Proteomic Investigation of the Sinulariolide-Treated Melanoma Cells A375: Effects on the Cell Apoptosis through Mitochondrial-Related Pathway and Activation of Caspase Cascade

 ,
,
Abstract
:1. Introduction
2. Results
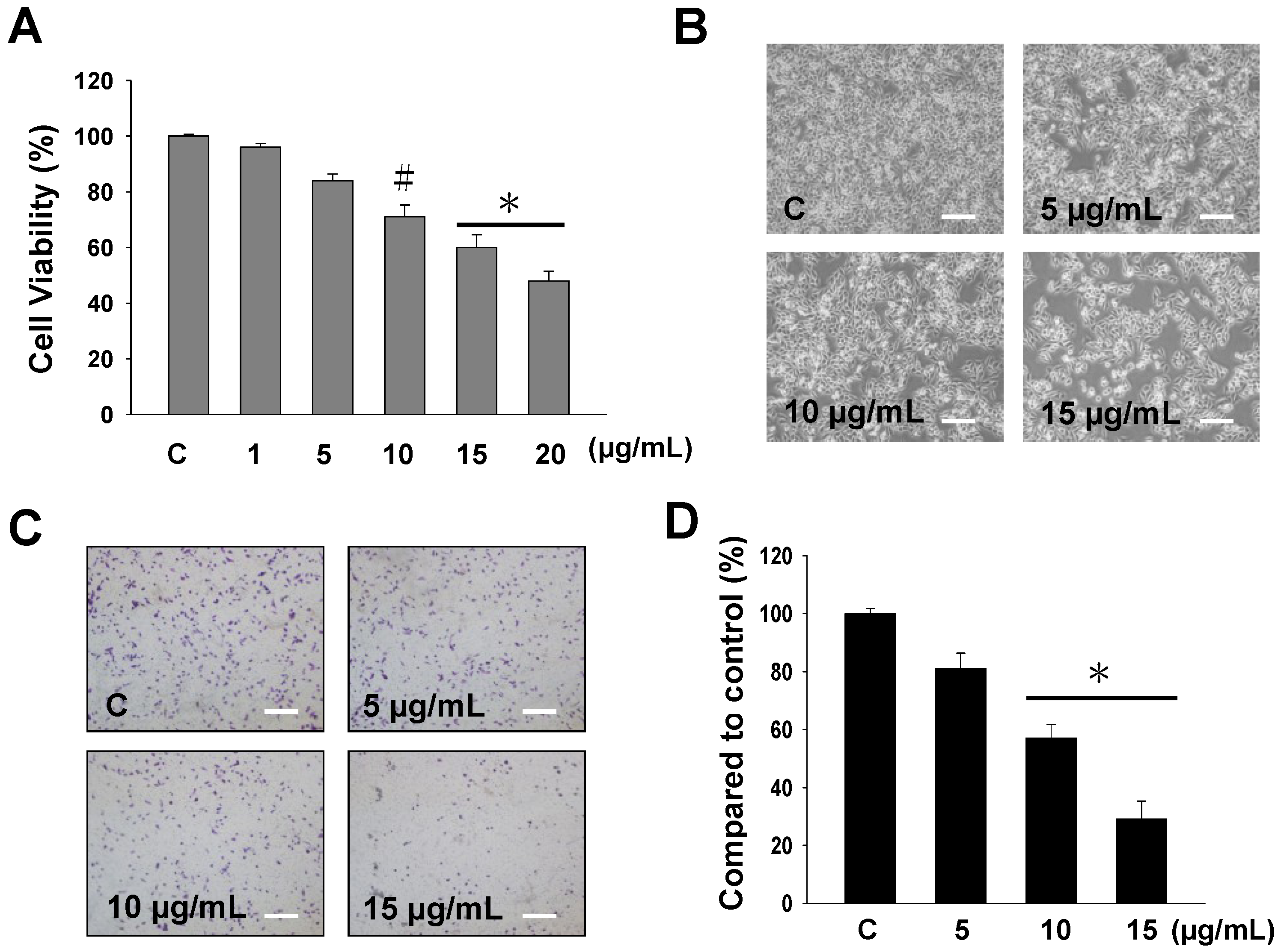
2.1. The Cytotoxicity and Anti-Migratory Effects of Sinulariolide on A375 Cells
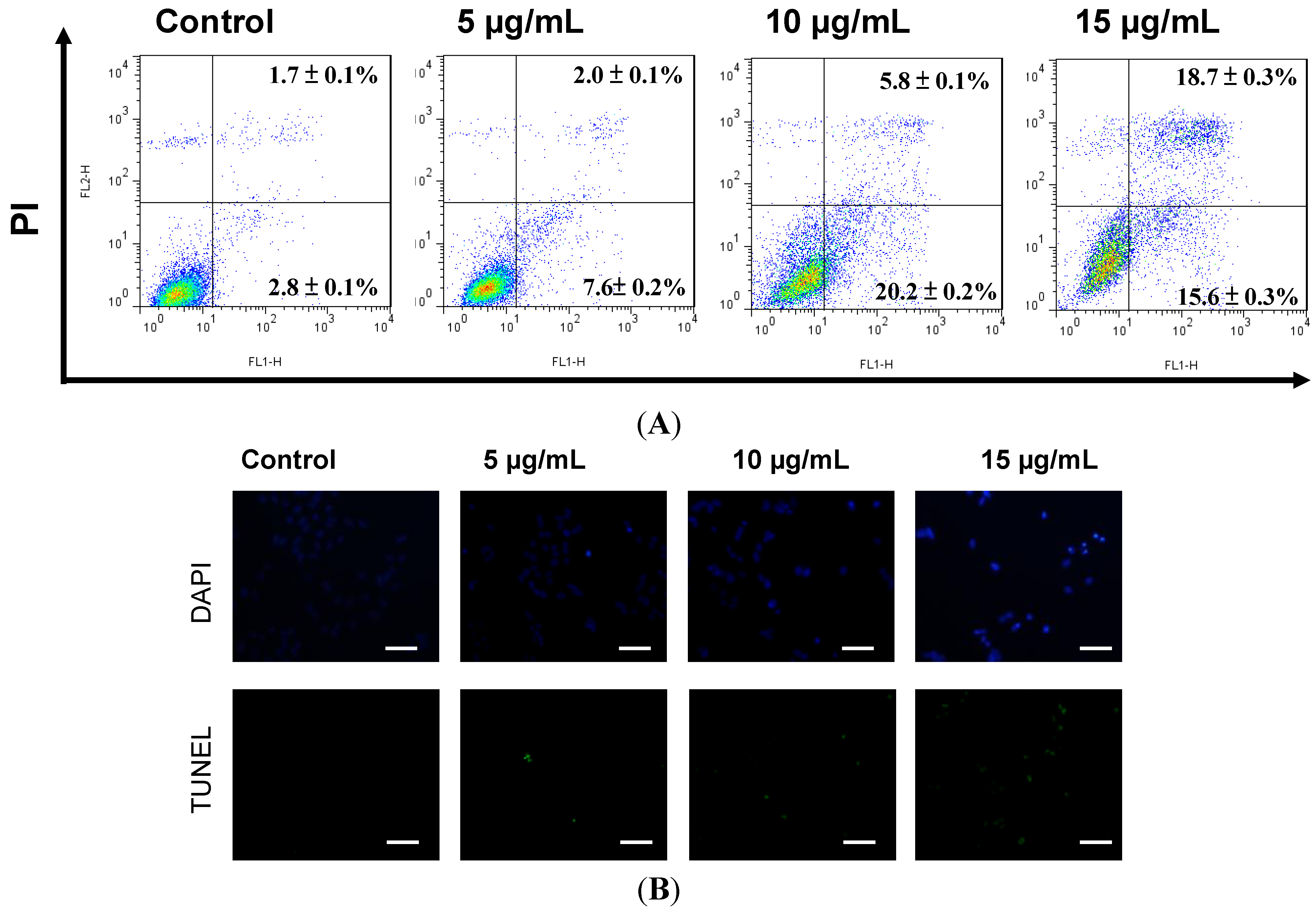
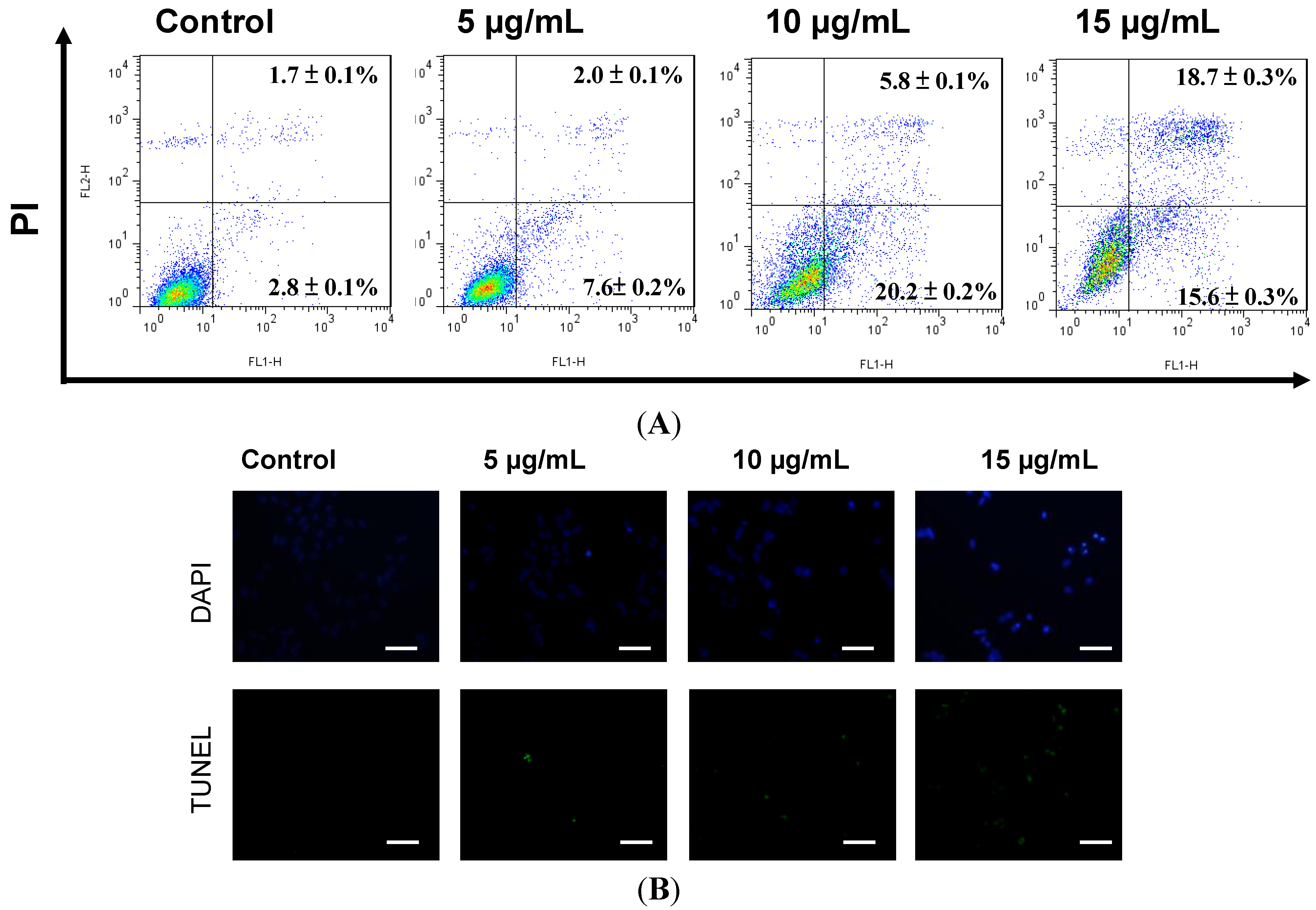
2.2. Apoptosis Was Triggered by Sinulariolide in A375 Cells
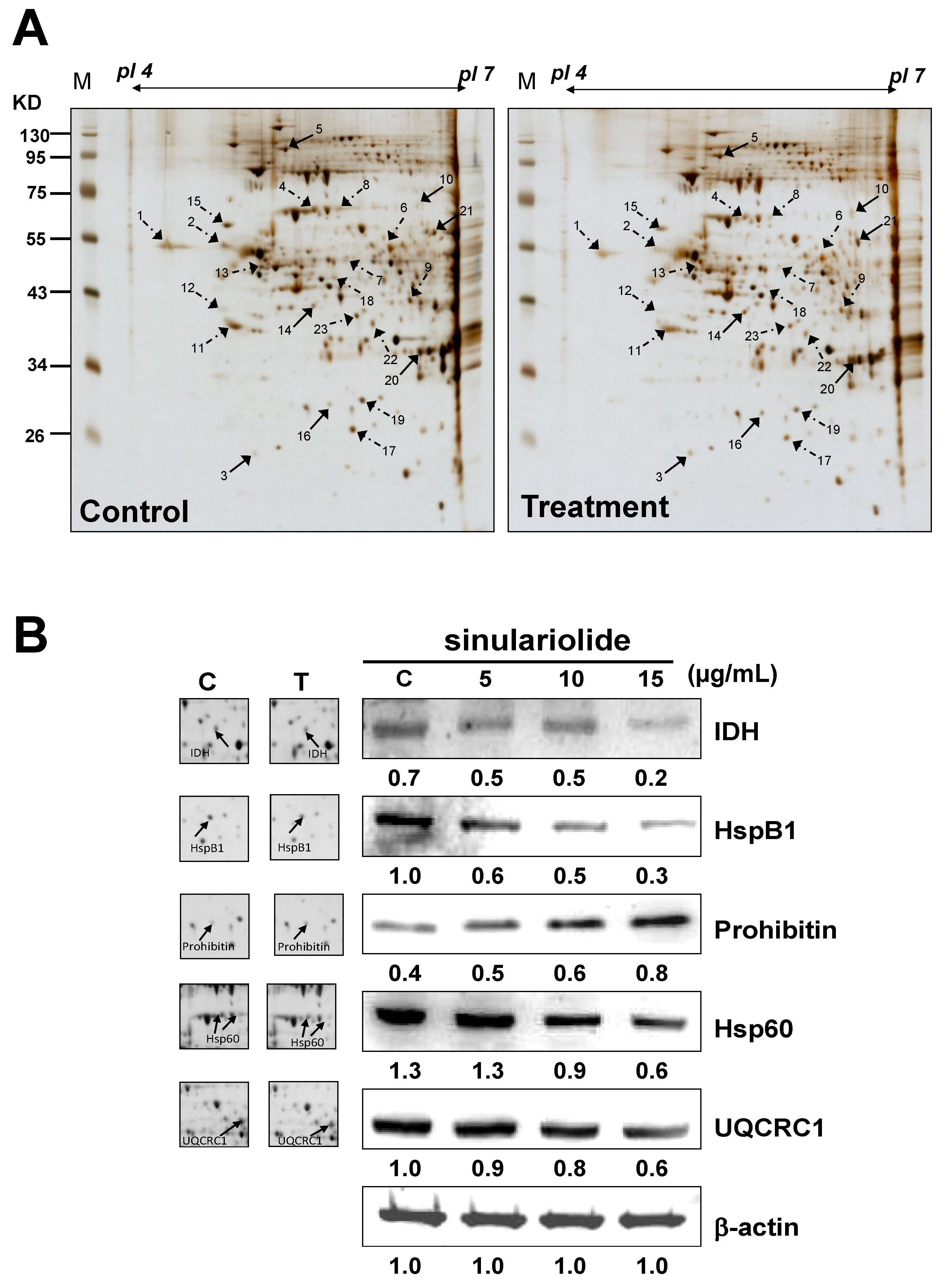
2.3. Proteomic Analysis of A375 Cells Treated with Sinulariolide
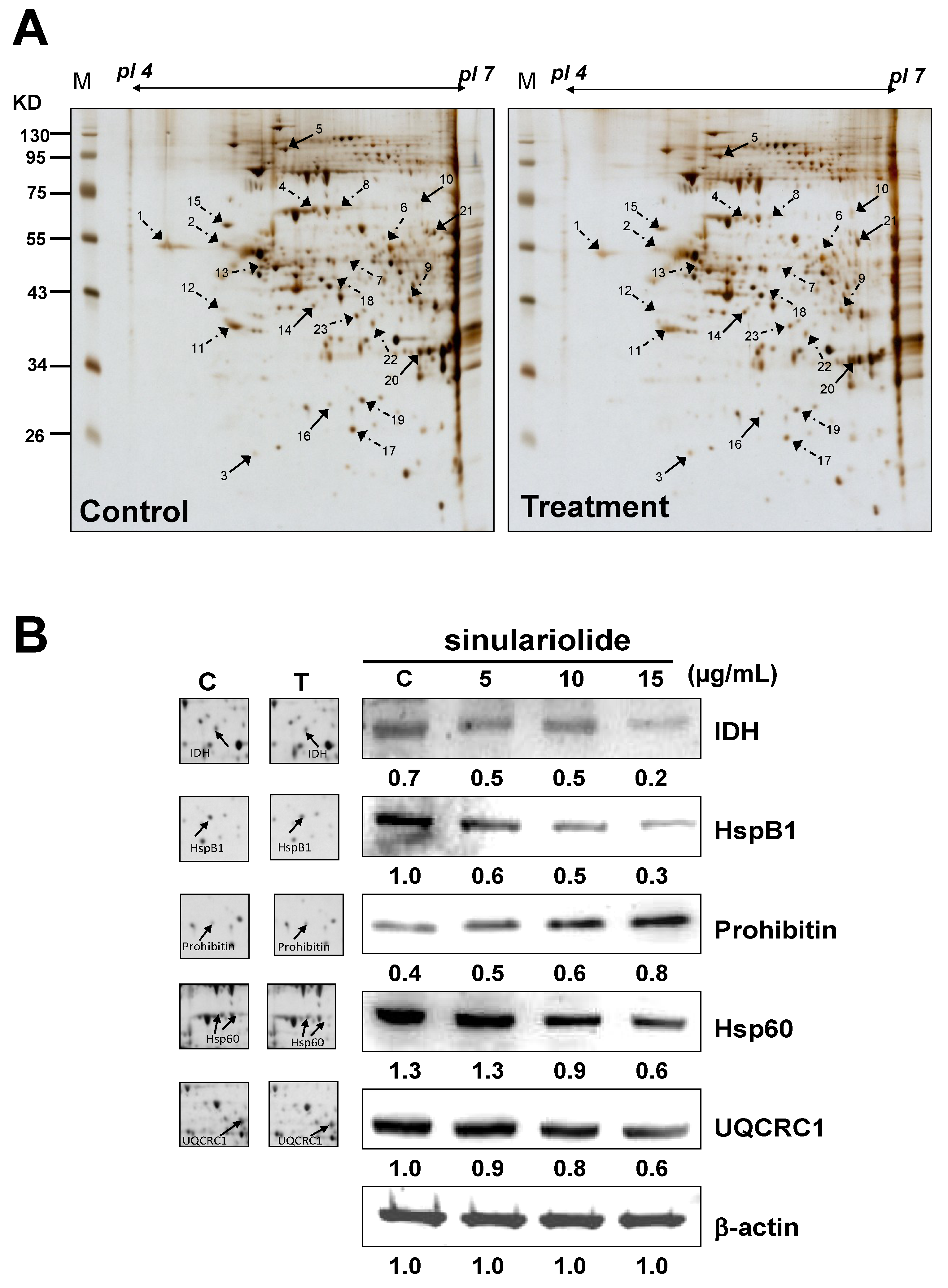

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Spot No. | Protein name | Accession No. | Calculated Mw/pI | Peptide matched | Sequence covered % | MASCOT score | Regulation (fold change) * |
|---|---|---|---|---|---|---|---|
| 1 | Calreticulin precursor | P27797 | 48.11/4.29 | 35 | 40 | 251 | −1.6 |
| 2 | Vimentin | P08670 | 53.61/5.06 | 26 | 39 | 272 | −2.2 |
| 3 | Rho GDP-dissociation inhibitor 1 (Rho-GDI) | P52565 | 23.19/5.02 | 5 | 26 | 68 | +2.3 |
| 4 | 60 kDa heat shock protein (Hsp60) | P10809 | 61.01/5.7 | 51 | 47 | 545 | −2.6 |
| 5 | Transitional endoplasmic reticulum ATPase | P55072 | 89.26/5.14 | 49 | 47 | 439 | +1.8 |
| 6 | Ubiquinol cytochrome c reductase complex core protein 1 (UQCRC1) | P31930 | 52.61/5.94 | 5 | 10 | 53 | −1.9 |
| 7 | Glutathione synthetase | P48637 | 52.35/5.67 | 5 | 10 | 123 | −2.8 |
| 8 | 60 kDa heat shock protein (Hsp60) | P10809 | 61.01/5.7 | 3 | 8 | 66 | −2.5 |
| 9 | DnaJ homolog subfamily B member 11 precursor | Q9UBS4 | 40.48/5.81 | 3 | 9 | 40 | −2.2 |
| 10 | Lamin-A/C (70 kDa lamin) | P02545 | 74.09/6.57 | 3 | 10 | 71 | +1.8 |
