Production, Characterization, and Antioxidant Activity of Fucoxanthin from the Marine Diatom Odontella aurita

Abstract
:1. Introduction
2. Results and Discussion
2.1. Growth Kinetics and Fucoxanthin Production in the Bubble Column Photobioreactor
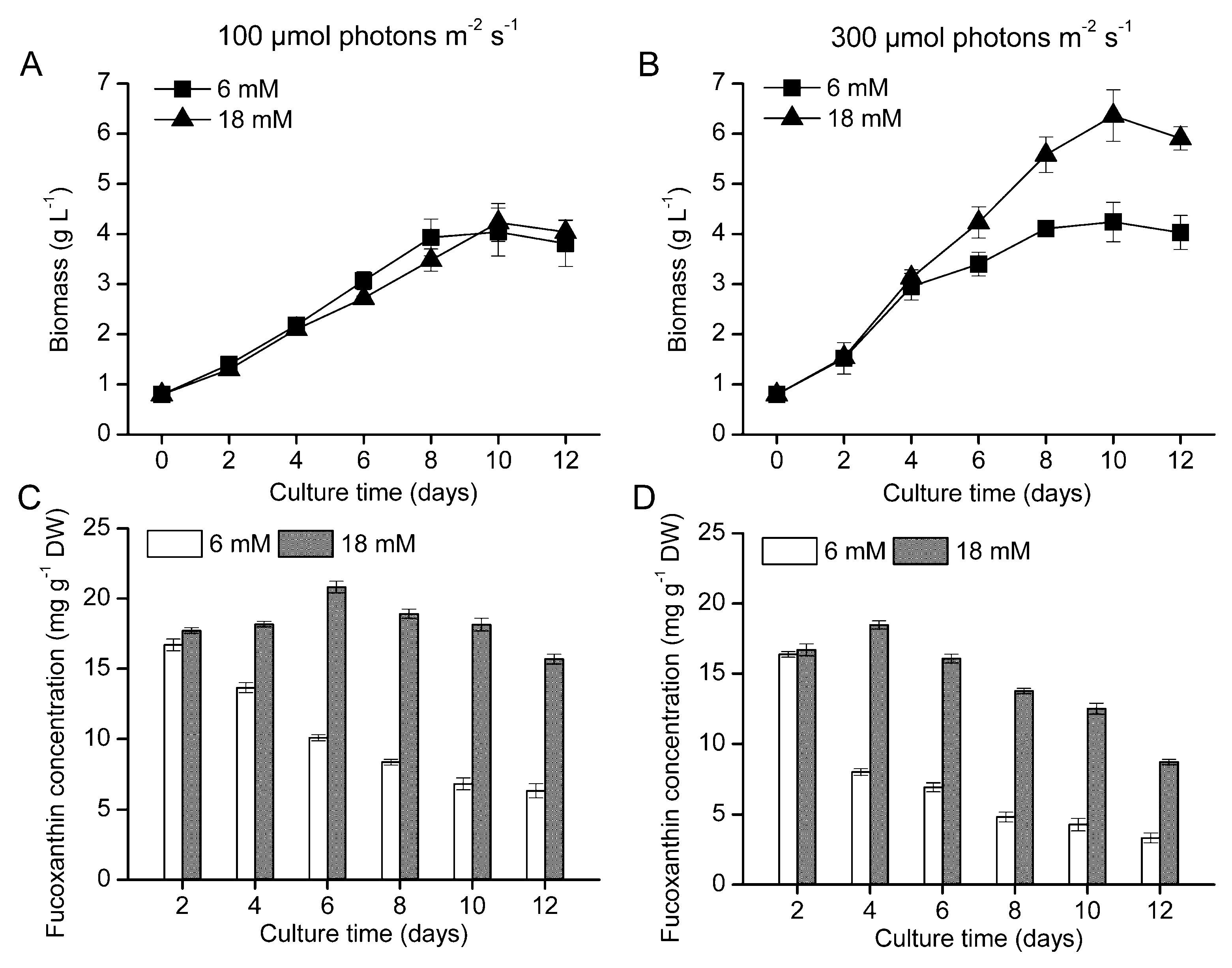

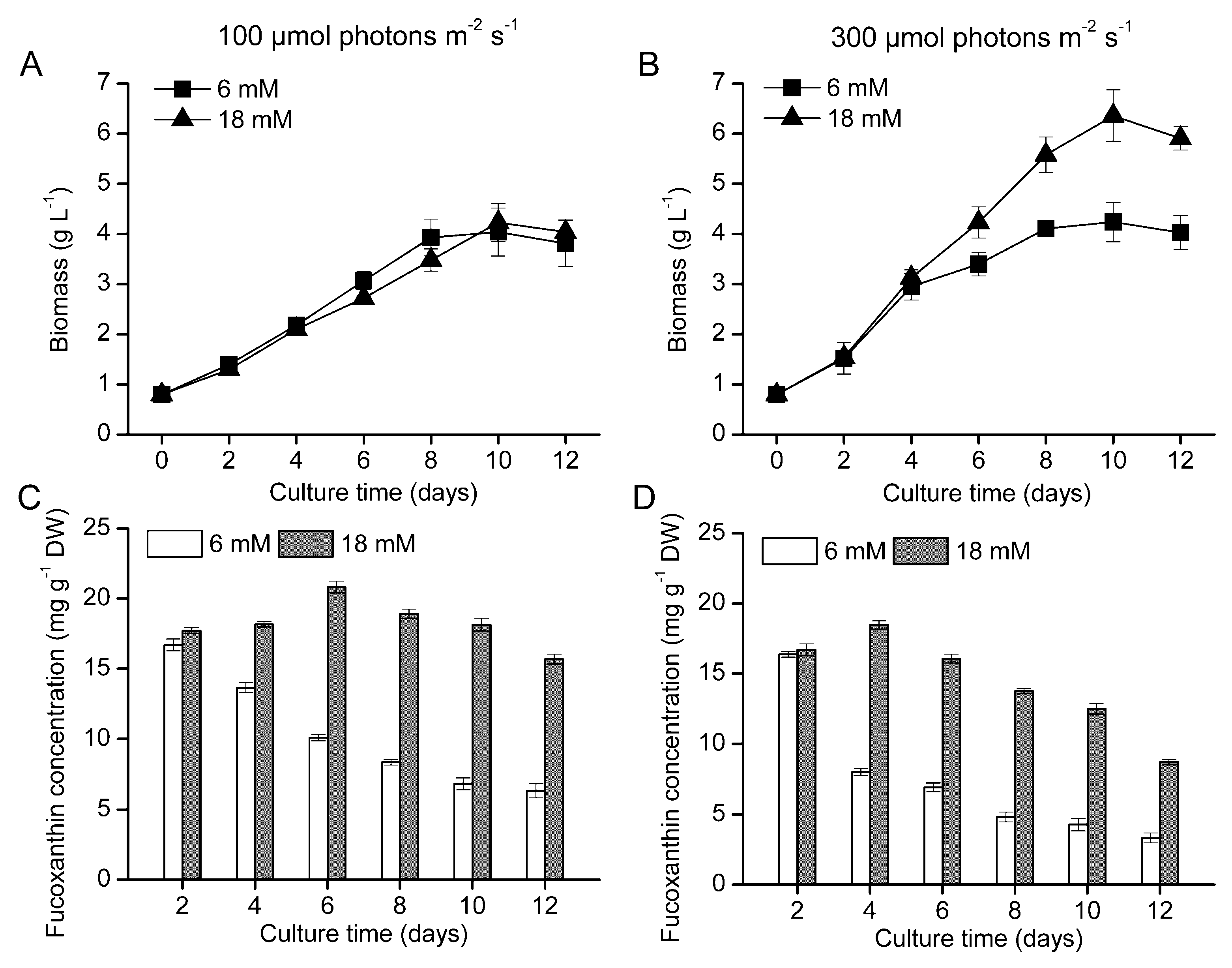{kind=link}
{kind=link}
{kind=link}
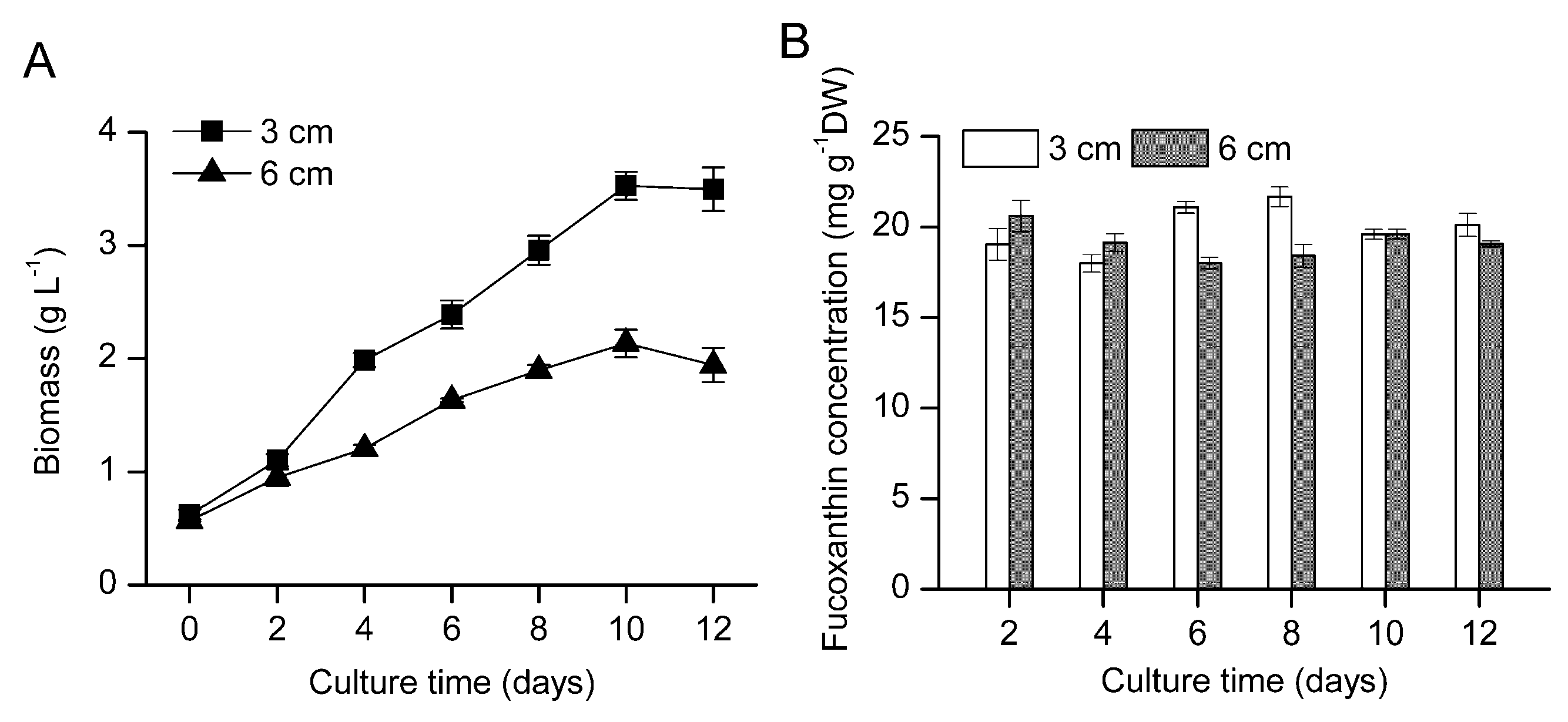
{kind=link}
{kind=link}
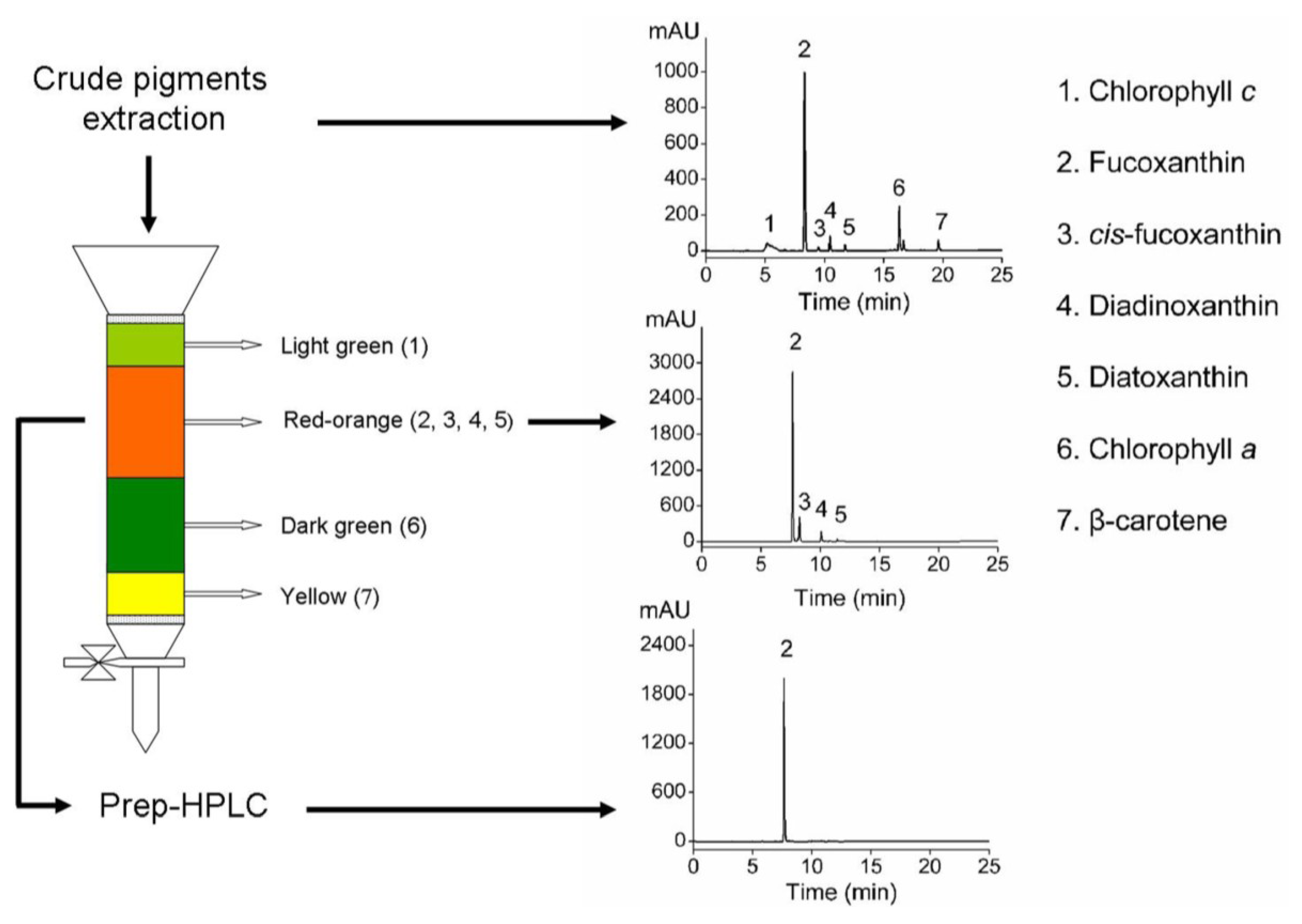
{kind=link}
| Fucoxanthin | ||||
|---|---|---|---|---|
| Light intensity (μmol photons m−2 s−1) | Nitrate concentration (mM) | Concentration (mg g−1) | Volumetric concentration (mg L−1) | Volumetric productivity (mg L−1 day−1) |
| 100 | 6 | 6.71 | 27.11 | 2.71 |
| 18 | 18.14 | 76.73 | 7.67 | |
| 300 | 6 | 4.28 | 18.15 | 1.82 |
| 18 | 12.51 | 79.56 | 7.96 | |
2.2. Mass Production Potential in the Pilot-Scale Flat Plate Photobioreactor

| Fucoxanthin | |||
|---|---|---|---|
| Light path (cm) | Concentration (mg g−1) | Volumetric concentration (mg L−1) | Volumetric productivity (mg L−1 day−1) |
| 3 | 19.59 | 69.15 | 6.92 |
| 6 | 19.61 | 41.97 | 4.20 |
| Species | Fresh or dried | Fucoxanthin concentration (mg g−1) | References | |
|---|---|---|---|---|
| Macroalgae | Eisenia bicyclis | Fresh | 0.26 | [7] 2 |
| Hizikia fusiformis | Fresh | 0.02 | [5] 2 | |
| Laminaria japonica | Fresh | 0.19 | [5] 2 | |
| Laminaria japonica | Fresh | 0.03 | [22] 2 | |
| Petalonia binghamiae | Fresh | 0.43–0.58 | [23] 3 | |
| Scytosiphon lomentaria | Fresh | 0.24–0.56 | [23] 3 | |
