Cephalopods as Vectors of Harmful Algal Bloom Toxins in Marine Food Webs



Abstract
:1. Introduction
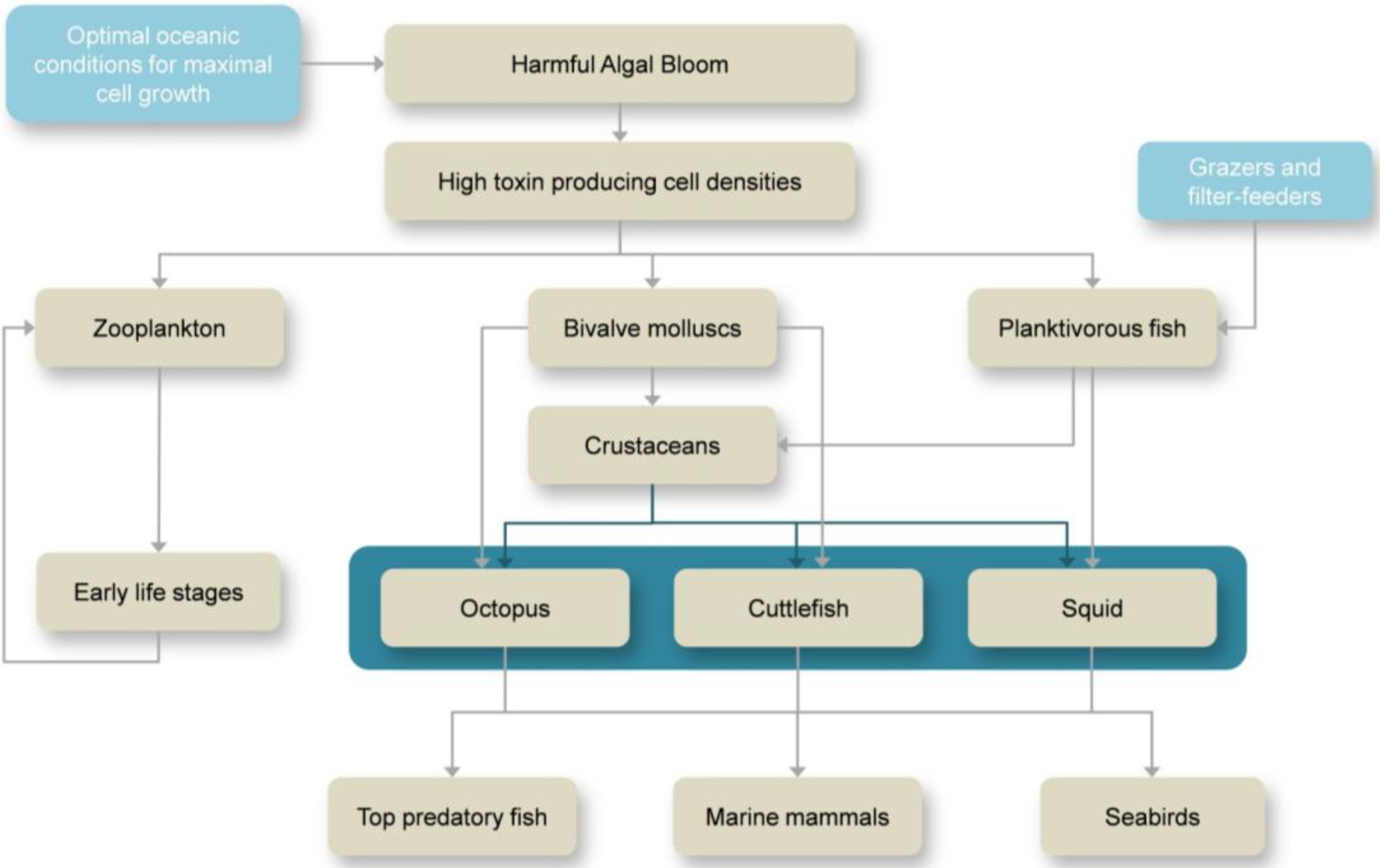
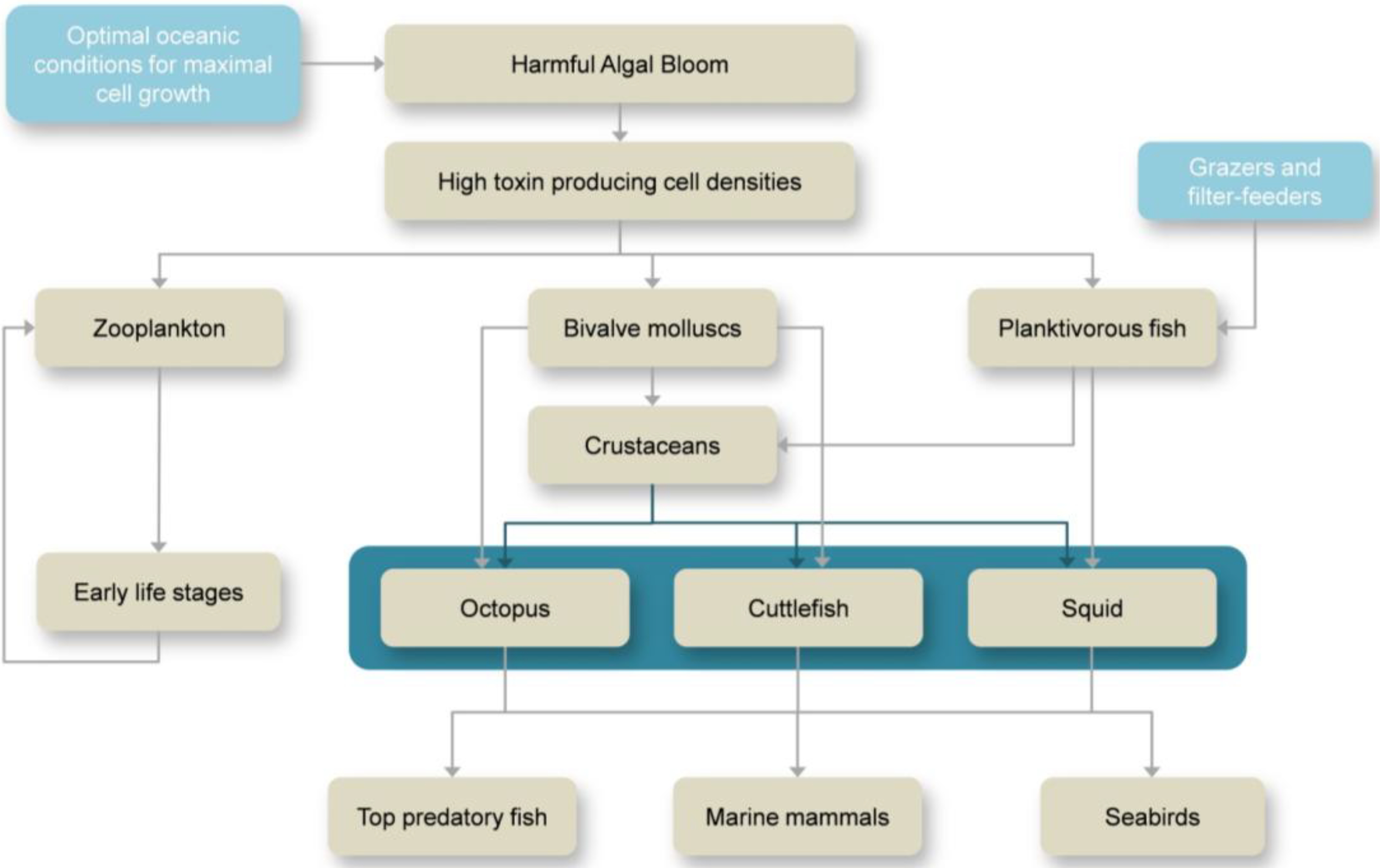
2. Overview on HABs and Marine Toxins

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Toxin | Species | Maximum levels found | Tissue/Organ | Reference |
|---|---|---|---|---|
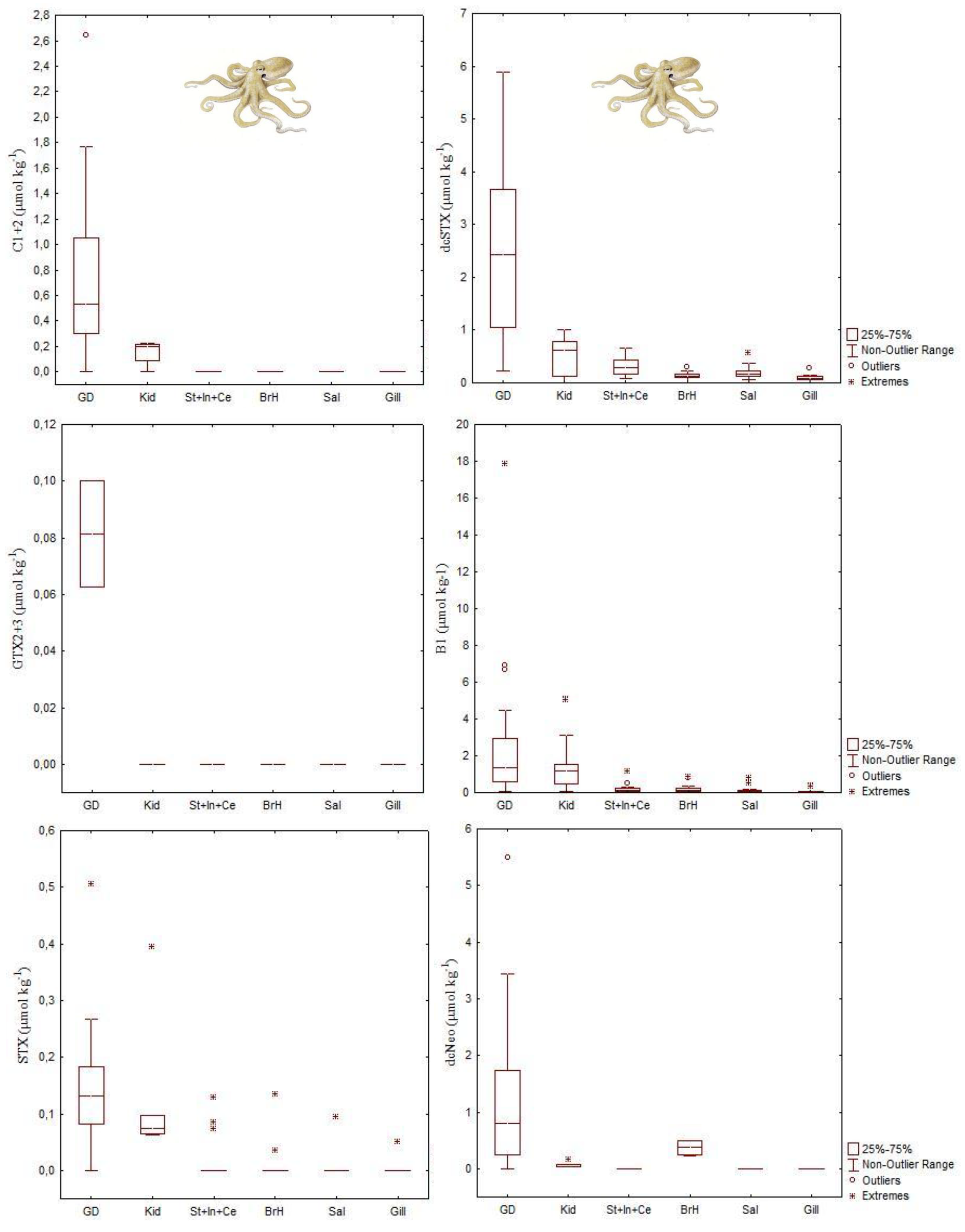
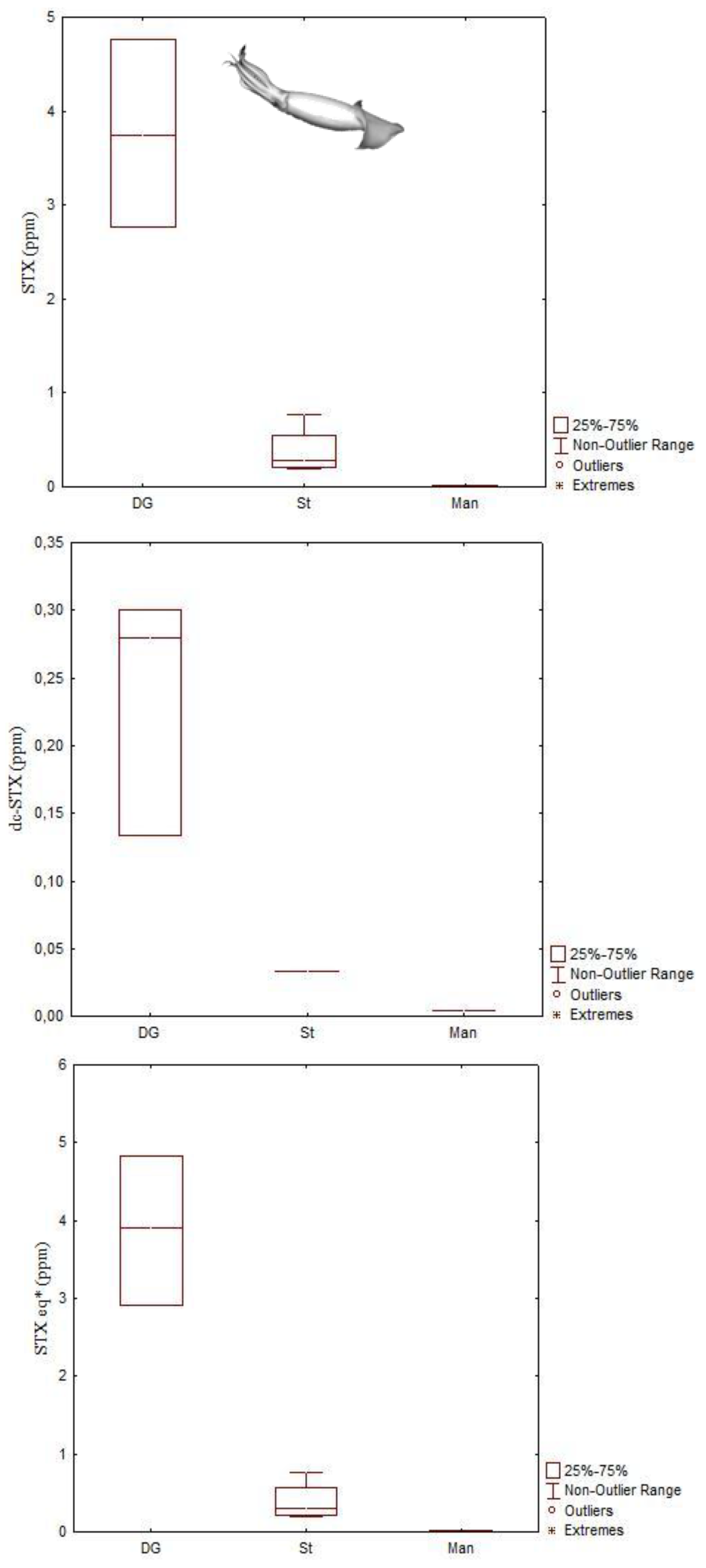
| DA | Octopus vulgaris | 166.2 μg DA g−1 | Digestive gland | [40] |
| Eledone moschata | 127 μg DA g−1 | Digestive gland | [41] | |
| Eledone cirrhosa | 18.8 μg DA g−1 | Digestive gland | [41] | |
| Sepia officinalis | 241.7 μg DA g−1 | Digestive gland | [42] | |
| Doryteuthis opalescens | 0.37 μg DA g−1 | Stomach | [43] | |
| Dosidicus gigas | 0.23 μg DA g−1 | Digestive gland | [44] | |
| STXs | Octopus vulgaris | 35 μg STX equivalents g−1 | Digestive gland | [45] |
| Dosidicus gigas | 4.83 μg STX equivalents g−1 | Digestive gland | [44] | |
| Octopus (Abdopus) sp. 5 | 2.46 μg STX equivalents g−1 | Arms | [46] | |
| PlTX | Octopod (species unknown) | 971 μg/kg OVTX-a and 115 μg/kg PlTX | Unspecified | [47] |
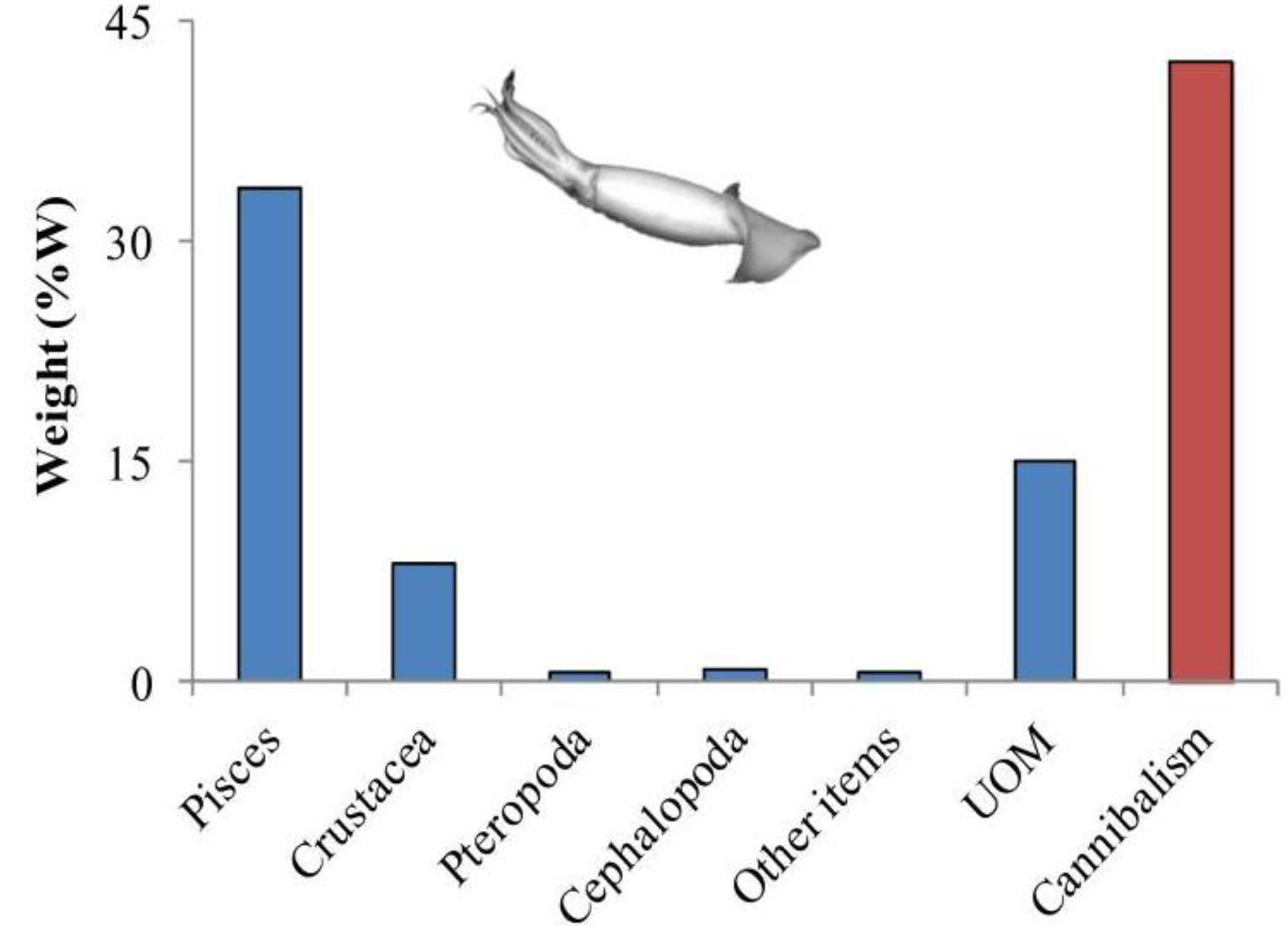
3. Cephalopod Life Strategies and Feeding Ecology
| Prey category | Occurrence index (OCI) | ||
|---|---|---|---|
| Viana | Cascais | Tavira | |
| ANNELIDA | 1.85 | 2.52 | - |
| POLYCHAETA unidentified | 1.85 | 2.52 | - |
| CRUSTACEA | 38.27 | 67.31 | 36.90 |
| ISOPODA unidentified | - | - | 0.18 |
| DECAPODA NATANTIA | 1.93 | 6.04 | 2.57 |
| Natantia unidentified | 1.93 | 3.43 | 2.57 |
| Caridae unidentified | - | 2.61 | - |
| DECAPODA REPTANTIA | 36.34 | 61.27 | 34.14 |
| ANOMURA | - | - | 0.33 |
| Paguridae unidentified | - | - | 0.26 |
| Pagurus prideauxi | - | - | 0.07 |
| BRACHYURA | 36.34 | 61.27 | 33.81 |
| Brachiura unidentified | 18.96 | 39.33 | 17.51 |
| Portunidae unidentified | 10.84 | 21.94 | 10.17 |
| Liocarcinus sp. | 5.17 | - | 5.03 |
| Polybius henzlow | 1.36 | - | 1.10 |
| MOLLUSCA | 7.13 | 14.89 | 23.52 |
| GASTROPODA unidentified | 1.49 | 1.87 | 0.61 |
| BIVALVIA | 1.75 | 7.43 | 16.56 |
| Carditidae unidentified | - | - | 1.07 |
| Cardiidae unidentified | - | - | 0.73 |
| Mytilus sp. | - | - | 1.72 |
| Venus sp. | - | - | 1.48 |
| Bivalvia unidentified | 1.75 | 7.43 | 11.56 |
