Interpreting the Possible Ecological Role(s) of Cyanotoxins: Compounds for Competitive Advantage and/or Physiological Aide?

{kind=link}
Abstract
:1. Introduction
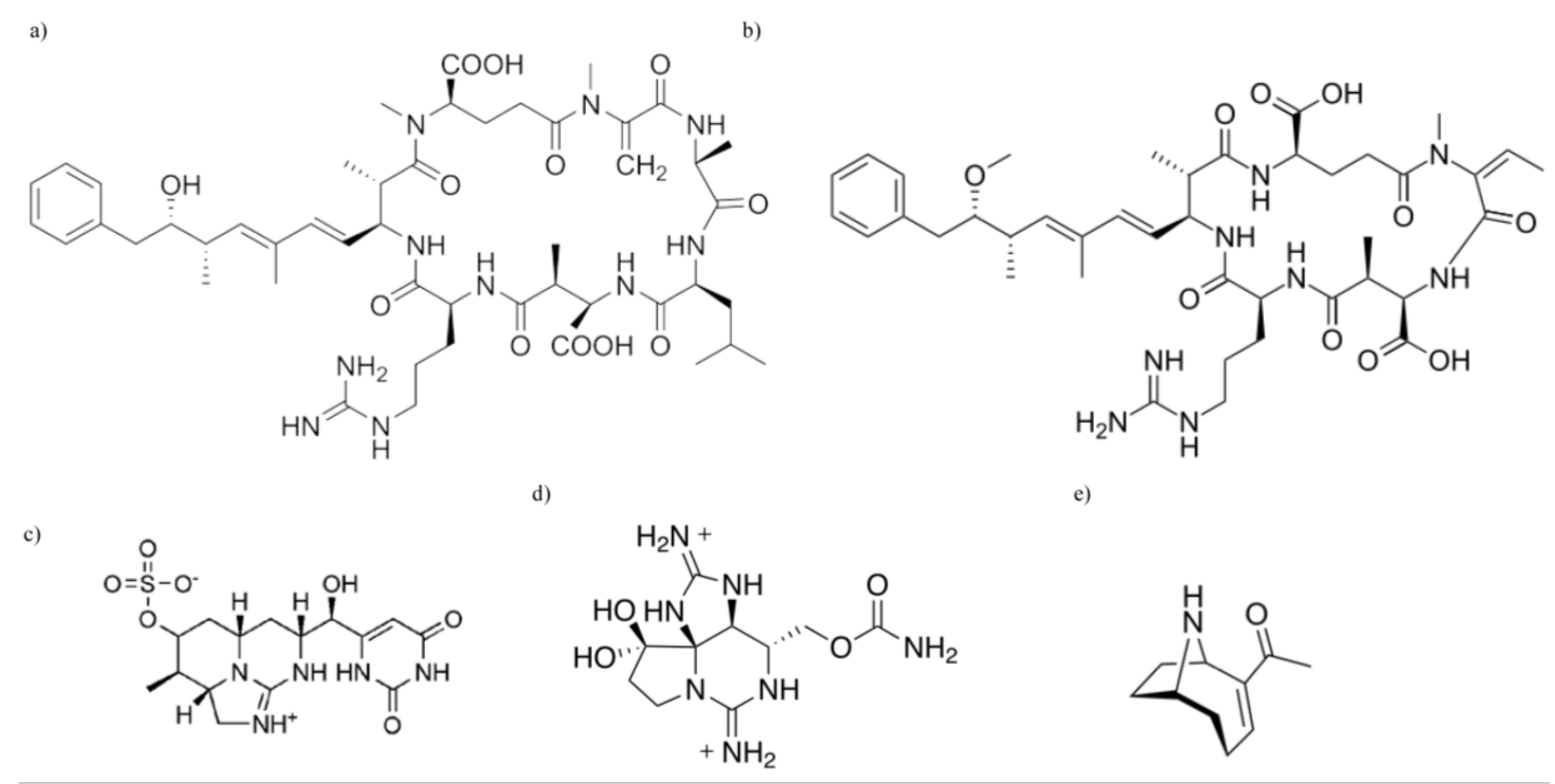
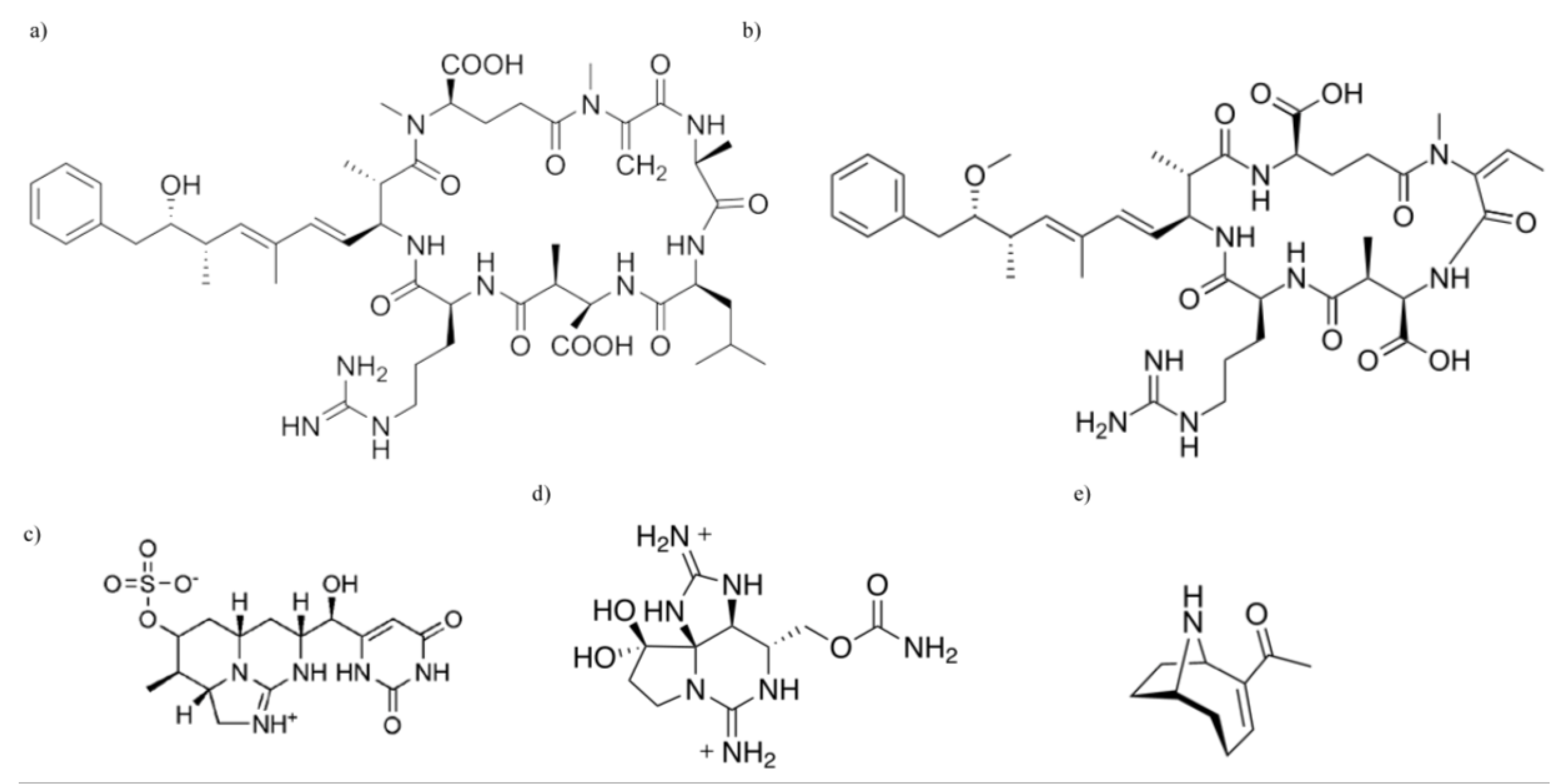
Cyanobacteria and Their Toxins
2. Determining Possible Ecological Roles
2.1. Toxins as Secondary Metabolites for Competitive Advantage
2.2. Toxins for Core Physiological Functions
3. Exploring the Evidence
3.1. Environmental Triggers for Toxin Production
3.1.1. Abiotic Factors
3.1.2. Biotic Factors
3.2. Toxicity for Competitive Advantage
3.2.1. Grazing Defence
3.2.2. Allelopathy
3.3. Toxic Compounds as Physiological Aides
3.3.1. Assistance in Nutrient Uptake
3.3.2. Iron Scavenging
3.3.3. Oxidative Stress and/or Carbon-Nitrogen Metabolism
3.3.4. Maintenance of Homeostasis
3.3.5. Roles as Infochemicals
4. Understanding the Role of Toxins: A Whole-of-System Approach
5. Implications for Future Management
6. Concluding Thoughts
Acknowledgments
Conflict of Interest
References
- Kinnear, S. Cylindrospermopsin: A decade of progress on bioaccumulation research. Mar. Drugs 2010, 8, 542–564. [Google Scholar] [CrossRef]
- Hitzfeld, B.C.; Lampert, C.S.; Spaeth, N.; Mountfort, D.; Kaspar, H.; Dietrich, D.R. Toxin production in cyanobacterial mats from ponds on the McMurdo Ice Shelf, Antarctica. Toxicon 2000, 38, 1731–1748. [Google Scholar] [CrossRef]
- Moreira, C.; Azevedo, J.; Antunes, A.; Vasconcelos, V. Cylindrospermopsin: Occurrence, methods of detection and toxicology. J. Appl. Microbiol. 2013, 114, 605–620. [Google Scholar] [CrossRef]
- Christoffersen, K. Ecological implications of cyanobacterial toxins in aquatic food webs. Phycologia 1996, 35, 42–50. [Google Scholar] [CrossRef]