| 11 | Nucleophosmin (NPM) | P06748 | 32.55/4.64 | 28 | 39 | 216 | −1.9 |
| 12 | Reticulocalbin-1 precursor | Q15293 | 38.86/4.86 | 4 | 9 | 84 | −2.4 |
| 13 | Vimentin | P08670 | 53.61/5.06 | 7 | 8 | 101 | −1.7 |
| 14 | Stomatin-like protein 2 | Q9UJZ1 | 38.51/6.88 | 29 | 46 | 331 | +2.0 |
| 15 | Protein disulfide-isomerase precursor (PDI) | P07237 | 57.08/4.76 | 23 | 35 | 205 | −2.1 |
| 16 | Prohibitin | P35232 | 29.78/5.57 | 45 | 67 | 660 | +1.8 |
| 17 | Platelet-activating factor acetylhydrolase IB subunit beta | P68402 | 25.55/5.57 | 4 | 9 | 46 | −2.2 |
| 18 | Heterogeneous nuclear ribonucleoprotein F | P52597 | 45.64/5.38 | 8 | 21 | 121 | −1.8 |
| 19 | Heat shock protein beta-1 (HspB1) | P04792 | 22.76/5.98 | 9 | 15 | 102 | −2.6 |
| 20 | Translation initiation factor eIF-2B subunit alpha | Q14232 | 33.69/6.9 | 1 | 4 | 47 | +2.5 |
| 21 | Retinal dehydrogenase 2 | O94788 | 56.68/5.79 | 3 | 2 | 54 | +2.3 |
| 22 | Isocitrate dehydrogenase (NAD) subunit alpha (IDH) | P50213 | 39.56/6.47 | 10 | 17 | 148 | −1.9 |
| 23 | Pyruvate dehydrogenase E1 component subunit beta | P11177 | 39.19/6.2 | 3 | 10 | 68 | −1.5 |
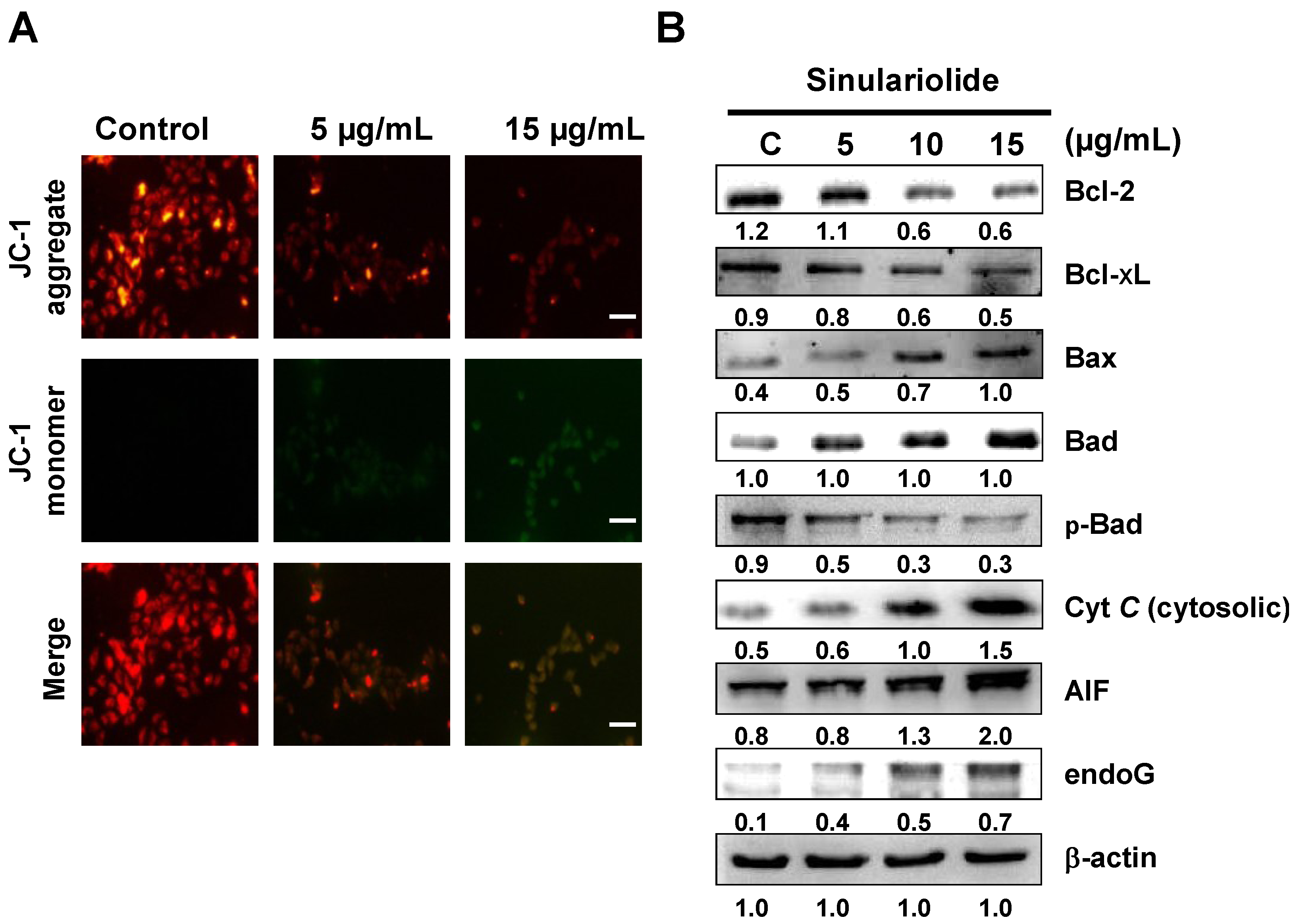
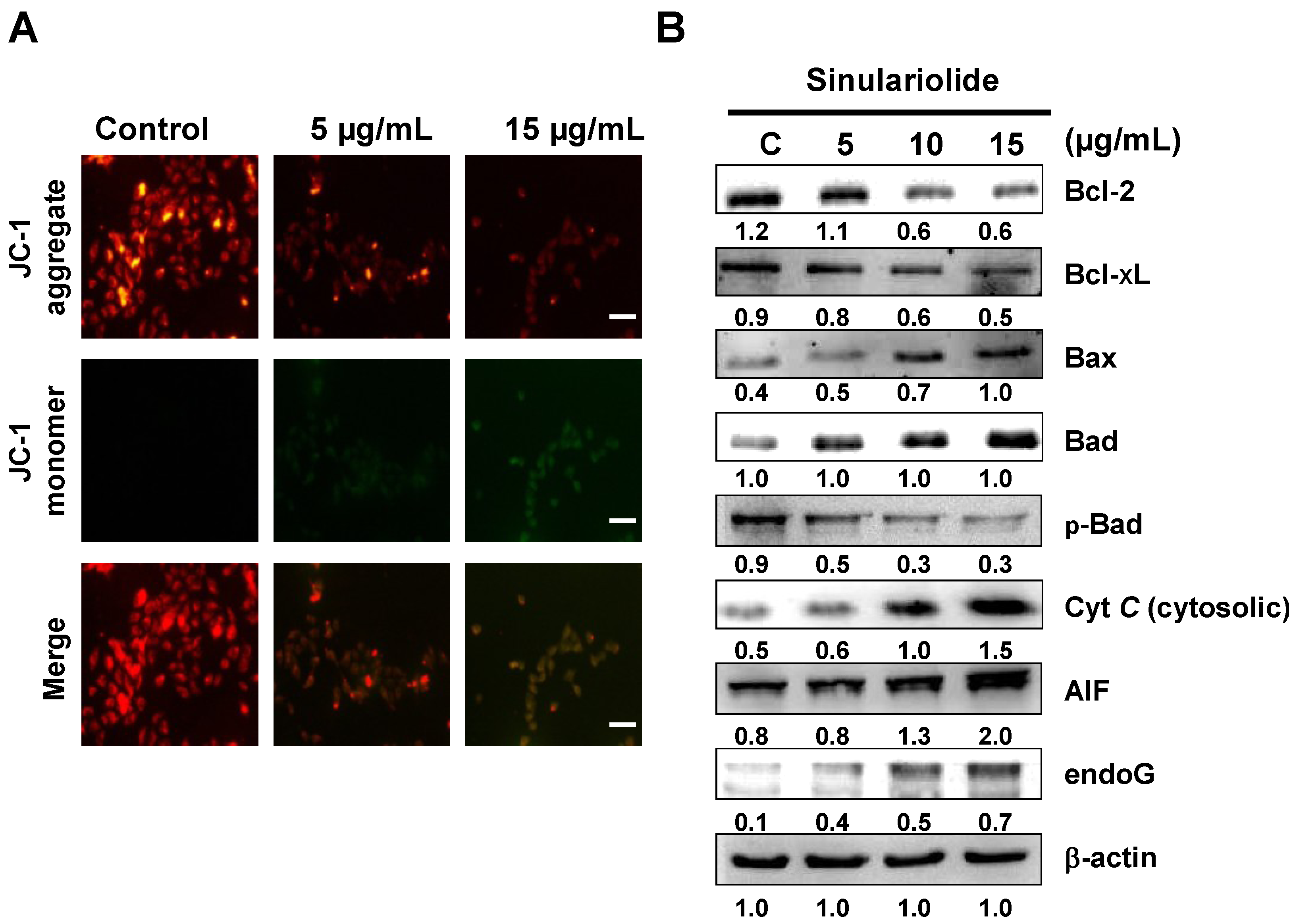
2.4. Sinulariolide Induced Mitochondrial Depolarization and Activated the Mitochondrial-Related Pathway, Resulting in Cell Apoptosis
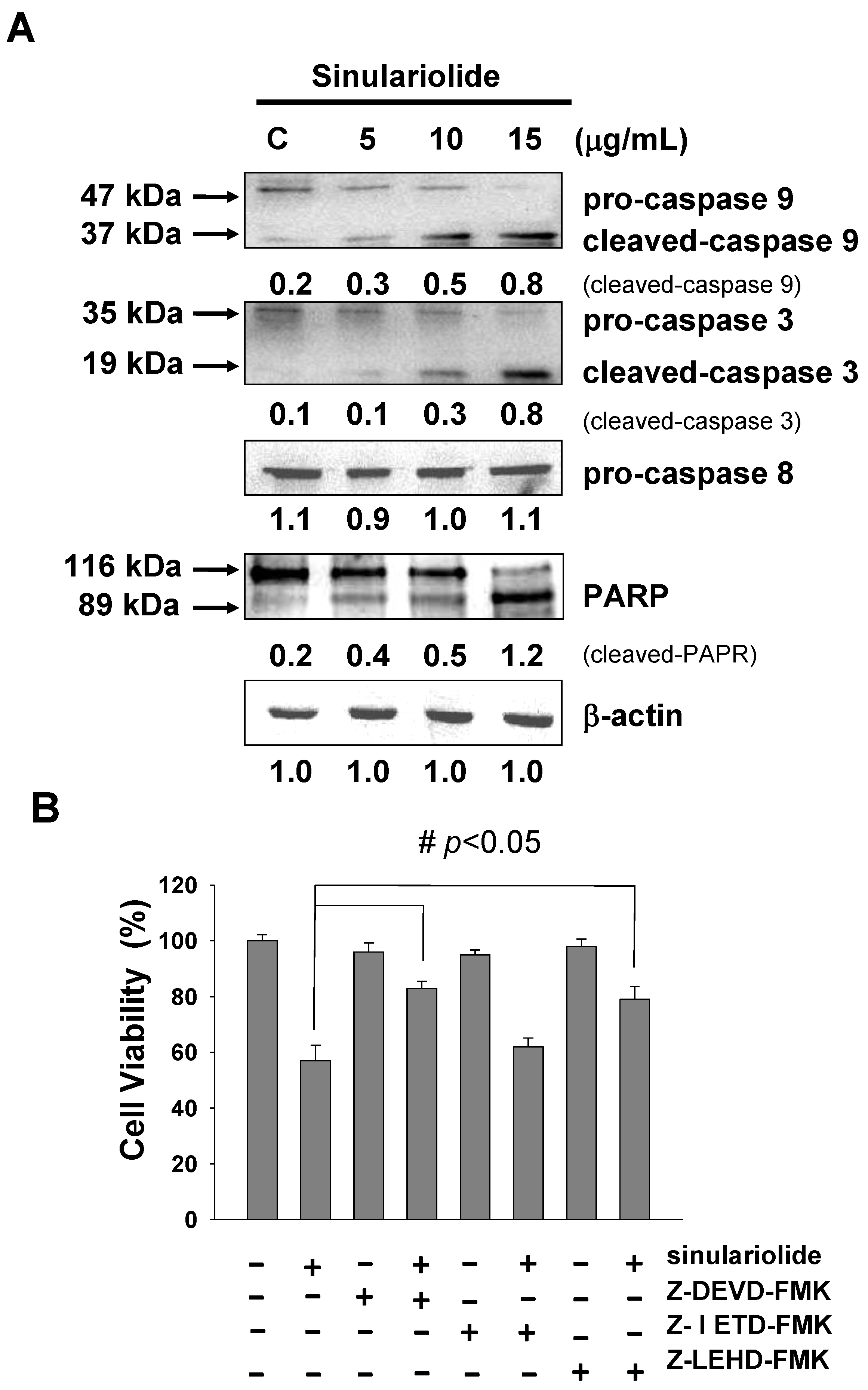
2.5. Sinulariolide Activates the Caspase-Dependent Pathway, Leading to Cell Apoptosis
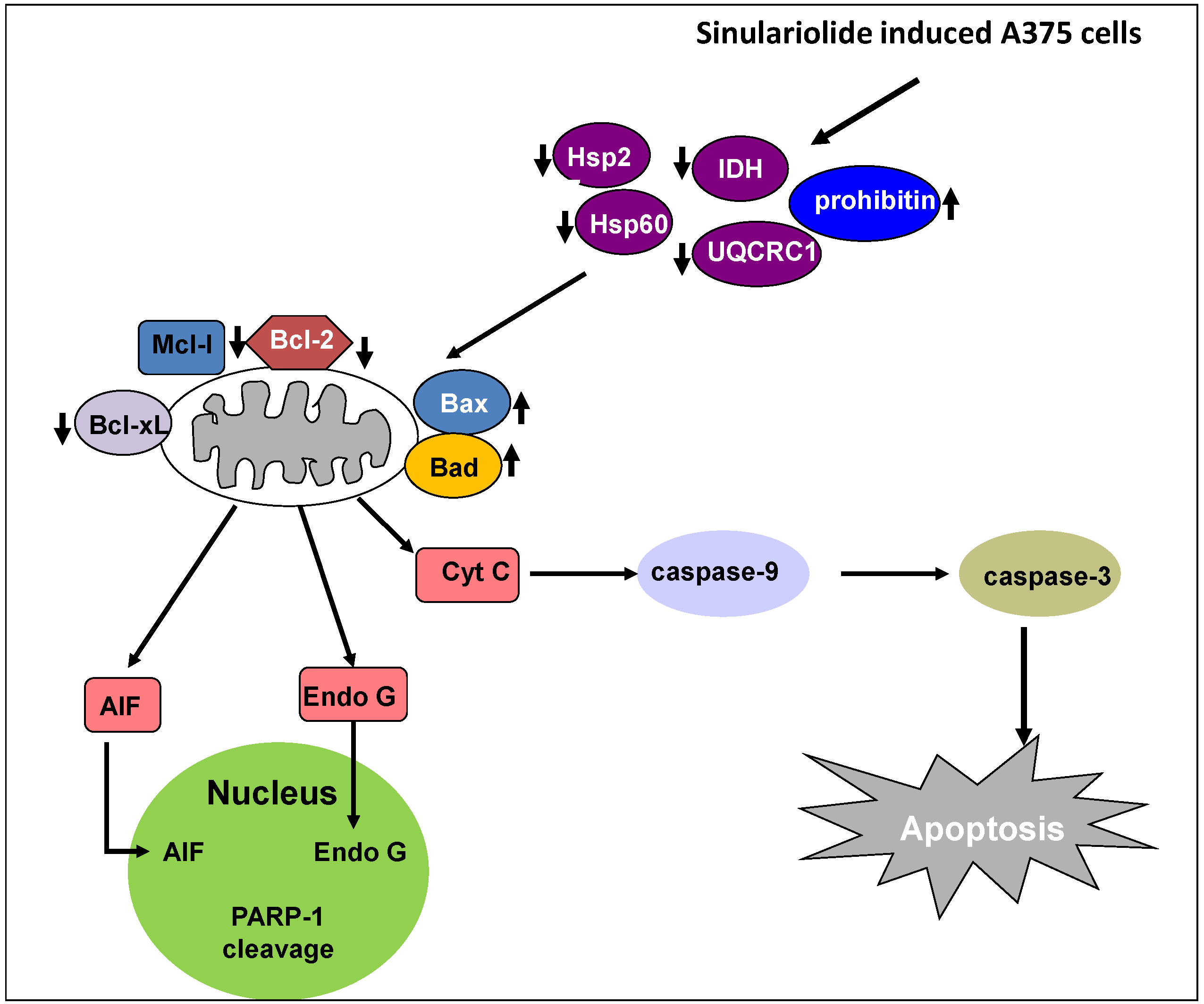
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Cell Culture
4.3. Determination of Cell Viability
4.4. Cell Migration Assay
4.5. Quantitative Detection of Apoptosis by Flow Cytometry
4.6. Protein Extraction and Concentration Estimation