| CEPHALOPODA | 3.90 | 5.59 | 6.35 |
| Sepiolidae unidentified | - | - | 1.39 |
| Sepia sp. | - | - | 0.73 |
| Illex coindetii | 0.71 | - | - |
| Octopus sp. | 0.18 | 0.83 | 0.36 |
| Cephalopoda unidentified | 3.01 | 4.75 | 3.87 |
| OSTEICHTHYA | 52.74 | 15.28 | 39.59 |
| Clupeidae unidentified | 16.70 | 3.29 | 12.94 |
| Gobidae unidentified | 5.80 | 4.53 | 1.43 |
| Osteichthya unidentified | 30.24 | 7.46 | 25.22 |


4. HAB-Toxins in Cephalopods
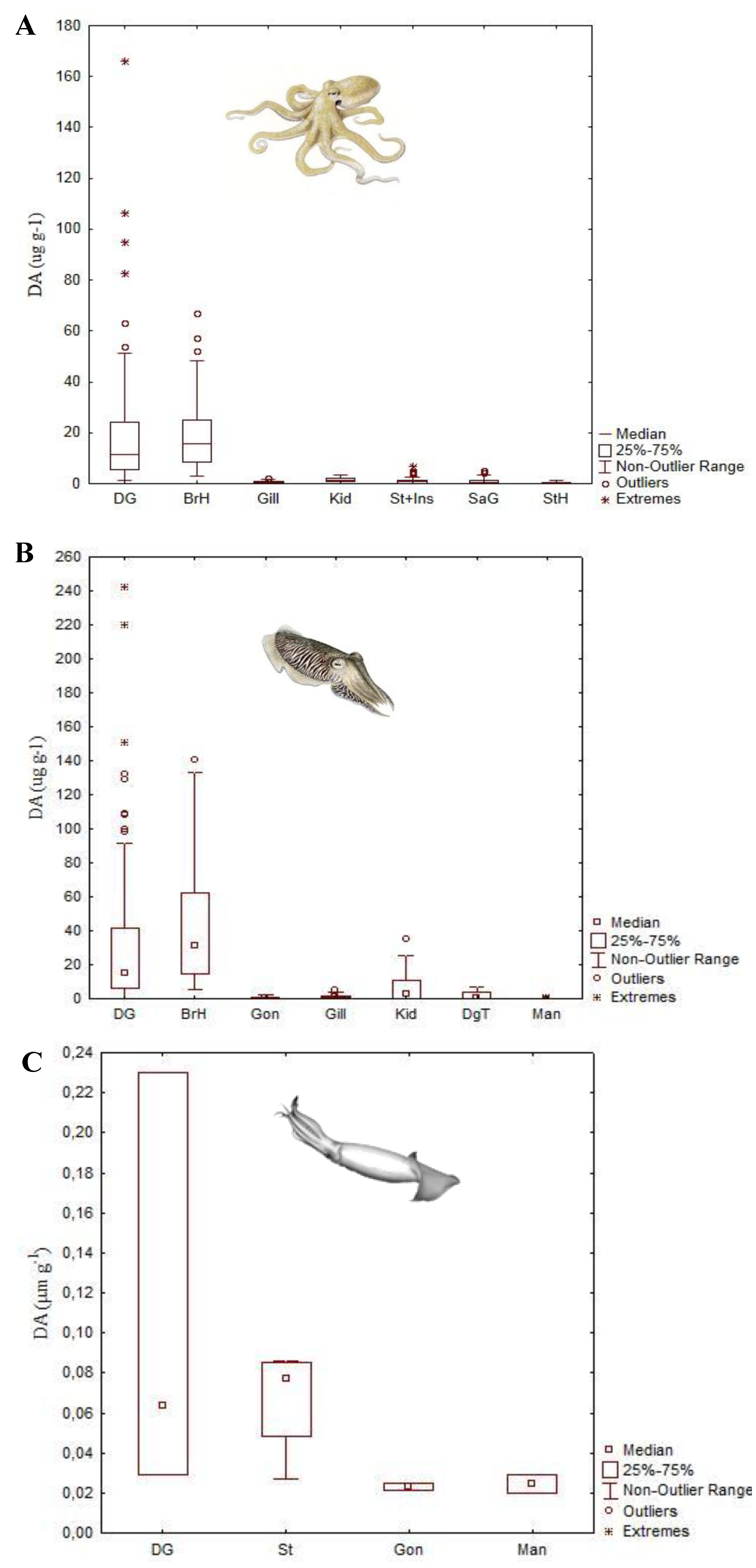
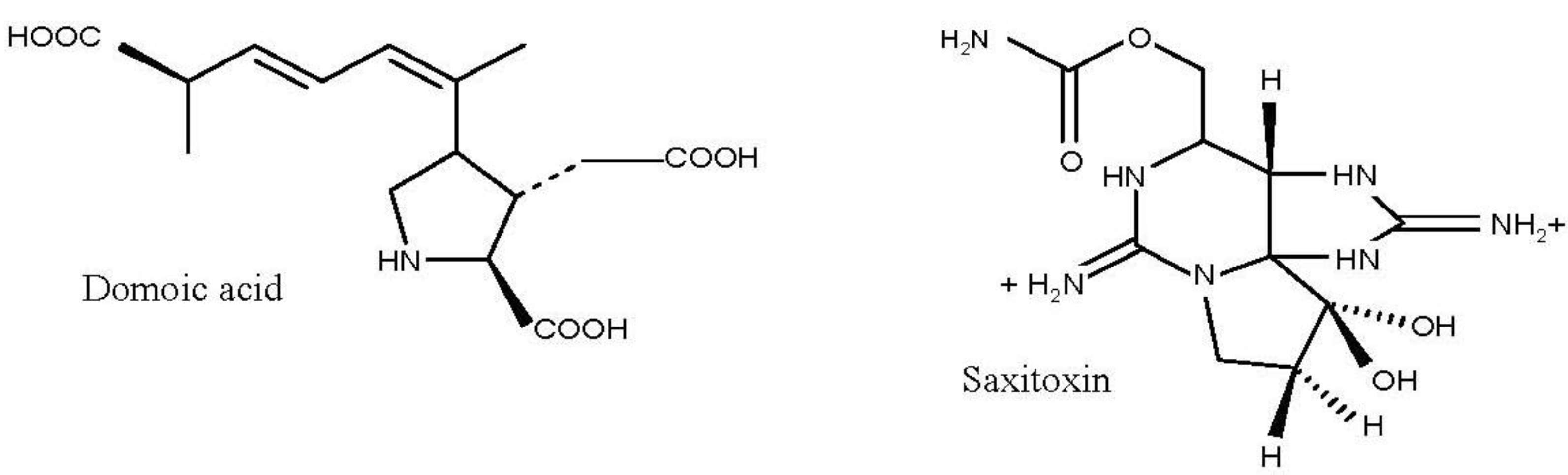
4.1. Amnesic Shellfish Toxins

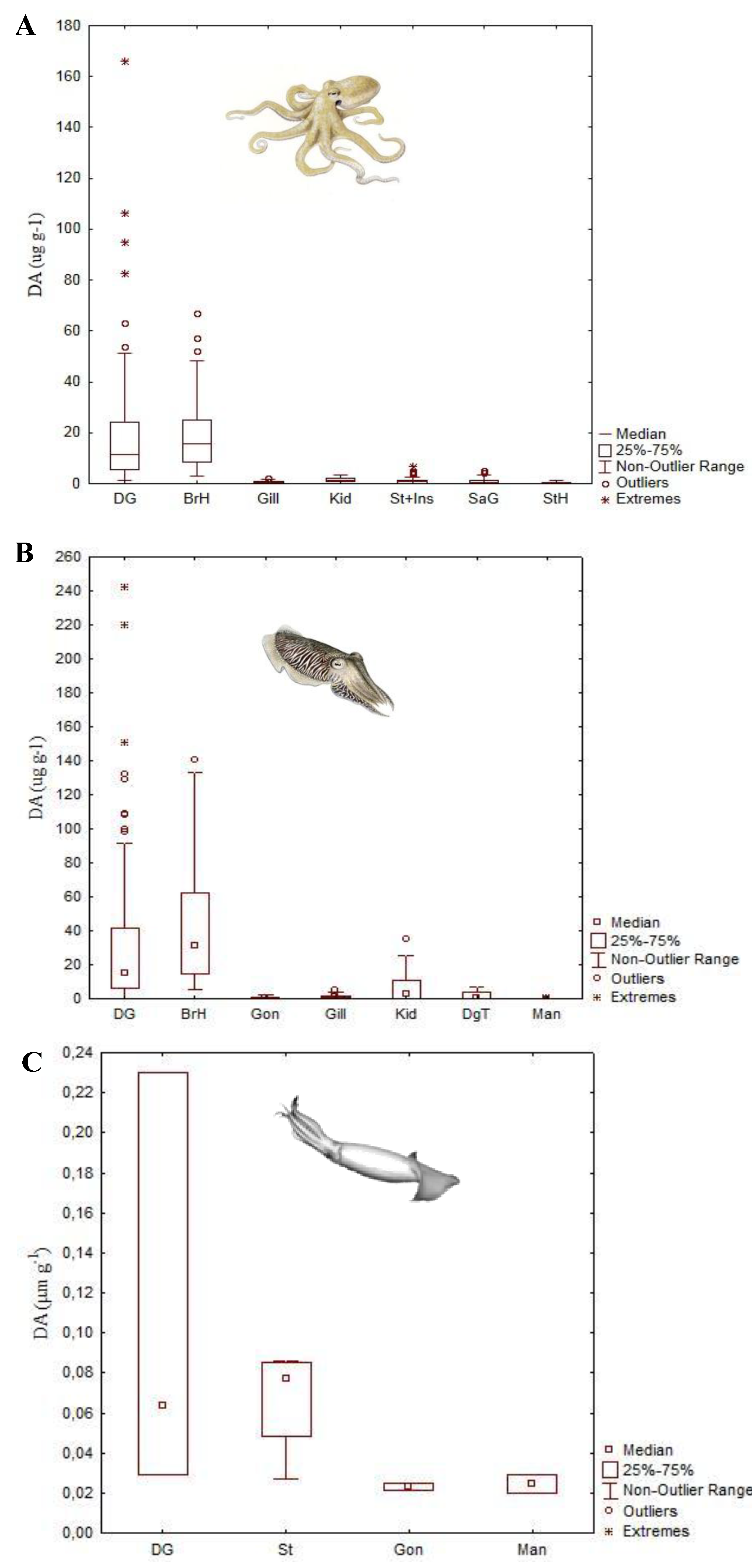
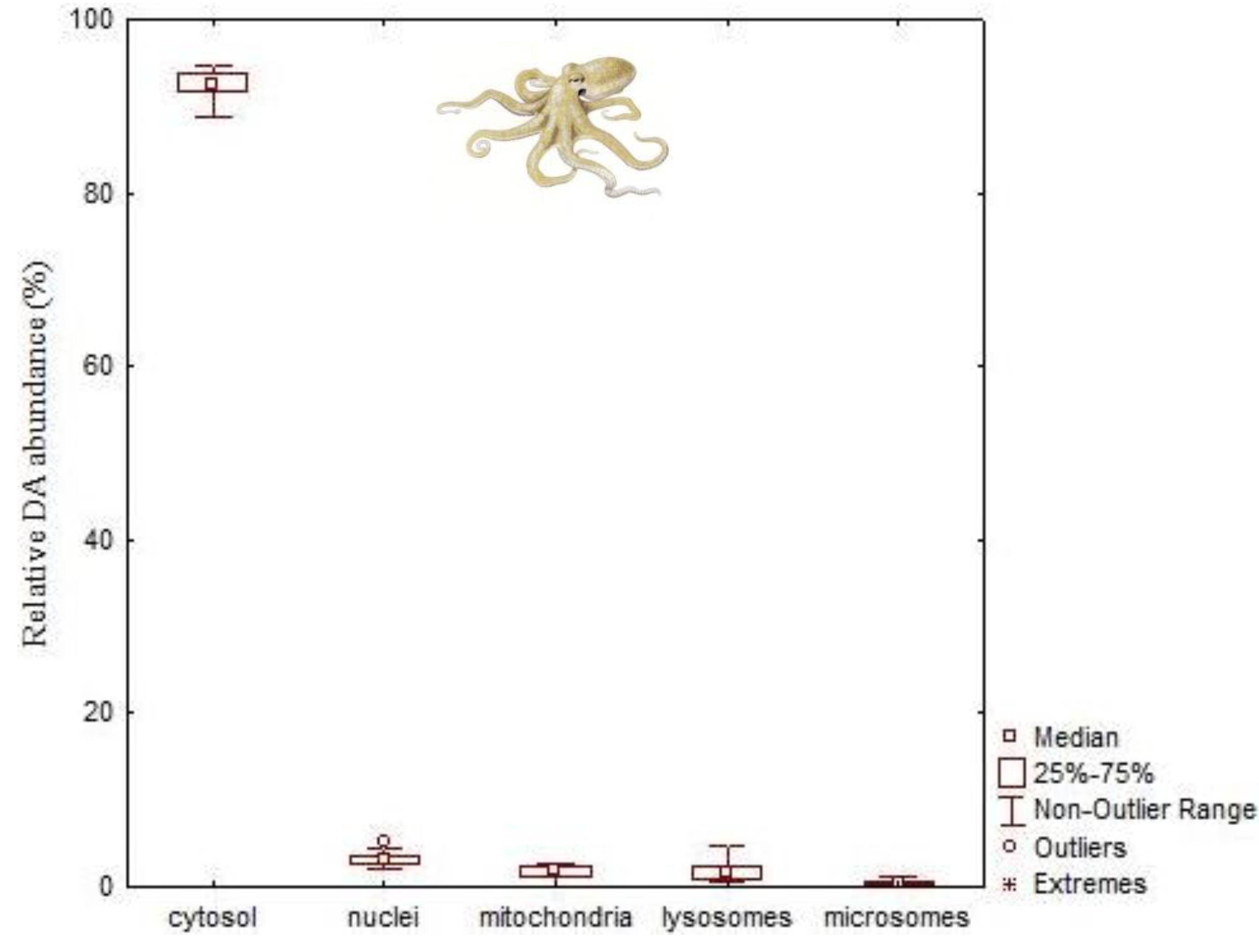
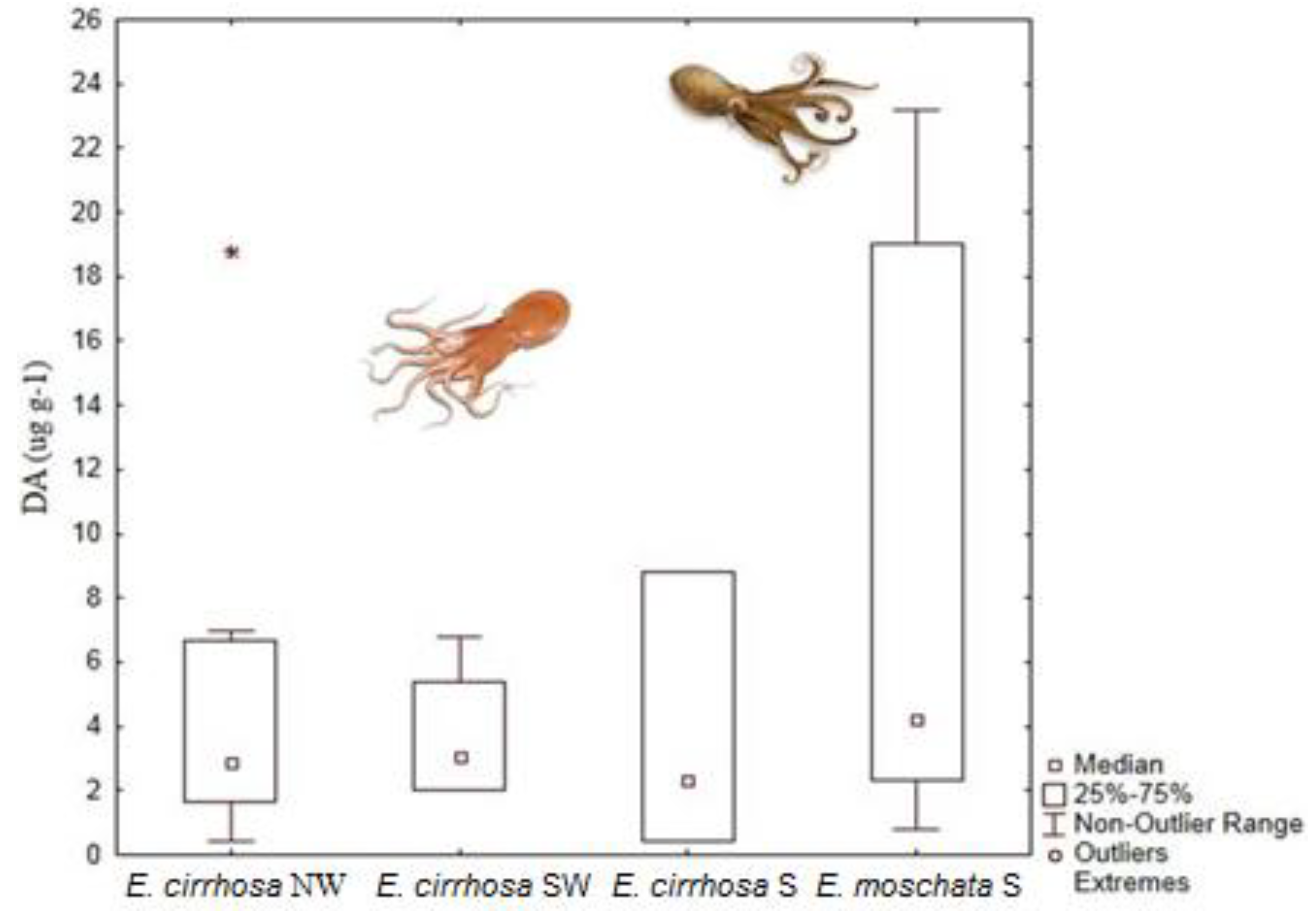

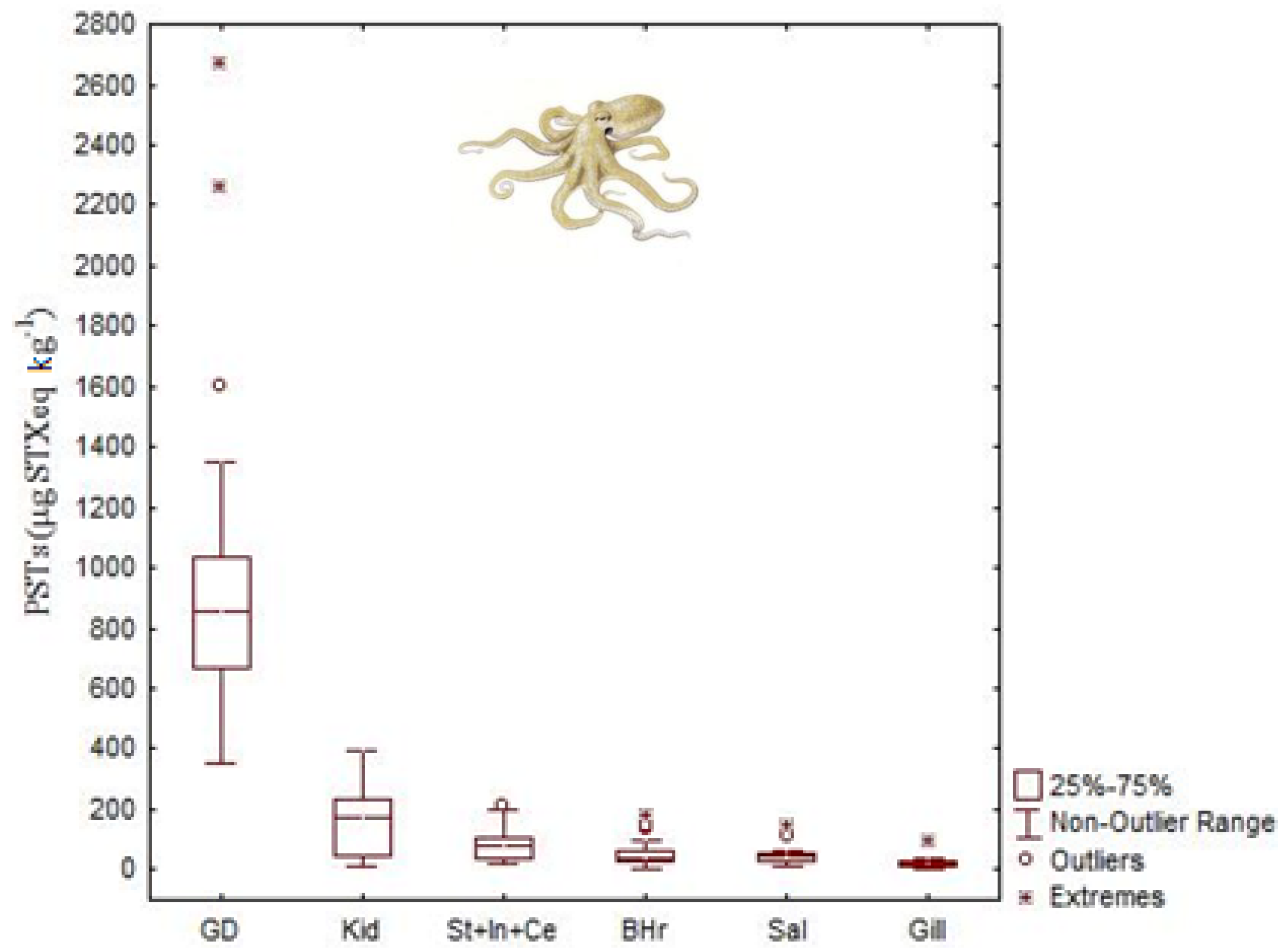
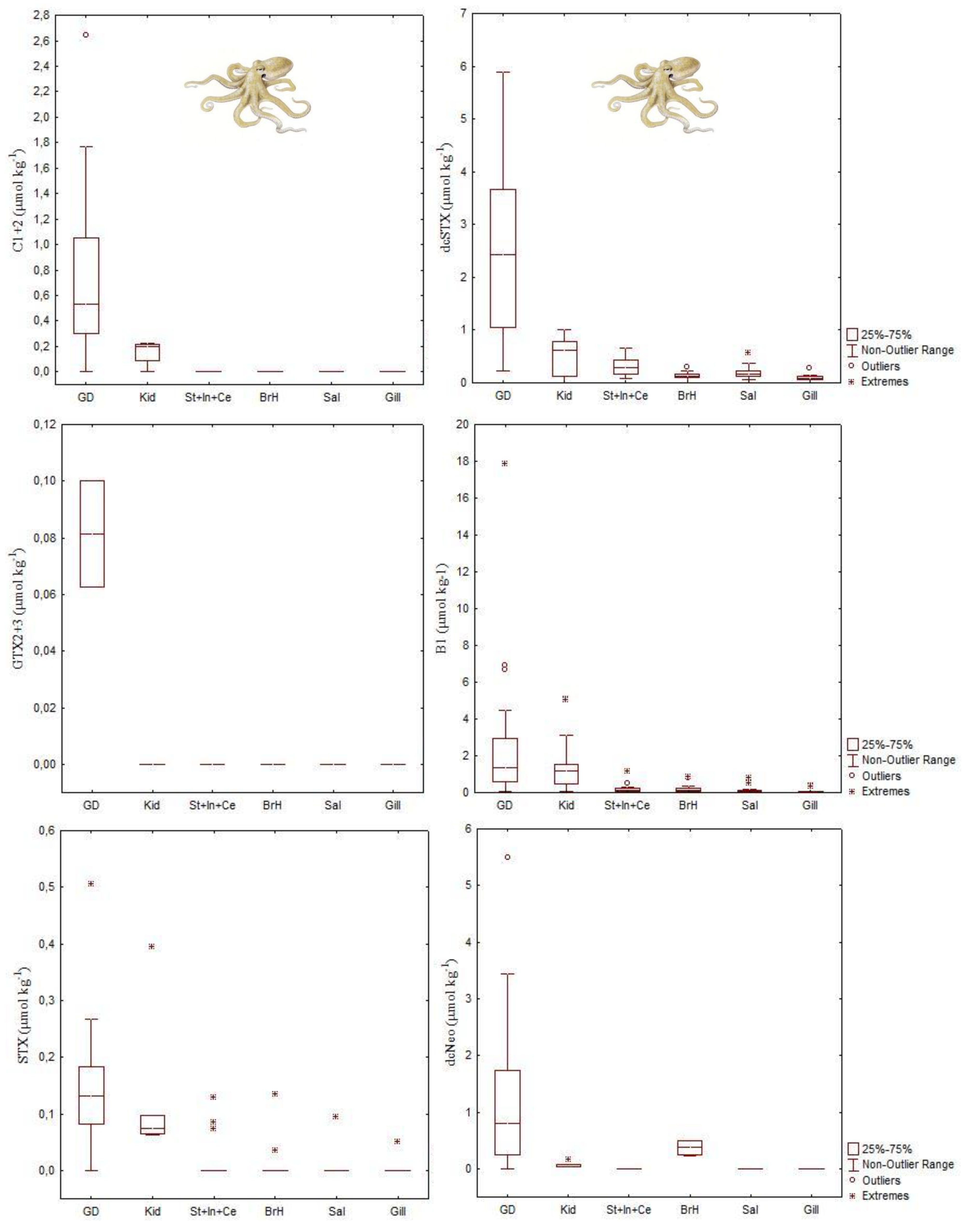
4.2. Paralytic Shellfish Toxins
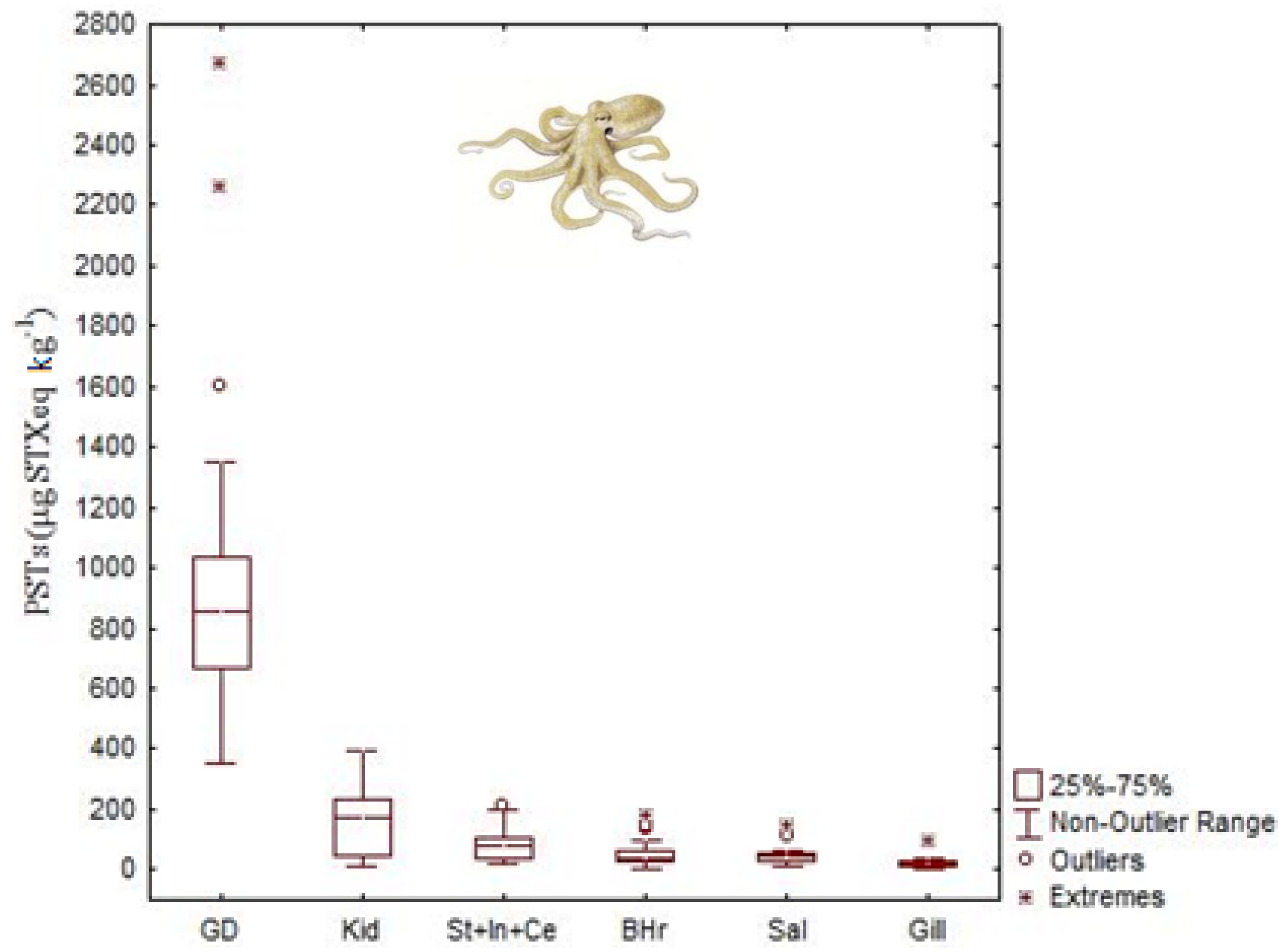

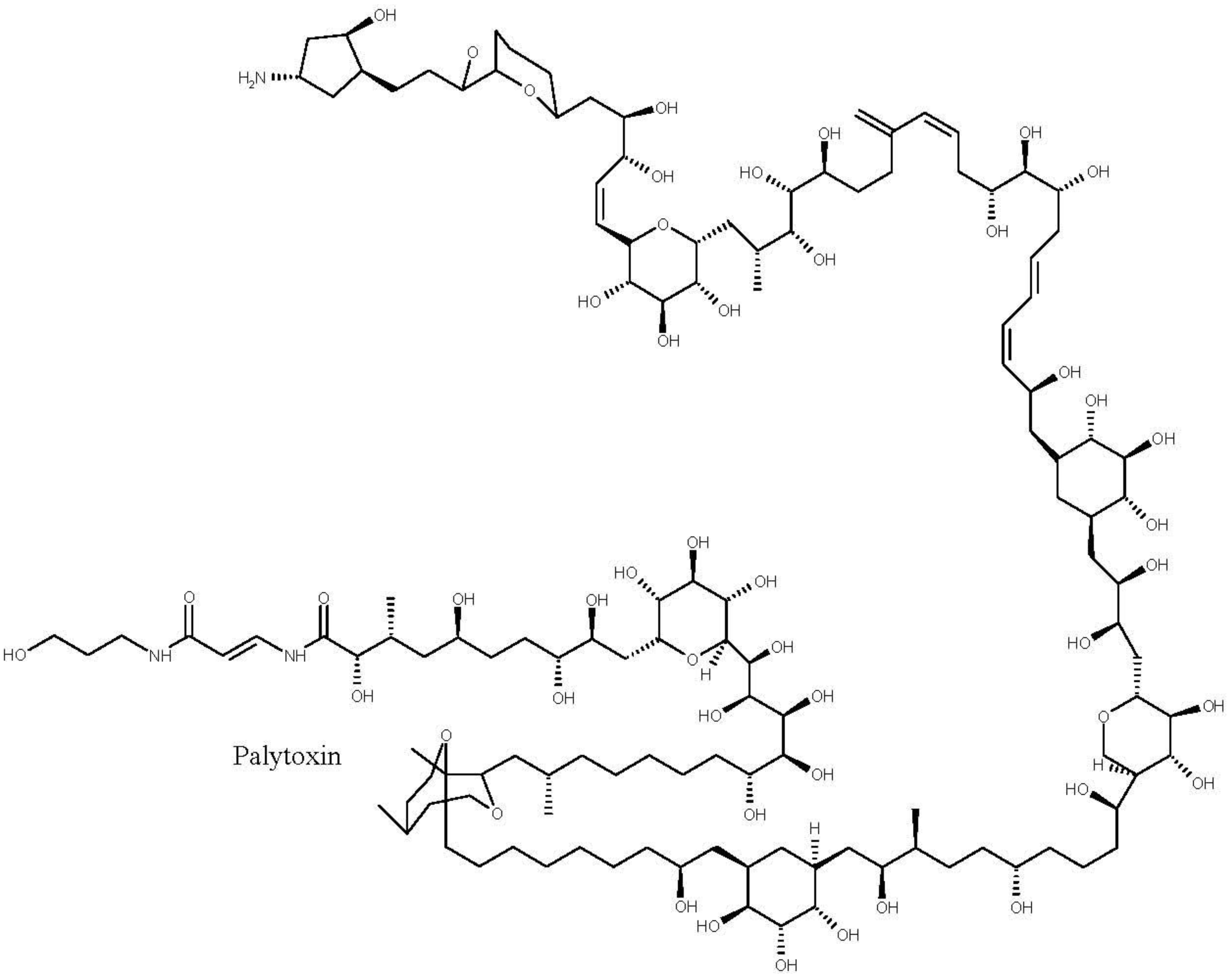
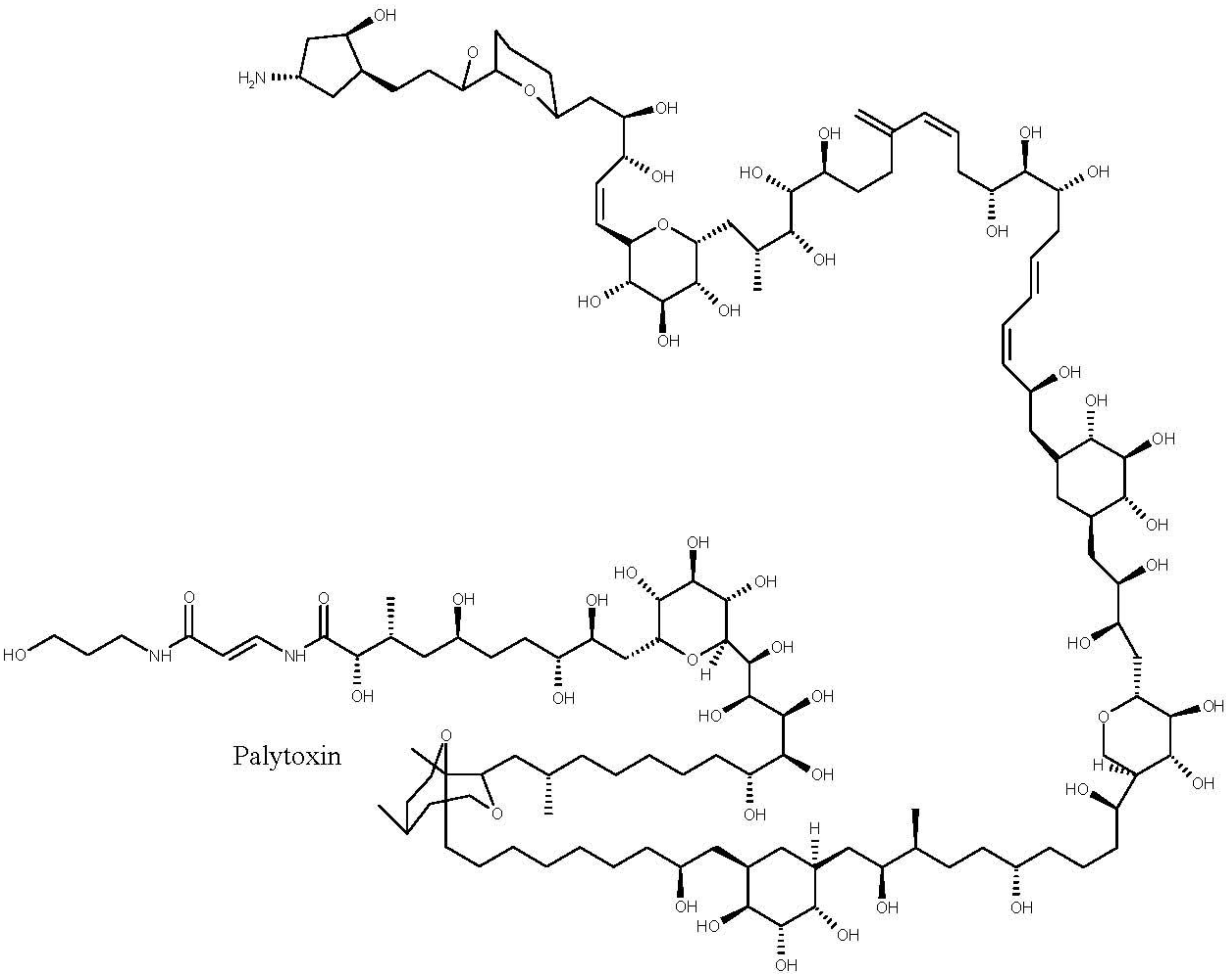
4.3. Palytoxins
5. Potential Effects of HAB-Toxins in Cephalopod Early Stages
6. Conclusions
Acknowledgments
Conflicts of Interest
References
- Smayda, T.J. What is a bloom? A commentary. Limnol. Oceanogr. 1997, 42, 1132–1136. [Google Scholar] [CrossRef]
- Landsberg, J.H. The effects of harmful algal blooms on aquatic organisms. Rev. Fish. Sci. 2002, 10, 113–390. [Google Scholar] [CrossRef]
- Smayda, T.J. Harmful algal blooms: Their ecophysiology and general relevance to phytoplankton blooms in the sea. Limnol. Oceanogr. 1997, 42, 1137–1153. [Google Scholar] [CrossRef]
- Kvitek, R.G.; DeGange, A.R.; Beitler, M.K. Paralytic shelfish poisoning toxins mediate feeding behavior of sea otters. Limnol. Oceanogr. 1991, 36, 393–404. [Google Scholar] [CrossRef]
- Lefebvre, K.; Elder, N.; Hershberger, P.; Trainer, V.; Stehr, C.; Scholz, N. Dissolved saxitoxin causes transient inhibition of sensorimotor function in larval Pacific herring (Clupea harengus pallasi). Mar. Biol. 2005, 147, 1393–1402. [Google Scholar] [CrossRef]
- Lefebvre, K.A.; Trainer, V.L.; Scholz, N.L. Morphological abnormalities and sensorimotor deficits in larval fish exposed to dissolved saxitoxin. Aquat. Toxicol. 2004, 66, 159–170. [Google Scholar] [CrossRef]
- Goldstein, T.; Zabka, T.S.; DeLong, R.L.; Wheeler, E.A.; Ylitalo, G.; Bargu, S.; Silver, M.; Leighfield, T.; van Dolah, F.; Langlois, G.; et al. The role of domoic acid in abortion and premature parturition of California sea lions (Zalophus californianus) on San Miguel Island, California. J. Wildl. Dis. 2009, 45, 91–108. [Google Scholar]