| Sargassum fusiforme | Dried | 0.01 | [22] 2 | |
| Sargassum binderib | Dried | 0.73 | [24] 2 | |
| Sargassum duplicatum | Dried | 1.01 | [24] 2 | |
| Sargassum plagyophyllum | Dried | 0.71 | [25] 2 | |
| Turbinaria turbinata | Dried | 0.59 | [25] 2 | |
| Undaria pinnatifida | Dried | 0.73 | [22] 2 | |
| Undaria pinnatifida | Fresh | 0.11 | [5] 2 | |
| Microalgae | Chaetoceros gracilis | Dried | 2.24 | [26] 2 |
| Cylindrotheca closterium | Dried | 5.23 | [27] 2 | |
| Isochrysis aff. Galbana | Dried | 18.23 | [26] 2 | |
| Isochrysis galbana | Dried | 6.04 | [26] 2 | |
| Phaeodactylum tricornutum | Dried | 8.55 | [26] 2 | |
| Phaeodactylum tricornutum | Dried | 15.42–16.51 | [7] 2 | |
| Nitzschia sp. | Dried | 4.92 | [26] 2 | |
| Odontella aurita | Dried | 21.67 | In this study |
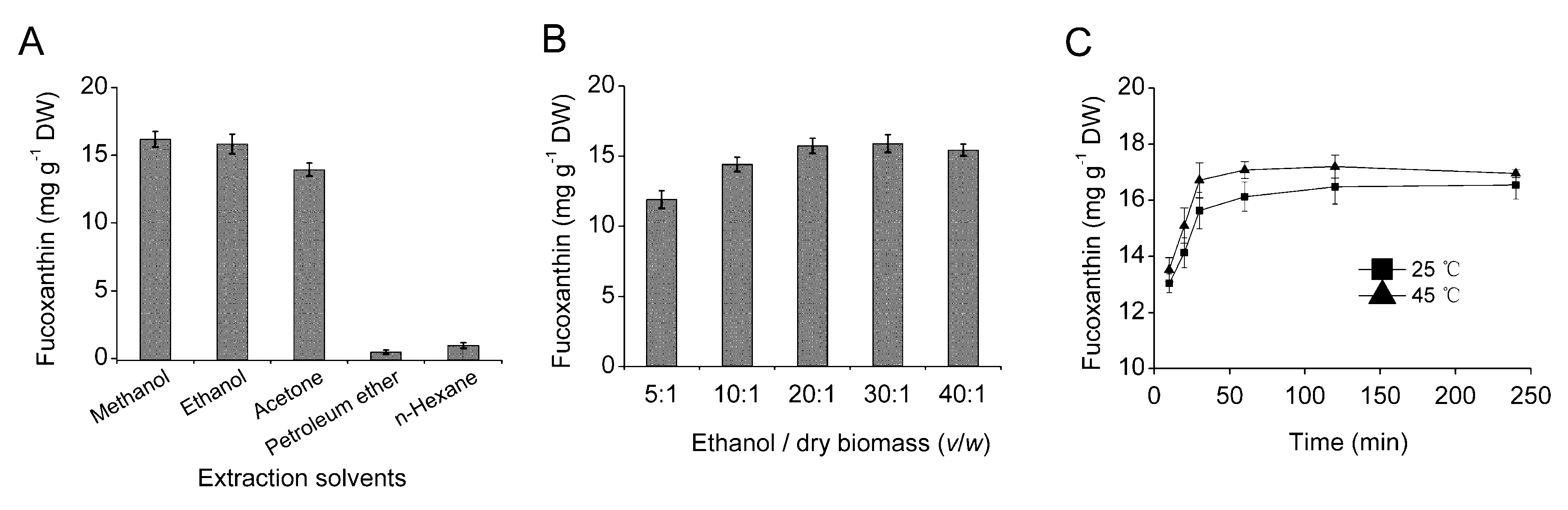
2.3. Optimization of Fucoxanthin Extraction Conditions
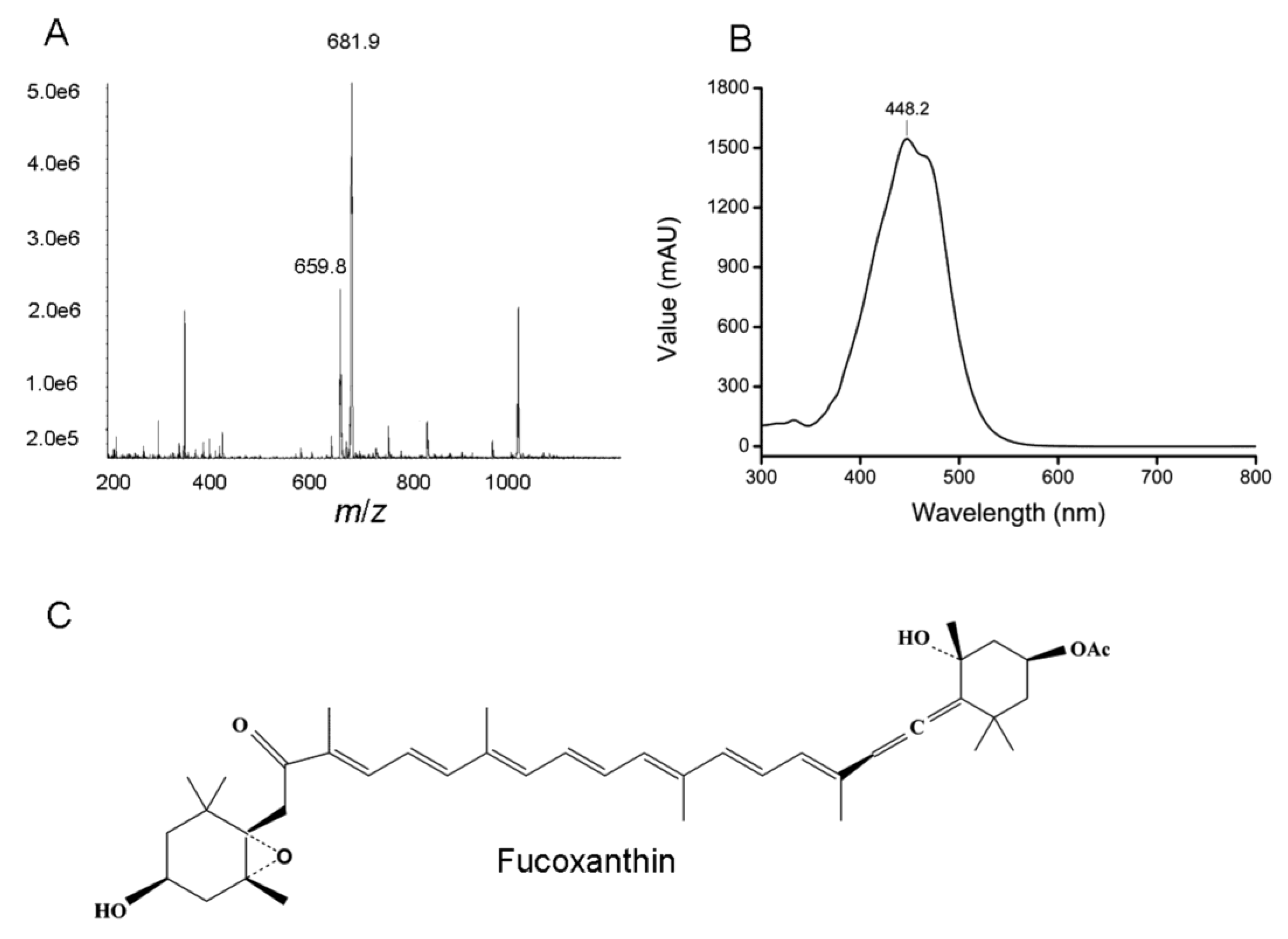
2.4. Purification and Identification of Fucoxanthin


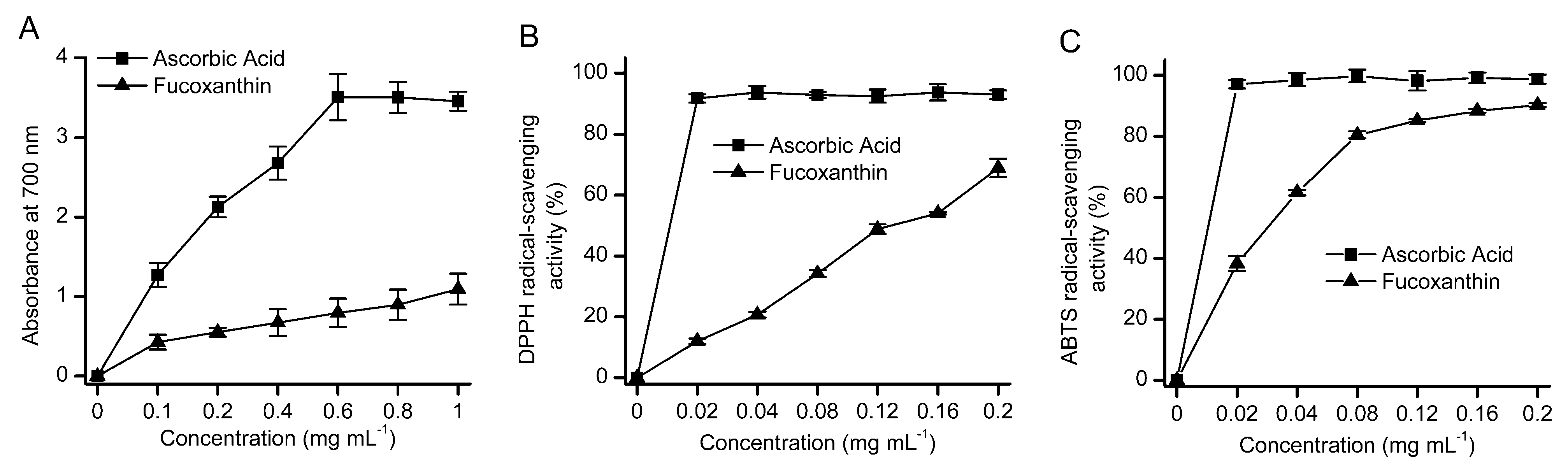
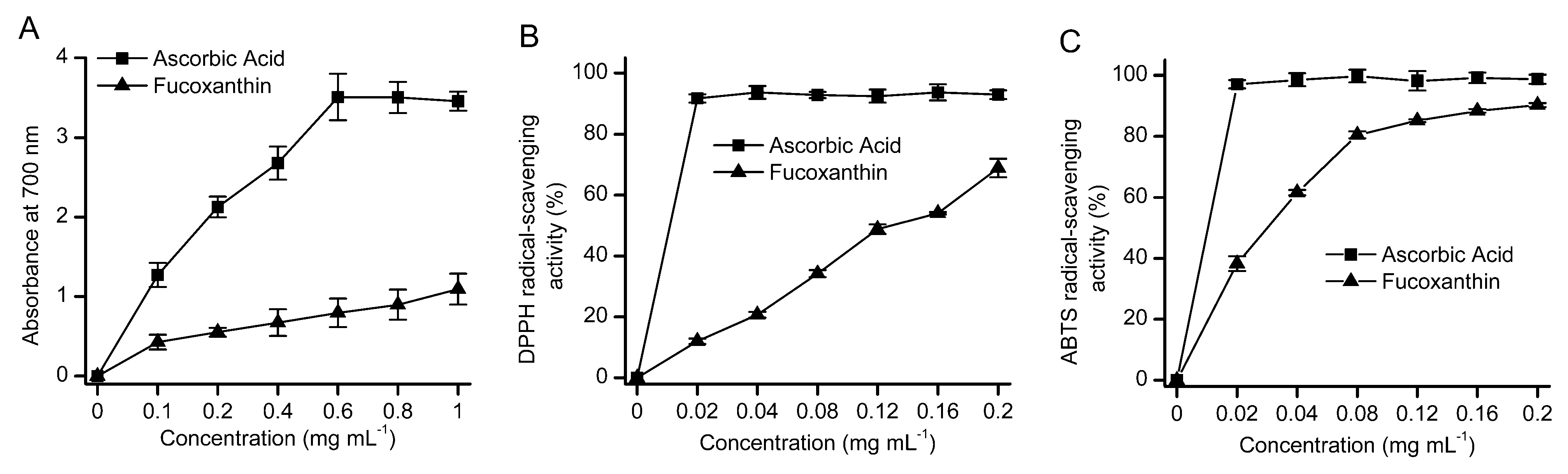
2.5. Antioxidant Activity
3. Experimental Section
3.1. Organism and Culture Conditions
3.2. Biomass Measurement
3.3. Pigment Extraction and Analysis
3.4. Purification of Fucoxanthin
3.5. HPLC and LC-MS Analysis
3.6. NMR Analysis
3.7. Assay for Antioxidant Activity
4. Conclusions
Acknowledgments
Conflict of Interest
References
- Owens, T.G.; Wold, E.R. Light-harvesting function in the diatom Phaeodactylum triconutum. Plant Physiol. 1986, 80, 732–738. [Google Scholar] [CrossRef]
