Cytotoxic and Apoptosis-Inducing Activity of Triterpene Glycosides from Holothuria scabra and Cucumaria frondosa against HepG2 Cells

Abstract
:1. Introduction
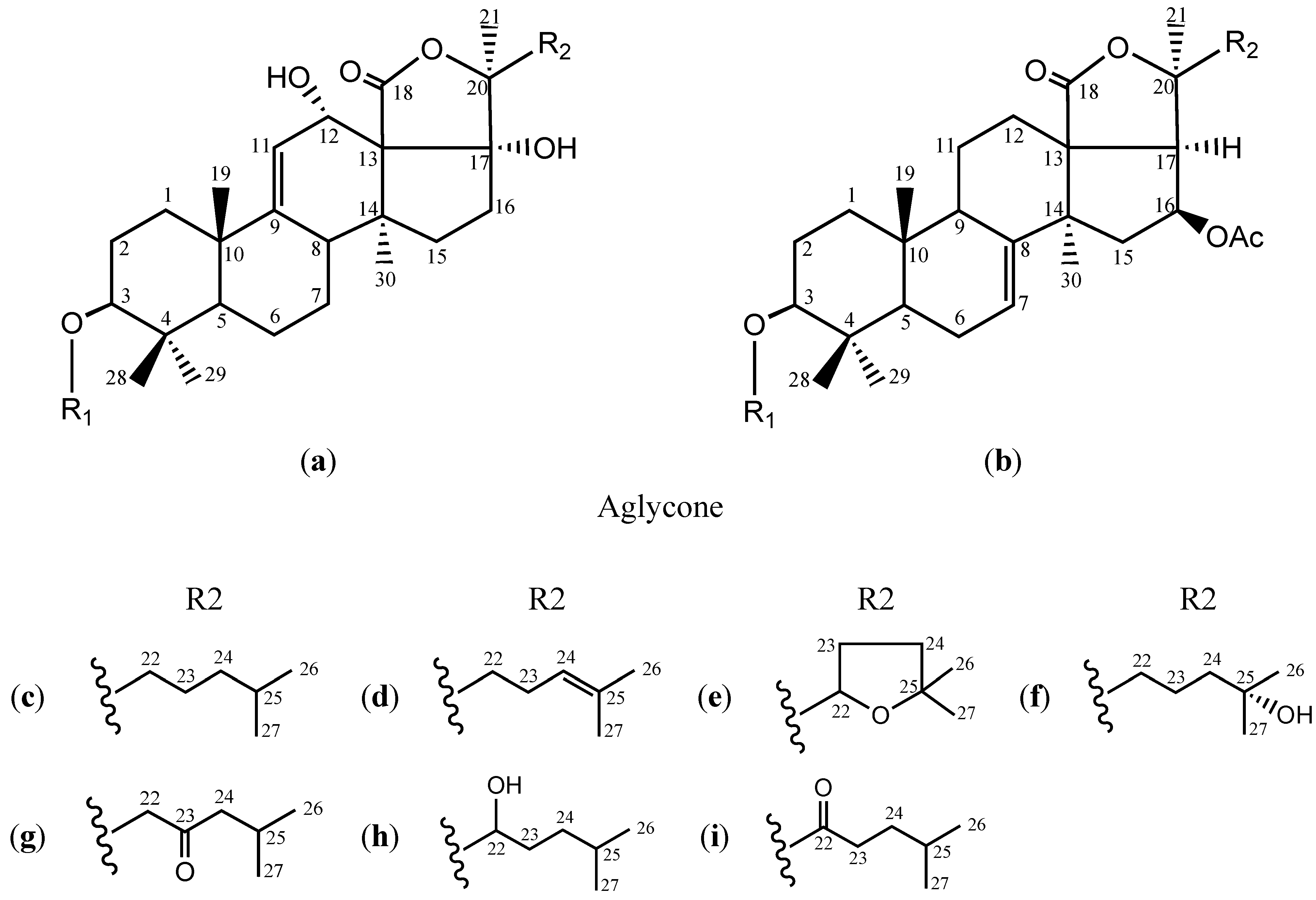

{kind=link}
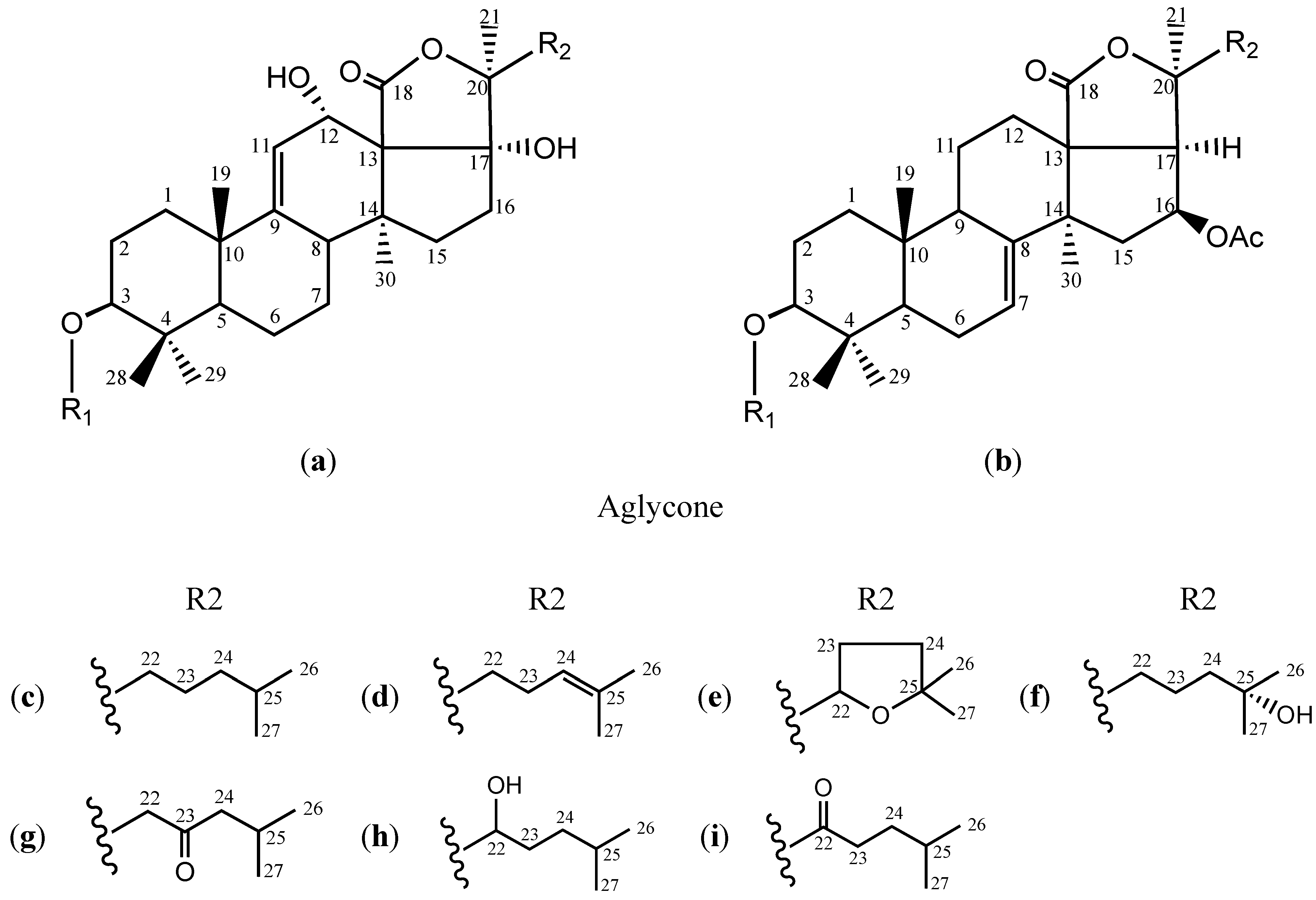
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Glycosides | Aglycone | R1 | R2 | MF | MR | Reference |
|---|---|---|---|---|---|---|
| Echinoside A (1) | a | S2 | c | C54H87O26NaS | 1206 | [22] |
| 24-Dehydro echinoside A (2) | S2 | d | C54H85O26NaS | 1204 | [23] | |
| Holothurin B (3) | S1 | e | C41H62O17NaS | 881 | [22] | |
| Holothurin B4 (4) | S1 | f | C41H65O17NaS | 884 | ||
| HS-1 (5) | S2 | g | C54H85O27NaS | 1220 | [24] | |
| Holothurin A (6) | S2 | e | C54H85O27NaS | 1220 | [22] | |
| Holothurin A1 (7) | S2 | h | C54H87O27NaS | 1222 | [24] | |
| Scabraside D (8) | S2 | f | C54H87O27NaS | 1222 | [23] | |
| Scabraside B (9) | S2 | i | C54H85O27NaS | 1220 | [25] | |
| Frondoside A1 (10) | b | S3 | c | C55H87O25NaS | 1202 | [26] |
| Frondoside A (11) | S4 | c | C60H95O29NaS | 1334 | [26] | |
| Frondoside A6 (12) | S5 | c | C60H95O29HS | 1312 | [26] | |
| 24-Dehydro frondoside A6 (13) | S5 | d | C60H93O29HS | 1310 |
2. Results and Discussion
2.1. Cytotoxicity against Tumor Cells and Structure-Activity Relationships
| Glycosides | IC50 (μg/mL) | Ratio (HL-7702/HepG2) | |||
|---|---|---|---|---|---|
| HeLa | K562 | HepG2 | HL-7702 | ||
| Echinoside A (1) | 1.25 ± 0.16 | 1.61 ± 0.11 | 1.50 ± 0.08 | 3.12 ± 0.65 * | 2.08 |
| 24-Dehydro echinoside A (2) | 2.06 ± 0.27 | 6.15 ± 0.24 | 1.95 ± 0.09 | 3.75 ± 0.40 ** | 1.92 |
| Holothurin B (3) | 2.05 ± 0.11 | 3.64 ± 0.04 | 1.79 ± 0.02 | 2.69 ± 0.45 * | 1.50 |
| Holothurin B4 (4) | 2.71 ± 0.54 | 3.55 ± 0.53 | 2.71 ± 0.12 | 4.58 ± 0.54 ** | 1.69 |
| HS-1 (5) | 3.20 ± 0.32 | 12.95 ± 1.54 | 6.10 ± 0.33 | 7.19 ± 0.20 ** | 1.18 |
| Holothurin A (6) | 3.76 ± 0.47 | 8.94 ± 0.01 | 3.46 ± 0.33 | 3.85 ± 0.32 | 1.11 |
| Holothurin A1 (7) | 2.84 ± 0.09 | 6.50 ± 0.01 | 2.90 ± 0.21 | 3.59 ± 0.05 ** | 1.24 |
