Bioactive Compounds Isolated from Neglected Predatory Marine Gastropods



Abstract
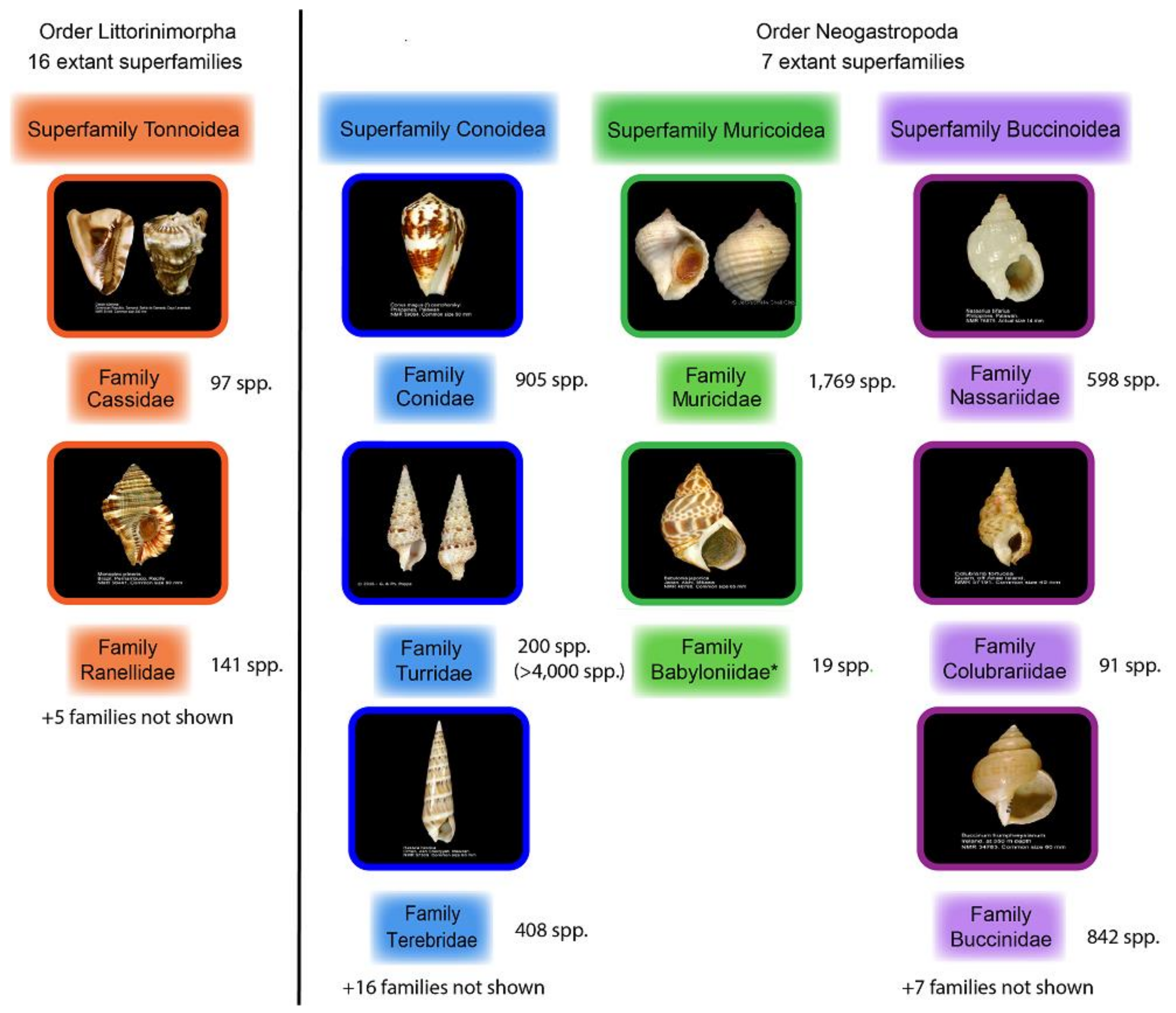
1. Introduction
2. Bioactive Compounds Isolated from Diverse Predatory Gastropods
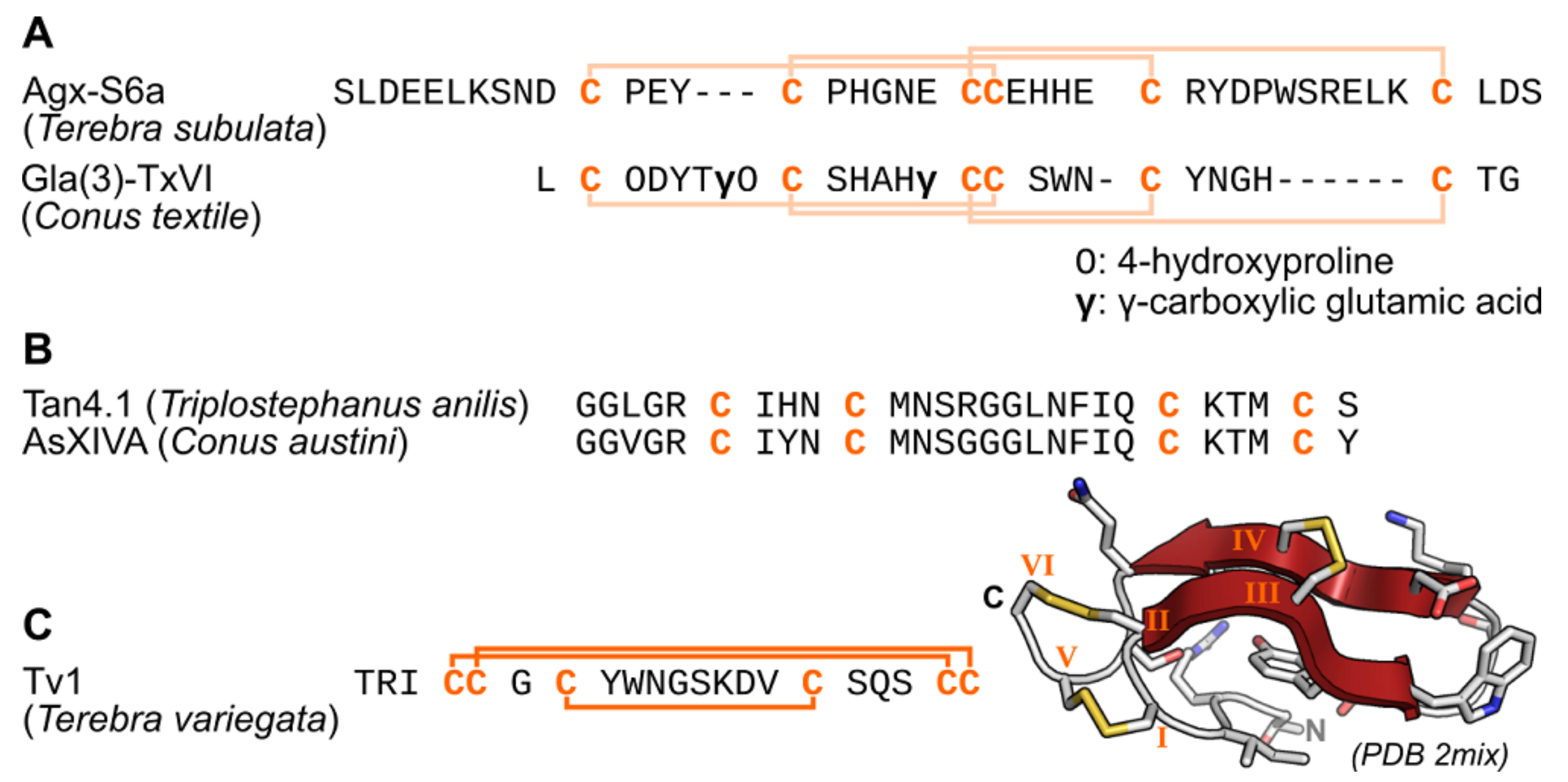
2.1. Terebrids—Auger Snails
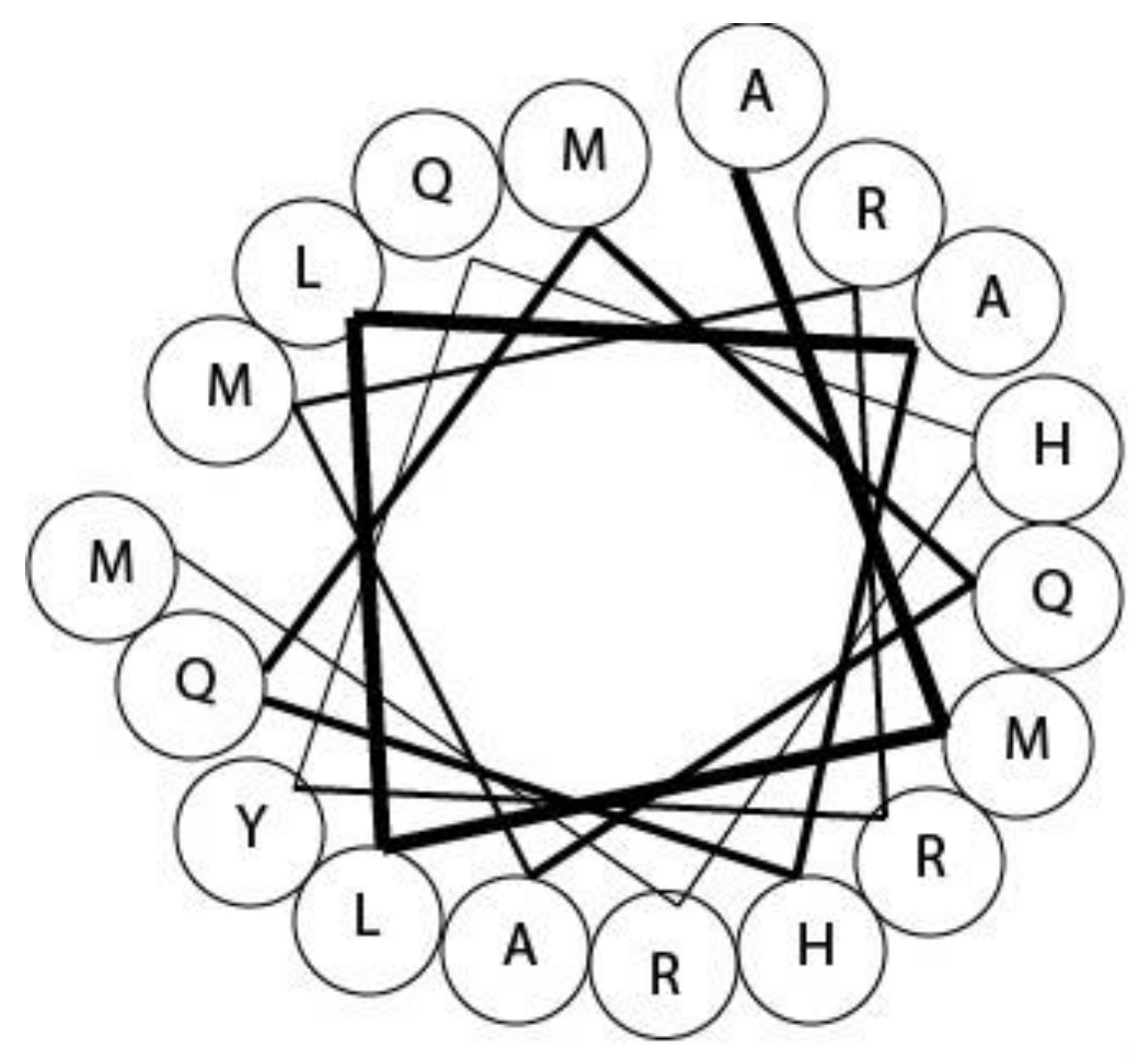
2.2. Turridae
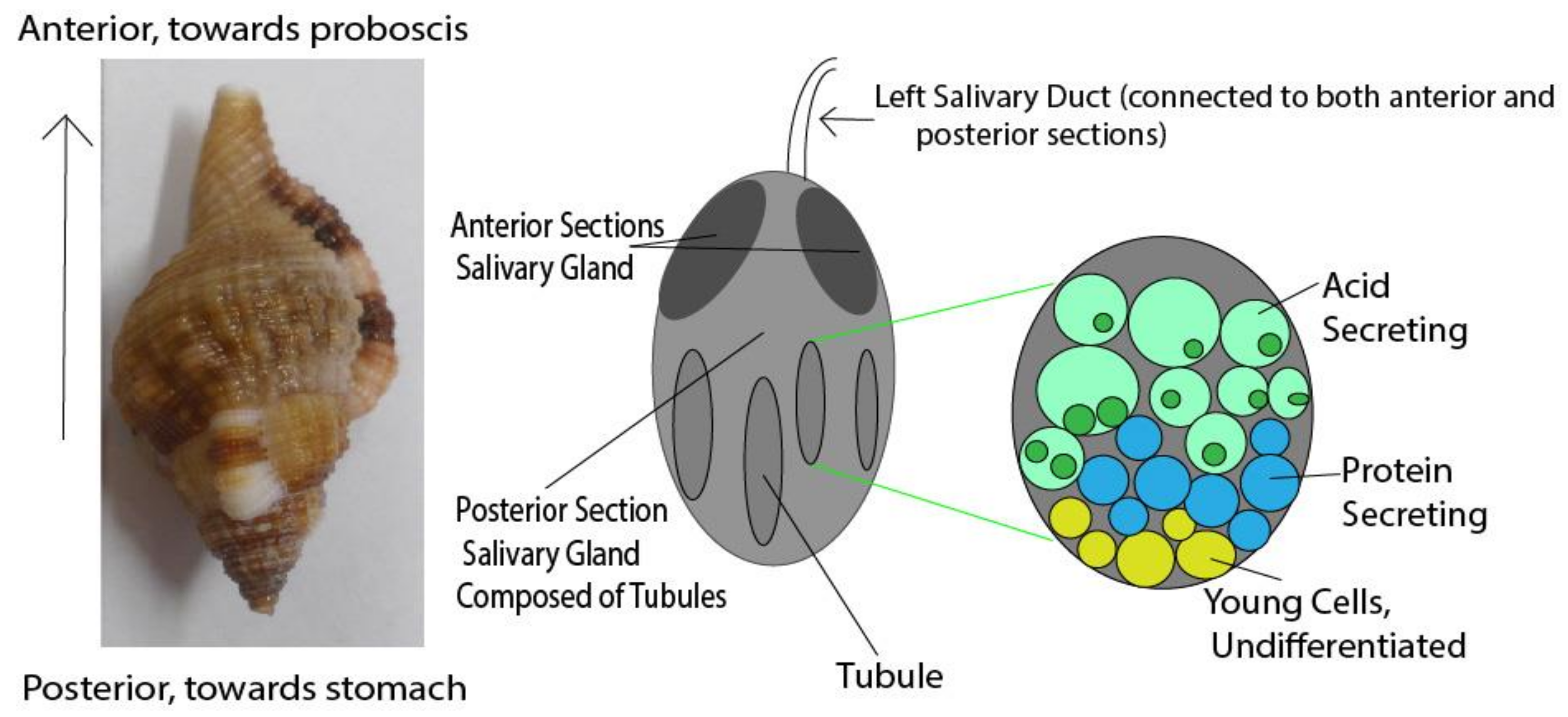
2.3. Buccinidae—True Whelks
2.4. Cassidae—Helmet Shells
2.5. Colubrariidae—Vampire Snail
2.6. Muricidae—Rock Snails
2.7. Family Ranellidae
2.7.1. Genus Charonia
2.7.2. Genus Monoplex
2.8. Bioaccumulated Toxins—Selected Examples
3. Pharmaceutical and Biotechnological Applications
3.1. Venom-Derived Peptides
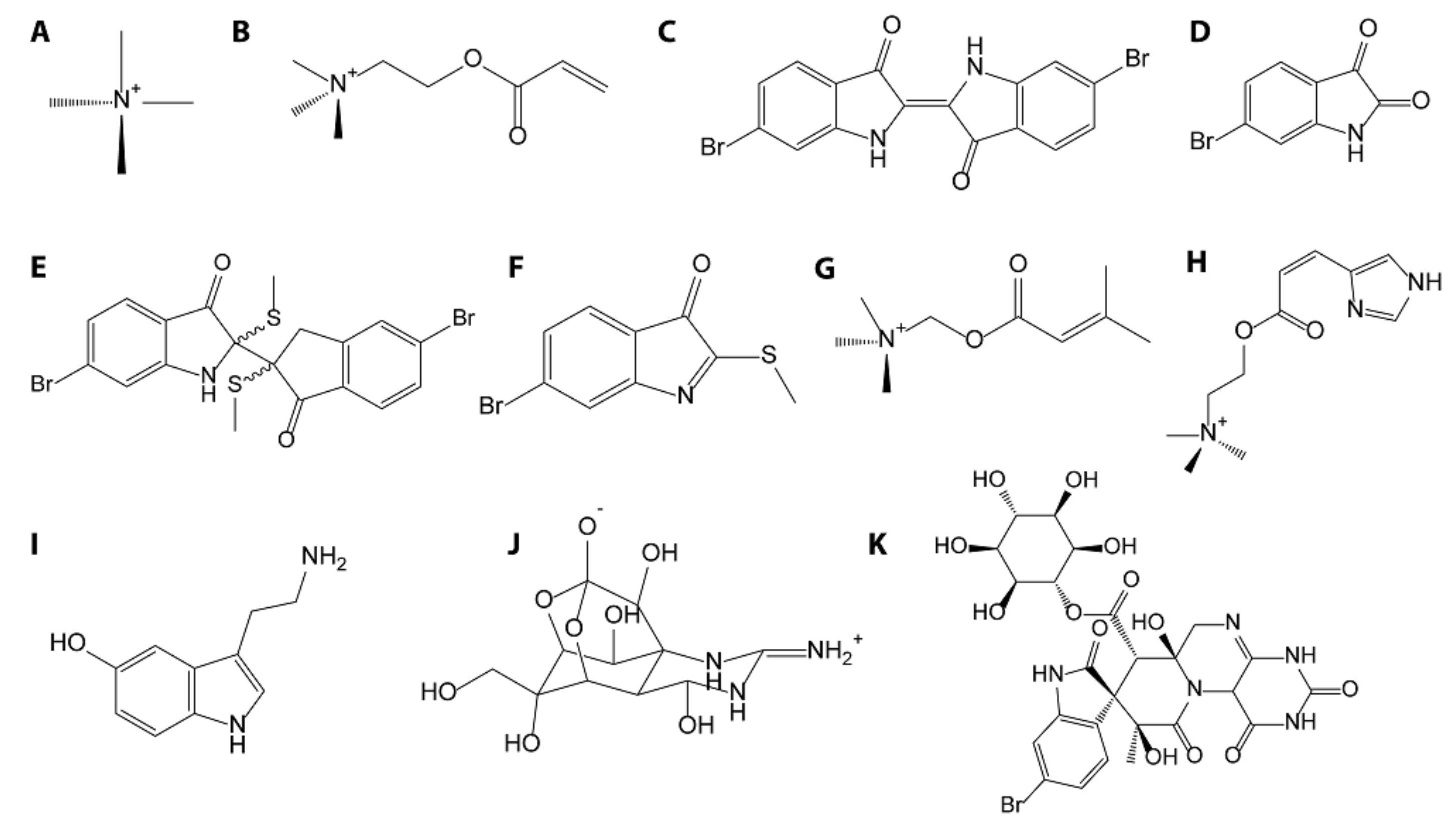
3.2. Brominated Indoles—Muricidae
3.3. Murexine
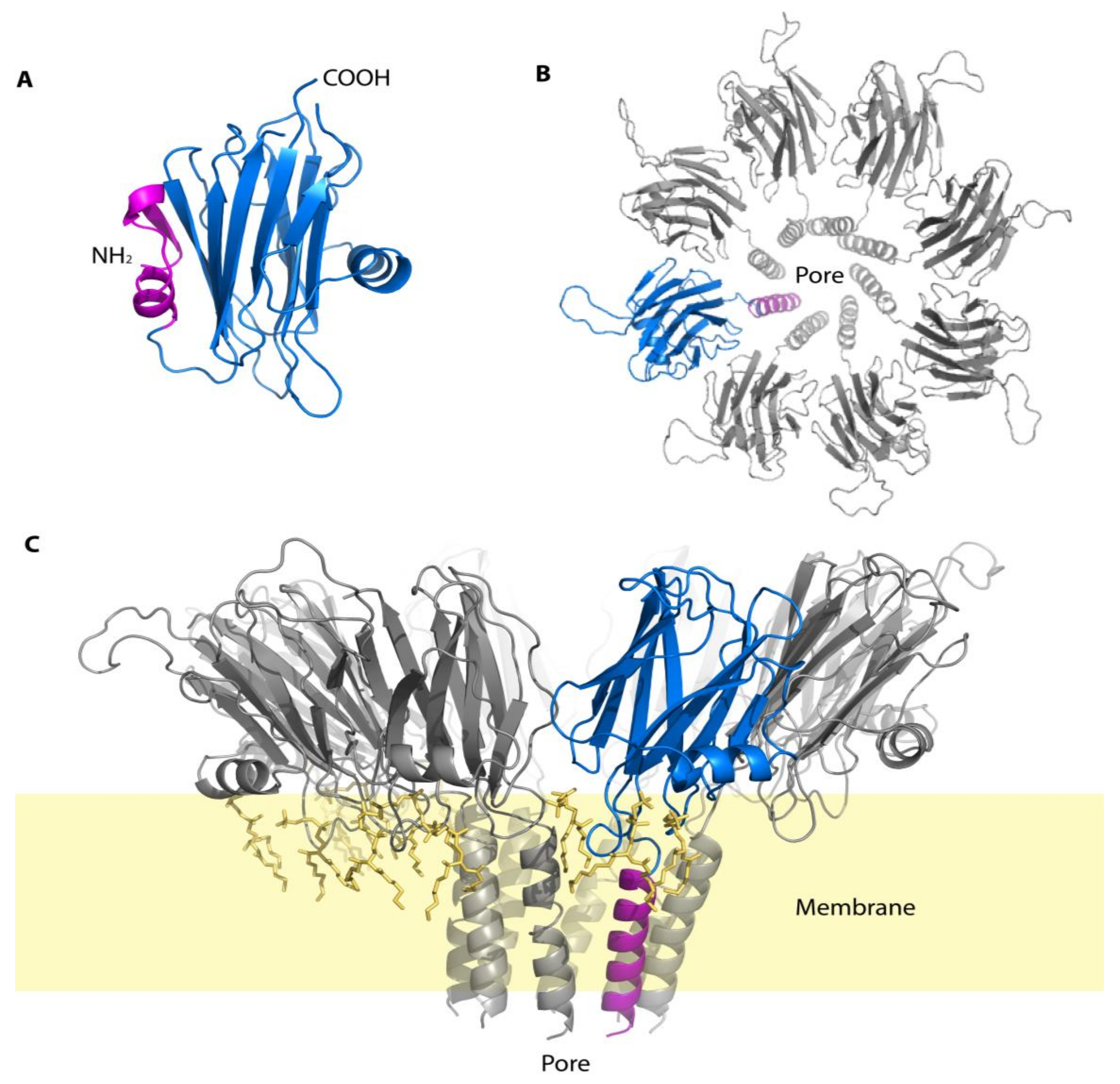
3.4. Echotoxins
4. Future Suggestions on Understudied Families
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Organism | Toxin | Accession Number |
|---|---|---|
| Monoplex echo | Echotoxin 2 | Q76CA2 |
| Cinguloterebra anilis | Tereporin-Ca1 | P0DN66 |
| Conus geographus | Conoporin 5 | W4VS02 |
| Actinia fragacea | Delta-actitoxin-Afr1a | B9W5G6 |
References
- Akondi, K.B.; Muttenthaler, M.; Dutertre, S.; Kaas, Q.; Craik, D.J.; Lewis, R.J.; Alewood, P.F. Discovery, synthesis, and structure-activity relationships of conotoxins. Chem. Rev. 2014, 114, 5815–5847. [Google Scholar] [CrossRef] [PubMed]
- Kaas, Q.; Craik, D.J. Conotoxins and other conopeptides. In Outstanding Marine Molecules: Chemistry, Biology, Analysis; Wiley-VCH Verlag GmbH and co.: Weinheim, Germany, 2014; Chapter 14; pp. 319–332. [Google Scholar]
- West, D.J.; Andrews, E.B.; Bowman, D.; McVean, A.R.; Thorndyke, M.C. Toxins from some poisonous and venemous marine snails. Comp. Biochem. Physiol. 1996, 113, 1–10. [Google Scholar]
- Duda, T.F., Jr.; Kohn, A.J.; Matheny, A.M. Cryptic species differentiated in Conus ebraeus, a widespread tropical marine gastropod. Biol. Bull. 2009, 217, 292–305. [Google Scholar] [CrossRef] [PubMed]
- Rockel, D.; Korn, W.; Kohn, A.J. The Indo-Pacific Region; Verlag: Wiesbaden, Germany, 1995; Volume 1. [Google Scholar]
- Terlau, H.; Olivera, B.M. Conus venoms: A rich source of novel ion channel-targeted peptides. Physiol. Rev. 2004, 84, 41–68. [Google Scholar] [CrossRef] [PubMed]
- Davis, J.; Jones, A.; Lewis, R.J. Remarkable inter- and intra-species complexity of conotoxins revealed by LC/MS. Peptides 2009, 30, 1222–1227. [Google Scholar] [CrossRef] [PubMed]
- Olivera, B.M. Conus venom peptides: Reflections from the biology of clades and species. Annu. Rev. Ecol. Syst. 2002, 33, 25–47. [Google Scholar] [CrossRef]
- Olivera, B.M.; Cruz, L.J. Conotoxins, in retrospect. Toxicon 2001, 39, 7–14. [Google Scholar] [CrossRef]
- Miljanich, G.P. Ziconotide: Neuronal calcium channel blocker for treating severe chronic pain. Curr. Med. Chem. 2004, 11, 3029–3040. [Google Scholar] [CrossRef] [PubMed]
- Holford, M.; Puillandre, N.; Terryn, Y.; Cruaud, C.; Olivera, B.; Bouchet, P. Evolution of the toxoglossa venom apparatus as inferred by molecular phylogeny of the Terebridae. Mol. Biol. Evol. 2009, 26, 15–25. [Google Scholar] [CrossRef] [PubMed]
- West, D.J.; Andrews, E.B.; McVean, A.R.; Osborne, D.J.; Thorndyke, M.C. Isolation of serotonin from the accessory glands of the marine snail Nucella lapillus. Toxicon 1994, 32, 1261–1264. [Google Scholar] [CrossRef]
- Barkalova, V.O.; Fedosov, A.E.; Kantor, Y.I. Morphology of the anterior digestive system of Tonnoideans (gastropoda: Caenogastropoda) with an emphasis on the foregut glands. Molluscan Res. 2016, 36, 54–73. [Google Scholar] [CrossRef]
- Ponte, G.; Modica, M.V. Salivary glands in predatory mollusks: Evolutionary considerations. Front. Physiol 2017, 8, 580. [Google Scholar] [CrossRef] [PubMed]
