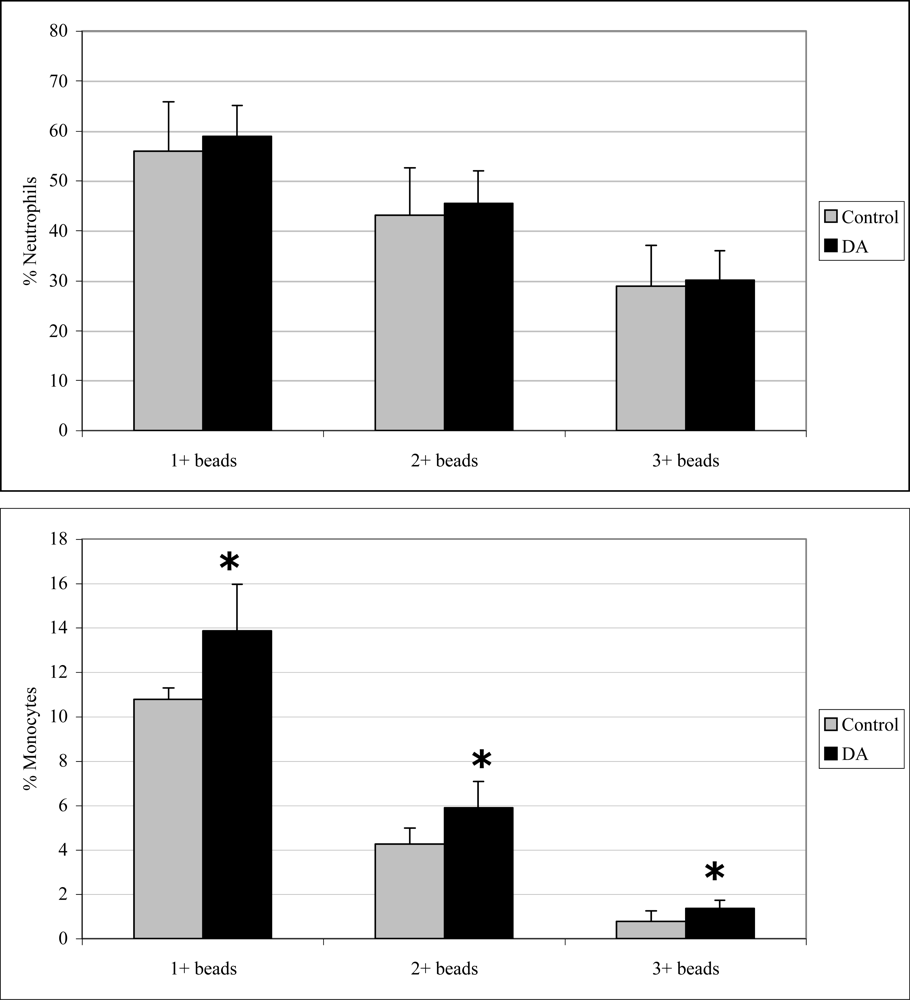
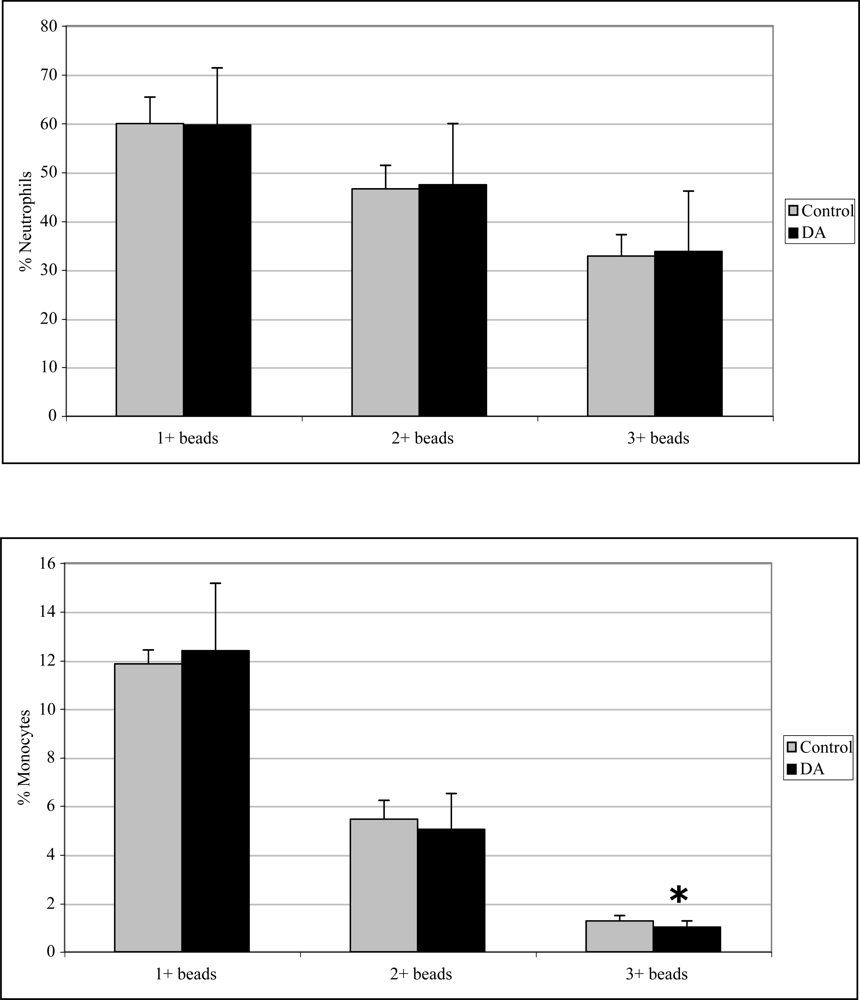
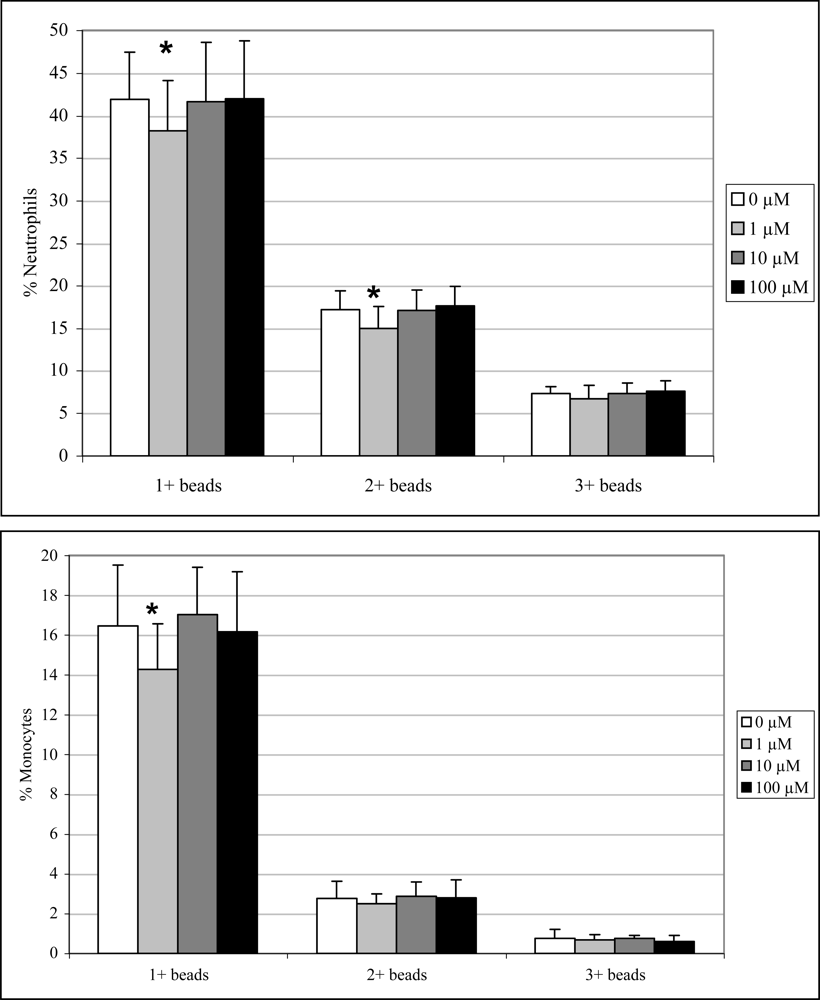
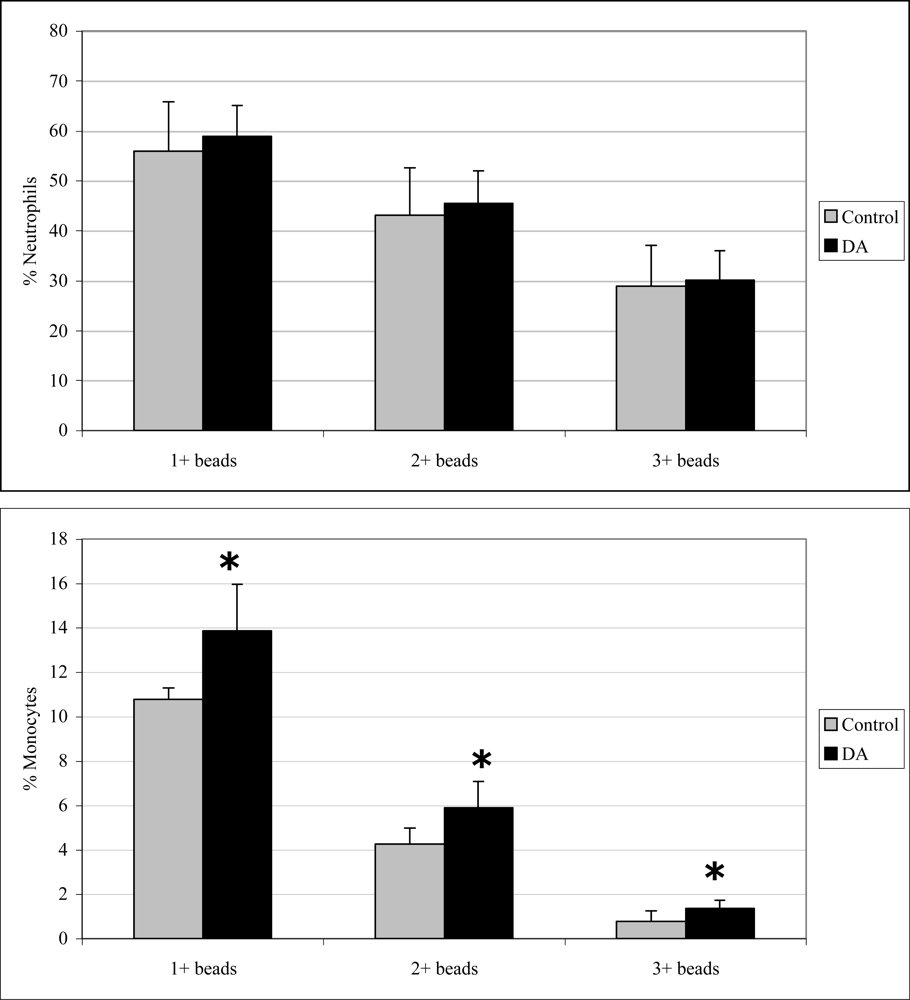
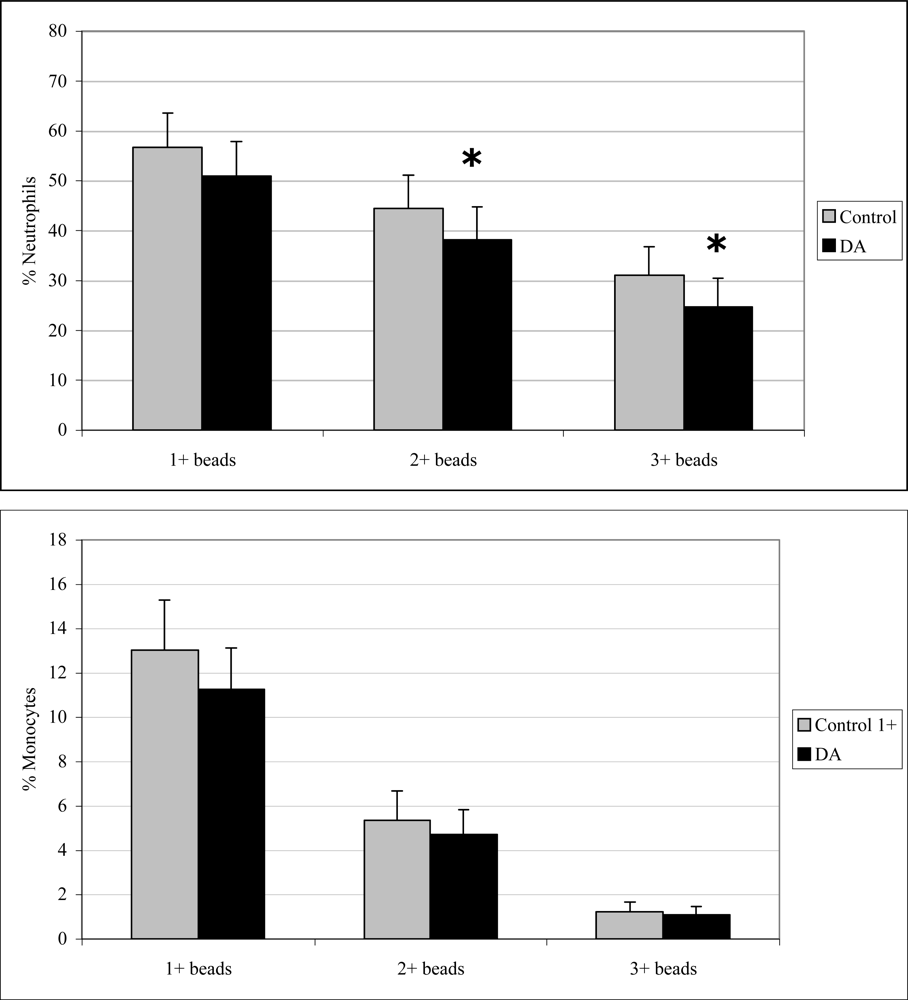
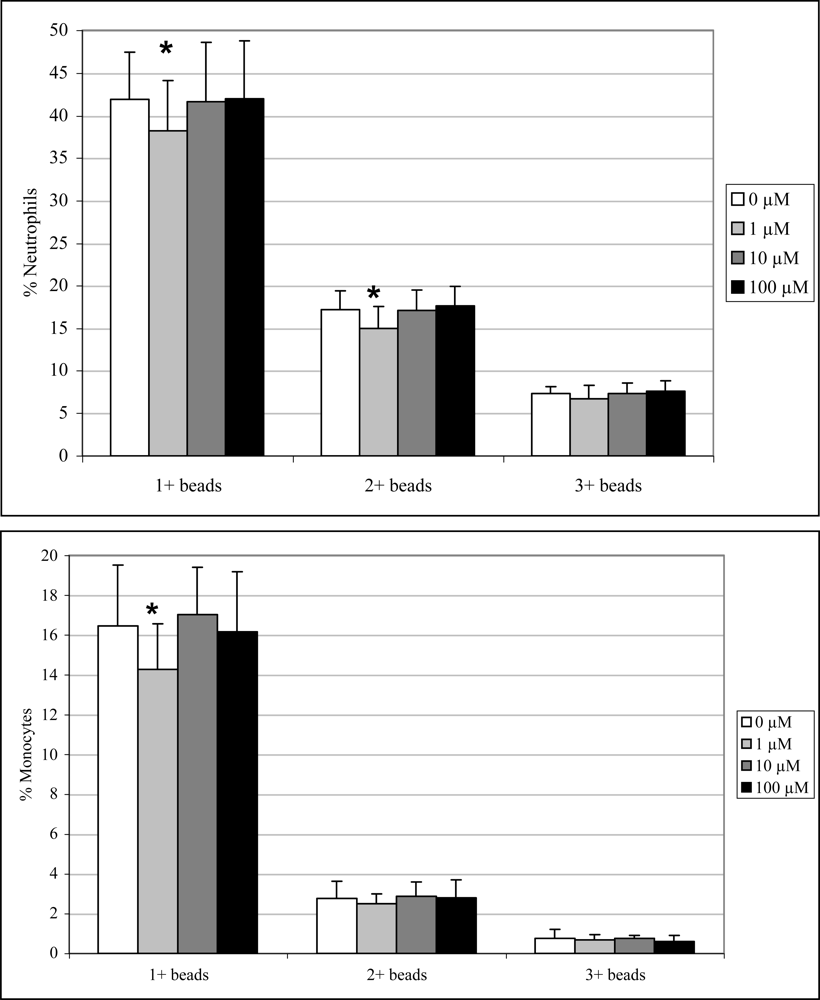
2.3 In vivo phagocytosis
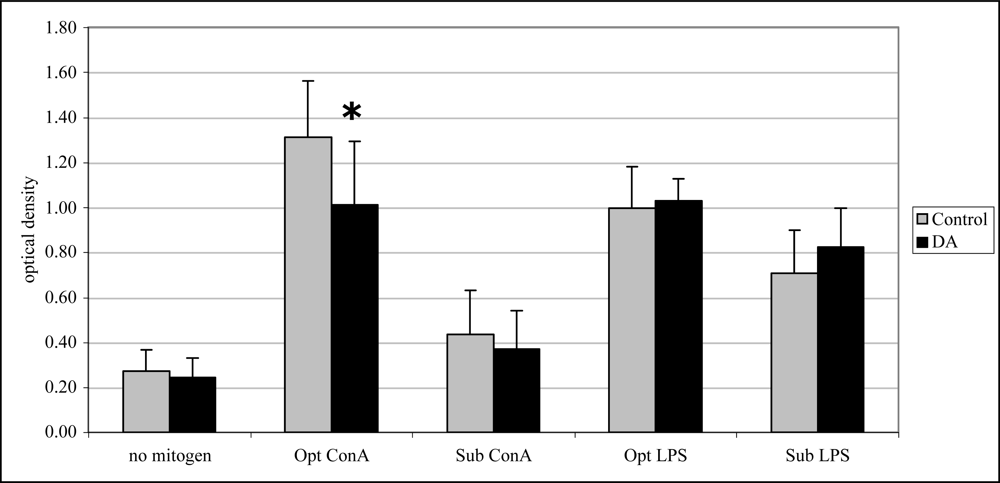
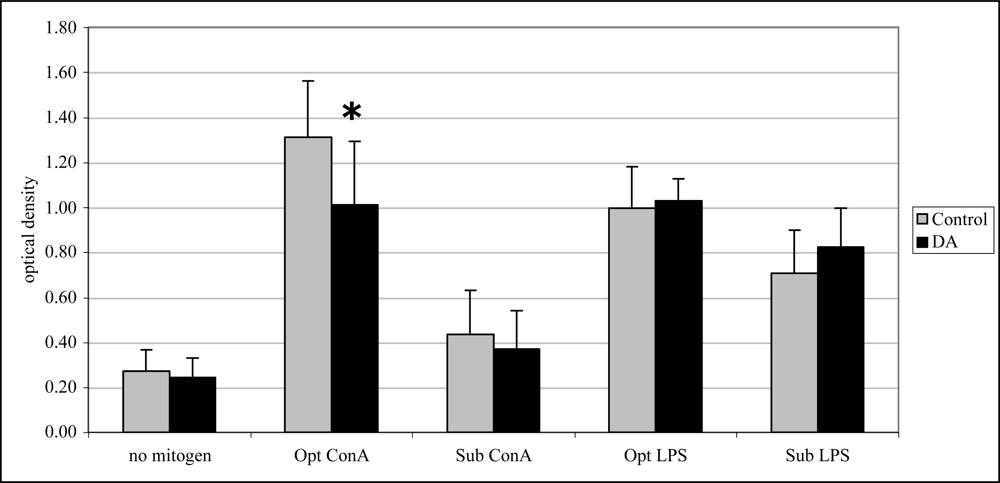
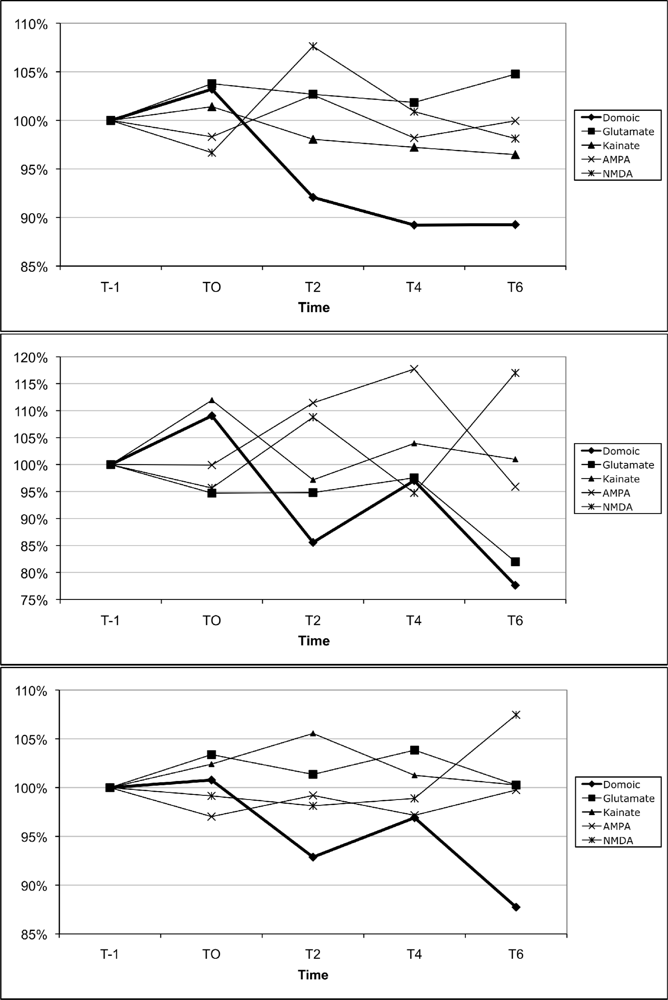
2.4 In vivo lymphocyte proliferation
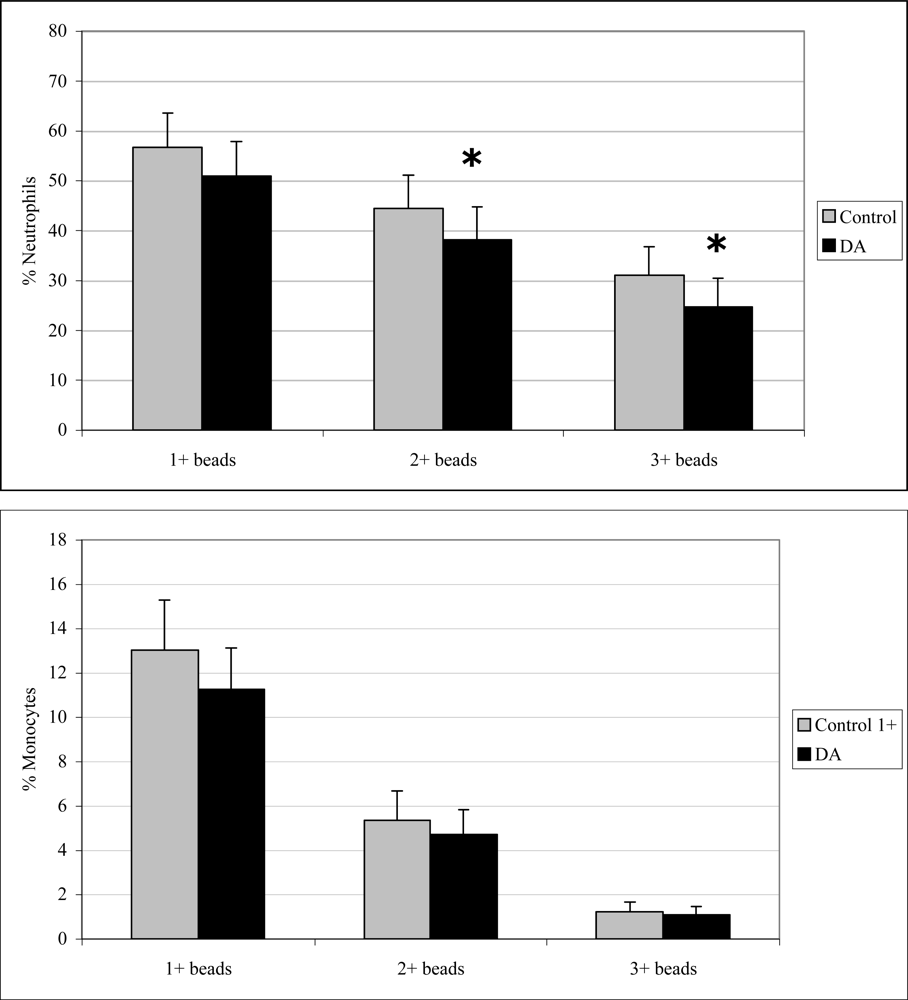
2.5 In vitro phagocytosis
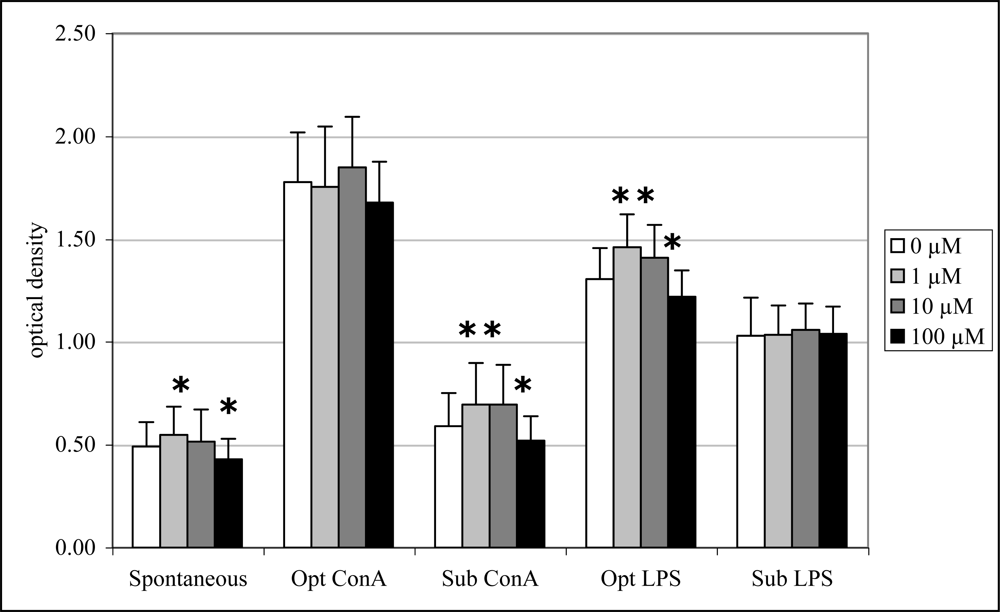
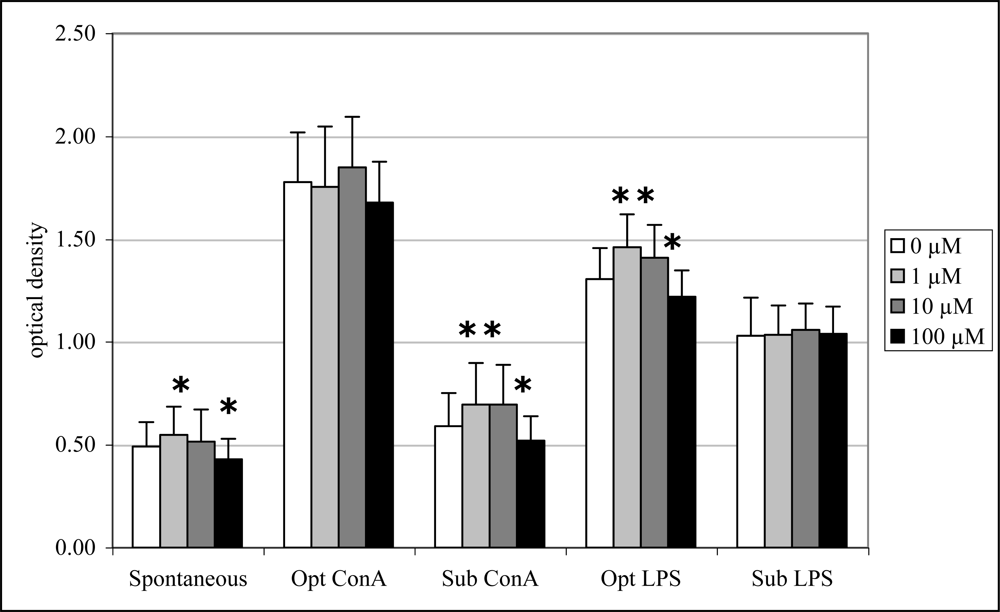
2.6 In vitro lymphocyte proliferation
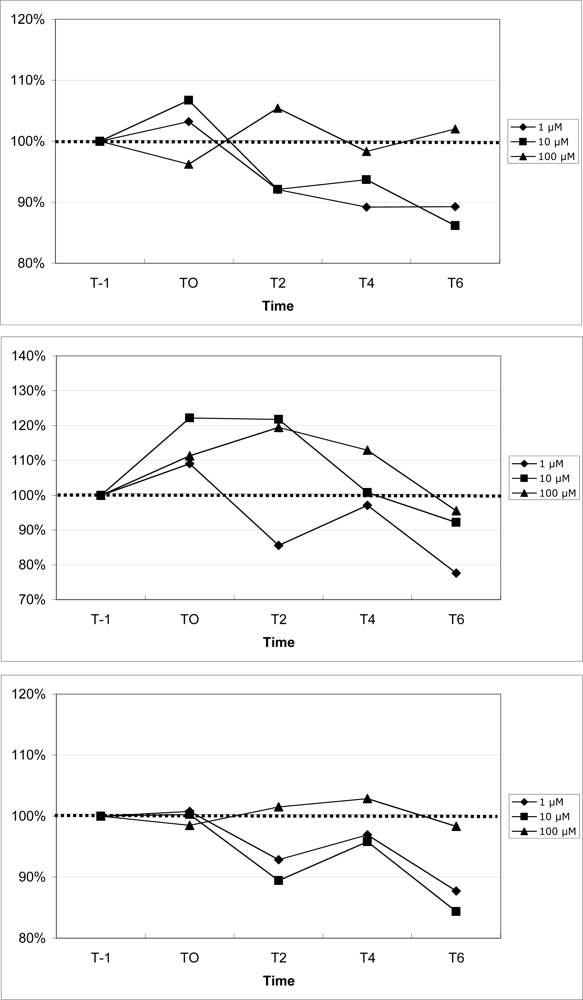
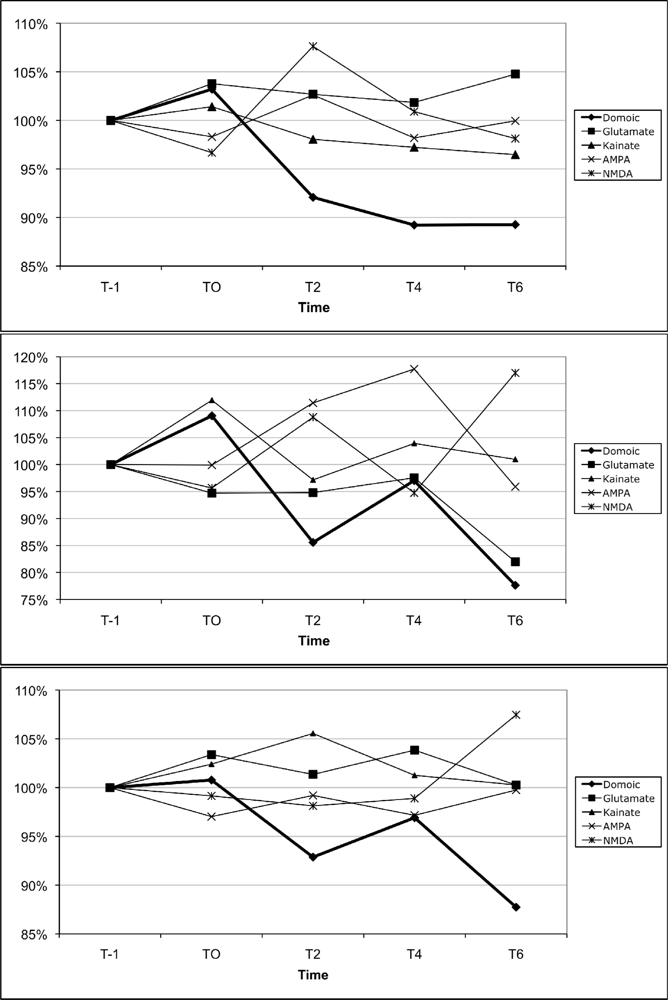
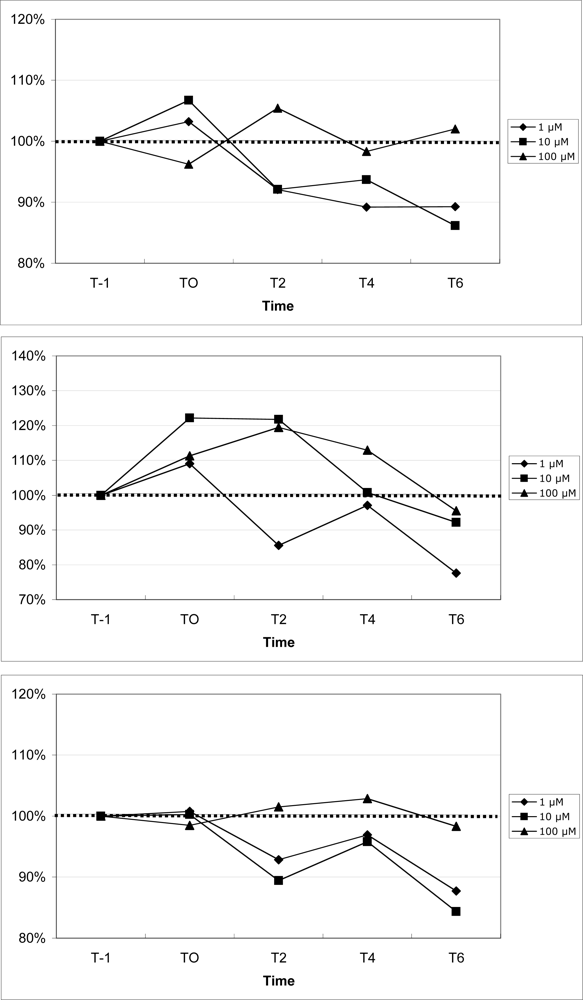
2.7 Mobilization of calcium in blood leukocytes
3. Discussion and Conclusion
3.1 In vivo exposure
3.2 In vitro exposure
3.3 Potential mechanism(s)/pathways(s) mediating in vitro DA-induced immunotoxicty
3.4 Differences between direct vs indirect effects
3.5. Relevancy of immune modulation
3.6 Conclusion
4. Experimental Section
4.1 In vivo exposure
4.2 Phagocytosis
4.3. Lymphocyte proliferation
4.4 In vitro exposure
4.5 Cytosolic calcium mobilization
4.6 Statistics
Acknowledgments
References and Notes
- Van Dolah, FM. Marine algal toxins: origins, health effects, and their increased occurrence. Environ. Health Perspect. 2000, 108(Suppl 1), 133–141. [Google Scholar]
- Sayer, A; Hu, Q; Bourdelais, AJ; Baden, DG; Gibson, JE. The effect of brevenal on brevetoxin-induced DNA damage in human lymphocytes. Arch. Toxicol 2005, 79(11), 683–688. [Google Scholar]
- Shen, PP; Zhao, SW; Zheng, WJ; Hua, ZC; Shi, Q; Liu, ZT. Effects of cyanobacteria bloom extract on some parameters of immune function in mice. Toxicol. Lett 2003, 143(1), 27–36. [Google Scholar]
- Walsh, CJ; Luer, CA; Noyes, DR. . Effects of environmental stressors on lymphocyte proliferation in Florida manatees, Trichechus manatus latirostris. Vet. Immunol. Immunopathol 2005, 103(3–4), 247–256. [Google Scholar]
