
Degree of Suppression of Mouse Myoblast Cell Line C2C12 Differentiation Varies According to Chondroitin Sulfate Subtype


{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
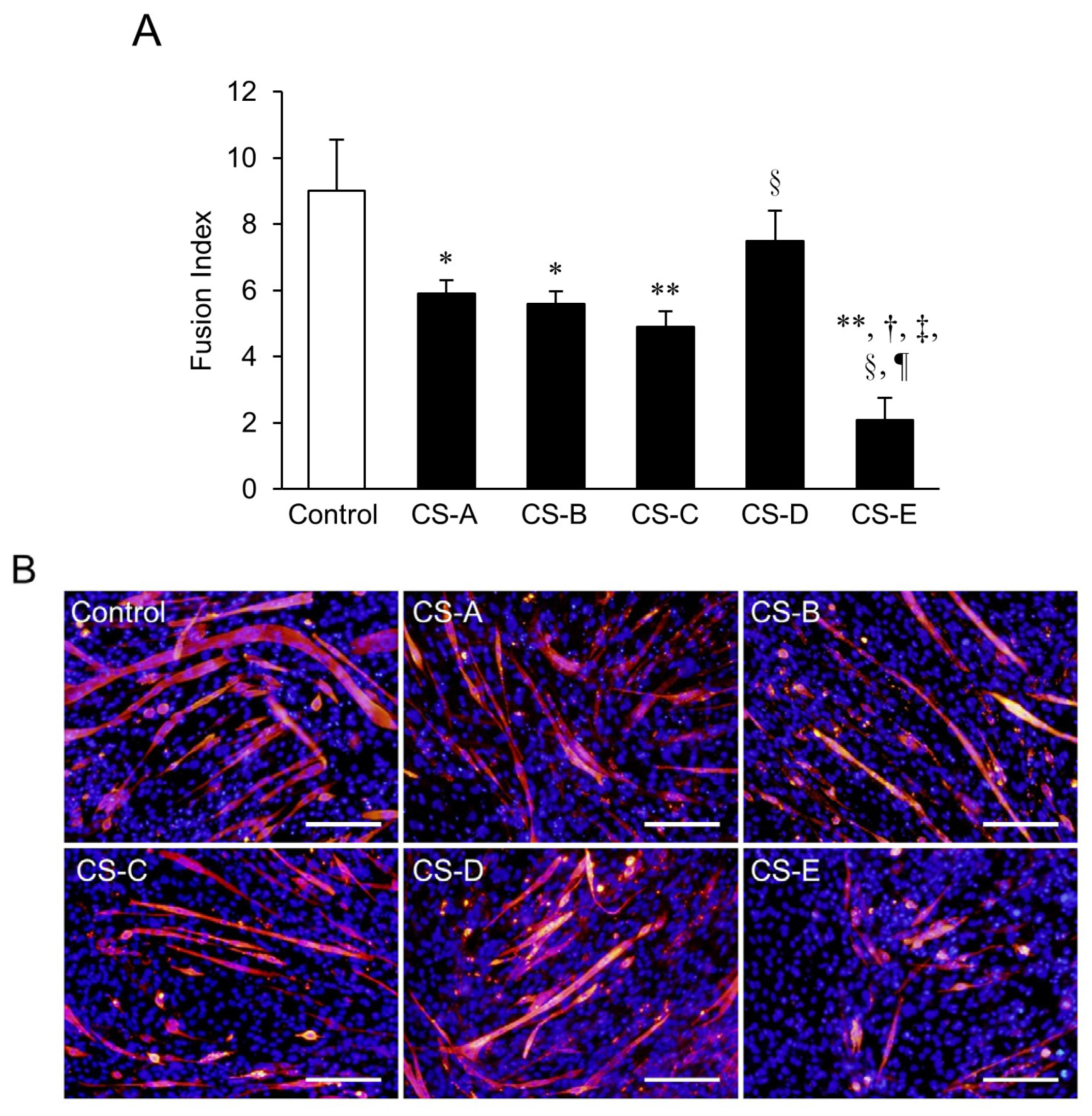
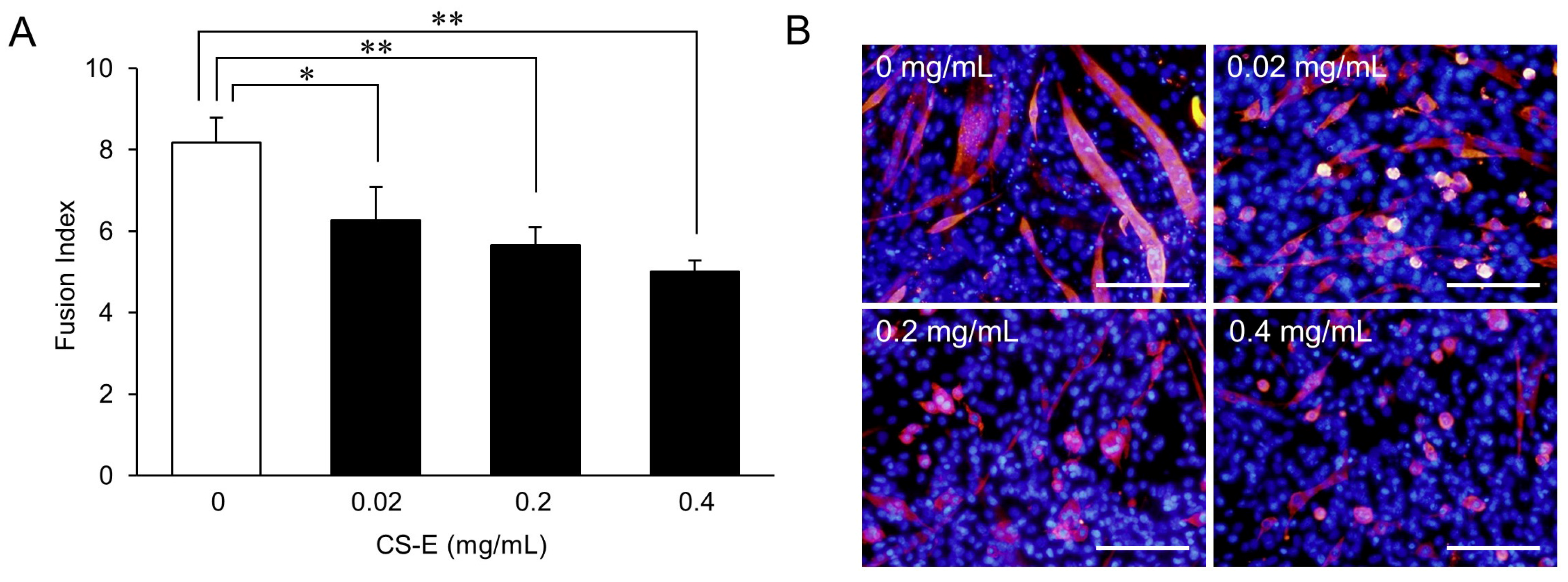
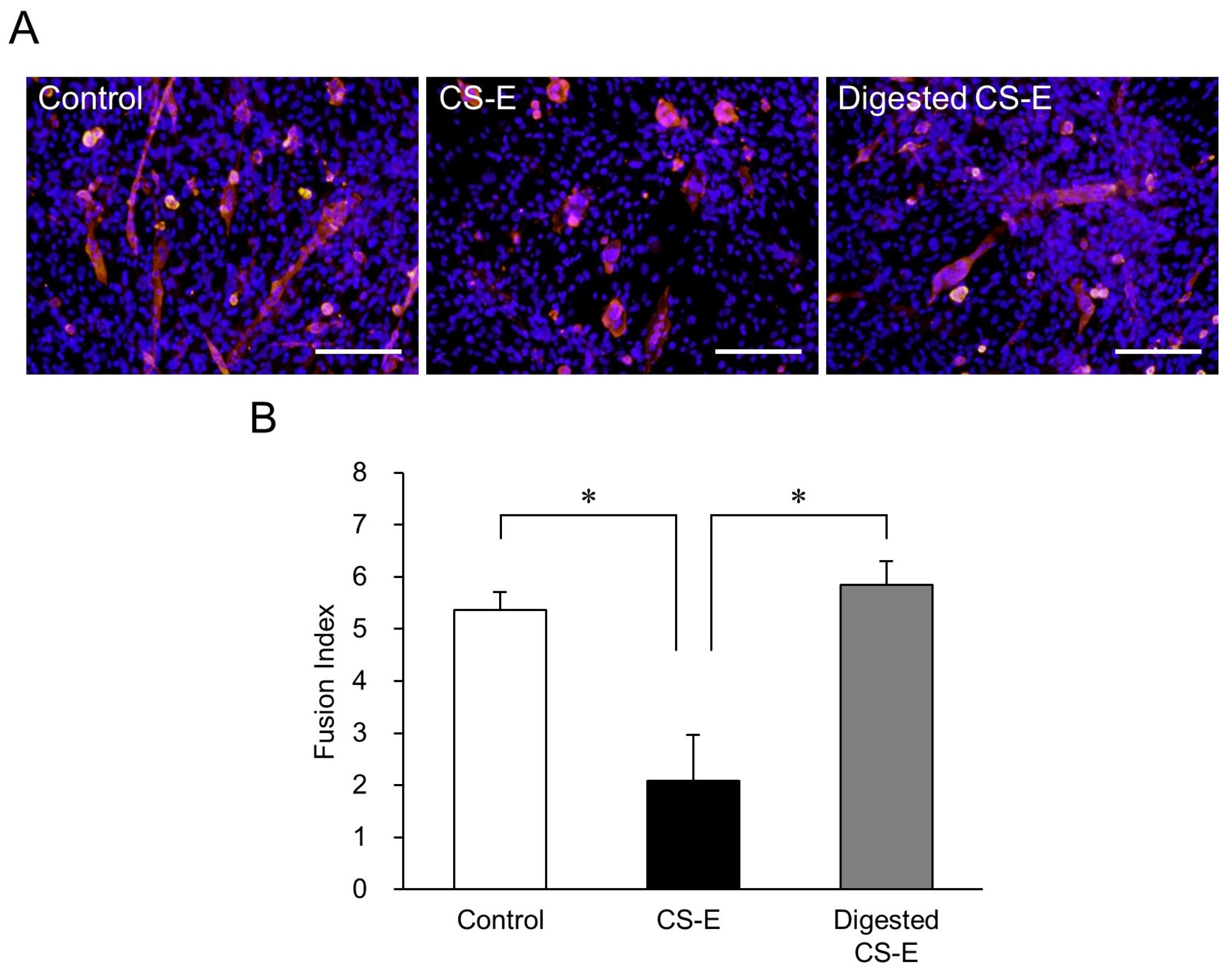
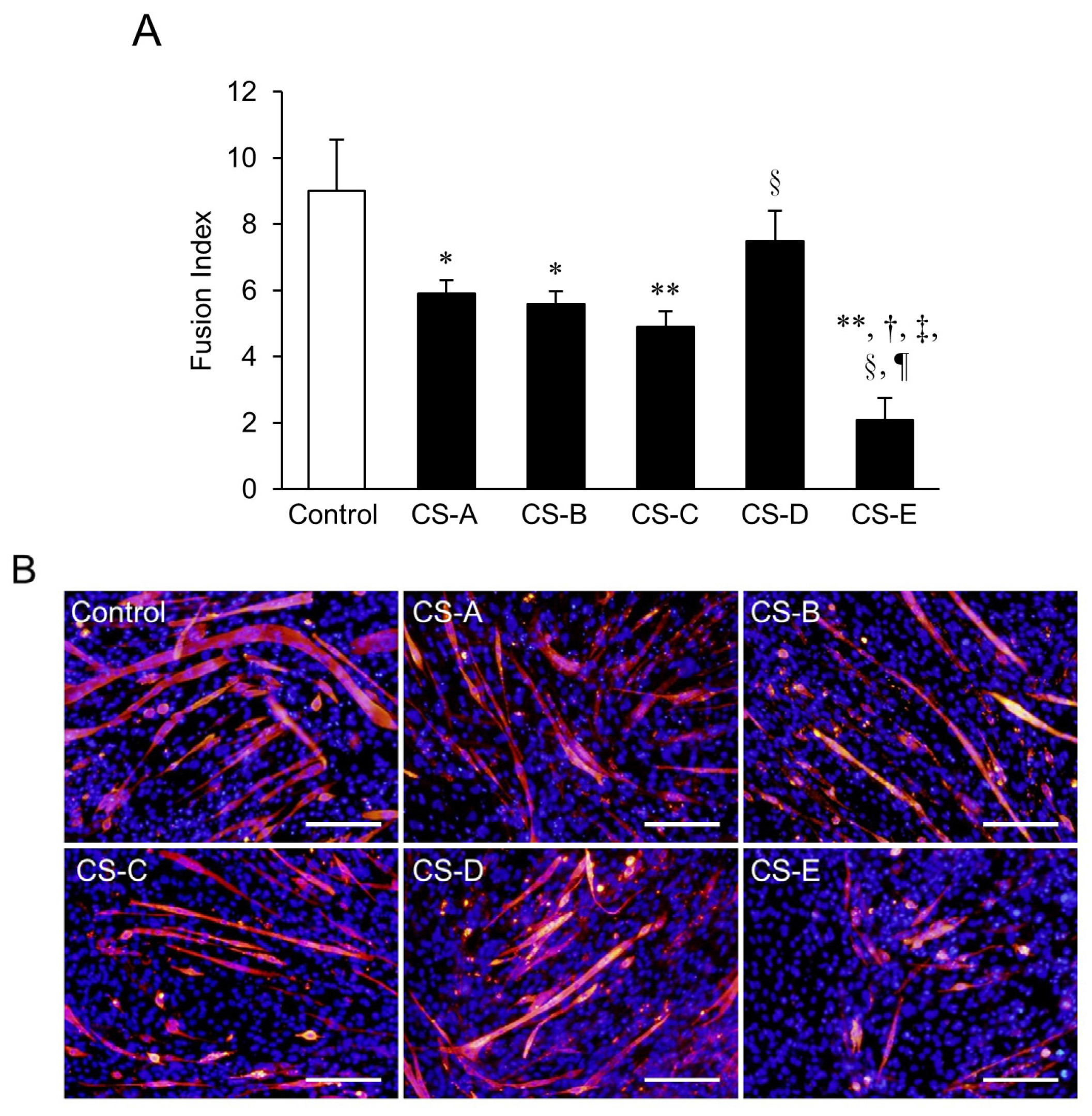
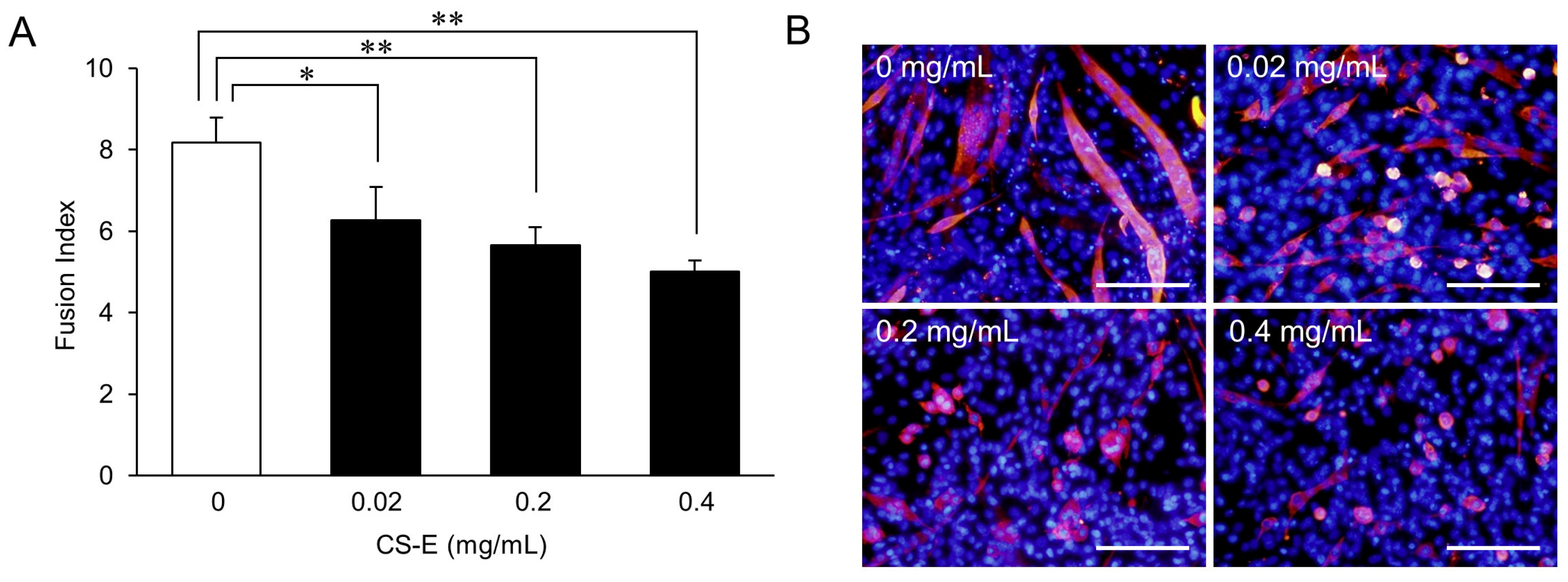
2. Results and Discussion
Suppression of Myotube Formation by CS
3. Materials and Methods
3.1. Cell Culture and Induction of Myogenic Differentiation
3.2. Immunocytochemistry
3.3. Assessment of Muscle Differentiation
3.4. Identification of CS Subtypes Affecting Differentiation
3.5. Dose-Response Effects of CS-E
3.6. Analysis of the Effects of the CS-E Glycan Region on Myotube Formation
3.7. Statistical Analysis
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Chen, E.H.; Grote, E.; Mohler, W.; Vignery, A. Cell-cell fusion. FEBS Lett. 2007, 581, 2181–2193. [Google Scholar] [CrossRef] [PubMed]
- Rochlin, K.; Yu, S.; Roy, S.; Baylies, M.K. Myoblast fusion: When it takes more to make one. Dev. Biol. 2010, 341, 66–83. [Google Scholar] [CrossRef] [PubMed]