| Scabraside D (8) | 3.84 ± 0.42 | 10.06 ± 1.06 | 3.33 ± 0.33 | 4.40 ± 0.22 ** | 1.32 |
| Scabraside B (9) | 4.44 ± 0.85 | 11.85 ± 2.26 | 7.29 ± 0.62 | 11.03 ± 1.37 * | 1.51 |
| Frondoside A1 (10) | 2.03 ± 0.17 | 2.21 ± 0.56 | 1.91 ± 0.03 | 4.96 ± 1.10 ** | 2.60 |
| Frondoside A (11) | 3.30 ± 0.05 | 3.76 ± 0.05 | 4.14 ± 0.57 | 5.42 ± 0.98 | 1.31 |
| Frondoside A6 (12) | 3.75 ± 0.51 | 6.59 ± 0.11 | 2.53 ± 0.66 | 6.05 ± 0.67 ** | 2.39 |
| 24-Dehydro frondoside A6 (13) | 3.16 ± 0.34 | 5.61 ± 1.05 | 3.57 ± 0.54 | 7.55 ± 0.83 ** | 2.12 |
| CDDP (positive control) | 3.41 ± 0.37 | 2.99 ± 0.24 | 2.05 ± 0.13 | 4.48 ± 0.41 *** | 2.18 |
2.2. Selective Cytotoxicity on Tumor and Normal Cells
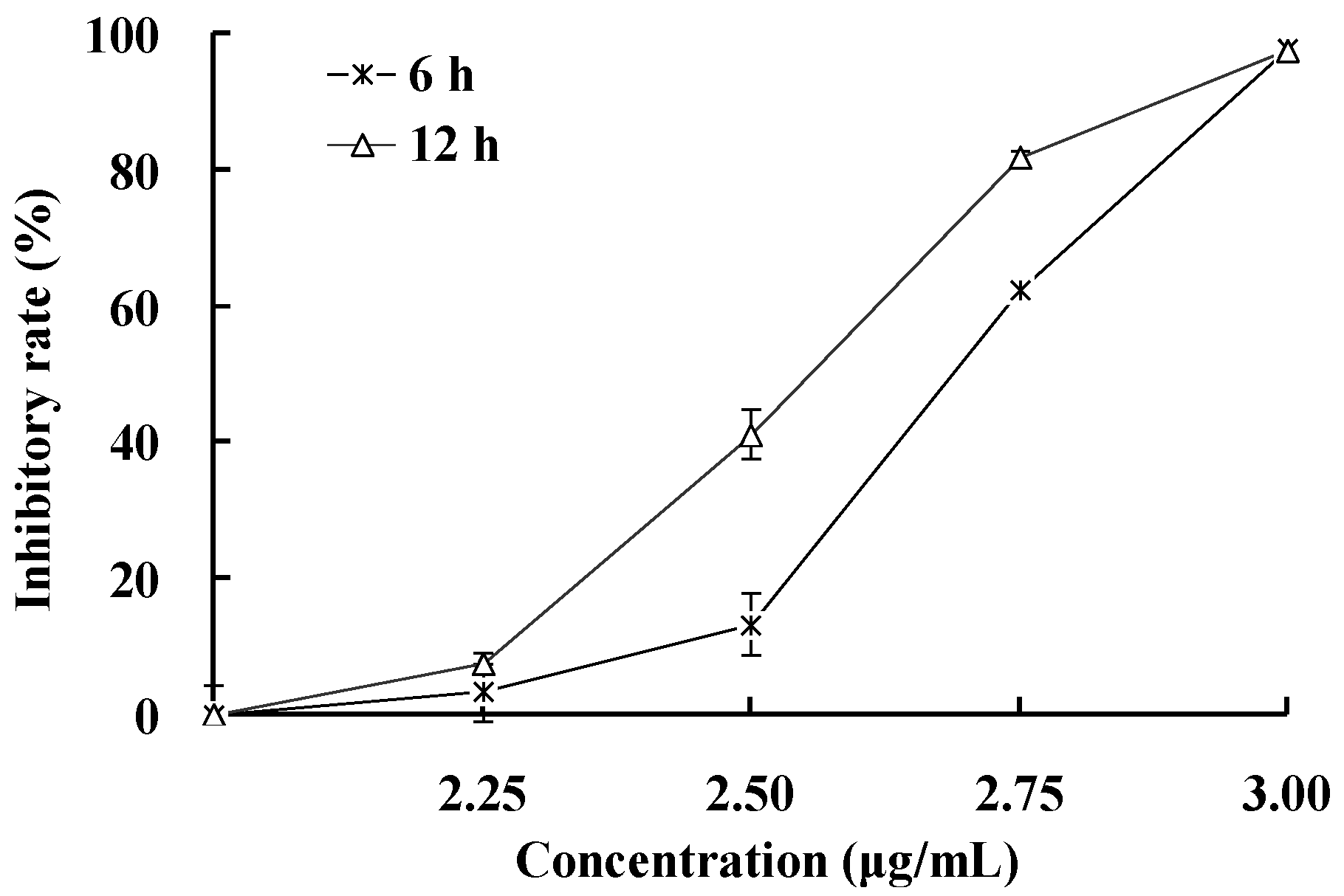
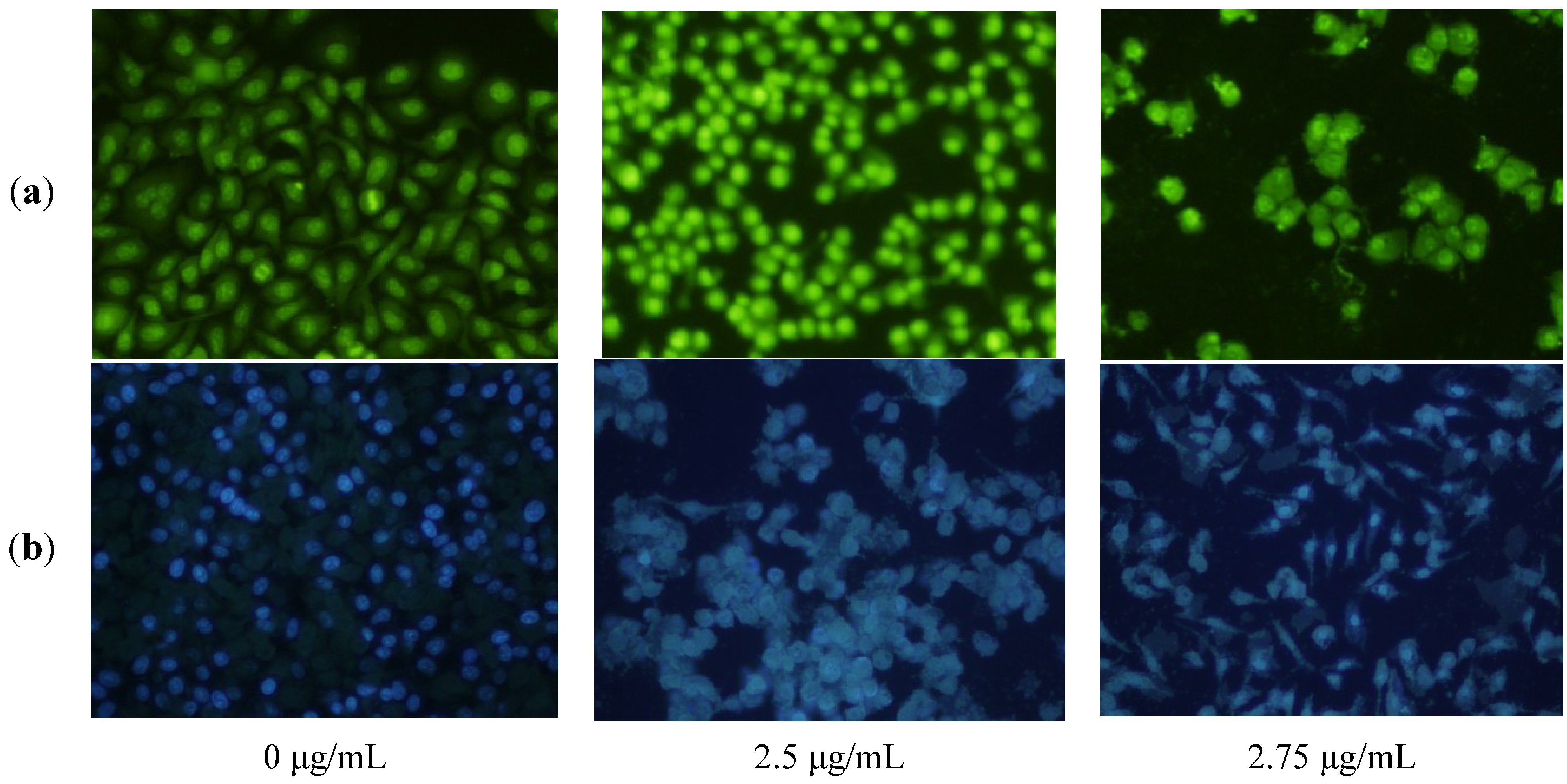
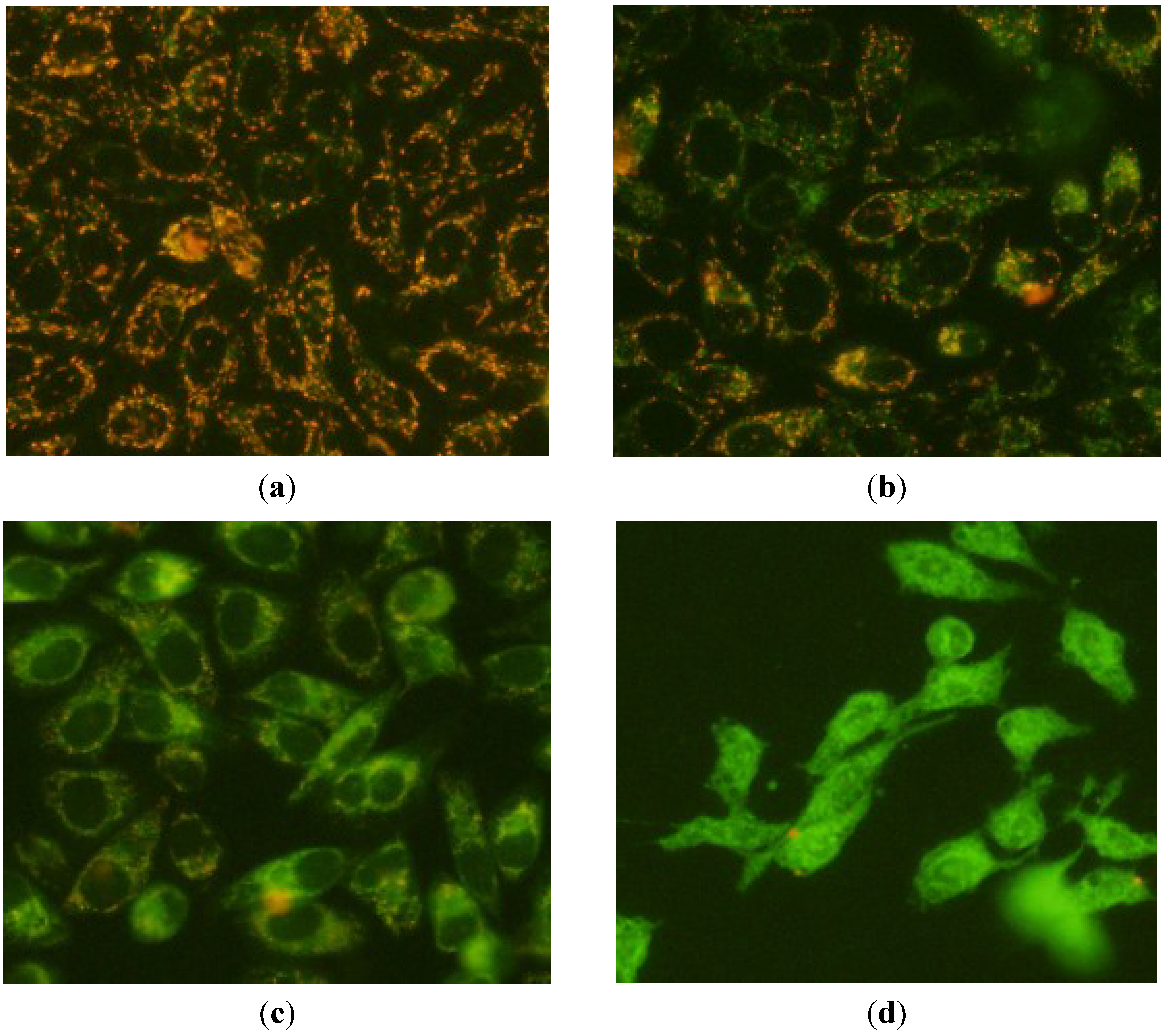
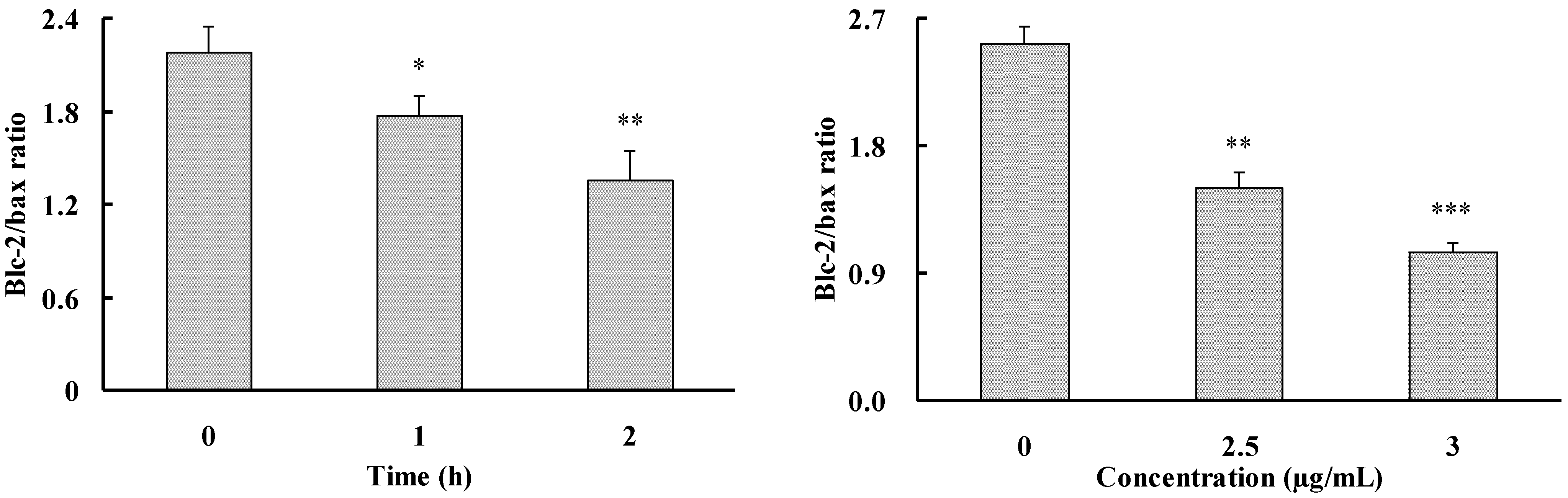
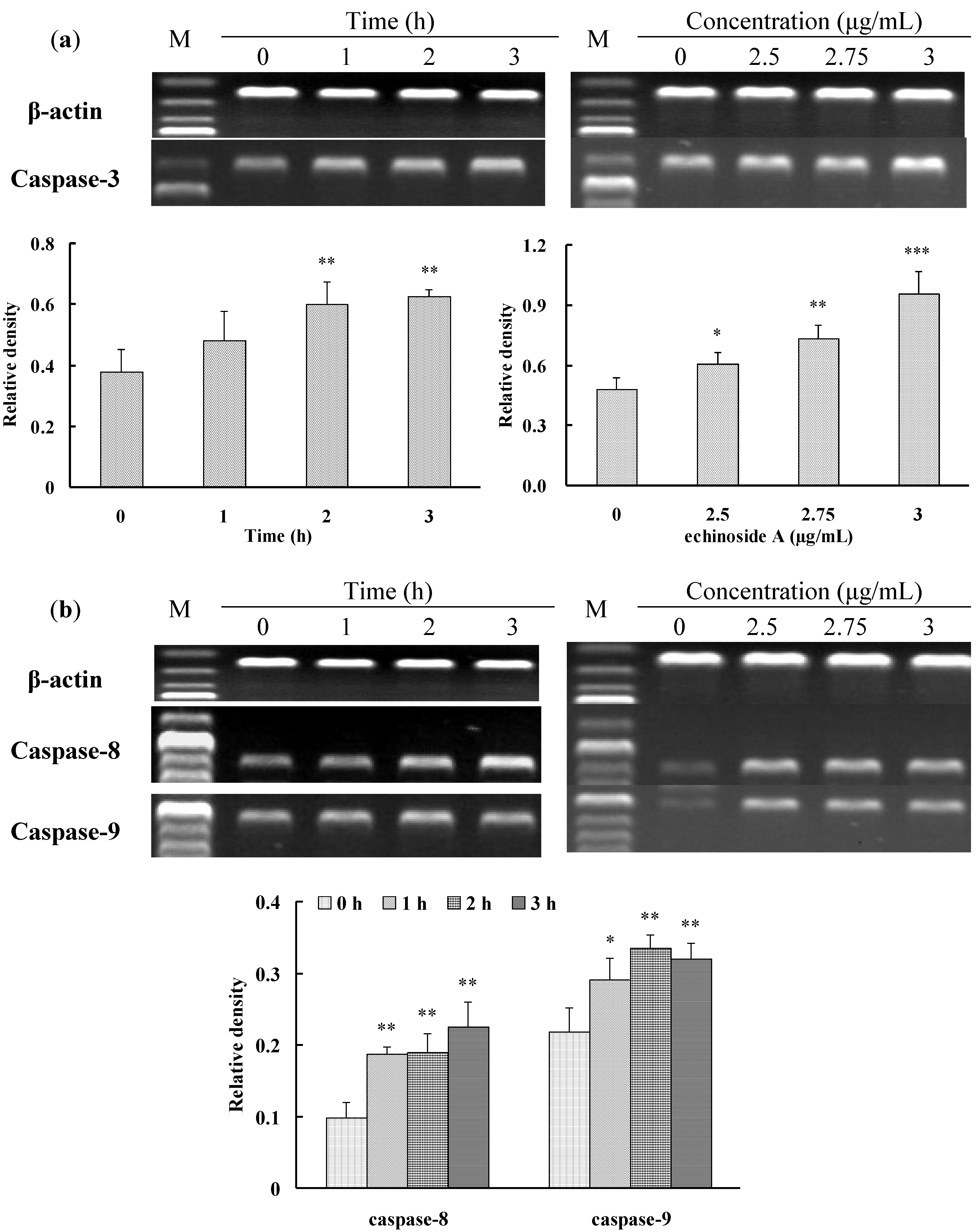
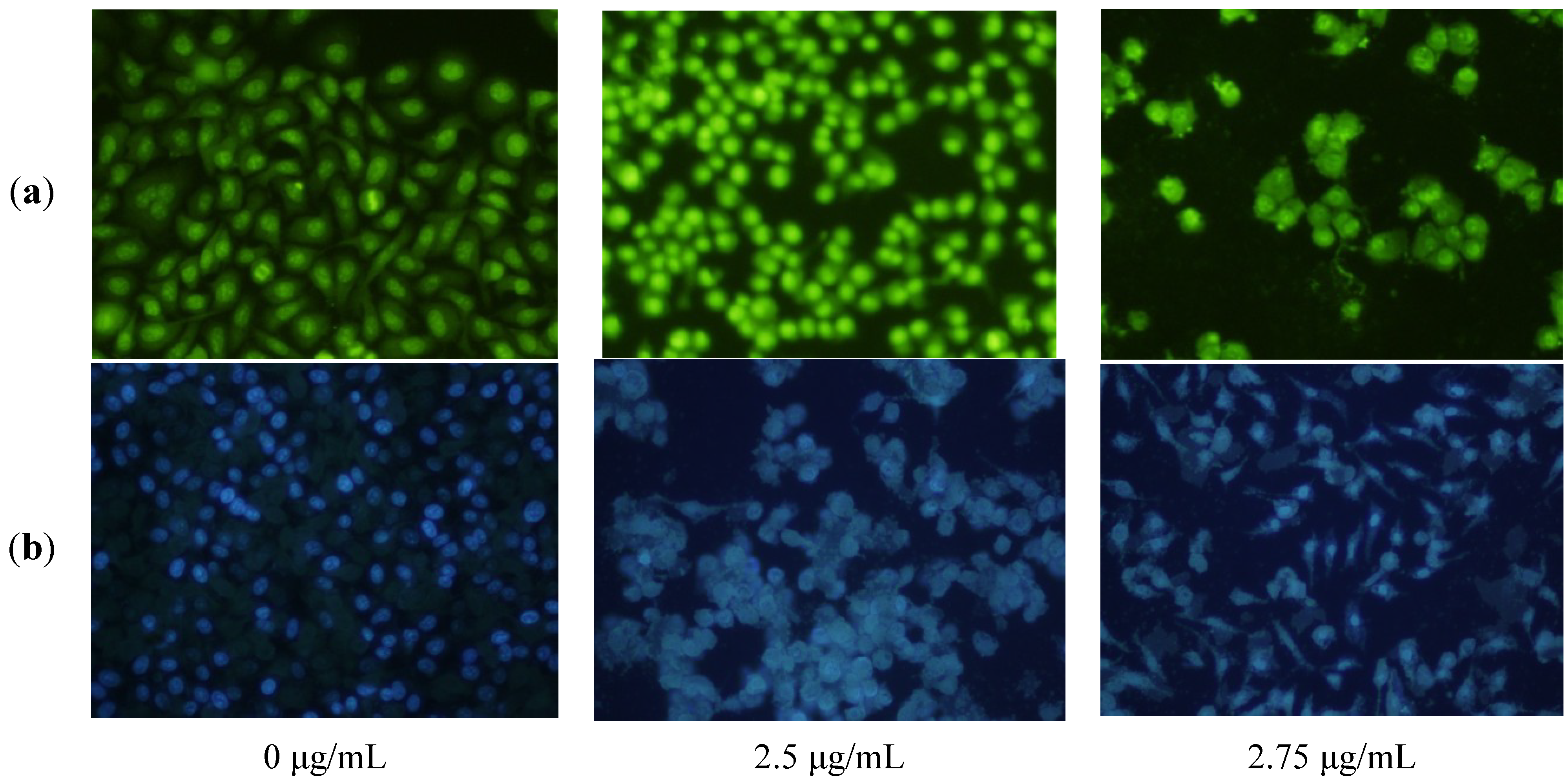
2.3. Apoptosis-Inducing Effect of 1 on HepG2 Cells

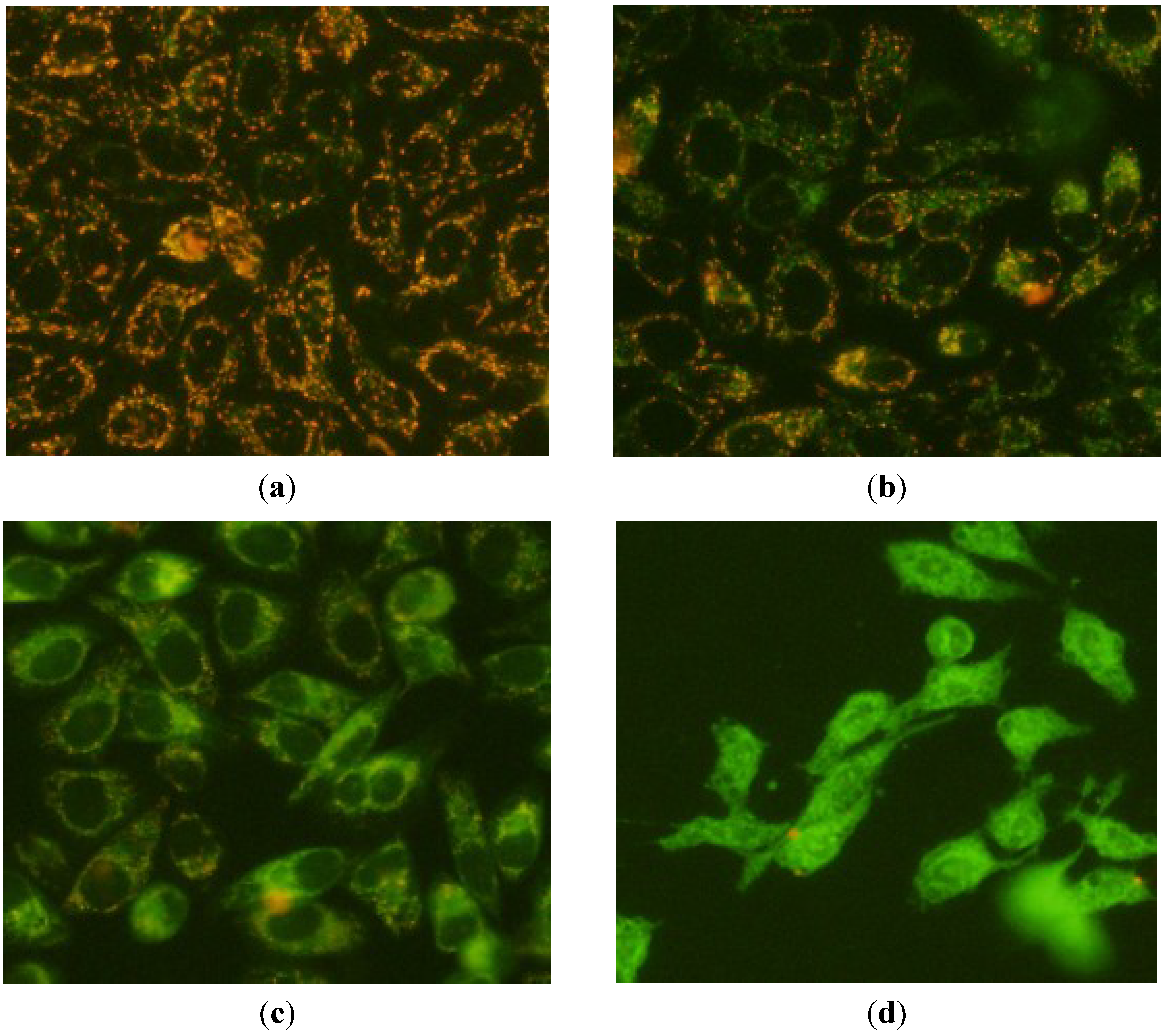
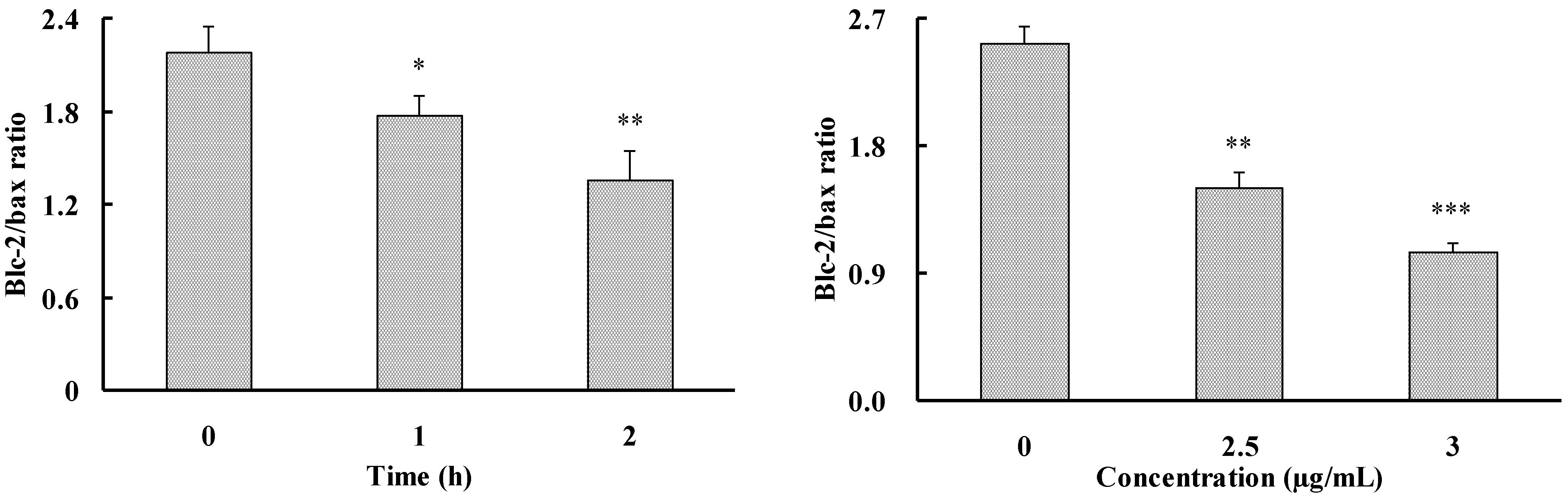
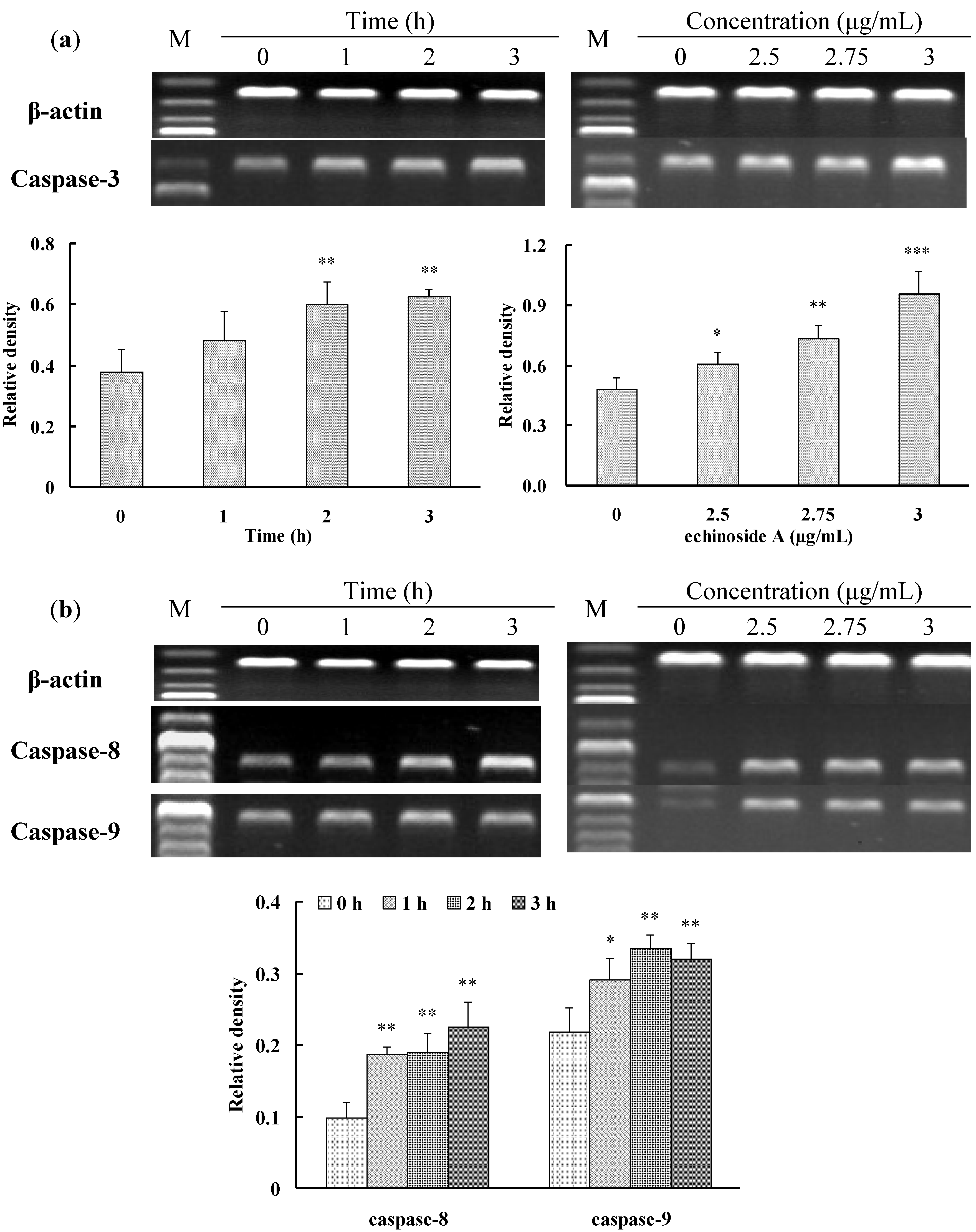
3. Experimental Section
3.1. General Details
3.2. Cell Lines
3.3. Cell Viability Assay
3.4. Fluorescence Microscope Observation
3.5. Measurement of the Mitochondrial Transmembrane Potentials (Δψm)
3.6. RT-PCR Analysis
| Gene | Primer Sequence | Product Size (bp) | Annealing Temperature (°C) | Cycle |
|---|---|---|---|---|
| β-Actin | 5′-CTGTCTGGCGGCACCACCAT-3′ 5′-GCAACTAAGTCATAGTCCGC-3′ | 254 | 54 | 19 |
| Bcl-2 | 5′-AGATGTCCAGCCAGCTGCACCTGAC-3′ 5′-AGATAGGCACCCAGGGTGATGCAAGCT-3′ | 367 | 57 | 33 |
| Bax | 5′-AAGCTGAGCGAGTGTCTCAAGCGC-3′ 5′-TCCCGCCACAAAGATGGTCACG-3′ | 366 | 66 | 26 |
| Caspase-3 | 5′-TTTGTTTGTGTGCTTCTGAGCC-3′ 5′-ATTCTGTTGCCACCTTTCGG-3′ | 400 | 54 | 27 |
| Caspase-8 | 5′-GGGACAGGAATGGAACACACTTGG-3′ 5′-TCAGGATGGTGAGAATATCATCGCC-3′ | 558 | 66 | 26 |
| Caspase-9 | 5′-AACAGGCAAGCAGCAAAGTT-3′ 5′-TCCATCTGTGCCGTAGACAG-3′ | 511 | 54 | 25 |