- Gentien, P.; Arzul, G. Exotoxin production by Gyrodinium cf. aureolum (Dinophyceae). J. Mar. Biol. Assoc. UK 1990, 70, 571–581. [Google Scholar] [CrossRef]
- Shilo, M.; Aschner, M. Factors governing the toxicity of cultures containing the Phytoflagellate Prymnesium parvum Carter. J. Gen. Microbiol. 1953, 8, 333–343. [Google Scholar] [CrossRef]
- Bourdelais, A.J.; Tomas, C.R.; Naar, J.; Kubanek, J.; Baden, D.G. New fish-killing alga in coastal Delaware produces neurotoxins. Environ. Health Perspect. 2002, 110, 465–470. [Google Scholar] [CrossRef]
- De la Riva, G.T.; Johnson, C.K.; Gulland, F.M.D.; Langlois, G.W.; Heyning, J.E.; Rowles, T.K.; Mazet, J.A.K. Association of an unusual marine mammal mortality event with Pseudo-nitzschia spp. blooms along the southern california coastline. J. Wildl. Dis. 2009, 45, 109–121. [Google Scholar]
- Fire, S.E.; Zhihong, W.; Berman, M.; Langlois, G.W.; Morton, S.L.; Sekula-Wood, E.; Benitez-Nelson, C.R. Trophic transfer of the harmful algal toxin domoic acid as a cause of death in a Minke whale (Balaenoptera acutorostrata) stranding in Southern California. Aquat. Mamm. 2010, 36, 342–350. [Google Scholar] [CrossRef]
- Geraci, J.R.; Anderson, D.M.; Timperi, R.J.; St. Aubin, D.J.; Early, G.A.; Prescott, J.H.; Mayo, C.A. Humpback whales (Megaptera novaeangliae) fatally poisoned by dinoflagellate toxin. Can. J. Fish. Aquat. Sci. 1989, 46, 1895–1898. [Google Scholar] [CrossRef]
- Flewelling, L.J.; Naar, J.P.; Abbott, J.P.; Baden, D.G.; Barros, N.B.; Bossart, G.D.; Bottein, M.-Y.D.; Hammond, D.G.; Haubold, E.M.; Heil, C.A. Brevetoxicosis: Red tides and marine mammal mortalities. Nature 2005, 435, 755–756. [Google Scholar] [CrossRef]
- Glibert, P.M.; Landsberg, J.H.; Evans, J.J.; Al-Sarawi, M.A.; Faraj, M.; Al-Jarallah, M.A.; Haywood, A.; Ibrahem, S.; Klesius, P.; Powell, C. A fish kill of massive proportion in Kuwait Bay, Arabian Gulf, 2001: The roles of bacterial disease, harmful algae, and eutrophication. Harmful Algae 2002, 1, 215–231. [Google Scholar] [CrossRef]
- White, A.W. Dinoflagellate toxins as probable cause of an Atlantic herring (Clupea harengus harengus) kill, and pteropods as apparent vector. J. Fish. Board Can. 1977, 34, 2421–2424. [Google Scholar] [CrossRef]