- Seale, P.; Rudnicki, M.A. A new look at the origin, function and “stem-cell” status of muscle satellite cells. Dev. Biol. 2000, 218, 115–124. [Google Scholar] [CrossRef] [PubMed]
- Relaix, F.; Zammit, P.S. Satellite cells are essential for skeletal muscle regeneration: The cell on the edge returns centre stage. Development 2012, 139, 2845–2856. [Google Scholar] [CrossRef] [PubMed]
- Hindi, S.M.; Tajrishi, M.M.; Kumar, A. Signaling mechanisms in mammalian myoblast fusion. Sci. Signal. 2013, 6. [Google Scholar] [CrossRef] [PubMed]
- Mikami, T.; Kitagawa, H. Biosynthesis and function of chondroitin sulfate. Biochim. Biophys. Acta 2013, 1830, 4719–4733. [Google Scholar] [CrossRef] [PubMed]
- Prydz, K. Determinants of glycosaminoglycan (GAG) structure. Biomolecules 2015, 5, 2003–2022. [Google Scholar] [CrossRef] [PubMed]
- Miyachi, K.; Wakao, M.; Suda, Y. Syntheses of chondroitin sulfate tetrasaccharide structures containing 4,6-disulfate patterns and analysis of their interaction with glycosaminoglycan-binding protein. Bioorg. Med. Chem. Lett. 2015, 25, 1552–1555. [Google Scholar] [CrossRef] [PubMed]