- Krienitz, L.; Ballot, A.; Kotut, K.; Wiegand, C.; Pütz, S.; Metcalf, J.S.; Codd, G.A.; Pflugmacher, S. Contribution of hot spring cyanobacteria to the mysterious deaths of lesser flamingos at Lake Bogoria, Kenya. FEMS Microbiol. Ecol. 2003, 43, 141–148. [Google Scholar]
- Kaplan, A.; harel, M.; Kaplan-Levy, R.; Hadas, O.; Sukenik, A.; Dittmann, E. The languages spoken in the water body (or the biological role of cyanobacteral toxins). Front. Microbiol. 2012, 3, 1–11. [Google Scholar]
- Neilan, B.A.; Pearson, L.A.; Muenchhoff, J.; Moffitt, M.C.; Dittmann, E. Environmental conditions that influence toxin biosynthesis in cyanobacteria. Environ. Microbiol. 2012, 15, 1239–1253. [Google Scholar]
- Kaebernick, M.; Neilan, B.A. Ecological and molecular investigations of cyanotoxin production. FEMS Microbiol. Ecol. 2001, 35, 1–9. [Google Scholar] [CrossRef]
- Kurmayer, R.; Christiansen, G. The genetic basis of toxin production in cyanobacteria. Freshw. Rev. 2009, 2, 31–50. [Google Scholar]
- Carmichael, W.W. Cyanobacteria secondary metabolites—The cyanotoxins. J. Appl. Microbiol. 1992, 72, 445–459. [Google Scholar]
- Vining, L.C. Functions of secondary metabolites. Annu. Rev. Microbiol. 1990, 44, 395–427. [Google Scholar] [CrossRef]
- Vining, L.C. Secondary metabolism, inventive evolution and biochemical diversity—A review. Gene 1992, 115, 135–140. [Google Scholar] [CrossRef]
- Lillehoj, E.B. Evolutionary basis and ecological role of toxic microbial secondary metabolites. J. Theor. Biol. 1982, 97, 325–332. [Google Scholar]
- Oullette, A.; Wilhelm, S. Toxic cyanobacteria: The evolving molecular toolbox. Front. Ecol. Environ. 2003, 1, 359–366. [Google Scholar] [CrossRef]