- Modica, M.V.; Holford, M. The neogastropoda: Evolutionary innovations of predatory marine snails with remarkable pharmacological potential. In Evolutionary Biology—Concepts, Molecular and Morphological; Pontarotti, P., Ed.; Springer: Berlin/Heidelberg, Germany, 2010. [Google Scholar]
- Tucker, J. Catalog of recent and fossil Turrids (mollusca: Gastropoda). Zootaxa 2004, 682, 1–1295. [Google Scholar]
- WoRMS. World Register of Marine Species. 2018. Available online: http://www.marinespecies.org/about.php (accessed on 1 November 2017).
- Imperial, J.S.; Kantor, Y.; Watkins, M.; Heralde, F.M., 3rd; Stevenson, B.; Chen, P.; Hansson, K.; Stenflo, J.; Ownby, J.P.; Bouchet, P.; et al. Venomous auger snail Hastula (Impages) hectica (linnaeus, 1758): Molecular phylogeny, foregut anatomy and comparative toxinology. J. Exp. Zool. B Mol. Dev. Evol. 2007, 308, 744–756. [Google Scholar] [CrossRef] [PubMed]
- Puillandre, N.; Holford, M. The terebridae and teretoxins: Combining phylogeny and anatomy for concerted discovery of bioactive compounds. BMC Chem. Biol. 2010, 10, 7. [Google Scholar] [CrossRef] [PubMed]
- Heralde, F.M., 3rd; Imperial, J.; Bandyopadhyay, P.K.; Olivera, B.M.; Concepcion, G.P.; Santos, A.D. A rapidly diverging superfamily of peptide toxins in venomous Gemmula species. Toxicon 2008, 51, 890–897. [Google Scholar] [CrossRef] [PubMed]
- Gorson, J.; Ramrattan, G.; Verdes, A.; Wright, E.M.; Kantor, Y.; Rajaram Srinivasan, R.; Musunuri, R.; Packer, D.; Albano, G.; Qiu, W.G.; et al. Molecular diversity and gene evolution of the venom arsenal of Terebridae predatory marine snails. Genome Biol. Evol. 2015, 7, 1761–1778. [Google Scholar] [CrossRef] [PubMed]
- Imperial, J.S.; Watkins, M.; Chen, P.; Hillyard, D.R.; Cruz, L.J.; Olivera, B.M. The augertoxins: Biochemical characterization of venom components from the toxoglossate gastropod Terebra subulata. Toxicon 2003, 42, 391–398. [Google Scholar] [CrossRef]
- Kaas, Q.; Westermann, J.C.; Craik, D.J. Conopeptide characterization and classifications: An analysis using conoserver. Toxicon 2010, 55, 1491–1509. [Google Scholar] [CrossRef] [PubMed]
- Czerwiec, E.; Kalume, D.E.; Roepstorff, P.; Hambe, B.; Furie, B.; Furie, B.C.; Stenflo, J. Novel gamma-carboxyglutamic acid-containing peptides from the venom of Conus textile. FEBS J. 2006, 273, 2779–2788. [Google Scholar] [CrossRef] [PubMed]
- Zugasti-Cruz, A.; Aguilar, M.B.; Falcon, A.; Olivera, B.M.; Heimer de la Cotera, E.P. Two new 4-Cys conotoxins (framework 14) of the vermivorous snail Conus austini from the Gulf of Mexico with activity in the central nervous system of mice. Peptides 2008, 29, 179–185. [Google Scholar] [CrossRef] [PubMed]
- Anand, P.; Grigoryan, A.; Bhuiyan, M.H.; Ueberheide, B.; Russell, V.; Quinonez, J.; Moy, P.; Chait, B.T.; Poget, S.F.; Holford, M. Sample limited characterization of a novel disulfide-rich venom peptide toxin from terebrid marine snail Terebra variegata. PLoS ONE 2014, 9, e94122. [Google Scholar] [CrossRef] [PubMed]
- Moon, J.; Gorson, J.; Wright, M.E.; Yee, L.; Khawaja, S.; Shin, H.Y.; Karma, Y.; Musunri, R.L.; Yun, M.; Holford, M. Characterization and recombinant expression of terebrid venom peptide from Terebra guttata. Toxins (Basel) 2016, 8, 63. [Google Scholar] [CrossRef] [PubMed]
- Kendel, Y.; Melaun, C.; Kurz, A.; Nicke, A.; Peigneur, S.; Tytgat, J.; Wunder, C.; Mebs, D.; Kauferstein, S. Venomous secretions from marine snails of the Terebridae family target acetylcholine receptors. Toxins (Basel) 2013, 5, 1043–1050. [Google Scholar] [CrossRef] [PubMed]
- Bouchet, P.; Kantor, Y.I.; Sysoev, A.; Puillandre, N. A new operational classification of the Conoidea (gastropoda). J. Moll. Stud. 2011, 46, 273–308. [Google Scholar] [CrossRef]
- Bouchet, P.; Waren, A. Revision of the Northeast Atlantic bathyal and abyssal Turridae (mollusca, gastropoda). J. Moll. Stud. 1980, 46, 1–119. [Google Scholar] [CrossRef]