3.7. Statistical Analysis
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Perez, E.A. Impact, mechanisms, and novel chemotherapy strategies for overcoming resistance to anthracyclines and taxanes inmetastatic breast cancer. Breast Cancer Res. Treat. 2009, 114, 195–201. [Google Scholar] [CrossRef]
- Elmore, S. Apoptosis: A review of programmed cell death. Toxicol. Pathol. 2007, 35, 495–516. [Google Scholar] [CrossRef]
- Haefner, B. Drugs from the deep: Marine natural products as drug candidates. Drug Discov. Today 2003, 8, 536–544. [Google Scholar] [CrossRef]
- Newman, D.; Cragg, G. Marine natural products and related compounds in clinical and advanced preclinical trials. J. Nat. Prod. 2004, 67, 1216–1238. [Google Scholar] [CrossRef]
- Villa, F.A.; Gerwick, L. Marine natural product drug discovery: Leads for treatment of inflammation, cancer, infections, and neurological disorders. Immunopharmacol. Immunotoxicol. 2010, 32, 228–237. [Google Scholar] [CrossRef]
- Kalinin, V.I.; Silchenko, A.S.; Avilov, S.A.; Stonik, V.A.; Smirnov, A.V. Sea cucumbers triterpene glycosides, the recent progress in structural elucidation and chemotaxonomy. Phytochem. Rev. 2005, 4, 221–236. [Google Scholar] [CrossRef]
- Cui, F.X.; Xue, C.H.; Li, Z.J.; Zhang, Y.Q.; Dong, P.; Fu, X.Y.; Gao, X. Characterization and subunit composition of collagen from the body wall of sea cucumber Stichopus japonicus. Food Chem. 2007, 100, 1120–1125. [Google Scholar] [CrossRef]
- Zhong, Y.; Khan, A.; Shahidi, F. Compositional characteristics and antioxidant properties of fresh and processed sea cucumber (Cucumaria frondosa). J. Agric. Food Chem. 2007, 55, 1188–1192. [Google Scholar] [CrossRef]
- Kim, S.K.; Himaya, S.W. Triterpene glycosides from sea cucumbers and their biological activities. Adv. Food Nutr. Res. 2012, 65, 297–319. [Google Scholar] [CrossRef]
