A Novel Atypical PKC-Iota Inhibitor, Echinochrome A, Enhances Cardiomyocyte Differentiation from Mouse Embryonic Stem Cells


 ,
, 
Abstract
:1. Introduction
2. Results
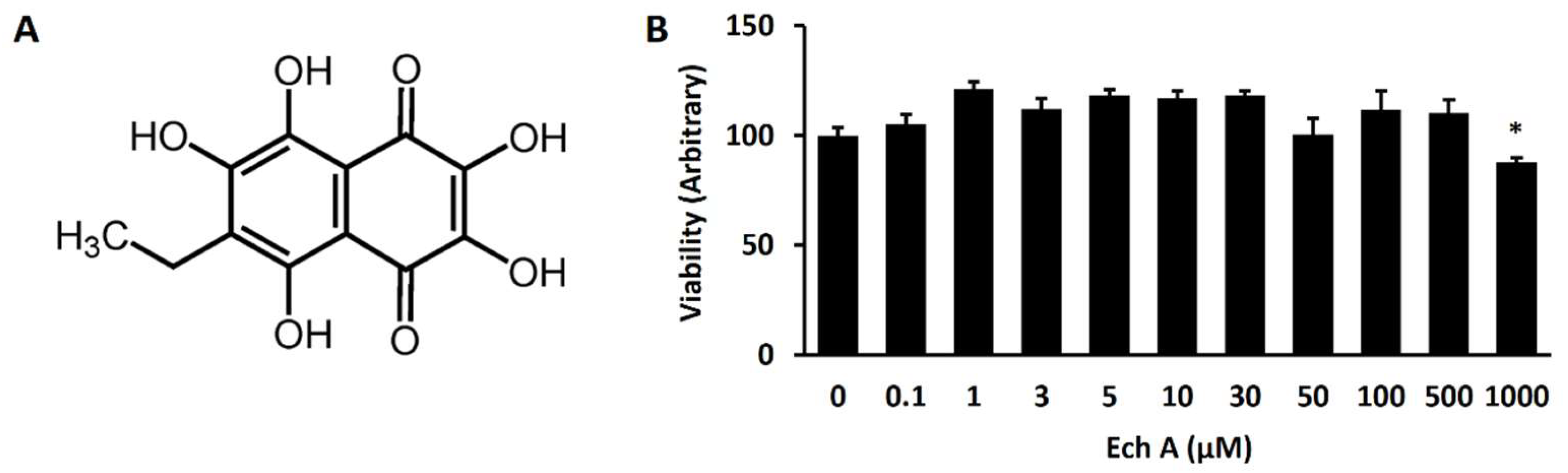
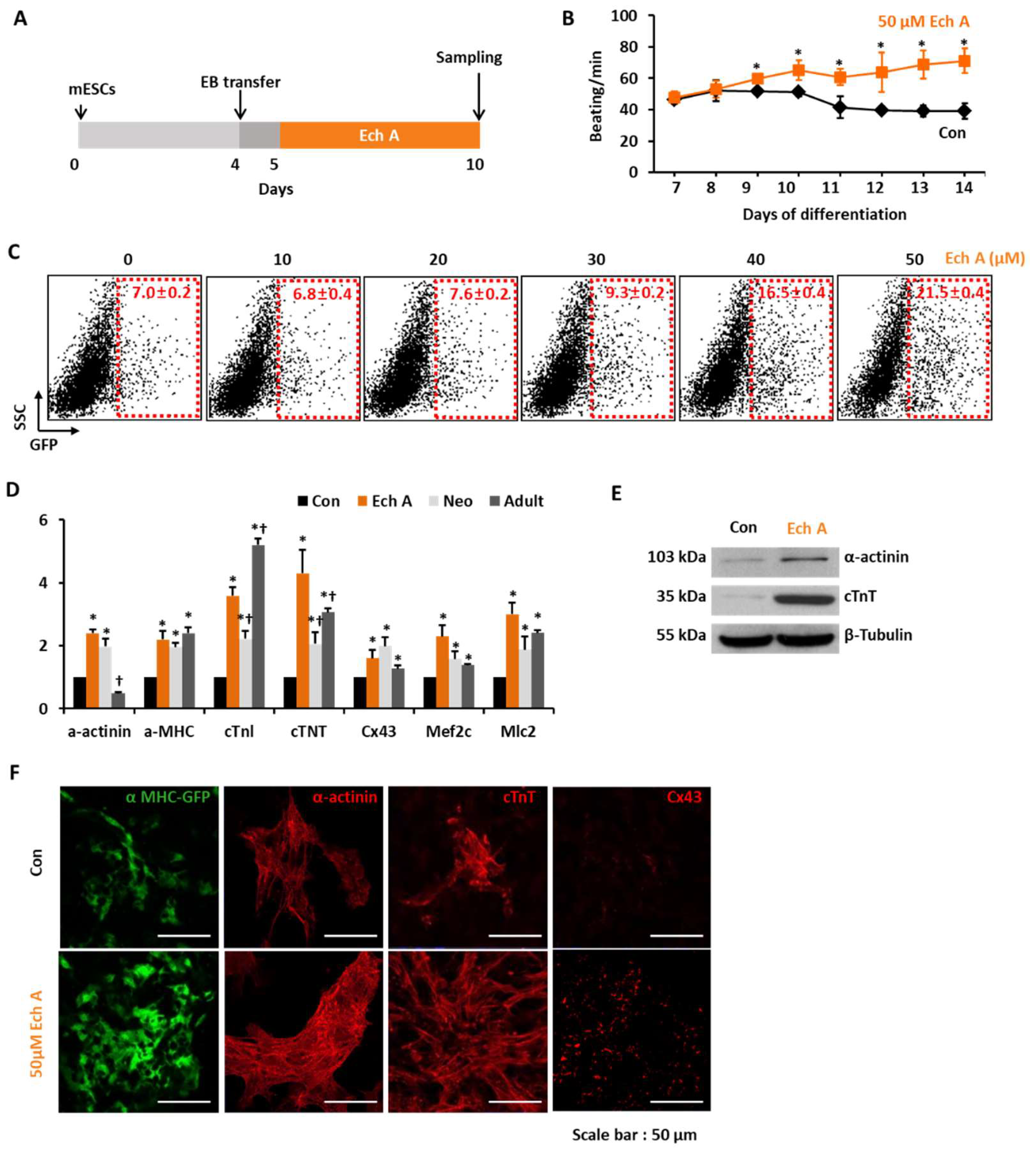
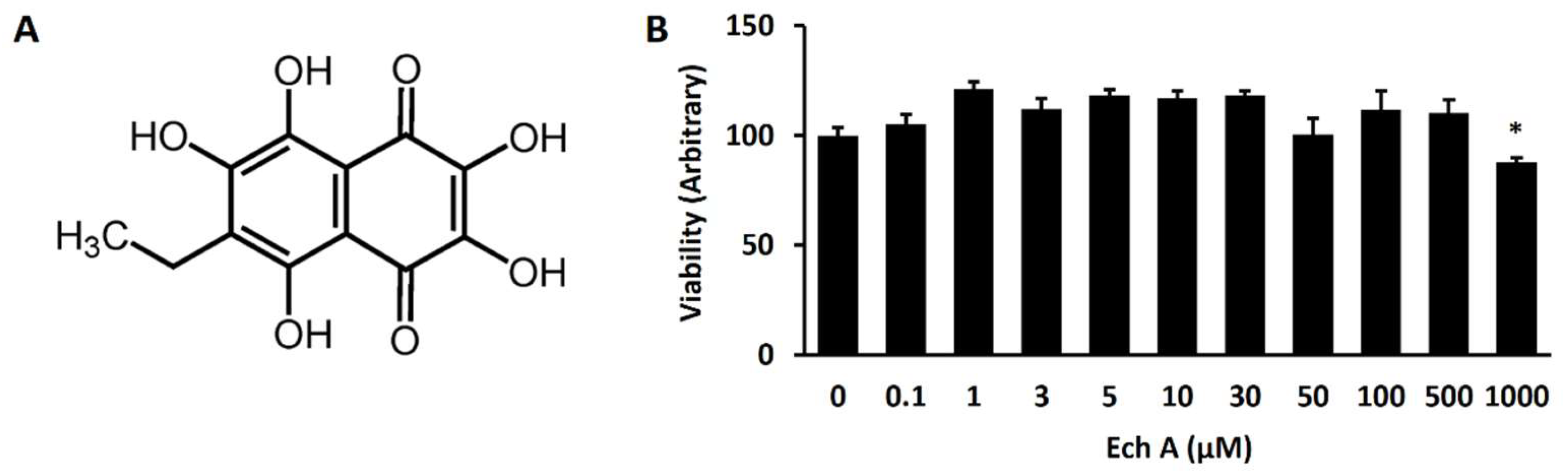
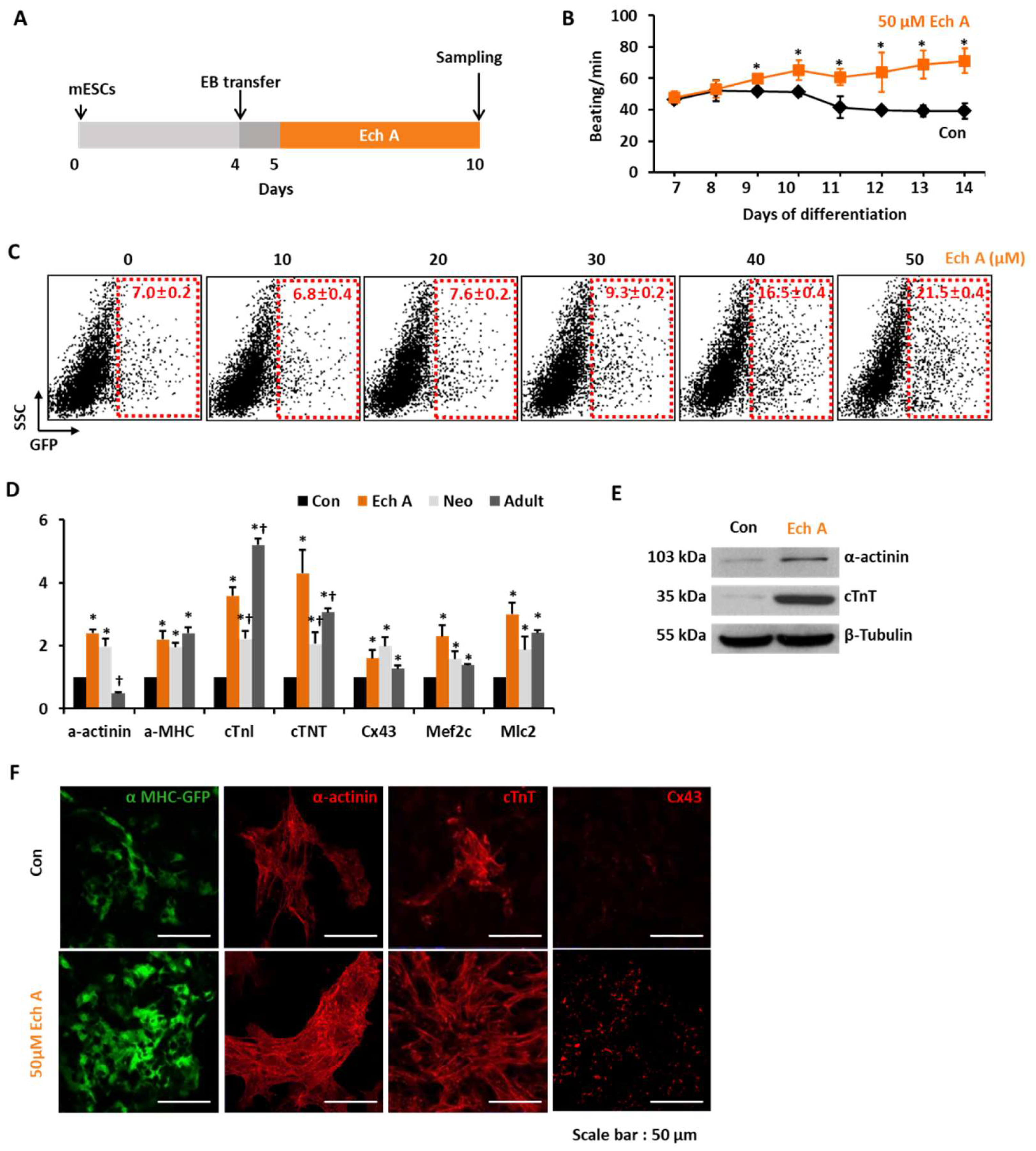
2.1. Ech A Enhances Cardiomyocyte Differentiation from mESCs
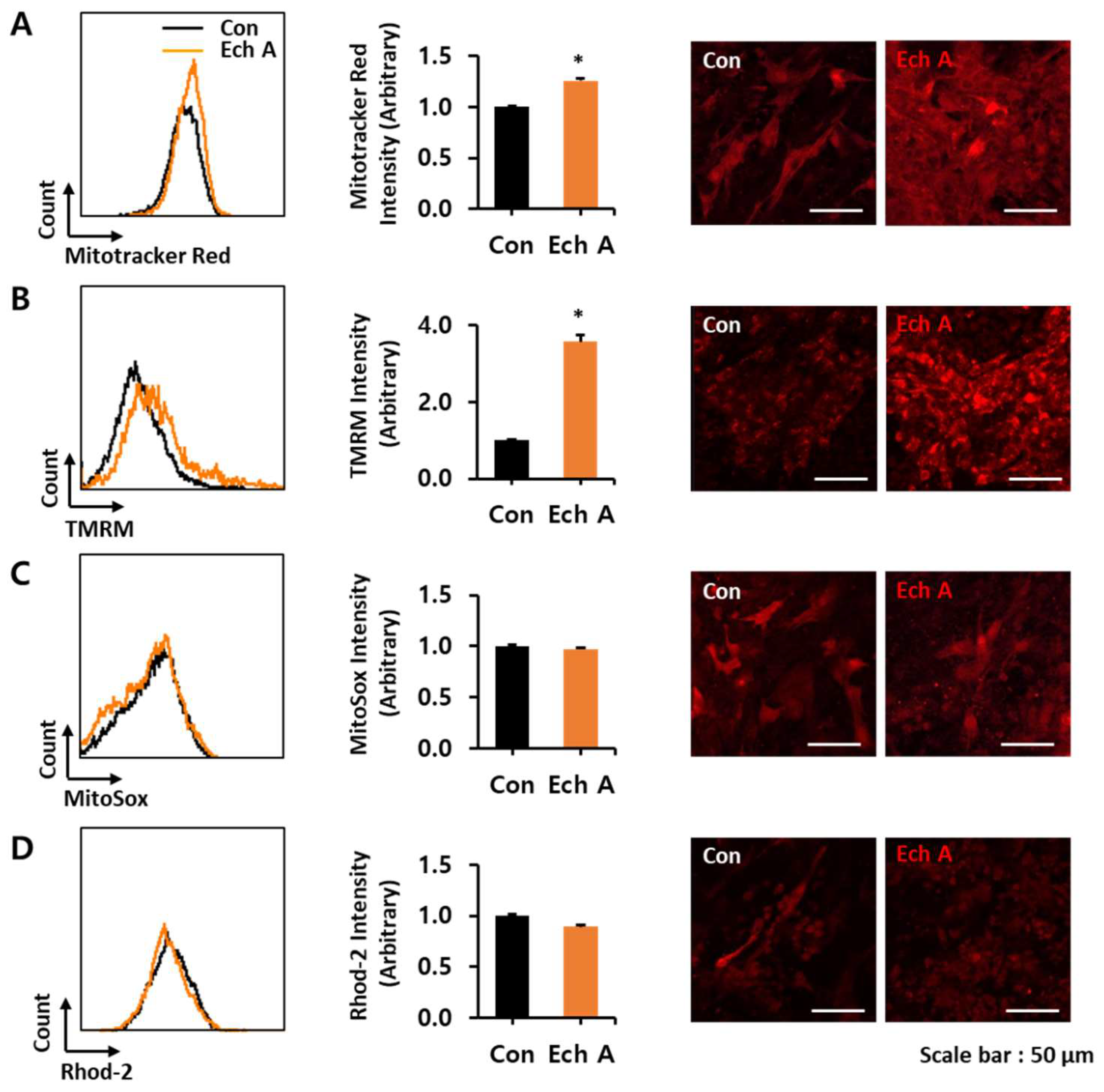
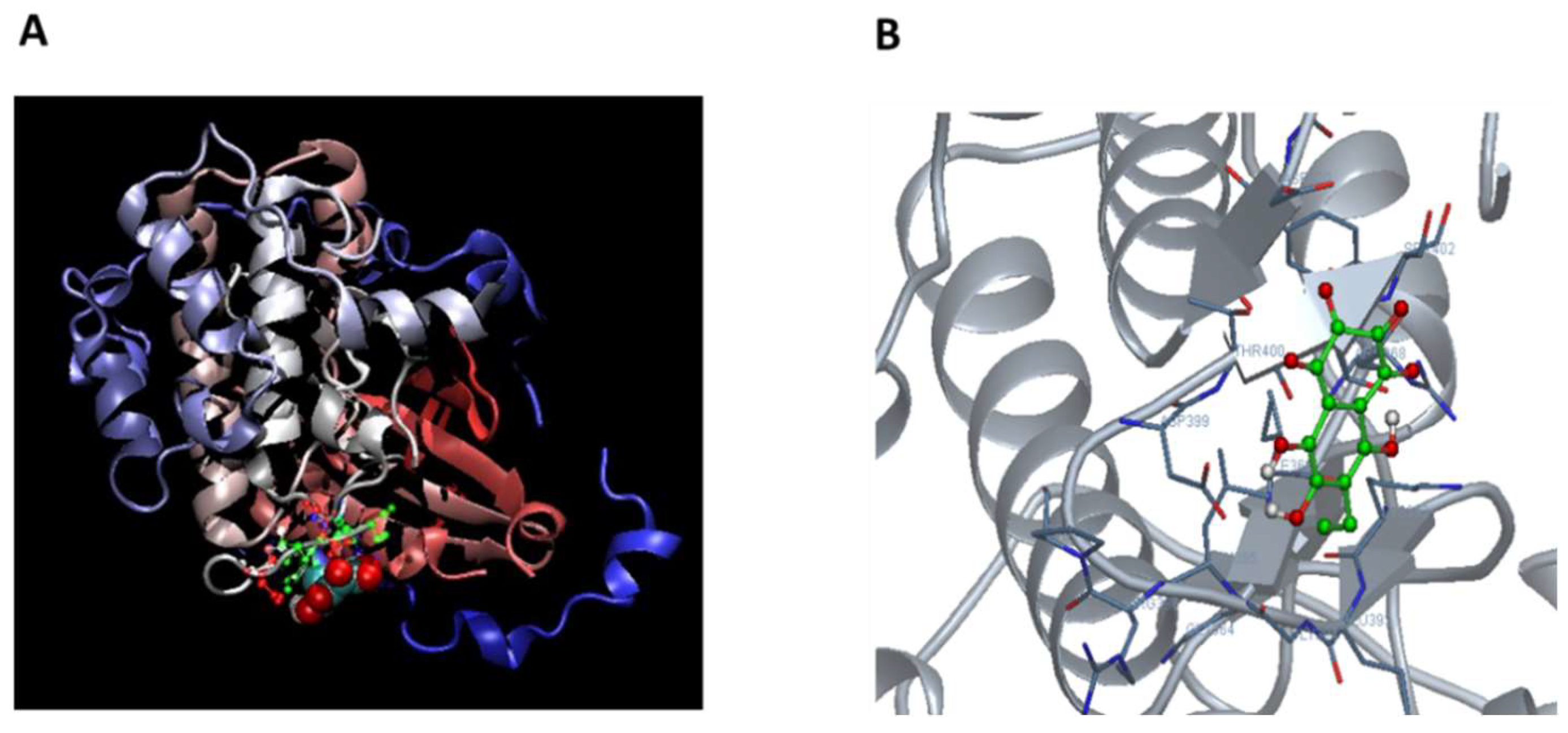
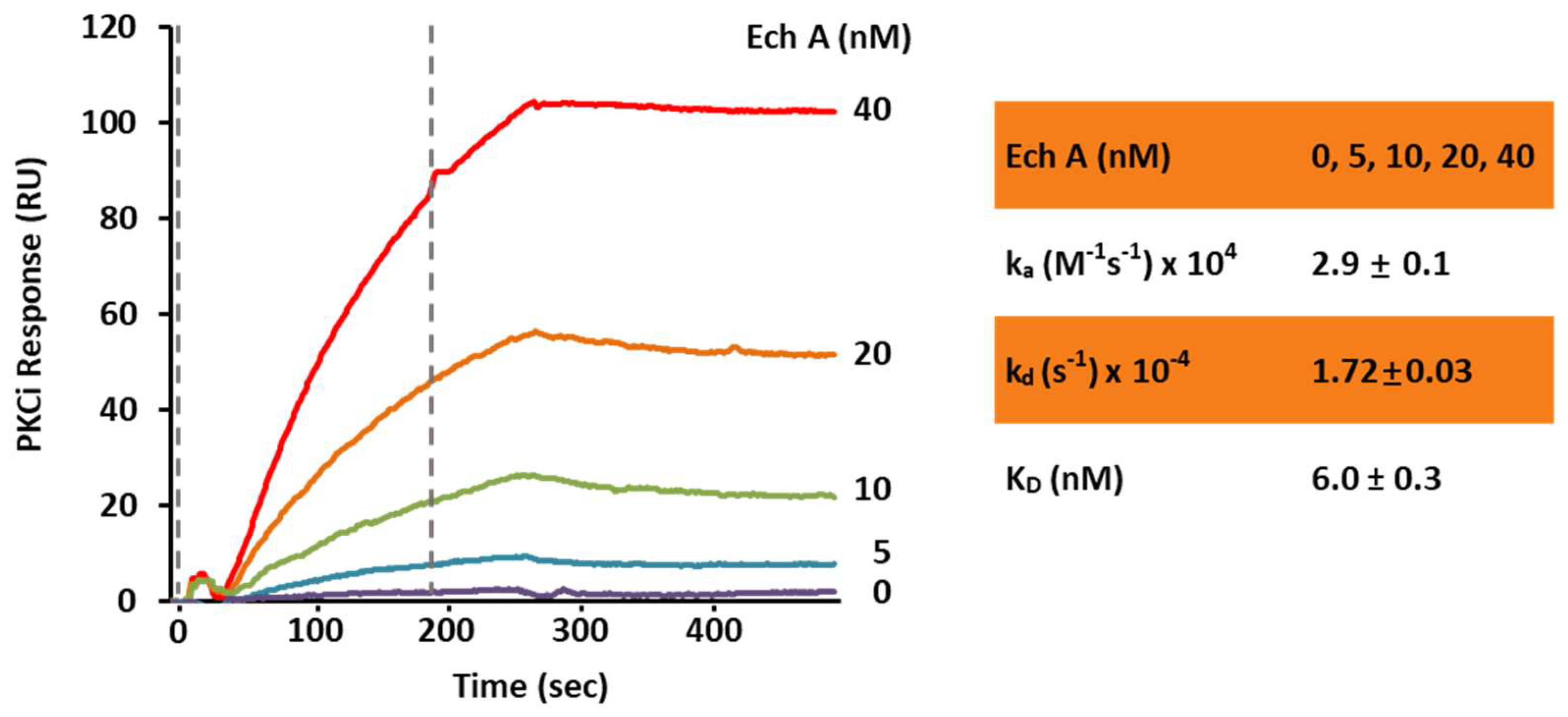
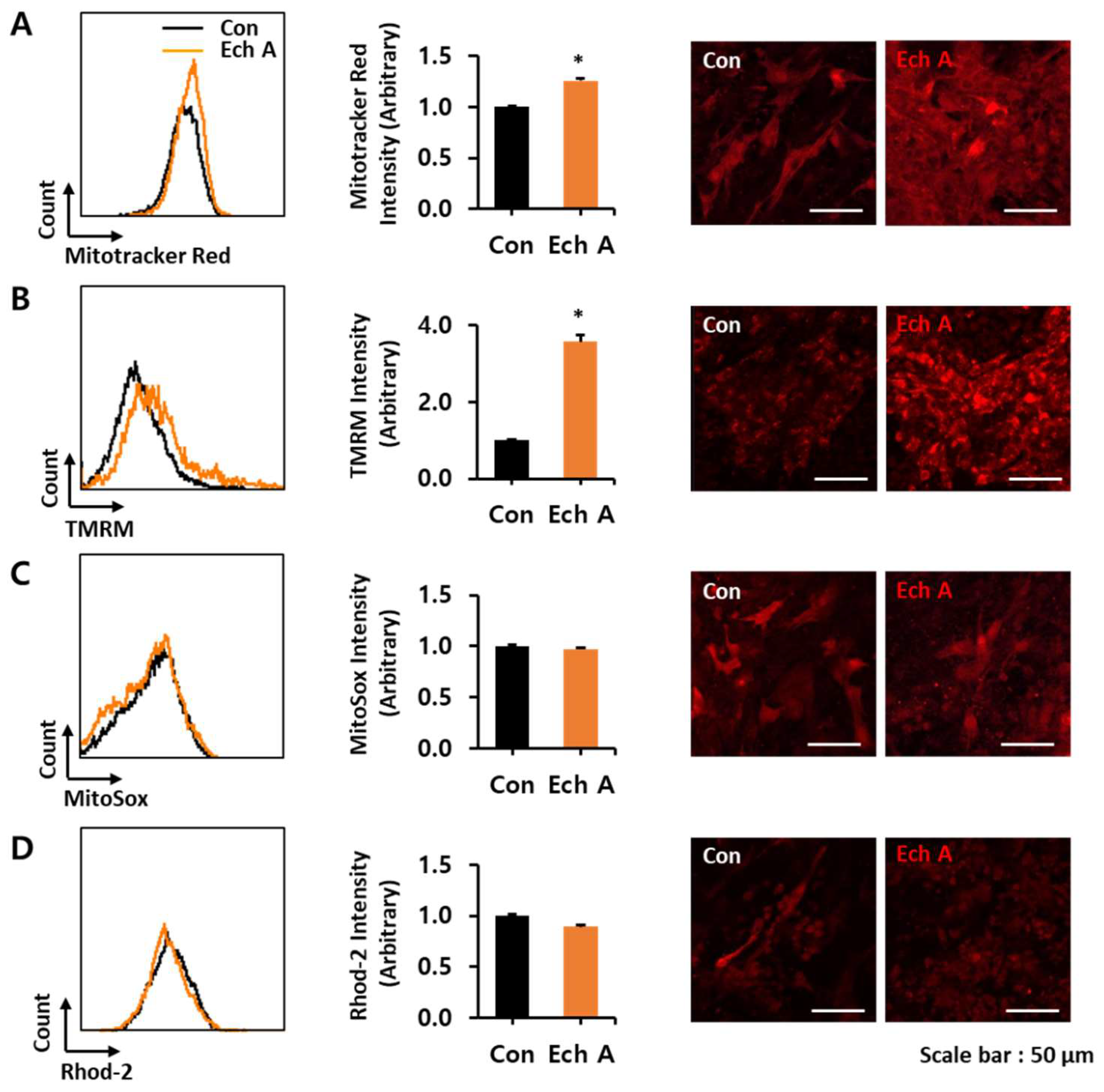
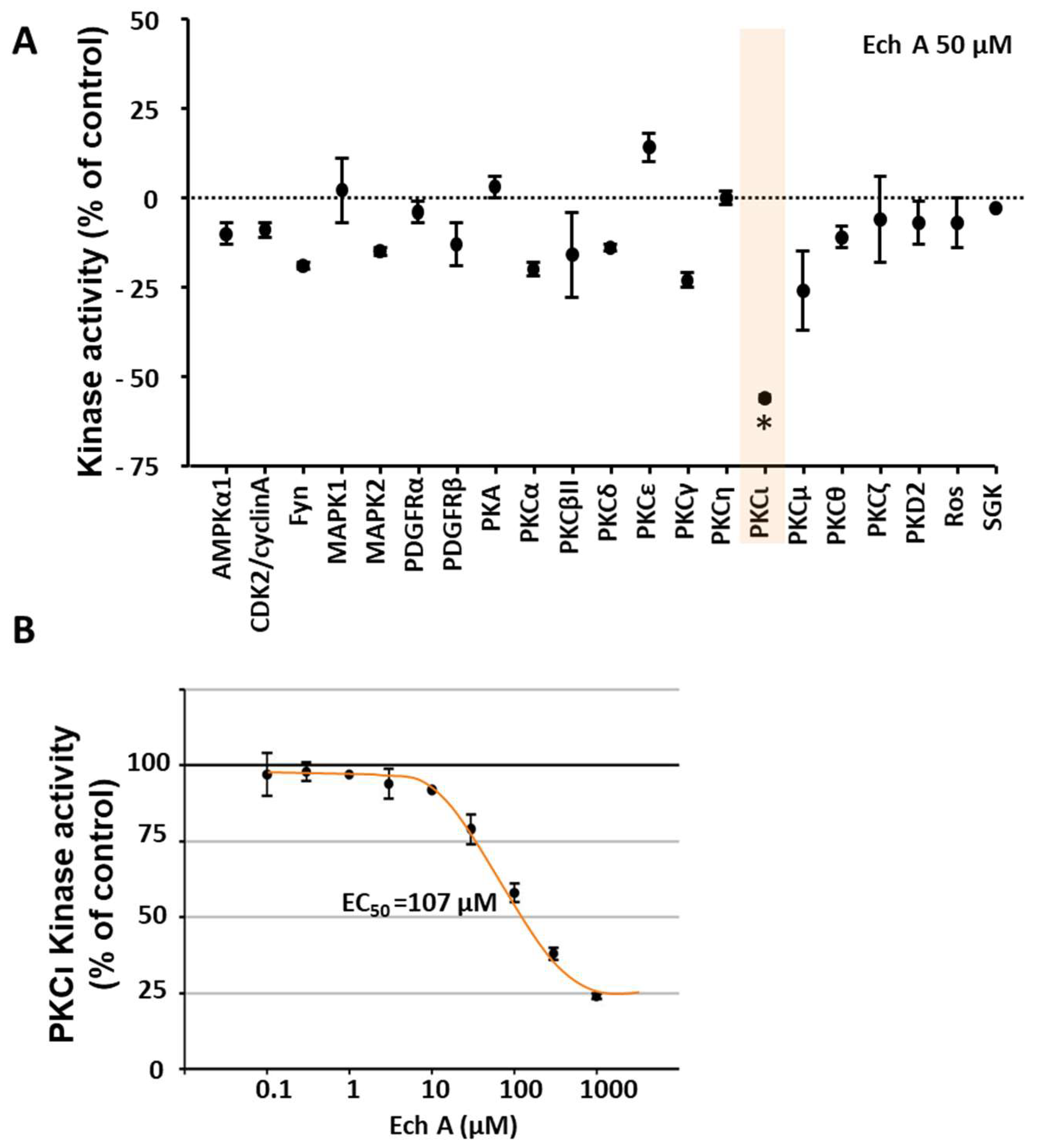
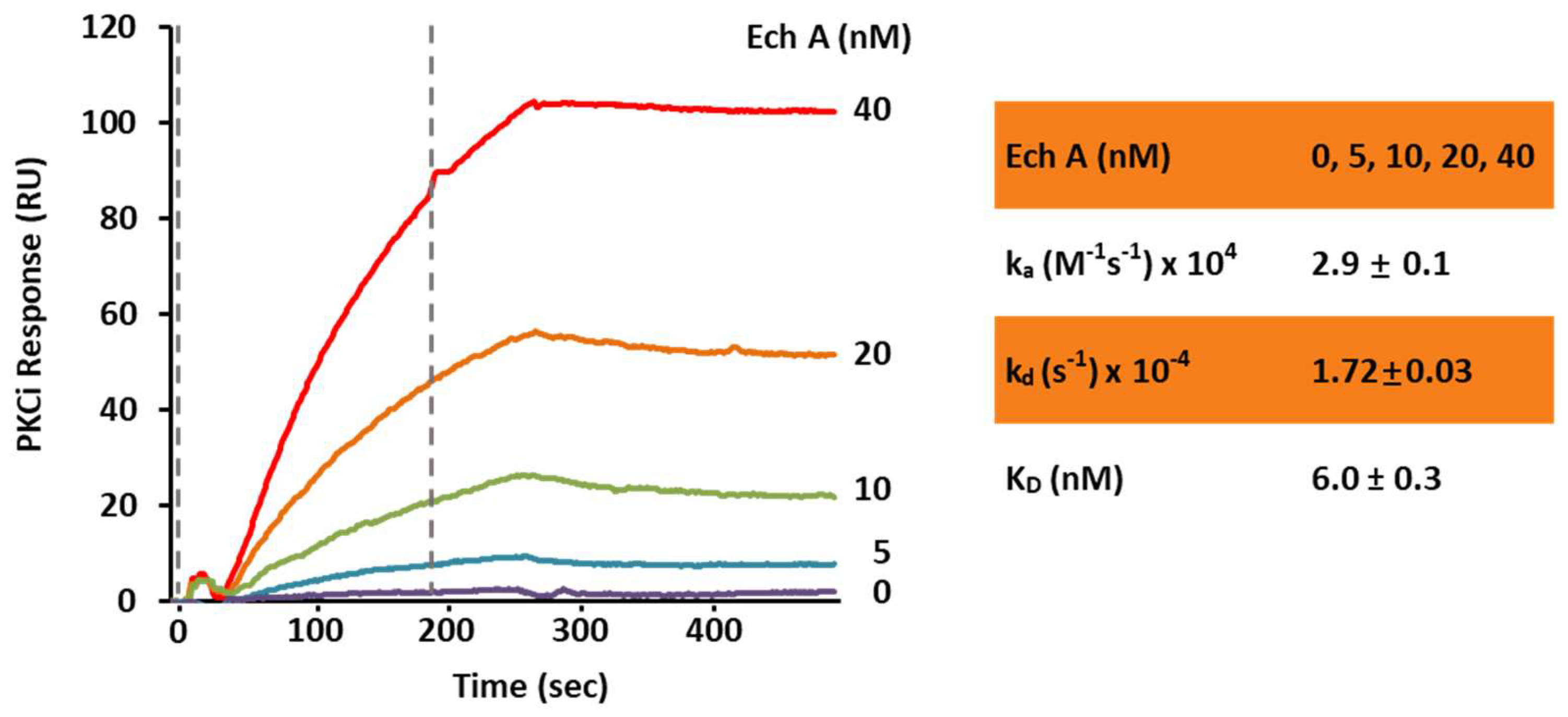
2.2. Ech A Enhances Cardiomyocyte Differentiation Through Direct Binding to Atypical PKCι and Inhibition of Its Activity
3. Discussion
4. Materials and Methods
4.1. Maintenance of mESCs
4.2. Induction of mESC-Derived Cardiomyocyte-Like Cell Using EBs
4.3. Cell Viability Assay
4.4. Quantitative Real-Time Polymerase Chain Reaction (PCR)
4.5. Western Blot Analyses
4.6. Immunocytochemistry
4.7. Flow Cytometry Analyses
4.8. Kinase Activity Assay
4.9. Computational Protein-Ligand Docking Simulation
4.10. Surface Plasmon Resonance Binding Assay
4.11. Statistical Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Hansson, E.M.; Lindsay, M.E.; Chien, K.R. Regeneration next: Toward heart stem cell therapeutics. Cell Stem Cell 2009, 5, 364–377. [Google Scholar] [CrossRef] [PubMed]
- Lin, Z.; Pu, W.T. Strategies for cardiac regeneration and repair. Sci. Transl. Med. 2014, 6, 239rv1. [Google Scholar] [CrossRef] [PubMed]