- Witkowski, JM; Siebert, J; Lukaszuk, K; Trawicka, L. Comparison of effect of a panel of membrane channel blockers on the proliferative, cytotoxic and cytoadherence abilities of human peripheral blood lymphocytes. Immunopharmacol 1993, 26(1), 53–63. [Google Scholar]
- Dizer, H; Fischer, B; Harabawy, AS; Hennion, MC; Hansen, PD. Toxicity of domoic acid in the marine mussel Mytilus edulis. Aquat. Toxicol 2001, 55(3–4), 149–156. [Google Scholar]
- Jones, TO; Whyte, JN; Ginther, NG; Townsend, LD; Iwama, GK. Haemocyte changes in the Pacific oyster, Crassostrea gigas, caused by exposure to domoic acid in the diatom Pseudonitzschia pungens f. multiseries. Toxicon 1995, 33(3), 347–353. [Google Scholar]
- Jeffery, B; Barlow, T; Moizer, K; Paul, S; Boyle, C. Amnesic shellfish poison. Food Chem. Toxicol 2004, 42(4), 545–557. [Google Scholar]
- Perl, TM; Bedard, L; Kosatsky, T; Hockin, JC; Todd, EC; Remis, RS. An outbreak of toxic encephalopathy caused by eating mussels contaminated with domoic acid. N. Engl. J. Med 1990, 322(25), 1775–1780. [Google Scholar]
- Lefebvre, KA; Powell, CL; Busman, M; Doucette, GJ; Moeller, PD; Silver, JB; Miller, PE; Hughes, MP; Singaram, S; Silver, MW; Tjeerdema, RS. Detection of domoic acid in northern anchovies and California sea lions associated with an unusual mortality event. Nat. Toxins 1999, 7(3), 85–92. [Google Scholar]
- Scholin, CA; Gulland, F; Doucette, GJ; Benson, S; Busman, M; Chavez, FP; Cordaro, J; DeLong, R; De Vogelaere, A; Harvey, J; Haulena, M; Lefebvre, K; Lipscomb, T; Loscutoff, S; Lowenstine, LJ; Marin, R., 3rd; Miller, PE; McLellan, WA; Moeller, PD; Powell, CL; Rowles, T; Silvagni, P; Silver, M; Spraker, T; Trainer, V; Van Dolah, FM. Mortality of sea lions along the central California coast linked to a toxic diatom bloom. Nature 2000, 403(6765), 80–84. [Google Scholar]
- Gulland, FM; Haulena, M; Fauquier, D; Langlois, G; Lander, ME; Zabka, T; Duerr, R. Domoic acid toxicity in Californian sea lions (Zalophus californianus): clinical signs, treatment and survival. Vet. Rec 2002, 150(15), 475–480. [Google Scholar]