- Sun, P.; Yi, Y.H.; Li, L.; Tang, H.F. The resources, chemical structures and structural characteristics of triterpene glycosides from sea cucumbers (order Aspidochirotid). Chin. J. Nat. Med. 2007, 5, 463–469. [Google Scholar]
- Stonik, V.A.; Kalinin, V.I.; Avilov, S.A. Toxins from sea cucumbers (holothuroids): Chemical structures, properties, taxonomic distribution, biosynthesis and evolution. J. Nat. Toxins 1999, 8, 235–248. [Google Scholar]
- Antonov, A.S.; Avilov, S.A.; Kalinovsky, A.I.; Anastyuk, S.D.; Dmitrenok, P.S.; Evtushenko, E.V.; Kalinin, V.I.; Smirnov, A.V.; Taboada, S.; Ballesteros, M.; et al. Triterpene glycosides from Antarctic sea cucumbers I. Structure of liouvillosides A1, A2, A3, B1 and B2 from the sea cucumber Staurocucumis liouvillei, new procedure for separation of highly polar glycoside fractions and taxonomic revision. J. Nat. Prod. 2008, 71, 1677–1685. [Google Scholar] [CrossRef]
- Maier, M.S.; Roccatagliata, A.J.; Kuriss, A.; Chludil, H.; Seldes, A.M.; Pujol, C.A.; Damonte, E.B. Two new cytotoxic and virucidal trisulfated triterpene glycosides from the Antarctic sea cucumber Staurocucumis liouvillei. J. Nat. Prod. 2001, 64, 732–736. [Google Scholar] [CrossRef]
- Chludil, H.D.; Muniain, C.C.; Seldes, A.M.; Maier, M.S. Cytotoxic and antifungal triterpene glycosides from the Patagonian sea cucumber Hemoiedema spectabilis. J. Nat. Prod. 2002, 65, 860–865. [Google Scholar] [CrossRef]
- Chludil, H.D.; Murray, A.P.; Seldes, A.M. Biologically active triterpene glycosides from sea cucumbers (Holothuroidea, Echinodermata). Stud. Nat. Prod. Chem. 2003, 28, 587–615. [Google Scholar] [CrossRef]
- Dang, N.H.; Thanh, N.V.; Kiem, P.V.; Huong, L.M.; Minh, C.V.; Kim, Y.H. Two new triterpene glycosides from the Vietnamese sea cucumber Holothuria scabra. Arch. Pharm. Res. 2007, 30, 1387–1391. [Google Scholar] [CrossRef]
- Zhang, S.; Tang, H.; Yi, Y. Cytotoxic triterpene glycosides from the sea cucumber Pseudocolochirus violaceus. Fitoterapia 2007, 78, 283–287. [Google Scholar] [CrossRef]