- Passier, R.; van Laake, L.W.; Mummery, C.L. Stem-cell-based therapy and lessons from the heart. Nature 2008, 453, 322–329. [Google Scholar] [CrossRef] [PubMed]
- Segers, V.F.; Lee, R.T. Stem-cell therapy for cardiac disease. Nature 2008, 451, 937–942. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burridge, P.W.; Keller, G.; Gold, J.D.; Wu, J.C. Production of de novo cardiomyocytes: Human pluripotent stem cell differentiation and direct reprogramming. Cell Stem Cell 2012, 10, 16–28. [Google Scholar] [CrossRef] [PubMed]
- Hong, S.P.; Song, S.; Cho, S.W.; Lee, S.; Koh, B.I.; Bae, H.; Kim, K.H.; Park, J.S.; Do, H.S.; Im, I.; et al. Generation of PDGFRalpha+ Cardioblasts from Pluripotent Stem Cells. Sci. Rep. 2017, 7, 41840. [Google Scholar] [CrossRef] [PubMed]
- Lebedev, A.V.; Ivanova, M.V.; Levitsky, D.O. Echinochrome, a naturally occurring iron chelator and free radical scavenger in artificial and natural membrane systems. Life Sci. 2005, 76, 863–875. [Google Scholar] [CrossRef] [PubMed]
- Thomson, R.H. Distribution of naturally occurring quinones. Pharm. Weekbl. Sci. 1991, 13, 70–73. [Google Scholar] [CrossRef] [PubMed]
- Shvilkin, A.V.; Serebriakov, L.I.; Tskitishvili, O.V.; Sadretdinov, S.M.; Kol’tsova, E.A.; Maksimov, O.B.; Mishchenko, N.P.; Novikov, V.L.; Levitskii, D.O.; Ruda, M. Effect of echinochrom on experimental myocardial reperfusion injury. Kardiologiia 1991, 31, 79–81. [Google Scholar] [PubMed]
- Buimov, G.A.; Maksimov, I.V.; Perchatkin, V.A.; Repin, A.N.; Afanas’ev, S.A.; Markov, V.A.; Karpov, R.S. Effect of the bioantioxidant histochrome on myocardial injury in reperfusion therapy on patients with myocardial infarction. Terapevticheskii Arkhiv 2002, 74, 12–16. [Google Scholar] [PubMed]