- Abidov, M.; Ramazanov, Z.; Seifulla, R.; Grachev, S. The effects of Xanthigen™ in the weight management of obese premenopausal women with non-alcoholic fatty liver disease and normal liver fat. Diabetes Obes. Metab. 2010, 12, 72–81. [Google Scholar] [CrossRef]
- Woo, M.N.; Jeon, S.M.; Kim, H.J.; Lee, M.K.; Shin, S.K.; Shin, Y.C.; Park, Y.B.; Choi, M.S. Fucoxanthin supplementation improves plasma and hepatic lipid metabolism and blood glucose concentration in high-fat fed C57BL/6N mice. Chem. Biol. Interact. 2010, 186, 316–322. [Google Scholar] [CrossRef]
- Kawakami, T.; Tsushima, M.; Katabami, Y.; Mine, M.; Ishida, A.; Matsuno, T. Effect of β-carotene, β-echinenone, astaxanthin, fucoxanthin, vitamin A and vitamin E on the biological defense of the sea urchin Pseudocentrotus depresus. J. Exp. Mar. Biol. Ecol. 1998, 226, 165–174. [Google Scholar] [CrossRef]
- Kanazawa, K.; Ozaki, Y.; Hashimoto, T.; Das, S.K.; Matsushita, S.; Hirano, M.; Okada, T.; Komoto, A.; Mori, N.; Nakatsuka, M. Commercial-scale preparation of biofunctional fucoxanthin from waste parts of brown sea algae Laminalia japonica. Food Sci. Technol. Res. 2008, 14, 573–582. [Google Scholar] [CrossRef]
- Kim, S.M.; Shang, Y.F.; Um, B.H. A preparative method for isolation of fucoxanthin from Eisenia bicyclis by centrifugal partition chromatography. Phytochem. Anal. 2010, 22, 322–329. [Google Scholar]
- Kim, S.M.; Jung, Y.J.; Kwon, O.N.; Cha, K.H.; Um, B.H.; Chung, D.; Pan, C.H. A potential commercial source of fucoxanthin extracted from the microalga Phaeodactylum tricornutum. Appl. Biochem. Biotechnol. 2012, 166, 1843–1855. [Google Scholar] [CrossRef]