- Lopez-Vera, E.; Heimer de la Cotera, E.P.; Maillo, M.; Riesgo-Escovar, J.R.; Olivera, B.M.; Aguilar, M.B. A novel structural class of toxins: The methionine-rich peptides from the venoms of Turrid marine snails (mollusca, conoidea). Toxicon 2004, 43, 365–374. [Google Scholar] [CrossRef] [PubMed]
- Watkins, M.; Hillyard, D.R.; Olivera, B.M. Genes expressed in a Turrid venom duct: Divergence and similarity to conotoxins. J. Mol. Evol. 2006, 62, 247–256. [Google Scholar] [CrossRef] [PubMed]
- Omaga, C.A.; Carpio, L.D.; Imperial, J.S.; Daly, N.L.; Gajewiak, J.; Flores, M.S.; Espino, S.S.; Christensen, S.; Filchakova, O.M.; Lopez-Vera, E.; et al. Structure and biological activity of a turripeptide from Unedogemmula bisaya venom. Biochemistry 2017, 56, 6051–6060. [Google Scholar] [CrossRef] [PubMed]
- Emmelin, N.; Fange, R. Comparison between biological effects of neurine and a salivary gland extract of Neptunea antiqua. Acta Zool. 1958, 39, 47–52. [Google Scholar] [CrossRef]
- Fange, R. Venoms and venom glands of marine molluscs. In Toxins, Drugs, and Pollutants in Marine Animals; Bolis, L., Zadunaisky, J., Gilles, R., Eds.; Springer-Verlag: Berlin, Germany, 1984; pp. 47–62. [Google Scholar]
- Shiomi, K.; Mizukami, M.; Shimakura, K.; Nagashima, Y. Toxins in the salivary gland of some marine carnivorous gastropods Comp. Biochem. Physiol. 1994, 107B, 427–432. [Google Scholar]
- Shiomi, K.; Kawashima, Y.; Mizukami, M.; Nagashima, Y. Properties of proteinaceous toxins in the salivary gland of the marine gastropod (Monoplex echo). Toxicon 2002, 40, 563–571. [Google Scholar] [CrossRef]
- Day, J.A. Feeding of the cymatiid gastropod, Argobuccinum argus, in relation to the structure of the proboscis and secretions of the proboscis gland. Am. Zool. 1969, 9, 909–916. [Google Scholar] [CrossRef][Green Version]
- Anthoni, U.; Bohlin, L.; Larsen, C.; Nielsen, P.; Nielsen, N.H.; Christophersen, C. Tetramine: Occurrence in marine organisms and pharmacology. Toxicon 1989, 27, 707–716. [Google Scholar] [CrossRef]
- Asano, M.; Ito, M. Occurrence of tetramine and choline compounds in the salivary gland of a marine gastropod Neptunea arthritica, Bernardi. Tohuku J. Agric. Res. 1959, 10, 209–227. [Google Scholar]
- Anthoni, U.; Bohlin, L.; Larsen, C.; Nielsen, P.; Nielsen, N.H.; Christophersen, C. The toxin tetramine from the “edible” whelk Neptunea antiqua. Toxicon 1989, 27, 717–723. [Google Scholar] [CrossRef]
- Reid, T.M.; Gould, I.M.; Mackie, I.M.; Ritchie, A.H.; Hobbs, G. Food poisoning due to the consumption of red whelks (Neptunea antiqua). Epidemiol. Infect. 1988, 101, 419–423. [Google Scholar] [CrossRef] [PubMed]
- Welsh, J.H. Composition and mode of action of some invertebrate venoms. Annu. Rev. Pharmacol. 1964, 4, 293–304. [Google Scholar] [CrossRef]
- Sarumathi, G.; Arumugam, M.; Kumaresan, S.; Balasubramanian, T. Studies on bioprospecting potential of a gastropod mollusc Cantharus tranquebaricus (Gmelin, 1791). Asian Pac. J. Trop. Biomed. 2012, 2, 759–764. [Google Scholar] [CrossRef]
- Endean, R. Aspects of Molluscan Pharmacology; Academic Press: New York, NY, USA, 1972; Volume 7. [Google Scholar]
- Himmelman, J.H.; Hamel, J.R. Diet, behavior, and reproduction of the whelk Buccinum undatum in the northern gulf of st. Lawrence, eastern canada. Mar. Biol. 1993, 116, 423–430. [Google Scholar] [CrossRef]
- Cornman, I. Toxic properties of the saliva of Cassis. Nature 1963, 200, 88–89. [Google Scholar] [CrossRef]
- Modica, M.V.; Lombardo, F.; Franchini, P.; Oliverio, M. The venomous cocktail of the vampire snail Colubraria reticulata (mollusca, gastropoda). BMC Genom. 2015, 16, 441. [Google Scholar] [CrossRef] [PubMed]