- Hampson, DR; Manalo, JL. The activation of glutamate receptors by kainic acid and domoic acid. Nat. Toxins 1998, 6(3–4), 153–158. [Google Scholar]
- Berman, FW; LePage, KT; Murray, TF. Domoic acid neurotoxicity in cultured cerebellar granule neurons is controlled preferentially by the NMDA receptor Ca2+ influx pathway. Brain Res 2002, 924(1), 20–29. [Google Scholar]
- Pulido, OM. Domoic acid toxicologic pathology: a review. Mar Drugs 2008, 6(2), 180–219. [Google Scholar]
- Boldyrev, AA; Carpenter, DO; Johnson, P. Emerging evidence for a similar role of glutamate receptors in the nervous and immune systems. J. Neurochem 2005, 95(4), 913–918. [Google Scholar]
- Skerry, TM; Genever, PG. Glutamate signalling in non-neuronal tissues. Trends Pharmacol. Sci 2001, 22(4), 174–181. [Google Scholar]
- Gill, S; Veinot, J; Kavanagh, M; Pulido, O. Human heart glutamate receptors - implications for toxicology, food safety, and drug discovery. Toxicol. Pathol 2007, 35(3), 411–417. [Google Scholar]
- Gill, SS; Mueller, RW; McGuire, PF; Pulido, OM. Potential target sites in peripheral tissues for excitatory neurotransmission and excitotoxicity. Toxicol. Pathol 2000, 28(2), 277–284. [Google Scholar]
- Gill, SS; Pulido, OM. Glutamate receptors in peripheral tissues: current knowledge, future research, and implications for toxicology. Toxicol. Pathol 2001, 29(2), 208–223. [Google Scholar]
- Dickman, KG; Youssef, JG; Mathew, SM; Said, SI. Ionotropic glutamate receptors in lungs and airways: molecular basis for glutamate toxicity. Am. J. Respir. Cell Mol. Biol 2004, 30(2), 139–144. [Google Scholar]
- Boldyrev, AA; Kazey, VI; Leinsoo, TA; Mashkina, AP; Tyulina, OV; Johnson, P; Tuneva, JO; Chittur, S; Carpenter, DO. Rodent lymphocytes express functionally active glutamate receptors. Biochem. Biophys. Res. Commun 2004, 324(1), 133–139. [Google Scholar]
- Ganor, Y; Besser, M; Ben-Zakay, N; Unger, T; Levite, M. Human T cells express a functional ionotropic glutamate receptor GluR3, and glutamate by itself triggers integrin-mediated adhesion to laminin and fibronectin and chemotactic migration. J. Immunol 2003, 170(8), 4362–4372. [Google Scholar]