- Pulz, O.; Gross, W. Valuable products from biotechnology of microalgae. Appl. Microbiol. Biotechnol. 2004, 65, 635–648. [Google Scholar] [CrossRef]
- Werner, D. Contributions of the Study of Diatoms to Biology. In The Biology of Diatoms; Werner, D., Ed.; Blackwell: Oxford, UK, 1977; pp. 1–3. [Google Scholar]
- Ibanez, E.; Herrero, M.; Mendiola, J.A.; Castro-Puyana, M. Extraction and Characterization of Bioactive Compounds with Health Benefits from Marine Resources: Macro and Micro Algae, Cyanobacteria, and Invertebrates. In Marine Bioactive Compounds: Sources, Characterization and Applications; Hayes, M., Ed.; Springer-Verlag: New York, NY, USA, 2011; pp. 55–98. [Google Scholar]
- Drebes, D.; Schnepf, E. Gyrodinium undulans Hulburt, a marine dinoflagellate feeding on the bloom-forming diatom Odontella aurita, and on copepod and rotifer eggs. Helgol. Meeresunters. 1998, 52, 1–14. [Google Scholar] [CrossRef]
- De la Peña, M.R. Cell growth and nutritive value of the tropical benthic diatom, Amphora sp., at varying levels of nutrients and light intensity, and different culture locations. J. Appl. Phycol. 2007, 19, 647–655. [Google Scholar]
- Grobbelaar, J.U. Algal Nutrition. In Handbook of Microalgal Culture: Biotechnology and Applied Phycology; Richmond, A., Ed.; Blackwell Science Ltd.: Oxford, UK, 2004; pp. 97–115. [Google Scholar]