- Jeong, S.H.; Kim, H.K.; Song, I.S.; Noh, S.J.; Marquez, J.; Ko, K.S.; Rhee, B.D.; Kim, N.; Mishchenko, N.P.; Fedoreyev, S.A.; et al. Echinochrome a increases mitochondrial mass and function by modulating mitochondrial biogenesis regulatory genes. Mar. Drugs 2014, 12, 4602–4615. [Google Scholar] [CrossRef] [PubMed]
- Jeong, S.H.; Kim, H.K.; Song, I.S.; Lee, S.J.; Ko, K.S.; Rhee, B.D.; Kim, N.; Mishchenko, N.P.; Fedoryev, S.A.; Stonik, V.A.; et al. Echinochrome A protects mitochondrial function in cardiomyocytes against cardiotoxic drugs. Mar. Drugs 2014, 12, 2922–2936. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.K.; Youm, J.B.; Jeong, S.H.; Lee, S.R.; Song, I.S.; Ko, T.H.; Pronto, J.R.; Ko, K.S.; Rhee, B.D.; Kim, N.; et al. Echinochrome A regulates phosphorylation of phospholamban Ser16 and Thr17 suppressing cardiac SERCA2A Ca(2)(+) reuptake. Pflugers Arch. 2015, 467, 2151–2163. [Google Scholar] [CrossRef] [PubMed]
- Cho, S.W.; Park, J.S.; Heo, H.J.; Park, S.W.; Song, S.; Kim, I.; Han, Y.M.; Yamashita, J.K.; Youm, J.B.; Han, J.; et al. Dual modulation of the mitochondrial permeability transition pore and redox signaling synergistically promotes cardiomyocyte differentiation from pluripotent stem cells. J. Am. Heart Assoc. 2014, 3, e000693. [Google Scholar] [CrossRef] [PubMed]
- Spitkovsky, D.; Sasse, P.; Kolossov, E.; Bottinger, C.; Fleischmann, B.K.; Hescheler, J.; Wiesner, R.J. Activity of complex III of the mitochondrial electron transport chain is essential for early heart muscle cell differentiation. FASEB J. 2004, 18, 1300–1302. [Google Scholar] [CrossRef] [PubMed]