- Lombardi, G; Dianzani, C; Miglio, G; Canonico, PL; Fantozzi, R. Characterization of ionotropic glutamate receptors in human lymphocytes. Br. J. Pharmacol 2001, 133(6), 936–944. [Google Scholar]
- Haas, HS; Pfragner, R; Siegl, V; Ingolic, E; Heintz, E; Schauenstein, K. Glutamate receptor-mediated effects on growth and morphology of human histiocytic lymphoma cells. Int. J. Oncol 2005, 27(3), 867–874. [Google Scholar]
- Luster, MI; Portier, C; Pait, DG; Rosenthal, GJ; Germolec, DR; Corsini, E; Blaylock, BL; Pollock, P; Kouchi, Y; Craig, W; White, KL; Munson, AE; Comment, CE. Risk assessment in immunotoxicology. II. Relationships between immune and host resistance tests. Fundam. Appl. Toxicol 1993, 21(1), 71–82. [Google Scholar]
- Luster, MI; Portier, C; Pait, DG; White, KL, Jr; Gennings, C; Munson, AE; Rosenthal, GJ. Risk assessment in immunotoxicology. I. Sensitivity and predictability of immune tests. Fundam. Appl. Toxicol 1992, 18(2), 200–210. [Google Scholar]
- Hymery, N; Sibiril, Y; Parent-Massin, D. Improvement of human dendritic cell culture for immunotoxicological investigations. Cell Biol. Toxicol 2006, 22(4), 243–255. [Google Scholar]
- Wetmore, L; Nance, DM. Differential and sex-specific effects of kainic acid and domoic acid lesions in the lateral septal area of rats on immune function and body weight regulation. Exp. Neurol 1991, 113(2), 226–236. [Google Scholar]
- Peng, YG; Ramsdell, JS. Brain Fos induction is a sensitive biomarker for the lowest observed neuroexcitatory effects of domoic acid. Fundam. Appl. Toxicol 1996, 31(2), 162–168. [Google Scholar]
- Maucher, JM; Ramsdell, JS. Ultrasensitive detection of domoic acid in mouse blood by competitive ELISA using blood collection cards. Toxicon 2005, 45(5), 607–613. [Google Scholar]