- Hu, Q. Environmental Effects on Cell Composition. In Handbook of Microalgal Culture: Biotechnology and Applied Phycology; Richmond, A., Ed.; Blackwell Science Ltd.: Oxford, UK, 2004; pp. 83–93. [Google Scholar]
- Harker, M.; Tsavalos, A.J.; Young, A.J. Factors responsible for astaxanthin formation in the Chlorophyte Haematococcus pluvialis. Bioresour. Technol. 1996, 55, 207–214. [Google Scholar] [CrossRef]
- Ben-Amotz, A. Dunaliella β-Carotene: From Science to Commerce. In Enigmatic Microorganisms and Life in Extreme Environments; Seckbach, J., Ed.; Kluwer: Dordrecht, Netherlands, 1999; pp. 401–410. [Google Scholar]
- Carreto, J.I.; Catoggio, J.A. Variations in pigment contents of the diatom Phaeodactylum tricornutum during growth. Mar. Biol. 1976, 36, 105–112. [Google Scholar] [CrossRef]
- Jin, E.; Polle, J.E.W.; Lee, H.K.; Hyum, S.M.; Chang, M. Xanthophylls in microalgae: From biosynthesis to biotechnological mass production and application. J. Microbiol. Biotechnol. 2003, 13, 165–174. [Google Scholar]
- Ben-Amotz, A.; Shaish, A. β-Carotene Bio-synthesis. In Dunaliella: Physiology, Biochemistry and Biotechnology; Avron, M., Ben-Amotz, A., Eds.; CRC Press: Boca Raton, FL, USA, 1992; pp. 206–216. [Google Scholar]
- Cysewski, G.R.; Todd-Lorenz, R. Industrial Production of Microalgal Cell-Mass and Secondary Products-Species of High Potential. In Handbook of Microalgal Culture: Biotechnology and Applied Phycology; Richmond, A., Ed.; Blackwell Publishing: Oxford, UK, 2004; pp. 281–288. [Google Scholar]