- Chung, S.; Dzeja, P.P.; Faustino, R.S.; Perez-Terzic, C.; Behfar, A.; Terzic, A. Mitochondrial oxidative metabolism is required for the cardiac differentiation of stem cells. Nat. Clin. Pract. Cardiovasc. Med. 2007, 4 (Suppl. 1), S60–S67. [Google Scholar] [CrossRef] [PubMed]
- Porter, G.A., Jr.; Hom, J.; Hoffman, D.; Quintanilla, R.; de Mesy Bentley, K.; Sheu, S.S. Bioenergetics, mitochondria, and cardiac myocyte differentiation. Prog. Pediatr. Cardiol. 2011, 31, 75–81. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tohyama, S.; Hattori, F.; Sano, M.; Hishiki, T.; Nagahata, Y.; Matsuura, T.; Hashimoto, H.; Suzuki, T.; Yamashita, H.; Satoh, Y.; et al. Distinct metabolic flow enables large-scale purification of mouse and human pluripotent stem cell-derived cardiomyocytes. Cell Stem Cell 2013, 12, 127–137. [Google Scholar] [CrossRef] [PubMed]
- Heo, H.J.; Kim, H.K.; Youm, J.B.; Cho, S.W.; Song, I.S.; Lee, S.Y.; Ko, T.H.; Kim, N.; Ko, K.S.; Rhee, B.D.; et al. Mitochondrial pyruvate dehydrogenase phosphatase 1 regulates the early differentiation of cardiomyocytes from mouse embryonic stem cells. Exp. Mol. Med. 2016, 48, e254. [Google Scholar] [CrossRef] [PubMed]
- Mahato, B.; Home, P.; Rajendran, G.; Paul, A.; Saha, B.; Ganguly, A.; Ray, S.; Roy, N.; Swerdlow, R.H.; Paul, S. Regulation of mitochondrial function and cellular energy metabolism by protein kinase C-lambda/iota: A novel mode of balancing pluripotency. Stem Cells 2014, 32, 2880–2892. [Google Scholar] [CrossRef] [PubMed]
- Mah, I.K.; Soloff, R.; Hedrick, S.M.; Mariani, F.V. Atypical PKC-iota Controls Stem Cell Expansion via Regulation of the Notch Pathway. Stem Cell Rep. 2015, 5, 866–880. [Google Scholar] [CrossRef] [PubMed]