- Ehrchen, J; Steinmuller, L; Barczyk, K; Tenbrock, K; Nacken, W; Eisenacher, M; Nordhues, U; Sorg, C; Sunderkotter, C; Roth, J. Glucocorticoids induce differentiation of a specifically activated, anti-inflammatory subtype of human monocytes. Blood 2007, 109(3), 1265–1274. [Google Scholar]
- Weller, RO; Engelhardt, B; Phillips, MJ. Lymphocyte targeting of the central nervous system: a review of afferent and efferent CNS-immune pathways. Brain Pathol 1996, 6(3), 275–288. [Google Scholar]
- Gill, SS; Hou, Y; Ghane, T; Pulido, OM. Regional susceptibility to domoic acid in primary astrocyte cells cultured from the brain stem and hippocampus. Mar. Drugs 2008, 6(1), 25–38. [Google Scholar]
- Calabrese, E. Hormesis: Why it is Important to Toxicology and Toxicologists. Environ. Toxicol. Chem. 2008, 1451–1474. [Google Scholar]
- Calabrese, E. Hormetic Dose-Response Relationships in Immunology: Occurrence, Quantitative Features of the Dose Response, Mechanistic Foundations, and Clinical Implications. Crit. Rev. Toxicol 2005, 35(2–3), 89–295. [Google Scholar]
- Mori, C; Morsey, B; Levin, M; Nambiar, PR; De Guise, S. Immunomodulatory effects of in vitro exposure to organochlorines on T-cell proliferation in marine mammals and mice. J. Toxicol. Environ. Health A 2006, 69(4), 283–302. [Google Scholar]
- Kannurpatti, SS; Joshi, PG; Joshi, NB. Calcium sequestering ability of mitochondria modulates influx of calcium through glutamate receptor channel. Neurochem. Res 2000, 25(12), 1527–1536. [Google Scholar]
- Peng, TI; Greenamyre, JT. Privileged access to mitochondria of calcium influx through N-methyl-D-aspartate receptors. Mol. Pharmacol 1998, 53(6), 974–980. [Google Scholar]
- Peng, TI; Jou, MJ; Sheu, SS; Greenamyre, JT. Visualization of NMDA receptor-induced mitochondrial calcium accumulation in striatal neurons. Exp. Neurol 1998, 149(1), 1–12. [Google Scholar]
- Hishikawa, T; Cheung, JY; Yelamarty, RV; Knutson, DW. Calcium transients during Fc receptor-mediated and nonspecific phagocytosis by murine peritoneal macrophages. J. Cell Biol 1991, 115(1), 59–66. [Google Scholar]