4.7. Two-Dimensional Gel Electrophoresis, Image Analysis and Protein Identification by LC-MS/MS
4.8. DAPI and TUNEL Stain
4.9. Mitochondria and Cytosol Fractionation
4.10. Assessment of Mitochondrial Membrane Potential (ΔΨm)
4.11. Western Blot Analysis
4.12. Statistical Analysis
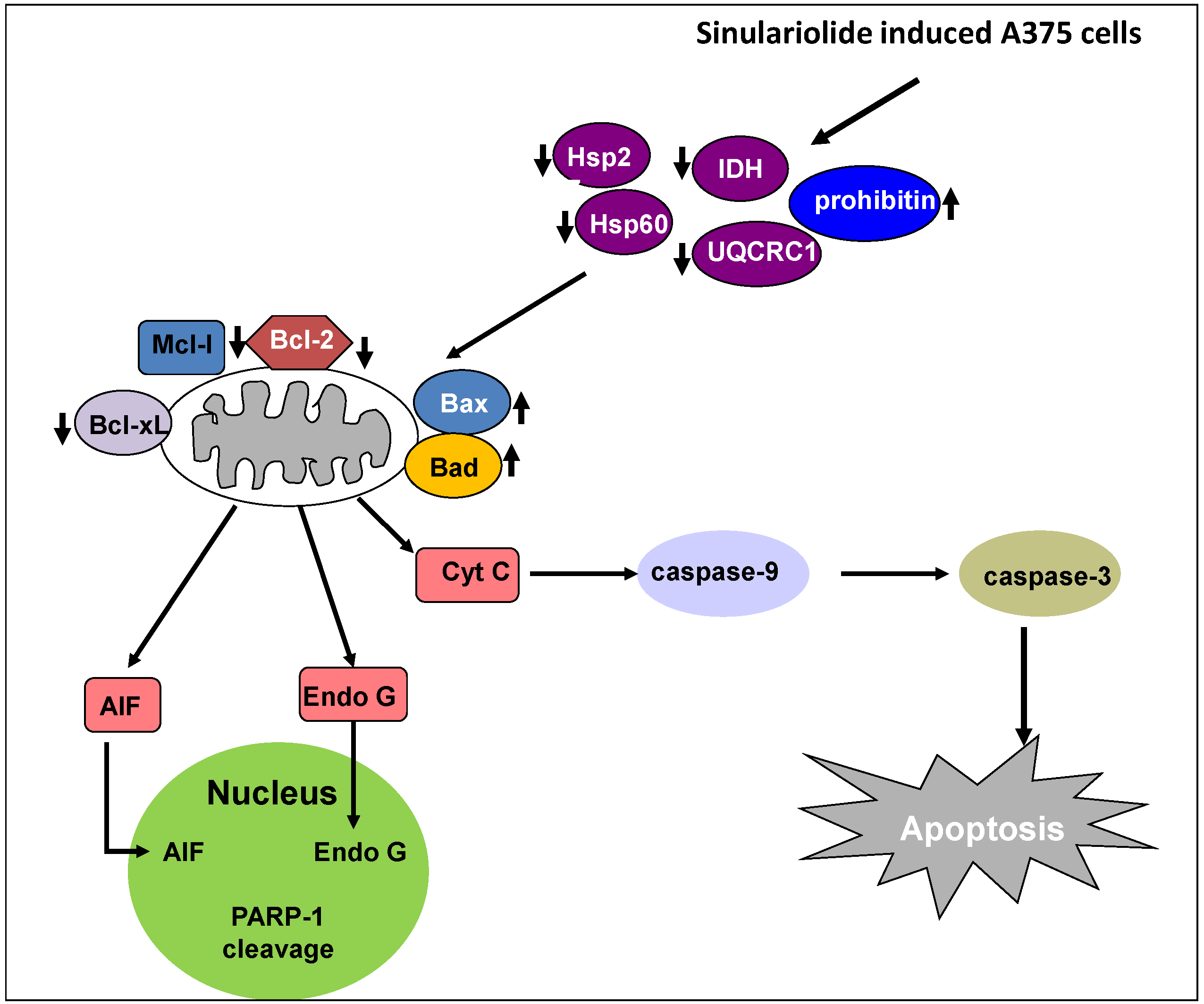
5. Conclusions
Acknowledgements
Conflict of Interest
References
- Parkin, D.M.; Bray, F.; Ferlay, J.; Pisani, P. Global cancer statistics, 2002. CA Cancer J. Clin. 2005, 55, 74–108. [Google Scholar] [CrossRef]
- Davar, D.; Tarhini, A.A.; Kirkwood, J.M. Adjuvant therapy for melanoma. Cancer J. 2012, 18, 192–202. [Google Scholar] [CrossRef]
- Tawbi, H.A.; Buch, S.C. Chemotherapy resistance abrogation in metastatic melanoma. Clin. Adv. Hematol. Oncol. 2010, 8, 259–266. [Google Scholar]
- Sasse, A.D.; Sasse, E.C.; Clark, L.G.; Ulloa, L.; Clark, O.A. Chemoimmunotherapy versus chemotherapy for metastatic malignant melanoma. Cochrane Database Syst. Rev. 2007, 1. [Google Scholar] [CrossRef]
- Fuglede, N.B.; Brinck-Claussen, U.O.; Deltour, I.; Boesen, E.H.; Dalton, S.O.; Johansen, C. Incidence of cutaneous malignant melanoma in Denmark, 1978–2007. Br. J. Dermatol. 2011, 165, 349–353. [Google Scholar] [CrossRef]
- Essner, R. Surgical treatment of malignant melanoma. Surg. Clin. North Am. 2003, 83, 109–156. [Google Scholar] [CrossRef]
- Chang, J.W. Cutaneous melanoma: Taiwan experience and literature review. Chang Gung Med. J. 2010, 33, 602–612. [Google Scholar]
- Treisman, J.; Garlie, N. Systemic therapy for cutaneous melanoma. Clin. Plast. Surg. 2010, 37, 127–146. [Google Scholar] [CrossRef]