- Voultsiadou, E. Therapeutic properties and uses of marine invertebrates in the ancient Greek world and early Byzantium. J. Ethnopharmacol. 2010, 130, 237–247. [Google Scholar] [CrossRef] [PubMed]
- Benkendorff, K.; Rudd, D.; Nongmaithem, B.D.; Liu, L.; Young, F.; Edwards, V.; Avila, C.; Abbott, C.A. Are the traditional medical uses of muricidae mulluscs substantiated by their pharmacological properties and bioactive compounds? Mar. Drugs 2015, 13, 5237–5275. [Google Scholar] [CrossRef] [PubMed]
- Benkendorff, K.; McIver, C.; Abbott, C. Bioactivity of the murex homeopathic remedy and of extracts from an Australian muricid mollusc against human cancer cells. J. Evid. Based Complement. Altern. Med. 2009, 2011, 1–12. [Google Scholar]
- Vine, K.L.; Locke, J.M.; Ranson, M.; Benkendorff, K.; Pyne, S.G.; Bremner, J.B. In vitro cytotoxicity evaluation of some substituted isatin derivatives. Bioorg. Med. Chem. 2007, 15, 931–938. [Google Scholar] [CrossRef] [PubMed]
- Edwards, V.; Benkendorff, K.; Young, F. Marine compounds selectively induce apoptosis in female reproductive cancer cells but not in primary-derived human reproductive granulosa cells. Mar. Drugs 2012, 10, 64–83. [Google Scholar] [CrossRef] [PubMed]
- Benkendorff, K. Natural product research in the Australian marine invertebrate Dicathais orbita. Mar. Drugs 2013, 11, 1370–1398. [Google Scholar] [CrossRef] [PubMed]
- Roseghini, M.; Severini, C.; Erspamer, G.F.; Erspamer, V. Choline esters and biogenic amines in the hypobranchial gland of 55 molluscan species of the neogastropod muricoidea superfamily. Toxicon 1996, 34, 33–55. [Google Scholar] [CrossRef]
- Andrews, E.B.; Elphick, M.R.; Thorndyke, M. Pharmacologically active constituents of the accessory salivary and hypobranchial glands of Nucella lapillus. J. Moll. Stud. 1991, 57, 136–138. [Google Scholar] [CrossRef]
- Andrews, E.B. The fine structure and function of the salivary glands of Nucella lapillus (gastropoda: Muricidae). J. Moll. Stud. 1991, 57, 111–126. [Google Scholar] [CrossRef]
- Andrews, E.B.; Page, A.M.; Taylor, J.D. The fine structure and function of the anterior foregut glands of Cymatium intermedius (cassoidea: Ranellidae). J. Moll. Stud. 1999, 65, 1–19. [Google Scholar] [CrossRef][Green Version]
- Hemingway, G.T. Evidence for a paralytic venom in the intertidal snail, Acanthina spirata (neogastropoda: Thaisidae). Comp. Biochem. Physiol. 1977, 60, 78–81. [Google Scholar] [CrossRef]
- Huang, C.L.; Mir, G.N. Pharmacological properties of hypobranchial gland of Thais haemastoma (clench). J. Pharm. Sci. 1971, 60, 1842–1846. [Google Scholar] [CrossRef] [PubMed]
- Morton, B. Foregut anatomy and predation by Charonia lampas (gastropoda: Prosobranchia: Neotaenioglossa) attacking Ophidiaster ophidianus (asteroidea: Ophidiasteridae) in the azores, with a review of triton feeding behavior. J. Nat. Hist. 2012, 46, 2621–2637. [Google Scholar] [CrossRef]
- Chau, R.; Kalaitzis, J.A.; Neilan, B.A. On the origins and biosynthesis of tetrodotoxin. Aquat. Toxicol. 2011, 104, 61–72. [Google Scholar] [CrossRef] [PubMed]
- Bose, U.; Wang, T.; Zhao, M.; Motti, C.A.; Hall, M.R.; Cummins, S.F. Multiomics analysis of the giant triton snail salivary gland, a crown-of-thorns starfish predator. Sci. Rep. 2017, 7, 6000. [Google Scholar] [CrossRef] [PubMed]
- Edgar, R.C. Muscle: A multiple sequence alignment method with reduced time and space complexity. BMC Bioinf. 2004, 5, 113. [Google Scholar] [CrossRef] [PubMed]
- Berg, J.M.; Tymoczko, J.L.; Stryer, L. Biochemistry; Freeman, W.H.: New York, NY, USA, 2002. [Google Scholar]
- Guimaraes, D.O.; Lopes, D.S.; Azevedo, F.V.; Gimenes, S.N.; Silva, M.A.; Ache, D.C.; Gomes, M.S.; Vecchi, L.; Goulart, L.R.; Yoneyama, K.A.; et al. In Vitro antitumor and antiangiogenic effects of bothropoidin, a metalloproteinase from Bothrops pauloensis snake venom. Int. J. Biol. Macromol. 2017, 97, 770–777. [Google Scholar] [CrossRef] [PubMed]