- Seidl, S.; Braun, U.; Roos, N.; Li, S.; Ludtke, T.H.; Kispert, A.; Leitges, M. Phenotypical analysis of atypical PKCs in vivo function display a compensatory system at mouse embryonic day 7.5. PLoS ONE 2013, 8, e62756. [Google Scholar] [CrossRef] [PubMed]
- Pillai, P.; Desai, S.; Patel, R.; Sajan, M.; Farese, R.; Ostrov, D.; Acevedo-Duncan, M. A novel PKC-iota inhibitor abrogates cell proliferation and induces apoptosis in neuroblastoma. Int. J. Biochem. Cell Biol. 2011, 43, 784–794. [Google Scholar] [CrossRef] [PubMed]
- Jatoi, A.; Grudem, M.E.; Dockter, T.J.; Block, M.S.; Villasboas, J.C.; Tan, A.; Deering, E.; Kasi, P.M.; Mansfield, A.S.; Botero, J.P.; et al. A proof-of-concept trial of protein kinase C iota inhibition with auranofin for the paclitaxel-induced acute pain syndrome. Support. Care Cancer 2017, 25, 833–838. [Google Scholar] [CrossRef] [PubMed]
- Chand, S.; Mehta, N.; Bahia, M.S.; Dixit, A.; Silakari, O. Protein kinase C-theta inhibitors: A novel therapy for inflammatory disorders. Curr. Pharm. Des. 2012, 18, 4725–4746. [Google Scholar] [CrossRef] [PubMed]
- Win, H.Y.; Acevedo-Duncan, M. Role of protein kinase C-iota in transformed non-malignant RWPE-1 cells and androgen-independent prostate carcinoma DU-145 cells. Cell Prolif. 2009, 42, 182–194. [Google Scholar] [CrossRef] [PubMed]
- Desai, S.R.; Pillai, P.P.; Patel, R.S.; McCray, A.N.; Win-Piazza, H.Y.; Acevedo-Duncan, M.E. Regulation of Cdk7 activity through a phosphatidylinositol (3)-kinase/PKC-iota-mediated signaling cascade in glioblastoma. Carcinogenesis 2012, 33, 10–19. [Google Scholar] [CrossRef] [PubMed]