- Bleehen, N.M.; Newlands, E.S.; Lee, S.M.; Thatcher, N.; Selby, P.; Calvert, A.H.; Rustin, G.J.; Brampton, M.; Stevens, M.F. Cancer research campaign phase II trial of temozolomide in metastatic melanoma. J. Clin. Oncol. 1995, 13, 910–913. [Google Scholar]
- Ibrahim, N.; Haluska, F.G. Molecular pathogenesis of cutaneous melanocytic neoplasms. Annu. Rev. Pathol. 2009, 4, 551–579. [Google Scholar] [CrossRef]
- Acquavella, N.; Kluger, H.; Rhee, J.; Farber, L.; Tara, H.; Ariyan, S.; Narayan, D.; Kelly, W.; Sznol, M. Toxicity and activity of a twice daily high-dose bolus interleukin 2 regimen in patients with metastatic melanoma and metastatic renal cell cancer. J. Immunother. 2008, 31, 569–576. [Google Scholar] [CrossRef]
- Liu, C.I.; Wang, R.Y.; Lin, J.J.; Su, J.H.; Chiu, C.C.; Chen, J.C.; Chen, J.Y.; Wu, Y.J. Proteomic profiling of the 11-dehydrosinulariolide-treated oral carcinoma cells Ca9–22: Effects on the cell apoptosis through mitochondrial-related and ER stress pathway. J. Proteomics 2012, 75, 5578–5589. [Google Scholar] [CrossRef]
- Su, C.C.; Su, J.H.; Lin, J.J.; Chen, C.C.; Hwang, W.I.; Huang, H.H.; Wu, Y.J. An investigation into the cytotoxic effects of 13-acetoxysarcocrassolide from the soft coral sarcophyton crassocaule on bladder cancer cells. Mar. Drugs 2011, 9, 2622–2642. [Google Scholar] [CrossRef]
- Su, T.R.; Lin, J.J.; Chiu, C.C.; Chen, J.Y.; Su, J.H.; Cheng, Z.J.; Hwang, W.I.; Huang, H.H.; Wu, Y.J. Proteomic investigation of anti-tumor activities exerted by sinularin against A2058 melanoma cells. Electrophoresis 2012, 33, 1139–1152. [Google Scholar] [CrossRef]
- Kondratyuk, T.P.; Park, E.J.; Yu, R.; van Breemen, R.B.; Asolkar, R.N.; Murphy, B.T.; Fenical, W.; Pezzuto, J.M. Novel marine phenazines as potential cancer chemopreventive and anti-inflammatory agents. Mar. Drugs 2012, 10, 451–464. [Google Scholar] [CrossRef]