- White, A.W. Marine zooplankton can accumulate and retain dinoflagellate toxins and cause fish kills. Limnol. Oceanogr. 1981, 26, 103–109. [Google Scholar] [CrossRef]
- White, A.W. Recurrence of kills of Atlantic herring (Clupea harengus harengus) caused by dinoflagellate toxins transferred through herbivorous zooplankton. Can. J. Fish. Aquat. Sci. 1980, 37, 2262–2265. [Google Scholar] [CrossRef]
- Daneri, G.A.; Carlini, A.R.; Rodhouse, P.G.K. Cephalopod diet of the southern elephant seal, Mirounga leonina, at King George Island, South Shetland Islands. Antarct. Sci. 2000, 12, 16–19. [Google Scholar]
- Clarke, M.; Goodall, N. Cephalopods in the diets of three odontocete cetacean species stranded at Tierra del Fuego, Globicephala melaena (Traill, 1809), Hyperoodon planifrons Flower, 1882 and Cephalorhynchus commersonii (Lacepede, 1804). Antarct. Sci. 1994, 6, 149–154. [Google Scholar]
- Pauly, D.; Trites, A.W.; Capuli, E.; Christensen, V. Diet composition and trophic levels of marine mammals. ICES J. Mar. Sci. 1998, 55, 467–481. [Google Scholar] [CrossRef]
- Brock, R.E. Preliminary study of the feeding habits of pelagic fish around Hawaiian fish aggregation devices enhance local fisheries productivity? Bull. Mar. Sci. 1985, 37, 40–49. [Google Scholar]
- Stillwell, C.E.; Kohler, N.E. Food, feeding habits, and estimates of daily ration of the shortfin mako (Isurus oxyrinchus) in the Northwest Atlantic. Can. J. Fish. Aquat. Sci. 1982, 39, 407–414. [Google Scholar] [CrossRef]
- Croxall, J.P.; Prince, P.A. Cephalopods as Prey I: Seabirds. Philos. Trans. R. Soc. Lond. B 1996, 351, 1023–1043. [Google Scholar] [CrossRef]
- Nixon, M. Capture of prey, diet and feeding of Sepia officinalis and Octopus vulgaris (Mollusca: Cephalopoda) from hatchling to adult. Vie et Milieu 1985, 35, 255–261. [Google Scholar]
- Hanlon, R.T.; Messenger, J.B. Cephalopod Behaviour; Cambridge University Press: Cambridge, UK, 1996; p. 232. [Google Scholar]
- Rodhouse, P.G.; Nigmatullin, C.M. Role as consumers. Philos. Trans. R. Soc. Lond. B 1996, 351, 1003–1022. [Google Scholar] [CrossRef]
- Debonnel, G.; Beauchesne, L.; Montigny, C. Domoic acid, the alleged “mussel toxin”, might produce its neurotoxic effect through kainate receptor activation: An electrophysiological study in the rat dorsal hippocampus. Can. J. Physiol. Pharmacol. 1989, 67, 29–33. [Google Scholar] [CrossRef]