- Peng, J.; Yuan, J.P.; Wu, C.F.; Wang, J.H. Fucoxanthin, a marine carotenoid present in brown seaweeds and diatoms: Metabolism and bioactivities relevant to human health. Mar. Drugs 2011, 9, 1806–1828. [Google Scholar] [CrossRef]
- Xiao, X.; Si, X.; Yuan, Z.; Xu, X.; Li, G. Isolation of fucoxanthin from edible brown algae by microwave-assisted extraction coupled with high-speed countercurrent chromatography. J. Sep. Sci. 2012, 35, 2313–2317. [Google Scholar] [CrossRef]
- Mori, K.; Ooi, T.; Hiraoka, M.; Oka, N.; Hamada, H.; Tamura, M.; Kusumi, T. Fucoxanthin and its metabolites in edible brown algae cultivated in deep seawater. Mar. Drugs 2004, 2, 63–72. [Google Scholar] [CrossRef]
- Jaswir, I.; Noviendri, D.; Salleh, H.M.; Miyashita, K. Fucoxanthin extractions of brown seaweeds and analysis of their lipid fraction in methanol. Food Sci. Technol. Res. 2012, 2, 251–257. [Google Scholar]
- Jaswir, I.; Noviendri, D.; Salleh, H.M.; Taher, M.; Miyashita, K.; Ramli, N. Analysis of fucoxanthin content and purification of all-trans-fucoxanthin from Turbinaria turbinata and Sargassum plagyophyllum by SiO2 open column chromatography and reversed phase-HPLC. J. Liq. Chromatogr. Relat. Technol. 2012, 36, 1340–1354. [Google Scholar]
- Kim, S.M.; Kang, S.W.; Kwon, O.N.; Chung, D.; Pan, C.H. Fucoxanthin as a major carotenoid in Isochrysis aff. galbana: Characterization of extraction for commercial application. J. Korea Soc. Appl. Biol. Chem. 2012, 55, 477–483. [Google Scholar]
- Pasquet, V.; Cherouvrier, J.R.; Farhat, F.; Thiery, V.; Piot, J.M.; Berard, J.B. Study on the microalgal pigments extraction process: Performance of microwave assisted extraction. Process Biochem. 2011, 46, 59–67. [Google Scholar] [CrossRef]