- Maoka, T.; Tokuda, H.; Suzuki, N.; Kato, H.; Etoh, H. Anti-oxidative, anti-tumor-promoting, and anti-carcinogensis activities of nitroastaxanthin and nitrolutein, the reaction products of astaxanthin and lutein with peroxynitrie. Mar. Drugs 2012, 10, 1391–1399. [Google Scholar] [CrossRef]
- Rabelo, L.; Monteiro, N.; Serquiz, R.; Santos, P.; Oliveira, R.; Oliveira, A.; Rocha, H.; Morais, A.H.; Uchoa, A.; Santos, E. A lactose-binding lectin from the marine sponge Cinachyrella apion (Cal) induces cell death in human cervical adenocarcinoma cells. Mar. Drugs 2012, 10, 727–743. [Google Scholar] [CrossRef]
- Neoh, C.A.; Wang, R.Y.; Din, Z.H.; Su, J.H.; Chen, Y.K.; Tsai, F.J.; Weng, S.H.; Wu, Y.J. Induction of apoptosis by sinulariolide from soft coral through mitochondrial-related and p38MAPK pathways on human bladder carcinoma cells. Mar. Drugs 2012, 10, 2893–2911. [Google Scholar] [CrossRef]
- Aceret, T.L.; Coll, J.C.; Uchio, Y.; Sammarco, P.W. Antimicrobial activity of the diterpenes flexibilide and sinulariolide derived from Sinularia flexibilis Quoy and Gaimard 1833 (Coelenterata: Alcyonacea, Octocorallia). Comp. Biochem. Physiol. C 1998, 120, 121–126. [Google Scholar]
- Ghosh, J.C.; Dohi, T.; Kang, B.H.; Altieri, D.C. Hsp60 regulation of tumor cell apoptosis. J. Biol. Chem. 2008, 283, 5188–5194. [Google Scholar]
- Gibert, B.; Hadchity, E.; Czekalla, A.; Aloy, M.T.; Colas, P.; Rodriguez-Lafrasse, C.; Arrigo, A.P.; Diaz-Latoud, C. Inhibition of heat shock protein 27 (HspB1) tumorigenic functions by peptide aptamers. Oncogene 2011, 30, 3672–3681. [Google Scholar] [CrossRef]