- McCray, A.N.; Desai, S.; Acevedo-Duncan, M. The interruption of PKC-iota signaling and TRAIL combination therapy against glioblastoma cells. Neurochem. Res. 2014, 39, 1691–1701. [Google Scholar] [CrossRef] [PubMed]
- Butler, A.M.; Scotti Buzhardt, M.L.; Erdogan, E.; Li, S.; Inman, K.S.; Fields, A.P.; Murray, N.R. A small molecule inhibitor of atypical protein kinase C signaling inhibits pancreatic cancer cell transformed growth and invasion. Oncotarget 2015, 6, 15297–15310. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ratnayake, W.S.; Apostolatos, A.H.; Ostrov, D.A.; Acevedo-Duncan, M. Two novel atypical PKC inhibitors; ACPD and DNDA effectively mitigate cell proliferation and epithelial to mesenchymal transition of metastatic melanoma while inducing apoptosis. Int. J. Oncol. 2017, 51, 1370–1382. [Google Scholar] [CrossRef] [PubMed]
- Shiba, Y.; Gomibuchi, T.; Seto, T.; Wada, Y.; Ichimura, H.; Tanaka, Y.; Ogasawara, T.; Okada, K.; Shiba, N.; Sakamoto, K.; et al. Allogeneic transplantation of iPS cell-derived cardiomyocytes regenerates primate hearts. Nature 2016, 538, 388–391. [Google Scholar] [CrossRef] [PubMed]
- Maculins, T.; Carter, N.; Dorval, T.; Hudson, K.; Nissink, J.W.; Hay, R.T.; Alwan, H. A Generic Platform for Cellular Screening Against Ubiquitin Ligases. Sci. Rep. 2016, 6, 18940. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bikadi, Z.; Hazai, E. Application of the PM6 semi-empirical method to modeling proteins enhances docking accuracy of AutoDock. J. Cheminform. 