- Warren, W.C.; Hillier, L.W.; Marshall Graves, J.A.; Birney, E.; Ponting, C.P.; Grutzner, F.; Belov, K.; Miller, W.; Clarke, L.; Chinwalla, A.T.; et al. Genome analysis of the platypus reveals unique signatures of evolution. Nature 2008, 453, 175–183. [Google Scholar] [CrossRef] [PubMed]
- Bose, U.; Suwansa-Ard, S.; Maikaeo, L.; Motti, C.A.; Hall, M.R.; Cummins, S.F. Neuropeptides encoded within a neural transcriptome of the giant triton snail Charonia tritonis, a crown-of-thorns starfish predator. Peptides 2017, 98, 3–14. [Google Scholar] [CrossRef] [PubMed]
- Cruz, L.J.; de Santos, V.; Zafaralla, G.C.; Ramilo, C.A.; Zeikus, R.; Gray, W.R.; Olivera, B.M. Invertebrate vasopressin/oxytocin homologs. Characterization of peptides from Conus geographus and Conus straitus venoms. J. Biol. Chem. 1987, 262, 15821–15824. [Google Scholar] [PubMed]
- van Soest, P.F.; Kits, K.S. Conopressin affects excitability, firing, and action potential shape through stimulation of transient and persistent inward currents in mulluscan neurons. J. Neurophysiol. 1998, 79, 1619–1632. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Kawashima, Y.; Nagai, H.; Ishida, M.; Nagashima, Y.; Shiomi, K. Primary structure of echotoxin 2, an actinoporin-like hemolytic toxin from the salivary gland of the marine gastropod Monoplex echo. Toxicon 2003, 42, 491–497. [Google Scholar] [CrossRef]
- West, D.J.; Andrews, E.B.; McVean, A.R.; Thorndyke, M.C.; Taylor, J.D. Presence of a toxin in the salivary glands of the marine snail Cymatium intermedius that targets nicotinic acetylcholine receptors. Toxicon 1998, 36, 25–29. [Google Scholar] [CrossRef]
- Bellomio, A.; Morante, K.; Barlic, A.; Gutierrez-Aguirre, I.; Viguera, A.R.; Gonzalez-Manas, J.M. Purification, cloning and characterization of fragaceatoxin C, a novel actinoporin from the sea anemone Actinia fragacea. Toxicon 2009, 54, 869–880. [Google Scholar] [CrossRef] [PubMed]
- Anderluh, G.; Macek, P. Cytolytic peptide and protein toxins from sea anemones (anthozoa: Actiniaria). Toxicon 2002, 40, 111–124. [Google Scholar] [CrossRef]
- Violette, A.; Biass, D.; Dutertre, S.; Koua, D.; Piquemal, D.; Pierrat, F.; Stocklin, R.; Favreau, P. Large-scale discovery of conopeptides and conoproteins in the injectable venom of a fish-hunting cone snail using a combined proteomic and transcriptomic approach. J. Proteom. 2012, 75, 5215–5225. [Google Scholar] [CrossRef] [PubMed]
- Luo, X.; Yu, R.C.; Wang, X.J.; Zhou, M.J. Toxin composition and toxicity dynamics of marine gastropod Nassarius spp. Collected from lianyungang, china. Food Addit. Contam. Part A Chem. Anal. Control Expo. Risk Assess. 2012, 29, 117–127. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.J.; Yu, R.C.; Luo, X.; Zhou, M.J.; Lin, X.T. Toxin-screening and identification of bacteria isolated from highly toxic marine gastropod Nassarius semiplicatus. Toxicon 2008, 52, 55–61. [Google Scholar] [CrossRef] [PubMed]
- Costa, P.R.; Costa, S.T.; Braga, A.C.; Rodrigues, S.M.; Vale, P. Relevance and challenges in monitoring marine biotoxins in non-bivalve vectors. Food Control 2017, 76, 24–33. [Google Scholar] [CrossRef]
- Borysko, L.; Ross, P.M. Adult exposure to the synthetic hormone 17alpha-ethynylestradiol affects offspring of the gastropods Nassarius burchardi and Nassarius jonasii. Ecotoxicol. Environ. Saf. 2014, 103, 91–100. [Google Scholar] [CrossRef] [PubMed]
- Choi, M.C.; Yu, P.K.; Hsieh, D.P.; Lam, P.K. Trophic transfer of paralytic shellfish toxins from clams (Ruditapes philippinarum) to gastropods (Nassarius festivus). Chemosphere 2006, 64, 1642–1649. [Google Scholar] [CrossRef] [PubMed]
- Coelho, J.P.; Pimenta, J.; Gomes, R.; Barroso, C.M.; Pereira, M.E.; Pardal, M.A.; Duarte, A. Can Nassarius reticulatus be used as a bioindicator for Hg contamination? Results from a longitudinal study of the Portuguese coastline. Mar. Pollut. Bull. 2006, 52, 674–680. [Google Scholar] [CrossRef] [PubMed]