- Perl, T.M.; Bédard, L.; Kosatsky, T.; Hockin, J.C.; Todd, E.C.D.; Remis, R.S. An outbreak of toxic encephalopathy caused by eating mussels contaminated with domoic acid. N. Engl. J. Med. 1990, 322, 1775–1780. [Google Scholar] [CrossRef]
- Todd, E.C.D. Domoic acid and amnesic shellfish poisoning—A review. J. Food Prot. 1993, 56, 69–83. [Google Scholar]
- Bates, S.S.; Bird, C.J.; Freitas, A.S.W.; Foxall, R.; Gilgan, M.; Hanic, L.A.; Johnson, G.R.; McCulloch, A.W.; Odense, P.; Pocklington, R.; et al. Pennate diatom Nitzschia pungens as the primary source of domoic acid, a toxin in shellfish from Eastern Prince Edward Island, Canada. Can. J. Fish. Aquat. Sci. 1989, 46, 1203–1215. [Google Scholar] [CrossRef]
- Bates, S.S. Domoic-acid-producing diatoms: Another genus added! J. Phycol. 2000, 36, 978–983. [Google Scholar] [CrossRef]
- Rao, D.V.S.; Quilliam, M.A.; Pocklington, R. Domoic acid—A neurotoxic amino acid produced by the marine diatom Nitzschia pungens in culture. Can. J. Fish. Aquat. Sci. 1988, 45, 2076–2079. [Google Scholar] [CrossRef]
- Garrison, D.L.; Conrad, S.M.; Eilers, P.P.; Waldron, E.M. Confirmation of domoic acid production by Pseudonitzschia australis (Bacillariophyceae) cultures. J. Phycol. 1992, 28, 604–607. [Google Scholar]
- Lundholm, N.; Moestrup, Ø.; Hasle, G.R.; Hoef-Emden, K. A study of the Pseudo-nitzschia pseudodelicatissima/cuspidata complex (Bacillariophyceae): What is P. pseudodelicatissima? J. Phycol. 2003, 39, 797–813. [Google Scholar] [CrossRef]
- Scholin, C.A.; Gulland, F.; Doucette, G.J.; Benson, S.; Busman, M.; Chavez, F.P.; Cordaro, J.; DeLong, R.; de Vogelaere, A.; Harvey, J. Mortality of sea lions along the central California coast linked to a toxic diatom bloom. Nature 2000, 403, 80–84. [Google Scholar] [CrossRef]
- Silvagni, P.A.; Lowenstine, L.J.; Spraker, T.; Lipscomb, T.P.; Gulland, F.M.D. Pathology of domoic acid toxicity in California Sea Lions (Zalophus californianus). Vet. Pathol. 2005, 42, 184–191. [Google Scholar] [CrossRef]
- Work, T.; Beale, A.; Fritz, L.; Quilliam, M.; Silver, M.; Buck, K.; Wright, J. Domoic Acid Intoxication of Brown Pelicans and Cormorants in Santa Cruz, California. In Toxic PhytoplanktonBlooms in the Sea; Smayda, T.J., Shimizu, Y., Eds.; Elsevier: Amsterdam, The Netherlands, 1993; pp. 643–649. [Google Scholar]