- Han, H.; Yi, Y.H.; Liu, B.S.; Wang, X.H.; Pan, M.X. Leucospilotaside C, a new sulfated triterpene glycoside from sea cucumber Holothuria leucospilota. Chin. Chem. Lett. 2008, 19, 1462–1464. [Google Scholar] [CrossRef]
- Kalinin, V.I.; Aminin, D.L.; Avilov, S.A.; Silchenko, A.S.; Stonik, V.A. Triterpene glycosides from sea cucucmbers (Holothurioidea, Echinodermata), biological activities and functions. In Studies in Natural Product Chemistry (Bioactive Natural Products); Rahman, A.-u., Ed.; Elsevier Science Publisher: Amsterdam, The Netherlands, 2008; pp. 135–196. [Google Scholar]
- Aminin, D.L.; Chaykina, E.L.; Agafonova, I.G.; Avilov, S.A.; Kalinin, V.I.; Stonik, V.A. Antitumor activity of the immunomodulatory lead Cumaside. Int. Immunopharmacol. 2010, 10, 648–654. [Google Scholar] [CrossRef]
- Aminin, D.L.; Silchenko, A.S.; Avilov, S.A.; Stepanov, V.G.; Kalinin, V.I. Immunomodulatory action of monosulfated triterpene glycosides from the sea cucumber Cucumaria okhotensis: Stimulation of activity of mouse peritoneal macrophages. Nat. Prod. Commun. 2010, 5, 1877–1880. [Google Scholar]
- Yan, B.; Li, L.; Yi, Y.H.; Tang, H.F.; Sun, P.; Zhang, S.Y. Study on bioactive triterpene glycosides of sea cucumber Holothuria scabra Jaeger. Acad. J. Second Mil. Med. Univ. 2005, 26, 626–631. (in Chinese). [Google Scholar]
- Han, H.; Li, L.; Yi, Y.H.; Wang, X.H.; Pan, X.M. Triterpene glycosides from sea cucumber Holothuria scabra with cytotoxic activity. Chin. Herb. Med. 2012, 4, 183–188. [Google Scholar]
- Han, H. Biologically Active Triterpene Glycosides from Sea Cucucmbers Holothuria leucospilota Brandt and Holothuria scabra Jaeger. Ph.D. Thesis, Second Military Medical University, Shanghai, China, 2007. [Google Scholar]
- Han, H.; Yi, Y.H.; Xu, Q.Z.; La, M.P.; Zhang, H.W. Two new cytotoxic triterpene glycosides from the sea cucumber Holothuria scabra. Planta Med. 2009, 75, 1608–1612. [Google Scholar] [CrossRef]
- La, M.P.; Yi, Y.H.; Li, L.; Lu, B.D.; Han, H.; Wand, Z.L.; Gong, W. Three tritserpene glycosides from sea cucumber Cucumaria frondosa Gunnerus. Chin. J. Nat. Med. 2008, 6, 254–258. [Google Scholar] [CrossRef]