- Sugahara, K.; Mikami, T.; Uyama, T.; Mizuguchi, S.; Nomura, K.; Kitagawa, H. Recent advances in the structural biology of chondroitin sulfate and dermatan sulfate. Curr. Opin. Struct. Biol. 2003, 13, 612–620. [Google Scholar] [CrossRef] [PubMed]
- Izumikawa, T.; Kanagawa, N.; Watamoto, Y.; Okada, M.; Saeki, M.; Sakano, M.; Sugahara, K.; Sugihara, K.; Asano, M.; Kitagawa, H. Impairment of embryonic cell division and glycosaminoglycan biosynthesis in glycuronyltransferase-I-deficient mice. J. Biol. Chem. 2010, 285, 12190–12196. [Google Scholar] [CrossRef] [PubMed]
- Sugiura, N.; Shioiri, T.; Chiba, M.; Sato, T.; Narimatsu, H.; Kimata, K.; Watanabe, H. Construction of a chondroitin sulfate library with defined structures and analysis of molecular interactions. J. Biol. Chem. 2012, 287, 43390–43400. [Google Scholar] [CrossRef] [PubMed]
- Mikami, T.; Koyama, S.; Yabuta, Y.; Kitagawa, H. Chondroitin sulfate is a crucial determinant for skeletal muscle development/regeneration and improvement of muscular dystrophies. J. Biol. Chem. 2012, 287, 38531–38542. [Google Scholar] [CrossRef] [PubMed]
- Chargé, S.B.; Rudnicki, M.A. Cellular and molecular regulation of muscle regeneration. Physiol. Rev. 2004, 84, 209–238. [Google Scholar] [CrossRef] [PubMed]
- Pavlath, G.K. Spatial and functional restriction of regulatory molecules during mammalian myoblast fusion. Exp. Cell Res. 2010, 316, 3067–3072. [Google Scholar] [CrossRef] [PubMed]
- Sugahara, K.; Yamada, S. Structure and function of oversulfated chondroitin sulfate variants: Unique sulfation patterns and neuroregulatory activities. Trends Glycosci. Glycotechnol. 2000, 12, 321–349. [Google Scholar] [CrossRef]
- Ueoka, C.; Kaneda, N.; Okazaki, I.; Nadanaka, S.; Muramatsu, T.; Sugahara, K. Neuronal cell adhesion, mediated by the heparin-binding neuroregulatory factor midkine, is specifically inhibited by chondroitin sulfate E. J. Biol. Chem. 2000, 275, 37407–37413. [Google Scholar] [CrossRef] [PubMed]