2009, 1, 15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morris, G.M.; Goodsell, D.S.; Halliday, R.S.; Huey, R.; Hart, W.E.; Belew, R.K.; Olson, A.J. Automated docking using a Lamarckian genetic algorithm and an empirical binding free energy function. J. Comput. Chem. 1998, 19, 1639–1662. [Google Scholar] [CrossRef] [Green Version]
- Solis, F.J.; Wets, R.J.-B. Minimization by Random Search Techniques. Math. Oper. Res. 1981, 6, 19–30. [Google Scholar] [CrossRef]
- Ali, A.G.; Mohamed, M.F.; Abdelhamid, A.O.; Mohamed, M.S. A novel adamantane thiadiazole derivative induces mitochondria-mediated apoptosis in lung carcinoma cell line. Bioorg. Med. Chem. 2017, 25, 241–253. [Google Scholar] [CrossRef] [PubMed]
- Serasinghe, M.N.; Seneviratne, A.M.; Smrcka, A.V.; Yoon, Y. Identification and characterization of unique proline-rich peptides binding to the mitochondrial fission protein hFis1. J. Biol. Chem. 2010, 285, 620–630. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Residue No | Amino Acid | Interaction Type |
|---|---|---|
| 392 | LYS | Polar |
| 395 | LEU | Hydrophobic |
| 399 | ASP | Polar |
| 400 | THR | Hydrogen bonds, polar |
| 401 | THR | Hydrogen bonds, polar |
| 402 | SER | Other |
| 420 | ASP | Polar |
| Gene | Primer Sequences |
|---|---|
| α-actinin | Forward 5′-AGCCAGGAACAGATGAACGA-3′ Reverse 5′-AAGTCGATGAAGGCCTGGAA-3′ |
| α-MHC | Forward 5′-GCCCAGTACCTCCGAAAGTC-3′ Reverse 5′-GCCTTAACATACTCCTCCTTGTC-3′ |
| cTnI | Forward 5′-CGTGGAAGCAAAAGTCACCA-3′ Reverse 5′-GTCCTCCTTCTTCACCTGCT-3′ |
| cTnT | Forward 5′-CAGAGGAGGCCAACGTAGAAG-3′ Reverse 5′-CTCCATCGGGGATCTTGGGT-3′ |
| Cx43 | Forward 5′-ACGGCAAGGTGAAGATGAGA-3′ Reverse 5′-GAGAGACACCAAGGACACCA-3′ |