- Stauber, J.L.; Jeffrey, S.W. Photosynthetic pigments in fifty-one species of marine diatoms. J. Phycol. 1988, 24, 158–172. [Google Scholar]
- Kim, K.N.; Heo, S.J.; Kang, S.M.; Ahn, G.; Jeon, Y.J. Fucoxanthin induces apoptosis in human leukemia HL-60 cells through a ROS-mediated Bcl-xL pathway. Toxicol. In Vitro 2010, 24, 1648–1654. [Google Scholar] [CrossRef]
- Klein, B.C.; Walter, C.; Lange, H.A.; Buchholz, R. Microalgae as natural sources for antoxidative compounds. J. Appl. Phycol. 2012, 24, 1133–1139. [Google Scholar] [CrossRef]
- Muller, L.; Frohlich, K.; Bohm, V. Comparative antioxidant activities of carotenoids measured by ferric reducing antioxidant power (FRAP), ABTS bleaching assay (αTEAC), DPPH assay and peroxyl radical scavenging assay. Food Chem. 2011, 129, 139–148. [Google Scholar] [CrossRef]
- Airanthi, M.W.A.; Hosokawa, M.; Miyashita, K. Comparative antioxidant activity of edible Japanese brown seaweeds. J. Food Sci. 2011, 76, 104–111. [Google Scholar] [CrossRef]
- Sachindra, N.M.; Sato, E.; Maeda, H.; Hosokawa, M.; Niwano, Y.; Kohno, M.; Miyashita, K. Radical scavenging and singlet oxygen quenching activity of marine carotenoid fucoxanthin and its metabolites. J. Agric. Food Chem. 2007, 55, 8516–8522. [Google Scholar] [CrossRef]
- Xia, S.; Li, A.F.; Wan, L.L.; Sang, M.; Zhang, C.W. Effects of nutrients and light on the growth and biochemical compositions of a marine microalga Odontella aurita. Chin. J. Oceanol. Limnol. 2013. [Google Scholar] [CrossRef]