- Kanagasabai, R.; Krishnamurthy, K.; Druhan, L.J.; Ilangovan, G. Forced expression of heat shock protein 27 (Hsp27) reverses P-glycoprotein (ABCB1)-mediated drug efflux and MDR1 gene expression in Adriamycin-resistant human breast cancer cells. J. Biol. Chem. 2011, 286, 33289–33300. [Google Scholar] [CrossRef]
- Liu, C.I.; Chen, C.C.; Chen, J.C.; Su, J.H.; Huang, H.H.; Chen, J.Y.; Wu, Y.J. Proteomic analysis of anti-tumor effects of 11-dehydrosinulariolide on CAL-27 cells. Mar. Drugs 2011, 9, 1254–1272. [Google Scholar] [CrossRef]
- Winter, A.; Kamarainen, O.; Hofmann, A. Molecular modeling of prohibitin domains. Proteins 2007, 68, 353–362. [Google Scholar] [CrossRef]
- Nijtmans, L.G.; de Jong, L.; Artal Sanz, M.; Coates, P.J.; Berden, J.A.; Back, J.W.; Muijsers, A.O.; van der Spek, H.; Grivell, L.A. Prohibitins act as a membrane-bound chaperone for the stabilization of mitochondrial proteins. EMBO J. 2000, 19, 2444–2451. [Google Scholar] [CrossRef]
- Fusaro, G.; Dasgupta, P.; Rastogi, S.; Joshi, B.; Chellappan, S. Prohibitin induces the transcriptional activity of p53 and is exported from the nucleus upon apoptotic signaling. J. Biol. Chem. 2003, 278, 47853–47861. [Google Scholar] [CrossRef]
- Rajalingam, K.; Wunder, C.; Brinkmann, V.; Churin, Y.; Hekman, M.; Sievers, C.; Rapp, U.R.; Rudel, T. Prohibitin is required for Ras-induced Raf-MEK-ERK activation and epithelial cell migration. Nat. Cell Biol. 2005, 7, 837–843. [Google Scholar] [CrossRef]
- Xu, S.G.; Yan, P.J.; Shao, Z.M. Differential proteomic analysis of a highly metastatic variant of human breast cancer cells using two-dimensional differential gel electrophoresis. J. Cancer Res. Clin. Oncol. 2010, 136, 1545–1556. [Google Scholar] [CrossRef]