- Hwang, P.A.; Tsai, Y.H.; Deng, J.F.; Cheng, C.A.; Ho, P.H.; Hwang, D.F. Identification of tetrodotoxin in a marine gastropod (Nassarius glans) responsible for human morbidity and mortality in Taiwan. J. Food Prot. 2005, 68, 1696–1701. [Google Scholar] [CrossRef] [PubMed]
- Zou, S.; Li, Q.; Kong, L. Monophyly, distance and character-based multigene barcoding reveal extraordinary cryptic diversity in Nassarius: A complex and dangerous community. PLoS ONE 2012, 7, e47276. [Google Scholar] [CrossRef] [PubMed]
- Minniti, F. Morphological and histochemical study of pharynx of Leiblein, salivary glands, and gland of Leiblein in the carnivorous gastropoda Amyclina tinei maravigna and Cyclope neritea larnarck (nassariidae: Prosobranchia stenoglossa). Zool. Anz. 1986, 217, 14–22. [Google Scholar]
- Huang, H.N.; Lin, J.; Lin, H.L. Identification and quantification of tetrodotoxin in the marine gastropod Nassarius by LC-MS. Toxicon 2008, 51, 774–779. [Google Scholar] [CrossRef] [PubMed]
- Galindo, L.A.; Puillandre, N.; Utge, J.; Lozouet, P.; Bouchet, P. The phylogeny and systematics of the Nassariidae revisited (gastropoda, buccinoidea). Mol. Phylogenet. Evol. 2016, 99, 337–353. [Google Scholar] [CrossRef] [PubMed]
- Neidleman, S.L.; Laskin, A.I. Advances in Applied Microbiology; Academic Press: Cambridge, MA, USA, 1997; Chapter 2; Volume 43. [Google Scholar]
- Teichert, R.W.; Olivera, B.M. Natural products and ion channel pharmacology. Future Med. Chem. 2010, 2, 731–744. [Google Scholar] [CrossRef] [PubMed]
- Williams, A.J.; Dave, J.R.; Phillips, J.B.; Lin, Y.; McCabe, R.T.; Tortella, F.C. Neuroprotective efficacy and therapeutic window of the high-affinity N-methyl-d-aspartate antagonist conantokin-G: In vitro (primary cerebellar neurons) and in vivo (rat model of transient focal brain ischemia) studies. J. Pharmacol. Exp. Ther. 2000, 294, 378–386. [Google Scholar] [PubMed]
- Balsara, R.; Dang, A.; Donahue, D.L.; Snow, T.; Castellino, F.J. Conantokin-g attenuates detrimental effects of nmdar hyperactivity in an ischemic rat model of stroke. PLoS ONE 2015, 10, e0122840. [Google Scholar] [CrossRef] [PubMed]
- Halai, R.; Craik, D.J. Conotoxins: Natural product drug leads. Nat. Prod. Rep. 2009, 26, 526–536. [Google Scholar] [CrossRef] [PubMed]
- Benkendorff, K.; Bremner, J.B.; Davis, A.R. Tyrian purple precursors in the egg masses of the Australian muricid, Dicathais orbita: A possible defensive role. J. Chem. Ecol. 2000, 26, 1037–1050. [Google Scholar] [CrossRef]
- Esmaeelian, B. Preclinical In Vitro and In Vivo Effects of Purified and Synthetic Bioactive Compounds from Marine Mollusc Dicathais orbita on Colorectal Cancer: Cancer Prevention and Toxicity Study; Flinders University: Adelaide, Australia, 2013. [Google Scholar]
- Esmaeelian, B.; Abbott, C.A.; Le Leu, R.K.; Benkendorff, K. 6-bromoisatin found in muricid mollusc extracts inhibits colon cancer cell proliferation and induces apoptosis, preventing early stage tumor formation in a colorectal cancer rodent model. Mar. Drugs 2014, 12, 17–35. [Google Scholar] [CrossRef] [PubMed]
- Esmaeelian, B.; Benkendorff, K.; Le Leu, R.K.; Abbott, C.A. Simultaneous assessment of the efficacy and toxicity of marine mollusc-derived brominated indoles in an in vivo model for early stage colon cancer. Integr. Cancer Ther. 2017, 1534735417699880. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.K.; Park, G.M. Indirubin-3-monoxime exhibits anti-inflammatory properties by down-regulating Nf-kappab and JNK signaling pathways in lipopolysaccharide-treated RAW264.7 cells. Inflamm. Res. 2012, 61, 319–325. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, T.B.; Rudd, D.; Smith, J.; Kotiw, M.; Mouatt, P.; Seymour, L.M.; Liu, L.; Benkendorff, K. Anti-inflammatory activity and structure-activity relationships of brominated indoles from a marine mollusc. Mar. Drugs 2017, 15, 133. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, T.B.; Rudd, D.; Benkendorff, K.; Mahdi, L.K.; Pratt, K.A.; Dooley, L.; Wei, C.; Kotiw, M. Brominated indoles from a marine mollusc inhibit inflammation in a murine model of acute lung injury. PLoS ONE 2017, 12, e0186904. [Google Scholar] [CrossRef] [PubMed]