- Shumway, S.E.; Allen, S.M.; Dee Boersma, P. Marine birds and harmful algal blooms: Sporadic victims or under-reported events? Harmful Algae 2003, 2, 1–17. [Google Scholar] [CrossRef]
- Costa, P.R.; Rosa, R.; Sampayo, M.A.M. Tissue distribution of the amnesic shellfish toxin, domoic acid, in Octopus vulgaris from the Portuguese Coast. Mar. Biol. 2004, 144, 971–976. [Google Scholar] [CrossRef]
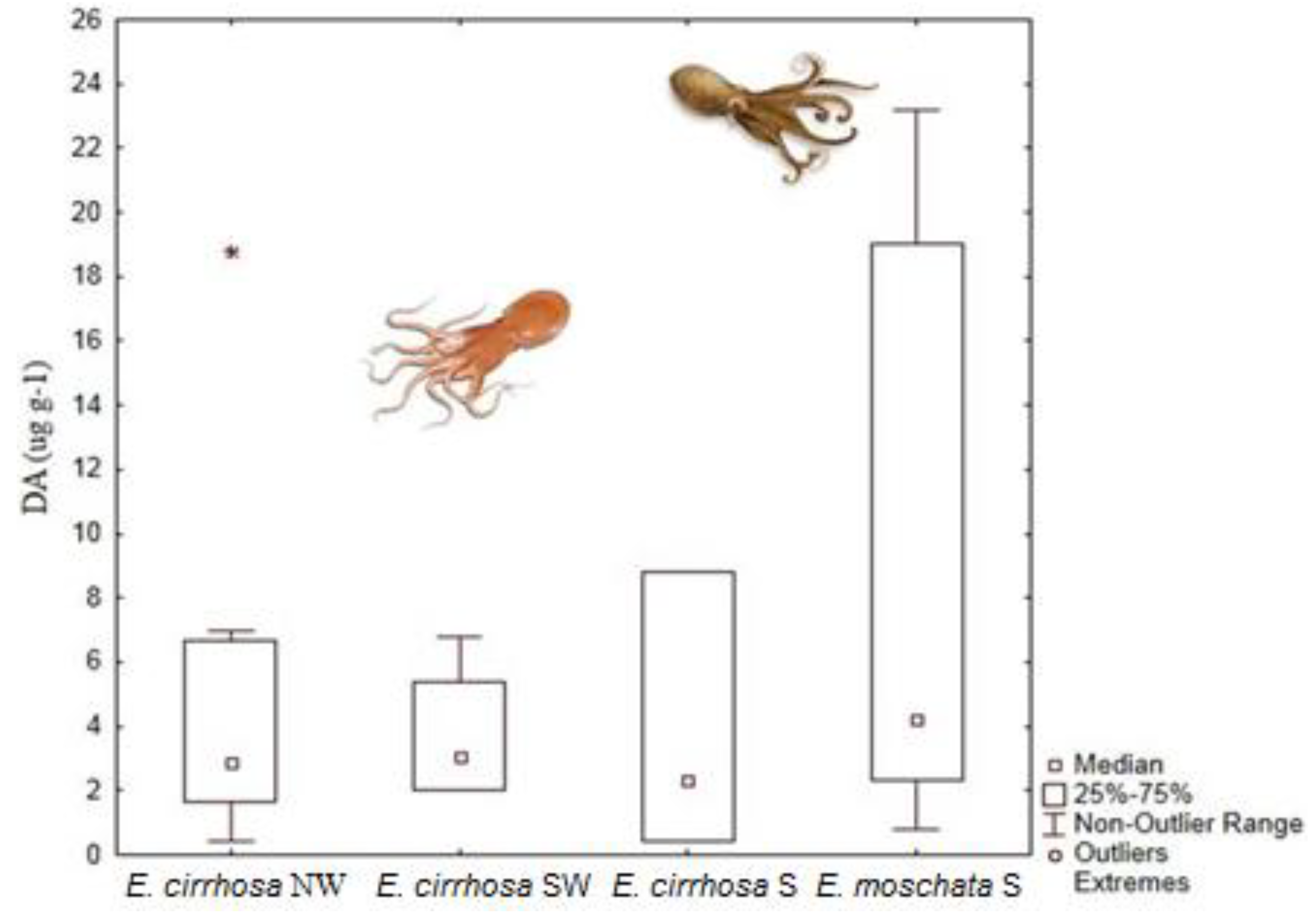
- Costa, P.R.; Rosa, R.; Pereira, J.; Sampayo, M.A.M. Detection of domoic acid, the amnesic shellfish toxin, in the digestive gland of Eledone cirrhosa and E. moschata (Cephalopoda, Octopoda) from the Portuguese Coast. Aquat. Living Resour. 2005, 18, 395–400. [Google Scholar] [CrossRef]
- Costa, P.R.; Rosa, R.; Duarte-Silva, A.; Brotas, V.; Sampayo, M.A.M. Accumulation, transformation and tissue distribution of domoic acid, the amnesic shellfish poisoning toxin, in the common cuttlefish, Sepia officinalis. Aquat. Toxicol. 2005, 74, 82–91. [Google Scholar] [CrossRef]
- Bargu, S.; Powell, C.L.; Wang, Z.; Doucette, G.J.; Silver, M.W. Note on the occurrence of Pseudo-nitzschia australis and domoic acid in squid from Monterey Bay, CA (USA). Harmful Algae 2008, 7, 45–51. [Google Scholar] [CrossRef]
- Braid, H.; Deeds, J.; DeGrasse, S.; Wilson, J.; Osborne, J.; Hanner, R. Preying on commercial fisheries and accumulating paralytic shellfish toxins: A dietary analysis of invasive Dosidicus gigas (Cephalopoda Ommastrephidae) stranded in Pacific Canada. Mar. Biol. 2012, 159, 25–31. [Google Scholar] [CrossRef]
- Monteiro, A.; Costa, P.R. Distribution and selective elimination of paralytic shellfish toxins in different tissues of Octopus vulgaris. Harmful Algae 2011, 10, 732–737. [Google Scholar] [CrossRef]
- Robertson, A.; Stirling, D.; Robillot, C.; Llewellyn, L.; Negri, A. First report of saxitoxin in octopi. Toxicon 2004, 44, 765–771. [Google Scholar] [CrossRef]
- Milandri, A.; Ceredi, A.; Riccardi, E.; Gasperetti, L.; Susini, F.; Casotti, M.; Faiman, L.; Pigozzi, S. Impact of Ostreopsis Ovata on Marine Benthic Communities: Accumulation of Palytoxins in Mussels, Sea Urchins and Octopuses from Italy. In Proceedings of ICHA14 Conference, Crete, Greek, 1–5 November 2010; pp. 24–27.