- Zou, Z.R.; Yi, Y.H.; Wu, H.M.; Wu, J.H.; Liaw, C.C.; Lee, K.H. Intecedensides A–C, three new cytotoxic triterpene glycosides from the sea cucumber Mensamaria intercedens Lampert. J. Nat. Prod. 2003, 66, 1055–1060. [Google Scholar] [CrossRef]
- Zou, Z.R.; Yi, Y.H.; Wu, H.M.; Yao, X.S.; Du, L.J.; Wu, J.H.; Liaw, C.C.; Lee, K.H. Intercedensides D–I, cytotoxic triterpene glycosides from the sea cucumber Mensamaria intercedens Lampert. J. Nat. Prod. 2005, 68, 540–546. [Google Scholar] [CrossRef]
- Kalinin, V.I.; Prokofieva, N.G.; Likhatskaya, G.N.; Schentsova, E.B.; Agafonova, I.G.; Avilov, S.A.; Drozdova, O.A. Hemolytic activities of triterpene glycosides from the holothurian order Dendrochirotida: Some trends in the evolution of this group of toxins. Toxicon 1996, 34, 475–483. [Google Scholar] [CrossRef]
- Avilov, S.A.; Silchenko, A.S.; Antonov, A.S.; Kalinin, V.I.; Kalinovsky, A.I.; Smirnov, A.V.; Dmitrenok, P.S.; Evtushenko, E.V.; Fedorov, S.N.; Savina, A.S.; et al. Synaptosides A and A1, triterpene glycosides from the sea cucumber Synapta maculata containing 3-O-methylglucuronic acid and their cytotoxic activity against tumor cells. J. Nat. Prod. 2008, 71, 525–531. [Google Scholar] [CrossRef]
- Jin, J.O.; Shastina, V.V.; Shin, S.W.; Xu, Q.; Park, J.I.; Rasskazov, V.A.; Avilov, S.A.; Fedorov, S.N.; Stonik, V.A.; Kwak, J.Y. Differential effects of triterpene glycosides, frondoside A and cucumarioside A2-2 isolated from sea cucumbers on caspase activation and apoptosis of human leukemia cells. FEBS Lett. 2009, 583, 697–702. [Google Scholar] [CrossRef]
- Silchenko, A.S.; Avilov, S.A.; Kalinin, V.I.; Kalinovsky, A.I.; Dmitrenok, P.S.; Fedorov, S.N.; Stepanov, V.G.; Dong, Z.; Stonik, V.A. Constituents of the sea cucumber Cucumaria okhotensis. Structures of okhotosides B1–B3 and cytotoxic activities of some glycosides from this species. J. Nat. Prod. 2008, 71, 351–356. [Google Scholar] [CrossRef]