- Li, Z.; Ma, X.Q.; LI, A.F.; Zhang, C.W. A novel potential source of β-carotene: Eustigmatos cf. polyphem (Eustigmatophyceae) and pilot β-carotene production in bubble column and flat panel photobioreactors. Bioresour. Technol. 2012, 117, 257–263. [Google Scholar]
- Deng, C.; Hu, Z.; Fu, H.; Hu, M.; Xu, X.; Chen, J. Chemical analysis and antioxidant activity in vitro of a β-d-glucan isolated from Dictyophora indusiata. Int. J. Biol. Macromol. 2012, 51, 70–75. [Google Scholar] [CrossRef]
- Osman, H.; Rahim, A.A.; Isa, N.M.; Bakhir, N.M. Antioxidant activity and phenolic content of Paederia foetida and Syzygium aqueum. Molecules 2009, 14, 970–978. [Google Scholar] [CrossRef]
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Xia, S.; Wang, K.; Wan, L.; Li, A.; Hu, Q.; Zhang, C. Production, Characterization, and Antioxidant Activity of Fucoxanthin from the Marine Diatom Odontella aurita. Mar. Drugs 2013, 11, 2667-2681. https://doi.org/10.3390/md11072667
Xia S, Wang K, Wan L, Li A, Hu Q, Zhang C. Production, Characterization, and Antioxidant Activity of Fucoxanthin from the Marine Diatom Odontella aurita. Marine Drugs. 2013; 11(7):2667-2681. https://doi.org/10.3390/md11072667
Chicago/Turabian StyleXia, Song, Ke Wang, Linglin Wan, Aifen Li, Qiang Hu, and Chengwu Zhang. 2013. "Production, Characterization, and Antioxidant Activity of Fucoxanthin from the Marine Diatom Odontella aurita" Marine Drugs 11, no. 7: 2667-2681. https://doi.org/10.3390/md11072667