- Quilliam, M.A.; Wright, J.L.C. The amnesic shellfish poisoning mystery. Anal. Chem. 1989, 61, 1053–1060. [Google Scholar]
- Wekell, J.C.; Hurst, J.; Lefebvre, K.A. The origin of the regulatory limits for PSP and ASP toxins in shellfish. J. Shellfish Res. 2004, 23, 927–930. [Google Scholar]
- Bargu, S.; Powell, C.L.; Coale, S.L.; Busman, M.; Doucette, G.J.; Silver, M.W. Krill: A potential vector for domoic acid in marine food webs. Mar. Ecol. Prog. Ser. 2002, 237, 209–216. [Google Scholar] [CrossRef]
- Lincoln, J.; Turner, J.; Bates, S.; Léger, C.; Gauthier, D. Feeding, egg production, and egg hatching success of the copepods Acartia tonsa and Temora longicornis on diets of the toxic diatom Pseudo-nitzschia multiseries and the non-toxic diatom Pseudo-nitzschia pungens. Hydrobiologia 2001, 453–454, 107–120. [Google Scholar] [CrossRef]
- Tester, P.A.; Pan, Y.; Doucette, G.J. Accumulation of Domoic Acid Activity in Copepods. In Proceedings of the 9th International Conference on Harmful Algal Blooms, Hobart, Australia, 7–11 February 2000; Hallegraeff, G.M.B., Blackburn, S.I., Bolch, C.J., Lewis, R.J., Eds.; UNESCO: Hobart, Australia, 2001; pp. 418–420. [Google Scholar]
- Lefebvre, K.; Silver, M.; Coale, S.; Tjeerdema, R. Domoic acid in planktivorous fish in relation to toxic Pseudo-nitzschia cell densities. Mar. Biol. 2002, 140, 625–631. [Google Scholar] [CrossRef]
- Lefebvre, K.A.; Dovel, S.L.; Silver, M.W. Tissue distribution and neurotoxic effects of domoic acid in a prominent vector species, the northern anchovy Engraulis mordax. Mar. Biol. 2001, 138, 693–700. [Google Scholar] [CrossRef]
- Vale, P.; Sampayo, M.A.M. Domoic acid in Portuguese shellfish and fish. Toxicon 2001, 39, 893–904. [Google Scholar] [CrossRef]
- Costa, P.R.; Garrido, S. Domoic acid accumulation in the sardine Sardina pilchardus and its relationship to Pseudo-nitzschia diatom ingestion. Mar. Ecol. Prog. Ser. 2004, 284, 261–268. [Google Scholar] [CrossRef]
- Costa, P.R.; Botelho, M.J.; Rodrigues, S.M. Accumulation of paralytic shellfish toxins in digestive gland of Octopus vulgaris during bloom events including the dinoflagellate Gymnodinium catenatum. Mar. Pollut. Bull. 2009, 58, 1747–1750. [Google Scholar] [CrossRef]
- Oshima, Y. Postcolumn derivatization liquid chromatographic method for paralytic shellfish toxins. J. AOAC Int. 1995, 78, 528–532. [Google Scholar]
- Robineau, B.; Gagné, J.A.; Fortier, L.; Cembella, A.D. Potential impact of a toxic dinoflagellate (Alexandrium excavatum) bloom on survival of fish and crustacean larvae. Mar. Biol. 1991, 108, 293–301. [Google Scholar] [CrossRef]
- Samson, J.C.; Shumway, S.E.; Weis, J.S. Effects of the toxic dinoflagellate, Alexandrium fundyense on three species of larval fish: A food-chain approach. J. Fish Biol. 2008, 72, 168–188. [Google Scholar] [CrossRef]
- Jester, R.; Baugh, K.; Lefebvre, K. Presence of Alexandrium catenella and paralytic shellfish toxins in finfish, shellfish and rock crabs in Monterey Bay, California, USA. Mar. Biol. 2009, 156, 493–504. [Google Scholar] [CrossRef]
- Costa, P.R.; Botelho, M.J.; Lefebvre, K.A. Characterization of paralytic shellfish toxins in seawater and sardines (Sardina pilchardus) during blooms of Gymnodinium catenatum. Hydrobiologia 2010, 655, 89–97. [Google Scholar] [CrossRef]
- Mianzan, H.W.; Pájaro, M.; Machinandiarena, L.; Cremonte, F. Salps: Possible vectors of toxic dinoflagellates? Fish. Res. 1997, 29, 193–197. [Google Scholar] [CrossRef]
- Lage, S.; Reis Costa, P. Paralytic shellfish toxins in the Atlantic horse mackerel (Trachurus trachurus) over a bloom of Gymnodinium catenatum: The prevalence of decarbamoylsaxitoxin in the marine food web. Sci. Mar. 2013, 77, 13–17. [Google Scholar] [CrossRef]
- Tanti, J.F.; Grémeaux, T.; Van Obberghen, E.; le Marchand-Brustel, Y. Effects of okadaic acid, an inhibitor of protein phosphatases-1 and -2A, on glucose transport and metabolism in skeletal muscle. J. Biol. Chem. 1991, 266, 2099–2103. [Google Scholar]
- Maneiro, I.; Frangópulos, M.; Guisande, C.; Fernández, M.; Reguera, B.; Riveiro, I. Zooplankton as a potential vector of diarrhetic shellfish poisoning toxins through the food web. Mar. Ecol. Prog. Ser. 2000, 201, 155–163. [Google Scholar] [CrossRef]
- Krock, B.; Seguel, C.G.; Valderrama, K.; Tillmann, U. Pectenotoxins and yessotoxin from arica bay, north chile as determined by tandem mass spectrometry. Toxicon 2009, 54, 364–367. [Google Scholar] [CrossRef]
- Paz, B.; Riobó, P.; Luisa Fernández, M.; Fraga, S.; Franco, J.M. Production and release of yessotoxins by the dinoflagellates Protoceratium reticulatum and Lingulodinium polyedrum in culture. Toxicon 2004, 44, 251–258. [Google Scholar] [CrossRef]
- Miles, C.O.; Samdal, I.A.; Aasen, J.A.; Jensen, D.J.; Quilliam, M.A.; Petersen, D.; Briggs, L.M.; Wilkins, A.L.; Rise, F.; Cooney, J.M. Evidence for numerous analogs of yessotoxin in Protoceratium reticulatum. Harmful Algae 2005, 4, 1075–1091. [Google Scholar] [CrossRef]
- Rhodes, L.; McNabb, P.; de Salas, M.; Briggs, L.; Beuzenberg, V.; Gladstone, M. Yessotoxin production by Gonyaulax spinifera. Harmful Algae 2006, 5, 148–155. [Google Scholar] [CrossRef]
- Riccardi, M.; Guerrini, F.; Roncarati, F.; Milandri, A.; Cangini, M.; Pigozzi, S.; Riccardi, E.; Ceredi, A.; Ciminiello, P.; Dell’Aversano, C. Gonyaulax spinifera from the Adriatic sea: Toxin production and phylogenetic analysis. Harmful Algae 2009, 8, 279–290. [Google Scholar] [CrossRef]
- Krock, B.; Tillmann, U.; John, U.; Cembella, A.D. Characterization of azaspiracids in plankton size-fractions and isolation of an azaspiracid-producing dinoflagellate from the North Sea. Harmful Algae 2009, 8, 254–263. [Google Scholar] [CrossRef]
- Wandscheer, C.B.; Vilariño, N.; Espiña, B.A.; Louzao, M.C.; Botana, L.M. Human muscarinic acetylcholine receptors are a target of the marine toxin 13-desmethyl C spirolide. Chem. Res. Toxicol. 2010, 23, 1753–1761. [Google Scholar] [CrossRef]
- Vasconcelos, V.; Azevedo, J.; Silva, M.; Ramos, V. Effects of marine toxins on the reproduction and early stages development of aquatic organisms. Mar. Drugs 2010, 8, 59–79. [Google Scholar] [CrossRef]
- Landsberg, J.H.; Steidinger, K. A historical review of Gymnodinium breve red tides implicated in mass mortalities of the manatee (Trichechus manatus latirostris) in Florida, USA. In Proceedings of the 8th International Conference on Harmful Algae, Vigo, Spain, 25–29 June 1997; pp. 97–100.
- Bagnis, R.; Chanteau, S.; Chungue, E.; Hurtel, J.; Yasumoto, T.; Inoue, A. Origins of ciguatera fish poisoning: A new dinoflagellate, Gambierdiscus toxicus Adachi and Fukuyo, definitively involved as a causal agent. Toxicon 1980, 18, 199–208. [Google Scholar] [CrossRef]
- Usami, M.; Satake, M.; Ishida, S.; Inoue, A.; Kan, Y.; Yasumoto, T. Palytoxin analogs from the dinoflagellate Ostreopsis siamensis. J. Am. Chem. Soc. 1995, 117, 5389–5390. [Google Scholar] [CrossRef]
- Wang, D.-Z. Neurotoxins from marine dinoflagellates: A brief review. Mar. Drugs 2008, 6, 349–371. [Google Scholar] [CrossRef]
- Shears, N.T.; Ross, P.M. Blooms of benthic dinoflagellates of the genus Ostreopsis; An increasing and ecologically important phenomenon on temperate reefs in New Zealand and worldwide. Harmful Algae 2009, 8, 916–925. [Google Scholar] [CrossRef]
- Rhodes, L.; Towers, N.; Briggs, L.; Munday, R.; Adamson, J. Uptake of palytoxin-like compounds by shellfish fed Ostreopsis siamensis (Dinophyceae). N. Z. J. Mar. Freshw. Res. 2002, 36, 631–636. [Google Scholar] [CrossRef]
- Ciminiello, P.; Dell’Aversano, C.; Fattorusso, E.; Forino, M.; Magno, G.S.; Tartaglione, L.; Grillo, C.; Melchiorre, N. The genoa 2005 outbreak. Determination of putative palytoxin in Mediterranean Ostreopsis ovata by a new liquid chromatography tandem mass spectrometry method. Anal. Chem. 2006, 78, 6153–6159. [Google Scholar] [CrossRef]
- Totti, C.; Accoroni, S.; Cerino, F.; Cucchiari, E.; Romagnoli, T. Ostreopsis ovata bloom along the Conero Riviera (Northern Adriatic Sea): Relationships with environmental conditions and substrata. Harmful Algae 2010, 9, 233–239. [Google Scholar] [CrossRef]
- Valverde, I.; Lago, J.; Vieites, J.M.; Cabado, A.G. In vitro approaches to evaluate palytoxin-induced toxicity and cell death in intestinal cells. J. Appl. Toxicol. 2008, 28, 294–302. [Google Scholar] [CrossRef]
- Gorbi, S.; Bocchetti, R.; Binelli, A.; Bacchiocchi, S.; Orletti, R.; Nanetti, L.; Raffaelli, F.; Vignini, A.; Accoroni, S.; Totti, C. Biological effects of palytoxin-like compounds from Ostreopsis cf. ovata: A multibiomarkers approach with mussels Mytilus galloprovincialis. Chemosphere 2012, 89, 623–632. [Google Scholar]
- Rosa, R.; Dierssen, H.M.; Gonzalez, L.; Seibel, B.A. Ecological biogeography of cephalopod molluscs in the atlantic ocean: Historical and contemporary causes of coastal diversity patterns. Glob. Ecol. Biogeogr. 2008, 17, 600–610. [Google Scholar] [CrossRef]
- Rosa, R.; Dierssen, H.M.; Gonzalez, L.; Seibel, B.A. Large-scale diversity patterns of cephalopods in the Atlantic open ocean and deep sea. Ecology 2008, 89, 3449–3461. [Google Scholar] [CrossRef]
- Rosa, R.; Gonzalez, L.; Dierssen, H.M.; Seibel, B.A. Environmental determinants of latitudinal size-trends in cephalopods. Mar. Ecol. Prog. Ser. 2012, 464, 153–165. [Google Scholar] [CrossRef]
- Wells, M.J. Oxygen extraction and jet propulsion in cephalopods. Can. J. Zool. 1988, 68, 815–824. [Google Scholar] [CrossRef]
- Wells, M.J.; Wells, J. The circulatory response to acute hypoxia in Octopus. J. Exp. Biol. 1983, 104, 59–71. [Google Scholar]
- Wells, M.J.; Wells, J. Blood flow in acute hypoxia in a cephalopod. J. Exp. Biol. 1986, 122, 345–353. [Google Scholar]
- Eno, C.N. The morphometrics of cephalopod gills. J. Mar. Biol. Assoc. UK 1994, 74, 687–706. [Google Scholar] [CrossRef]
- Hanlon, R.T.; Chiao, C.C.; Mäthger, L.M.; Barbosa, A.; Buresch, K.C.; Chubb, C. Cephalopod dynamic camouflage: Bridging the continuum between background matching and disruptive coloration. Philos. Trans. R. Soc. Lond. B 2009, 364, 429–437. [Google Scholar] [CrossRef]
- Rosa, R.; Marques, A.M.; Nunes, M.L.; Bandarra, N.; Reis, C.S. Spatial-temporal changes in dimethyl acetal (octadecanal) levels of Octopus vulgaris (Mollusca, Cephalopoda): Relation to feeding ecology. Sci. Mar. 2004, 68, 227–236. [Google Scholar]
- Markaida, U. Food and feeding of jumbo squid Dosidicus gigas in the Gulf of California and adjacent waters after the 1997–98 El Niño event. Fish. Res. 2006, 79, 16–27. [Google Scholar] [CrossRef]
- Castro, B.G.; Guerra, A. The diet of Sepia officinalis (Linaeus, 1785) and Sepia elegans (Orbigny, 1835) (Cephalopoda, Sepioidea) from the Ria de Vigo (NW Spain). Sci. Mar. 1990, 54, 375–388. [Google Scholar]
- Coelho, M.; Domingues, P.; Balguerias, E.; Fernandez, M.; Andrade, J.P. A comparative study of the diet of Loligo vulgaris (Lamarck, 1799) (Mollusca: Cephalopoda) from the south coast of Portugal and the Saharan Bank (central-east Atlantic). Fish. Res. 1997, 29, 245–255. [Google Scholar] [CrossRef]
- Cortez, T.; Castro, B.G.; Guerra, A. Feeding dynamics of Octopus mimus (Mollusca: Cephalopoda) in northern Chile waters. Mar. Biol. 1995, 123, 497–503. [Google Scholar] [CrossRef]
- Smale, M.J.; Buchan, P.R. Biology of Octopus vulgaris off the east coast of South Africa. Mar. Biol. 1981, 65, 1–12. [Google Scholar] [CrossRef]
- Ambrose, R.F.; Nelson, B. Predation by Octopus vulgaris in the Mediterrean. PSZNI Mar. Ecol. 1983, 4, 251–261. [Google Scholar] [CrossRef]
- Altman, J.S. The behaviour of Octopus vulgaris Lam, in its natural habitat: A pilot study. Underw. Assoc. Rep. 1967, 1966, 77–83. [Google Scholar]
- Nigmatullin, C.M.; Ostapenko, A. Feeding of Octopus vulgaris Lam. from the Northwest African Coast. ICES CM 1976, 1976, 15. [Google Scholar]
- Guerra, A. Sobre la alimentacion y el comportamiento alimentario de Octopus vulgaris. Invest. Pesq. 1978, 42, 351–364. [Google Scholar]
- Sanchez, P.; Obarti, R.; Okutani, T.; O’Dor, R.K.; Kubodera, T. The Biology and Fishery of Octopus vulgaris Caught with Clay Pots on the Spanish Mediterranean Coast. In Recent Advances in Cephalopod Fisheries Biology; Tokay University Press: Tokyo, Japan, 1993; pp. 477–487. [Google Scholar]
- Quetglas, A.; Alemany, F.; Carbonell, A.; Merella, P.; Sanchez, P. Biology and fishery of Octopus vulgaris Cuvier, 1797, caught by trawlers in Mallorca (Balearic Sea, Western Mediterraneam). Fish. Res. 1998, 36, 237–249. [Google Scholar] [CrossRef]
- Villanueva, R. Decapod crab zoeae as food for rearing cephalopod paralarvae. Aquaculture 1994, 128, 143–152. [Google Scholar] [CrossRef]
- Boyle, P.; Rodhouse, P.G. Cephalopods: Ecology and Fisheries; Blackwell Publishing: Oxford, UK, 2005. [Google Scholar]
- Rosa, R.; Graham, P.; O’Dor, R. Advances in Squid Biology, Ecology and Fisheries, Volume I—Myopsid Squids; Nova Publishers: New York, NY, USA, 2013; in press. [Google Scholar]
- Rosa, R.; Graham, P.; O’Dor, R. Advances in Squid Biology, Ecology and Fisheries, Volume II—Oegopsid Squids; Nova Publishers: New York, NY, USA, in press.