- Deepa, S.S.; Umehara, Y.; Higashiyama, S.; Itoh, N.; Sugahara, K. Specific molecular interactions of oversulfated chondroitin sulfate E with various heparin-binding growth factors. Implications as a physiological binding partner in the brain and other tissues. J. Biol. Chem. 2002, 277, 43707–43716. [Google Scholar] [CrossRef] [PubMed]
- Takagaki, K.; Munakata, H.; Kakizaki, I.; Iwafune, M.; Itabashi, T.; Endo, M. Domain structure of chondroitin sulfate E octasaccharides binding to type V collagen. J. Biol. Chem. 2002, 277, 8882–8889. [Google Scholar] [CrossRef] [PubMed]
- Millay, D.P.; O’Rourke, J.R.; Sutherland, L.B.; Bezprozvannaya, S.; Shelton, J.M.; Bassel-Duby, R.; Olson, E.N. Myomaker is a membrane activator of myoblast fusion and muscle formation. Nature 2013, 499, 301–305. [Google Scholar] [CrossRef] [PubMed]
- Millay, D.P.; Sutherland, L.B.; Bassel-Duby, R.; Olson, E.N. Myomaker is essential for muscle regeneration. Genes Dev. 2014, 28, 1641–1646. [Google Scholar] [CrossRef] [PubMed]