- Kalinin, V.I. System-theoretical (holistic) approach to the modeling of structural functional relationships of biomolecules and their evolution: An example of triterpene glycosides from sea cucumbers (Echinodermata, Holothurioidea). J. Theor. Biol. 2000, 206, 151–168. [Google Scholar] [CrossRef]
- Zhao, Q.; Xue, Y.; Wang, J.F.; Li, H.; Long, T.T.; Li, Z.; Wang, Y.M.; Dong, P.; Xue, C.H. In vitro and in vivo anti-tumour activities of echinoside A and ds-echinoside A from Pearsonothuria graeffei. J. Sci. Food Agric. 2012, 92, 965–974. [Google Scholar] [CrossRef]
- Zhao, Q.; Xue, Y.; Liu, Z.D.; Li, H.; Wang, J.F.; Li, Z.J.; Wang, Y.M.; Dong, P.; Xue, C.H. Differential effects of sulfated triterpene glycosides, holothurin A1, and 24-dehydroechinoside A, on antimetastatic activity via regulation of the MMP-9 signal pathway. J. Food Sci. 2010, 75, H280–H288. [Google Scholar] [CrossRef]
- Li, M.; Miao, Z.H.; Chen, Z.; Chen, Q.; Gui, M.; Lin, L.P.; Sun, P.; Yi, Y.H.; Ding, J. Echinoside A, a new marine-derived anticancer saponin, targets topoisomerase2α by unique interference with its DNA binding and catalytic cycle. Ann. Oncol. 2010, 21, 597–607. [Google Scholar] [CrossRef]
- Ziedan, N.I.; Kadri, H.; Westwekll, A.D. The development of pro-apoptotic cancer therapeutics. Mini-Rev. Med. Chem. 2008, 8, 711–718. [Google Scholar] [CrossRef]
- San-Miguel, B.; Alvarez, M.; Culebras, J.M.; Gonzalez-Gallego, J.; Tunon, M.J. N-Acetyl-cysteine protects liver from apoptotic death in an animal model of fulminant hepatic failure. Apoptosis 2006, 11, 1945–1957. [Google Scholar] [CrossRef]
- Delhalle, S.; Duvoix, A.; Schnekenburger, M.; Morceau, F.; Dicato, M.; Diederich, M. An introduction to the molecular mechanisms of apoptosis. Ann. N. Y. Acad. Sci. 2003, 1010, 1–8. [Google Scholar] [CrossRef]