- Stevens, R.W.; Morita, H.; Nakane, N. Indole Derivatives as Antiallergy and Antiinflammatory Agents. U.S. Patent US 5290788A, 1 March 1994. [Google Scholar]
- Erspamer, V.; Glasser, A. The pharmacological actions of murexine (urocanylcholine). Br. J. Pharmacol. Chemother. 1957, 12, 176–184. [Google Scholar] [CrossRef] [PubMed]
- Erspamer, V.; Glasser, A. The pharmacological actions of some murexine-like substances. Br. J. Pharmacol. Chemother. 1958, 13, 378–384. [Google Scholar] [CrossRef] [PubMed]
- Peraro, M.D.; van der Goot, F.G. Pore-forming toxins: Ancient, but never really out of fashion. Nat. Rev. 2016, 14, 77–92. [Google Scholar] [CrossRef] [PubMed]
- Dyer, P.D.R.; Shepherd, T.R.; Gollings, A.S.; Shorter, S.A.; Gorringe-Pattrick, M.A.M.; Tang, C.K.; Cattoz, B.N.; Baillie, L.; Griffiths, P.C.; Richardson, S.C.W. Disarmed anthrax toxin delivers antisense oligonucleotides and sirna with high efficiency and low toxicity. J. Control Release 2015, 220, 316–328. [Google Scholar] [CrossRef] [PubMed]
- Barbier, J.; Gillet, D. Engineering of Bacterial Toxins for Research and Medicine. In The Comprehensive Sourcebook of Bacterial Protein Toxins; Institute of Biology and Technology of Saclay: Gif sur Yvette, France, 2015; Chapter 35. [Google Scholar]
- Liao, X.; Rabideau, A.E.; Pentelute, B.L. Delivery of antibody mimics into mammalian cells via anthrax toxin protective antigen. ChemBioChem 2014, 15, 2458–2466. [Google Scholar] [CrossRef] [PubMed]
- Rabideau, A.E.; Pentelute, B.L. Delivery of non-native cargo into mammalian cells using anthrax lethal toxin. ACS Chem. Biol. 2016, 11, 1490–1501. [Google Scholar] [CrossRef] [PubMed]
- Verdermen, W.; Luginbuhl, M.; Honegger, A.; Pluckthum, A. Effecient cell-specific uptake of binding proteins into the cytoplasm through engineered modular transport systems. J. Control. Release 2015, 200, 13–22. [Google Scholar] [CrossRef] [PubMed]
- Dosio, F.; Stella, B.; Cerioni, S.; Gastaldi, D.; Arpicco, S. Advances in anticancer antibody-drug conjugates and immunotoxins. Recent Pat. Anticancer Drug Discov. 2014, 9, 35–65. [Google Scholar] [CrossRef] [PubMed]
- Kasianowicz, J.J.; Balijepalli, A.K.; Ettedgui, J.; Forstater, J.H.; Wang, H.; Zhang, H.; Robertson, J.W. Analytical applications for pore-forming proteins. Biochim. Biophys. Acta 2016, 1858, 593–606. [Google Scholar] [CrossRef] [PubMed]
- Mukherjee, P.; Faber, A.C.; Shelton, L.M.; Baek, R.C.; Chiles, T.C.; Seyfried, T.N. Thematic review series: Sphingolipids. Ganglioside GM3 suppresses the proangiogenic effects of vascular endothelial growth factor and ganglioside GD1A. J. Lipid Res. 2008, 49, 929–938. [Google Scholar] [CrossRef] [PubMed]
- Yu, R.K.; Tsai, Y.T.; Ariga, T.; Yanagisawa, M. Structures, biosynthesis, and functions of gangliosides—An overview. J. Oleo Sci. 2011, 60, 537–544. [Google Scholar] [CrossRef] [PubMed]
- Lorizate, M.; Brugger, B.; Akiyama, H.; Glass, B.; Muller, B.; Anderluh, G.; Wieland, F.T.; Krausslich, H.G. Probing HIV-1 membrane liquid order by laurdan staining reveals producer cell-dependent differences. J. Biol. Chem. 2009, 284, 22238–22247. [Google Scholar] [CrossRef] [PubMed]
- Yamaji-Hasegawa, A.; Hullin-Matsuda, F.; Greimel, P.; Kobayashi, T. Pore-forming toxins: Properties, diversity, and uses as tools to image sphingomyelin and ceramide phosphoethanolamine. Biochim. Biophys. Acta 2016, 1858, 576–592. [Google Scholar] [CrossRef] [PubMed]