- Lew, DP; Andersson, T; Hed, J; Di Virgilio, F; Pozzan, T; Stendahl, O. Ca2+-dependent and Ca2+-independent phagocytosis in human neutrophils. Nature 1985, 315(6019), 509–511. [Google Scholar]
- Christensen, RN; Ha, BK; Sun, F; Bresnahan, JC; Beattie, MS. Kainate induces rapid redistribution of the actin cytoskeleton in ameboid microglia. J. Neurosci. Res. 2006, 84(1), 170–181. [Google Scholar]
- Reynaud, S; Duchiron, C; Deschaux, P. 3-methylcholanthrene inhibits lymphocyte proliferation and increases intracellular calcium levels in common carp (Cyprinus carpio L). Aquat. Toxicol 2003, 63(3), 319–331. [Google Scholar]
- Poulopoulou, C; Markakis, I; Davaki, P; Nikolaou, C; Poulopoulos, A; Raptis, E; Vassilopoulos, D. Modulation of voltage-gated potassium channels in human T lymphocytes by extracellular glutamate. Mol. Pharmacol 2005, 67(3), 856–867. [Google Scholar]
- Webster, JI; Tonelli, L; Sternberg, EM. Neuroendocrine regulation of immunity. Annu. Rev. Immunol 2002, 20, 125–163. [Google Scholar]
- Qiu, S; Curras-Collazo, MC. Histopathological and molecular changes produced by hippocampal microinjection of domoic acid. Neurotoxicol. Teratol 2006, 28(3), 354–362. [Google Scholar]
- Jakobsen, B; Tasker, A; Zimmer, J. Domoic acid neurotoxicity in hippocampal slice cultures. Amino Acids 2002, 23(1–3), 37–44. [Google Scholar]
- Umegaki, H; Yamamoto, A; Suzuki, Y; Iguchi, A. Stimulation of the hippocampal glutamate receptor systems induces stress-like responses. Neuro. Endocrinol. Lett 2006, 27(3), 339–343. [Google Scholar]
- Petroni, KC; Shen, L; Guyre, PM. Modulation of human polymorphonuclear leukocyte IgG Fc receptors and Fc receptor-mediated functions by IFN-gamma and glucocorticoids. J. Immunol 1988, 140(10), 3467–3472. [Google Scholar]
- Long, F; Wang, YX; Liu, L; Zhou, J; Cui, RY; Jiang, CL. Rapid nongenomic inhibitory effects of glucocorticoids on phagocytosis and superoxide anion production by macrophages. Steroids 2005, 70(1), 55–61. [Google Scholar]
- Goulding, NJ; Euzger, HS; Butt, SK; Perretti, M. Novel pathways for glucocorticoid effects on neutrophils in chronic inflammation. Inflamm. Res 1998, 47(Suppl 3), S158–165. [Google Scholar]