- Li, X.; Roginsky, A.B.; Ding, X.Z.; Woodward, C.; Collin, P.; Newman, R.A.; Bell, R.H., Jr.; Adrian, T.E. Review of the apoptosis pathways in pancreatic cancer and the anti-apoptotic effects of the novel sea cucumber compound, Frondoside A. Ann. N. Y. Acad. Sci. 2008, 1138, 181–198. [Google Scholar] [CrossRef]
- Sun, H.X.; Zheng, Q.F.; Tu, J. Induction of apoptosis in HeLa cells by 3β-hydroxy-12-oleanen-27-oic acid from the rhizomes of Astilbe chinensis. Bioorgan. Med. Chem. 2006, 4, 1189–1198. [Google Scholar]
- Huang, X.H.; Xie, H.T.; Sun, R.Y. Introduction and application of new edition of software of the new drug statistical treatment (NDST-21). Chin. J. Clin. Pharmacol. Ther. 2002, 7, 169–174. (in Chinese). [Google Scholar]
- Wang, J.J.; Chen, X.F.; Zhou, Z.H.; Li, J.H.; Sun, H.X. The inhibitory effect of 3β-hydroxy-12-oleanen-27-oic acid on growth and motility of human hepatoma hepG2 cells through JNK and Akt signaling pathway. Evid. Based Complement. Altern. Med. 2013, 2013, 1–17. [Google Scholar]
- Zhang, Y.B.; Ye, Y.P.; Wu, X.D.; Sun, H.X. Astilbotriterpenic acid induces growth arrest and apoptosis in HeLa cells through mitochondria-related pathways and reactive oxygen species (ROS) production. Chem. Biodivers. 2009, 6, 218–230. [Google Scholar] [CrossRef]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Wang, J.; Han, H.; Chen, X.; Yi, Y.; Sun, H. Cytotoxic and Apoptosis-Inducing Activity of Triterpene Glycosides from Holothuria scabra and Cucumaria frondosa against HepG2 Cells. Mar. Drugs 2014, 12, 4274-4290. https://doi.org/10.3390/md12084274
Wang J, Han H, Chen X, Yi Y, Sun H. Cytotoxic and Apoptosis-Inducing Activity of Triterpene Glycosides from Holothuria scabra and Cucumaria frondosa against HepG2 Cells. Marine Drugs. 2014; 12(8):4274-4290. https://doi.org/10.3390/md12084274
Chicago/Turabian StyleWang, Juanjuan, Hua Han, Xiangfeng Chen, Yanghua Yi, and Hongxiang Sun. 2014. "Cytotoxic and Apoptosis-Inducing Activity of Triterpene Glycosides from Holothuria scabra and Cucumaria frondosa against HepG2 Cells" Marine Drugs 12, no. 8: 4274-4290. https://doi.org/10.3390/md12084274