- Matsuzaki, K.; Kato, K.; Yanagisawa, K. Abeta polymerization through interaction with membrane gangliosides. Biochim. Biophys. Acta 2010, 1801, 868–877. [Google Scholar] [CrossRef] [PubMed]
- Harasewych, M.G. Anatomy and biology of Mitra cornea lamarck, 1811 (mollusca, caenogastropoda, mitridae) from the azores. Acoreana 2009, 6, 121–135. [Google Scholar]
- West, T.L. Feeding behavior and functional morphology of the epiproboscis of Mitra idae. Bull. Mar. Sci. 1990, 46, 761–779. [Google Scholar]
- Taylor, J.D. The diet of coral-reef Mitridae (gastropoda) from guam; with a review of other species of the family. J. Nat. Hist. 1989, 23, 261–278. [Google Scholar] [CrossRef]







| Family | Species | Toxin |
|---|---|---|
| Conidae | Various | Conopeptides, CRiSPs, metallopreases |
| Terebridae | Hastula hectica | Teretoxins |
| Terebra subulata | Teretoxins | |
| Terebra argus | Teretoxins | |
| Terebra guttata | Teretoxins | |
| Cinguloterebra anilis | Teretoxins, cytolytic proteins/actinoporins | |
| Terebra consobrina | Teretoxins, cytolytic proteins/actinoporins | |
| Turridae | Lophiotoma olangoensis | Turritoxins |
| Polystira albida | Turritoxins | |
| Gemmula periscelida | Turritoxins | |
| Buccinidae | Buccinum leucostoma | Tetramine, cholines/murexines |
| Buccinum schantaricum | Tetramine, unknown nervous system depressant? Stimulant? | |
| Buccinum undatum | Cholines/murexines | |
| Neptunea antiqua | Tetramine, unknown nervous system depressant | |
| Neptunea arthritica | Tetramine | |
| Neptunea intersculpta | Tetramine | |
| Neptunea. kuroshiro | Tetramine | |
| Neptunea lyrata | Tetramine | |
| Cantharus tranquebaricus | Antimicrobial, cytolytic activity? | |
| Babylonidae | Babylonia japonica | Surugatoxin |
| Muricidae | Hexaplex trunculus | Choline esters/murexines, brominated indoles |
| Ocenebra erinaceus | Choline esters/murexines, brominated indoles | |
| Bolinus brandaris | Choline esters/murexines, brominated indoles | |
| Nucella lapillus | CRiSPs? Choline esters/murexines, brominated indoles, unknown nervous system depressant? Stimulant? | |
| Stramonita haemastoma | Unknown nervous system depressant? Stimulant? | |
| Acanthinucella spirata | Cholines/murexines, unknown nervous system depressant | |
| Cassidae | Cassis tuberosa | Unknown nervous system depressant |
| Cassis madagascarensis | Unknown nervous system depressant | |
| Cassis flammea | Unknown nervous system depressant | |
| Colubraridae | Cumia reticulata | Metalloproteases, cytolytic proteins/actinoporins |
| Ranellidae | Charonia lampas | Cytolytic proteins/actinoporins, unknown neurostimulant |
| Charonia tritonis | CRiSPs, metalloproteases, echotoxins | |
| Fusitriton oregonensis | Tetramine | |
| Monoplex intermedius | Unknown neurostimulant | |
| Monoplex parthenopeus echo | Echotoxins |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Turner, A.H.; Craik, D.J.; Kaas, Q.; Schroeder, C.I. Bioactive Compounds Isolated from Neglected Predatory Marine Gastropods. Mar. Drugs 2018, 16, 118. https://doi.org/10.3390/md16040118
Turner AH, Craik DJ, Kaas Q, Schroeder CI. Bioactive Compounds Isolated from Neglected Predatory Marine Gastropods. Marine Drugs. 2018; 16(4):118. https://doi.org/10.3390/md16040118
Chicago/Turabian StyleTurner, Ashlin H., David J. Craik, Quentin Kaas, and Christina I. Schroeder. 2018. "Bioactive Compounds Isolated from Neglected Predatory Marine Gastropods" Marine Drugs 16, no. 4: 118. https://doi.org/10.3390/md16040118
APA StyleTurner, A. H., Craik, D. J., Kaas, Q., & Schroeder, C. I. (2018). Bioactive Compounds Isolated from Neglected Predatory Marine Gastropods. Marine Drugs, 16(4), 118. https://doi.org/10.3390/md16040118