- Goulding, NJ; Euzger, HS; Butt, SK; Perretti, M. Novel pathways for glucocorticoid effects on neutrophils in chronic inflammation. Inflamm. Res. 1998, 47(Suppl 3), S158–S165. [Google Scholar]
- De, A; Blotta, HM; Mamoni, RL; Louzada, P; Bertolo, MB; Foss, NT; Moreira, AC; Castro, M. Effects of dexamethasone on lymphocyte proliferation and cytokine production in rheumatoid arthritis. J. Rheumatol. 2002, 29(1), 46–51. [Google Scholar]
- Musiani, M; Gentile, L; Valentini, M; Modesti, A; Musiani, P. Lymphocyte proliferative response in brown bears: cytokine role and glucocorticoid effect. J. Exp. Zool 1998, 280(6), 421–428. [Google Scholar]
- Ryan, JC; Morey, JS; Ramsdell, JS; Van Dolah, FM. Acute phase gene expression in mice exposed to the marine neurotoxin domoic acid. Neuroscience 2005, 136(4), 1121–1132. [Google Scholar]
- Germolec, DR. Sensitivity and predictivity in immunotoxicity testing: immune endpoints and disease resistance. Toxicol. Lett 2004, 149(1–3), 109–114. [Google Scholar]
- Hermanowicz, A; Nawarska, Z; Borys, D; Maslankiewicz, A. The neutrophil function and infectious diseases in workers occupationally exposed to organochloride insecticides. Int. Arch. Occup. Environ. Health 1982, 50(4), 329–340. [Google Scholar]
- Loose, LD; Silkworth, JB; Charbonneau, T; Blumenstock, F. Environmental chemical-induced macrophage dysfunction. Environ. Health Perspect 1981, 39, 79–91. [Google Scholar]
- Cotran, RS; Kumar, V; Collins, T. Robbins pathologic basis of disease, 6th ed; 1999; Philadelphia; W.B. Saunders Company. [Google Scholar]
- Kuby, J. Immunology, 3rd ed; 1997; New York; W.H. Freeman and Company. [Google Scholar]
- Rzeski, W; Ikonomidou, C; Turski, L. Glutamate antagonists limit tumor growth. Biochem. Pharmacol 2002, 64(8), 1195–1200. [Google Scholar]
- Stepulak, A; Sifringer, M; Rzeski, W; Brocke, K; Gratopp, A; Pohl, EE; Turski, L; Ikonomidou, C. AMPA antagonists inhibit the extracellular signal regulated kinase pathway and suppress lung cancer growth. Cancer Biol. Ther. 2007, 6(12), 1908–1915. [Google Scholar]