- Rosa, R.; Seibel, B.A. Metabolic physiology of the humboldt squid, Dosidicus gigas: Implications for vertical migration in a pronounced oxygen minimum zone. Prog. Oceanogr. 2010, 86, 72–80. [Google Scholar] [CrossRef]
- Markaida, U.; Sosa-Nishizaki, O. Food and feeding habits of jumbo squid Dosidicus gigas (Cephalopoda: Ommastrephidae) from the Gulf of California, Mexico. J. Mar. Biol. Assoc. UK 2003, 83, 507–522. [Google Scholar] [CrossRef]
- Rosa, R.; Costa, P.R.; Bandarra, N.; Nunes, M.L. Changes in tissue biochemical composition and energy reserves associated with sexual maturation of Illex coindetii and Todaropsis eblanae. Biol. Bull. 2005, 208, 100–113. [Google Scholar] [CrossRef]
- Grisley, M.S.; Boyle, P.R. Recognition of food in Octopus digestive tract. J. Exp. Mar. Biol. Ecol. 1988, 118, 7–32. [Google Scholar] [CrossRef]
- Costa, P.R.; Pereira, J. Ontogenic differences in the concentration of domoic acid in the digestive gland of male and female Octopus vulgaris. Aquat. Biol. 2010, 9, 221–225. [Google Scholar] [CrossRef]
- García, B.G.; Giménez, F.A. Influence of diet on ongrowing and nutrient utilization in the common octopus (Octopus vulgaris). Aquaculture 2002, 211, 171–182. [Google Scholar] [CrossRef]
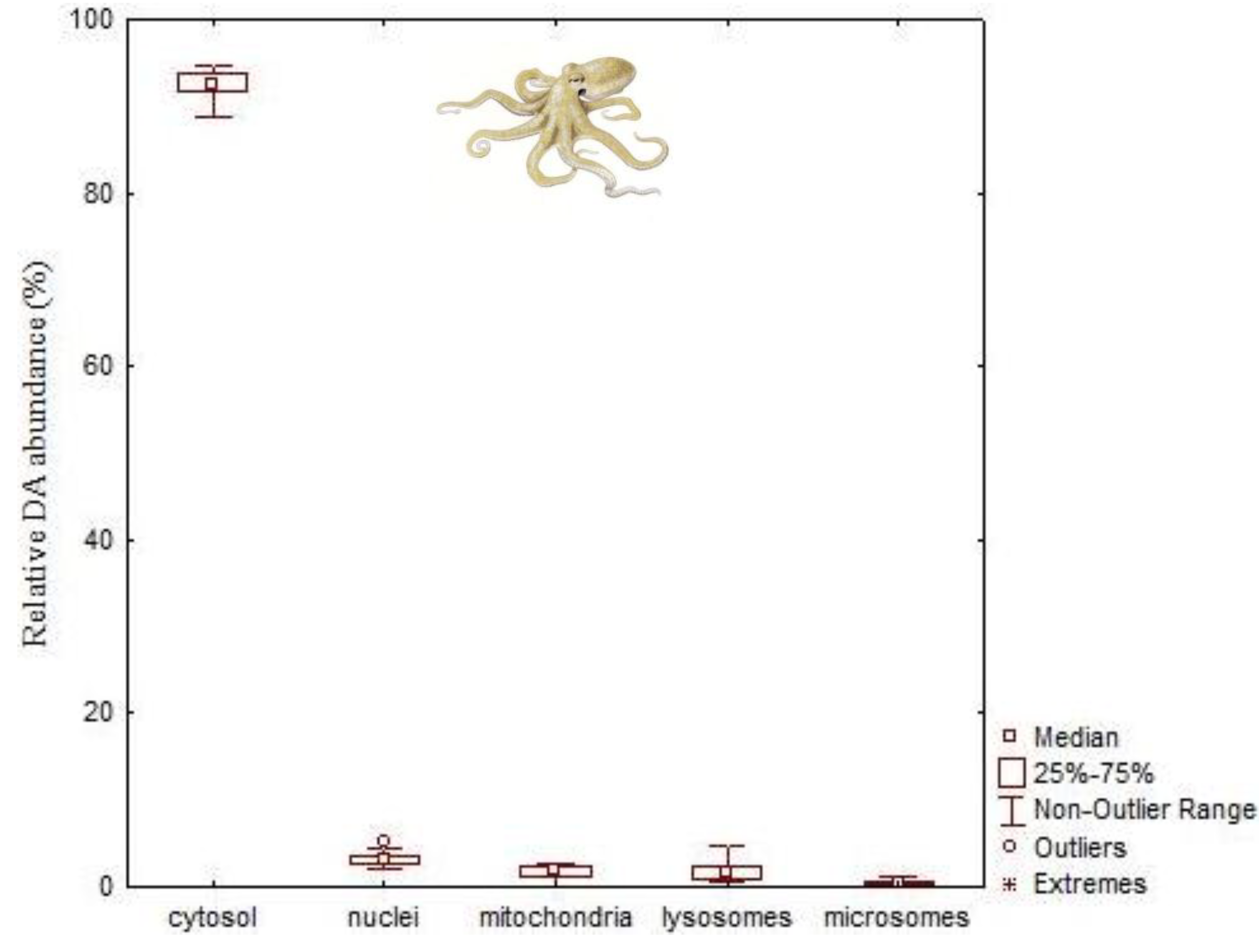
- Lage, S.; Raimundo, J.; Brotas, V.; Costa, P.R. Detection and sub-cellular distribution of the amnesic shellfish toxin, domoic acid, in the digestive gland of Octopus vulgaris during periods of toxin absence. Mar. Biol. Res. 2012, 8, 784–789. [Google Scholar] [CrossRef]
- Cuénot, K.; Bruntz, V. Recherches chimiques sur les coeurs branchiaux des Céphalopodes. Démonstration du rôle excreteur des cellules qui éliminenent le carmin ammoniacal des injections physiologiques. Arch. Zool. Exp. Gén. 1908, 9, 49–53. [Google Scholar]
- Schipp, R.; Hevert, F. Ultrafiltration in the branchial heart appendage of dibranchiate cephalopods: A comparative ultrastructural and physiological study. J. Exp. Biol. 1981, 92, 23–35. [Google Scholar]
- Nakahara, M.; Koyanagi, T.; Ueda, T.; Shimizu, C. Peculiar accumulation of cobalt-60 by the branchial heart of Octopus. Bull. Jpn. Soc. Sci. Fish. 1979, 45, 539. [Google Scholar]
- Nakahara, M.; Shimizu, C. Cobalt-binding substances in the branchial heart of Octopus vulgaris. Nippon Suisan Gakkaishi 1985, 51, 1195–1199. [Google Scholar] [CrossRef]
- Miramand, P.; Guary, J. High concentrations of some heavy metals in tissues of the Mediterranean octopus. Bull. Environ. Contam. Toxicol. 1980, 24, 783–788. [Google Scholar] [CrossRef]
- Martin, A.W.; Aldrich, F.A. Comparison of hearts and branchial heart appendages in some cephalopods. Can. J. Zool. 1970, 48, 751–756. [Google Scholar] [CrossRef]
- Boyle, P.R. Eledone Cirrhosa. In Cephalopod Life Cycles: Species Accounts; Boyle, P.R., Ed.; Academic Press: London, UK, 1983; Volume 1, pp. 365–386. [Google Scholar]
- Sanchez, P. Regime alimentaire d’Eledone cirrosa (Lamarck, 1798) (Mollusca, Cephalopoda) dans la mer Catalane. Rapp. Comm. Int. Mer Medit. 1981, 27, 209–212. [Google Scholar]
- Grisley, M.; Boyle, P.; Pierce, G.; Key, L. Factors affecting prey handling in lesser octopus (Eledone cirrhosa) feeding on crabs (Carcinus maenas). J. Mar. Biol. Assoc. UK 1999, 79, 1085–1090. [Google Scholar] [CrossRef]
- Mangold, K. Eledone Moschata. In Cephalopod Life Cycles: Species Accounts; Boyle, P.R., Ed.; Academic Press: London, UK, 1983; Volume 1, pp. 387–400. [Google Scholar]
- Davis, R.; Jaquet, N.; Gendron, D.; Markaida, U.; Bazzino, G.; Gilly, W. Diving behavior of sperm whales in relation to behavior of a major prey species, the jumbo squid, in the Gulf of California, Mexico. Mar. Ecol. Prog. Ser. 2007, 333, 291–302. [Google Scholar] [CrossRef]
- Fire, S.E.; Wang, Z.; Leighfield, T.A.; Morton, S.L.; McFee, W.E.; McLellan, W.A.; Litaker, R.W.; Tester, P.A.; Hohn, A.A.; Lovewell, G.; et al. Domoic acid exposure in pygmy and dwarf sperm whales (Kogia spp.) from southeastern and mid-Atlantic U.S. waters. Harmful Algae 2009, 8, 658–664. [Google Scholar] [CrossRef]
- Gleibs, S.; Mebs, D. Distribution and sequestration of palytoxin in coral reef animals. Toxicon 1999, 37, 1521–1527. [Google Scholar] [CrossRef]
- Mebs, D. Occurrence and sequestration of toxins in food chains. Toxicon 1998, 36, 1519–1522. [Google Scholar] [CrossRef]
- Li, S.-C.; Wang, W.-X. Radiotracer studies on the feeding of two marine bivalves on the toxic and nontoxic dinoflagellate Alexandrium tamarense. J. Exp. Mar. Biol. Ecol. 2001, 263, 65–75. [Google Scholar] [CrossRef]
- Pimentel, M.S.; Trübenbach, K.; Faleiro, F.; Boavida-Portugal, J.; Repolho, T.; Rosa, R. Impact of ocean warming on the early ontogeny of cephalopods: A metabolic approach. Mar. Biol. 2012, 159, 2051–2059. [Google Scholar] [CrossRef]
- Rosa, R.; Pimentel, M.S.; Boavida-Portugal, J.; Teixeira, T.; Trübenbach, K.; Diniz, M. Ocean warming enhances malformations, premature hatching, metabolic suppression and oxidative stress in the early life stages of a keystone squid. PLoS One 2012, 7, e38282. [Google Scholar]
- Liu, H.; Kelly, M.; Campbell, D.; Fang, J.; Zhu, J. Accumulation of domoic acid and its effect on juvenile king scallop Pecten maximus (Linnaeus, 1758). Aquaculture 2008, 284, 224–230. [Google Scholar] [CrossRef]
- Tiedeken, J.A.; Ramsdell, J.S.; Ramsdell, A.F. Developmental toxicity of domoic acid in zebrafish (Danio rerio). Neurotoxicol. Teratol. 2005, 27, 711–717. [Google Scholar] [CrossRef]
- Ajuzie, C.C. Palatability and fatality of the dinoflagellate Prorocentrum lima to Artemia salina. J. Appl. Phycol. 2007, 19, 513–519. [Google Scholar] [CrossRef]
- Zhenxing, W.; Yinglin, Z.; Mingyuan, Z.; Zongling, W.; Dan, W. Effects of toxic Alexandrium species on the survival and feeding rates of brine shrimp, Artemia salina. Acta Ecol. Sin. 2006, 26, 3942–3947. [Google Scholar] [CrossRef]
- Perez, M.; Sulkin, S. Palatability of autotrophic dinoflagellates to newly hatched larval crabs. Mar. Biol. 2005, 146, 771–780. [Google Scholar] [CrossRef]
- Sulkin, S.; Hinz, S.; Rodriguez, M. Effects of exposure to toxic and non-toxic dinoflagellates on oxygen consumption and locomotion in stage 1 larvae of the crabs Cancer oregonensis and C. magister. Mar. Biol. 2003, 142, 205–211. [Google Scholar]
- Gosselin, S.; Fortier, L.; Gagné, J. Vulnerability of marine fish larvae to the toxic dinoflagellate Protogonyaulax tamarensis. Mar. Ecol. Prog. Ser. 1989, 57, 1–10. [Google Scholar] [CrossRef]
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Lopes, V.M.; Lopes, A.R.; Costa, P.; Rosa, R. Cephalopods as Vectors of Harmful Algal Bloom Toxins in Marine Food Webs. Mar. Drugs 2013, 11, 3381-3409. https://doi.org/10.3390/md11093381
Lopes VM, Lopes AR, Costa P, Rosa R. Cephalopods as Vectors of Harmful Algal Bloom Toxins in Marine Food Webs. Marine Drugs. 2013; 11(9):3381-3409. https://doi.org/10.3390/md11093381
Chicago/Turabian StyleLopes, Vanessa M., Ana Rita Lopes, Pedro Costa, and Rui Rosa. 2013. "Cephalopods as Vectors of Harmful Algal Bloom Toxins in Marine Food Webs" Marine Drugs 11, no. 9: 3381-3409. https://doi.org/10.3390/md11093381