- Jung, K.H.; Park, J.W. Suppression of mitochondrial NADP(+)-dependent isocitrate dehydrogenase activity enhances curcumin-induced apoptosis in HCT116 cells. Free Radic. Res. 2011, 45, 431–438. [Google Scholar] [CrossRef]
- Tuquet, C.; Dupont, J.; Mesneau, A.; Roussaux, J. Effects of tamoxifen on the electron transport chain of isolated rat liver mitochondria. Cell Biol. Toxicol. 2000, 16, 207–219. [Google Scholar] [CrossRef]
- Denicourt, C.; Dowdy, S.F. Targeting apoptotic pathways in cancer cells. Science 2004, 305, 1411–1413. [Google Scholar] [CrossRef]
- Liao, C.T.; Chang, J.T.; Wang, H.M.; Ng, S.H.; Hsueh, C.; Lee, L.Y.; Lin, C.H.; Chen, I.H.; Huang, S.F.; Cheng, A.J.; et al. Analysis of risk factors of predictive local tumor control in oral cavity cancer. Ann. Surg. Oncol. 2008, 15, 915–922. [Google Scholar] [CrossRef]
- Nicholson, D.W.; Thornberry, N.A. Life and death decisions. Science 2003, 299, 214–215. [Google Scholar] [CrossRef]
- Amarante-Mendes, G.P.; Naekyung Kim, C.; Liu, L.; Huang, Y.; Perkins, C.L.; Green, D.R.; Bhalla, K. Bcr-Abl exerts its antiapoptotic effect against diverse apoptotic stimuli through blockage of mitochondrial release of cytochrome c and activation of caspase-3. Blood 1998, 91, 1700–1705. [Google Scholar]
- Hsieh, P.W.; Chang, F.R.; McPhail, A.T.; Lee, K.H.; Wu, Y.C. New cembranolide analogues from the formosan soft coral Sinularia flexibilis and their cytotoxicity. Nat. Prod. Res. 2003, 17, 409–418. [Google Scholar] [CrossRef]
- Su, T.R.; Tsai, F.J.; Lin, J.J.; Huang, H.H.; Chiu, C.C.; Su, J.H.; Yang, Y.T.; Chen, J.Y.; Wong, B.S.; Wu, Y.J. Induction of apoptosis by 11-dehydrosinulariolide via mitochondrial dysregulation and ER stress pathways in human melanoma cells. Mar. Drugs 2012, 10, 1883–1898. [Google Scholar] [CrossRef]