- Carvalho, PS; Catian, R; Moukha, S; Matias, WG; Creppy, EE. Comparative study of Domoic Acid and Okadaic Acid induced-chromosomal abnormalities in the Caco-2 cell line. Int. J. Environ. Res. Public Health 2006, 3(1), 4–10. [Google Scholar]
- Mathew, A; Kurane, I; Green, S; Vaughn, DW; Kalayanarooj, S; Suntayakorn, S; Ennis, FA; Rothman, AL. Impaired T cell proliferation in acute dengue infection. J. Immunol 1999, 162(9), 5609–5615. [Google Scholar]
- Brousseau, P; Payette, Y; Tryphonas, H; Blakley, B; Boermans, H; Flipo, D; Fournier, M. Manual of Immunological Methods; 1999; Boca Raton; CRC Press. [Google Scholar]
- Brousseau, P; Pellerin, J; Morin, Y; Cyr, D; Blakley, B; Boermans, H; Fournier, M. Flow cytometry as a tool to monitor the disturbance of phagocytosis in the clam Mya arenaria hemocytes following in vitro exposure to heavy metals. Toxicology 2000, 142(2), 145–156. [Google Scholar]
- Burleson, GR; Dean, JH; Munson, AE (Eds.) Methods in Immunotoxicoogy; Vol. 2, 1995; Wiley-Liss, Inc: New York.
- Suzuki, CA; Hierlihy, SL. Renal clearance of domoic acid in the rat. Food Chem. Toxicol. 1993, 31(10), 701–706. [Google Scholar]
- Samples Availability: Available from the authors.

Share and Cite
Levin, M.; Leibrecht, H.; Ryan, J.; Van Dolah, F.; De Guise, S. Immunomodulatory Effects of Domoic Acid Differ Between In vivo and In vitro Exposure in Mice. Mar. Drugs 2008, 6, 636-659. https://doi.org/10.3390/md6040636
Levin M, Leibrecht H, Ryan J, Van Dolah F, De Guise S. Immunomodulatory Effects of Domoic Acid Differ Between In vivo and In vitro Exposure in Mice. Marine Drugs. 2008; 6(4):636-659. https://doi.org/10.3390/md6040636
Chicago/Turabian StyleLevin, Milton, Heather Leibrecht, James Ryan, Frances Van Dolah, and Sylvain De Guise. 2008. "Immunomodulatory Effects of Domoic Acid Differ Between In vivo and In vitro Exposure in Mice" Marine Drugs 6, no. 4: 636-659. https://doi.org/10.3390/md6040636