| Mef2c | Forward 5′-ACCAGGACAAGGAATGGGAG-3′ Reverse 5′-GGCGGCATGTTATGTAGGTG-3′ |
| MLC2 | Forward 5′-GGCACCAAAGAAAGCCAAGA-3′ Reverse 5′-GGACCTGGAGCCTCTTTGAT-3′ |
| GAPDH | Forward 5′-CACCATCTTCCAGGAGCGAG-3′ Reverse 5′-CCTTCTCCATGGTGGTGAAGAC-3′ |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, H.K.; Cho, S.W.; Heo, H.J.; Jeong, S.H.; Kim, M.; Ko, K.S.; Rhee, B.D.; Mishchenko, N.P.; Vasileva, E.A.; Fedoreyev, S.A.; et al. A Novel Atypical PKC-Iota Inhibitor, Echinochrome A, Enhances Cardiomyocyte Differentiation from Mouse Embryonic Stem Cells. Mar. Drugs 2018, 16, 192. https://doi.org/10.3390/md16060192
Kim HK, Cho SW, Heo HJ, Jeong SH, Kim M, Ko KS, Rhee BD, Mishchenko NP, Vasileva EA, Fedoreyev SA, et al. A Novel Atypical PKC-Iota Inhibitor, Echinochrome A, Enhances Cardiomyocyte Differentiation from Mouse Embryonic Stem Cells. Marine Drugs. 2018; 16(6):192. https://doi.org/10.3390/md16060192
Chicago/Turabian StyleKim, Hyoung Kyu, Sung Woo Cho, Hye Jin Heo, Seung Hun Jeong, Min Kim, Kyung Soo Ko, Byoung Doo Rhee, Natalia P. Mishchenko, Elena A. Vasileva, Sergey A. Fedoreyev, and et al. 2018. "A Novel Atypical PKC-Iota Inhibitor, Echinochrome A, Enhances Cardiomyocyte Differentiation from Mouse Embryonic Stem Cells" Marine Drugs 16, no. 6: 192. https://doi.org/10.3390/md16060192
APA StyleKim, H. K., Cho, S. W., Heo, H. J., Jeong, S. H., Kim, M., Ko, K. S., Rhee, B. D., Mishchenko, N. P., Vasileva, E. A., Fedoreyev, S. A., Stonik, V. A., & Han, J. (2018). A Novel Atypical PKC-Iota Inhibitor, Echinochrome A, Enhances Cardiomyocyte Differentiation from Mouse Embryonic Stem Cells. Marine Drugs, 16(6), 192. https://doi.org/10.3390/md16060192