- Chiu, C.C.; Chen, J.Y.; Lin, K.L.; Huang, C.J.; Lee, J.C.; Chen, B.H.; Chen, W.Y.; Lo, Y.H.; Chen, Y.L.; Tseng, C.H.; Lin, S.R. p38 MAPK and NF-kappaB pathways are involved in naphtho(1,2-b)furan-4,5-dione induced anti-proliferation and apoptosis of human hepatoma cells. Cancer Lett. 2010, 295, 92–99. [Google Scholar] [CrossRef]
- Lu, C.M.; Wu, Y.J.; Chen, C.C.; Hsu, J.L.; Chen, J.C.; Chen, J.Y.; Huang, C.H.; Ko, Y.C. Identification of low-abundance proteins via fractionation of the urine proteome with weak anion exchange chromatography. Proteome Sci. 2011, 9. [Google Scholar] [CrossRef]
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Li, H.-H.; Su, J.-H.; Chiu, C.-C.; Lin, J.-J.; Yang, Z.-Y.; Hwang, W.-I.; Chen, Y.-K.; Lo, Y.-H.; Wu, Y.-J. Proteomic Investigation of the Sinulariolide-Treated Melanoma Cells A375: Effects on the Cell Apoptosis through Mitochondrial-Related Pathway and Activation of Caspase Cascade. Mar. Drugs 2013, 11, 2625-2642. https://doi.org/10.3390/md11072625
Li H-H, Su J-H, Chiu C-C, Lin J-J, Yang Z-Y, Hwang W-I, Chen Y-K, Lo Y-H, Wu Y-J. Proteomic Investigation of the Sinulariolide-Treated Melanoma Cells A375: Effects on the Cell Apoptosis through Mitochondrial-Related Pathway and Activation of Caspase Cascade. Marine Drugs. 2013; 11(7):2625-2642. https://doi.org/10.3390/md11072625
Chicago/Turabian StyleLi, Hsing-Hui, Jui-Hsin Su, Chien-Chih Chiu, Jen-Jie Lin, Zih-Yan Yang, Wen-Ing Hwang, Yu-Kuei Chen, Yu-Hsuan Lo, and Yu-Jen Wu. 2013. "Proteomic Investigation of the Sinulariolide-Treated Melanoma Cells A375: Effects on the Cell Apoptosis through Mitochondrial-Related Pathway and Activation of Caspase Cascade" Marine Drugs 11, no. 7: 2625-2642. https://doi.org/10.3390/md11072625