- Fongmoon, D.; Shetty, A.K.; Basappa; Yamada, S.; Sugiura, M.; Kongtawelert, P.; Sugahara, K. Chondroitinase-mediated degradation of rare 3-O-sulfated glucuronic acid in functional oversulfated chondroitin sulfate K and E. J. Biol. Chem. 2007, 282, 36895–36904. [Google Scholar] [CrossRef] [PubMed]

© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Warita, K.; Oshima, N.; Takeda-Okuda, N.; Tamura, J.-i.; Hosaka, Y.Z. Degree of Suppression of Mouse Myoblast Cell Line C2C12 Differentiation Varies According to Chondroitin Sulfate Subtype. Mar. Drugs 2016, 14, 193. https://doi.org/10.3390/md14100193
Warita K, Oshima N, Takeda-Okuda N, Tamura J-i, Hosaka YZ. Degree of Suppression of Mouse Myoblast Cell Line C2C12 Differentiation Varies According to Chondroitin Sulfate Subtype. Marine Drugs. 2016; 14(10):193. https://doi.org/10.3390/md14100193
Chicago/Turabian StyleWarita, Katsuhiko, Nana Oshima, Naoko Takeda-Okuda, Jun-ichi Tamura, and Yoshinao Z. Hosaka. 2016. "Degree of Suppression of Mouse Myoblast Cell Line C2C12 Differentiation Varies According to Chondroitin Sulfate Subtype" Marine Drugs 14, no. 10: 193. https://doi.org/10.3390/md14100193