- Codd, G.A. Cyanobacterial toxins: Occurrence, properties and biological significance. Water Sci. Technol. 1995, 32, 149–156. [Google Scholar]
- Raven, J.A. Cyanotoxins: A poison that frees phosphate. Curr. Biol. 2010, 20, R850–R852. [Google Scholar] [CrossRef]
- Utkilen, H.; Gjolme, N. Iron-stimulated toxin production in Microcystis aeruginosa. Appl. Environ. Microbiol. 1995, 61, 797–800. [Google Scholar]
- Gan, N.; Xiao, Y.; Zhu, L.; Wu, Z.; Liu, J.; Hu, C.; Song, L. The role of microcystins in maintaining colonies of bloom-forming Microcystis spp. Environ. Microbiol. 2012, 14, 730–742. [Google Scholar] [CrossRef]
- Pomati, F.; Rossetti, C.; Manarolla, G.; Burns, B.P.; Neilan, B.A. Interactions between intracellular Na+ levels and saxitoxin production in Cylindrospermopsis raciborskii T3. Microbiology 2004, 150, 455–461. [Google Scholar] [CrossRef]
- Griffiths, D.J.; Saker, M.L. The Palm Island mystery disease 20 years on: A review of research on the cyanotoxin cylindrospermopsin. Environ. Toxicol. 2003, 18, 78–93. [Google Scholar] [CrossRef]
- Sivonen, K.; Jones, G. Cyanobacterial Toxins. In Toxic Cyanobacteria in Water: A Guide to Their Public Health Consequences, Monitoring and Management; Chorus, I., Batram, J., Eds.; E and FN Spoon: London, UK, 1999. [Google Scholar]
- Bácsi, I.; Vasas, G.; Surányi, G.; M-Hamvas, M.; Máthé, C.; Tóth, E.; Grigorszky, I.; Gáspár, A.; Tóth, S.; Borbely, G. Alteration of cylindrospermopsin production in sulfate- or phosphate-starved cyanobacterium Aphanizomenon ovalisporum. FEMS Microbiol. Lett. 2006, 259, 303–310. [Google Scholar] [CrossRef]
- Bar-Yosef, Y.; Sukenik, A.; Hadas, O.; Viner-Mozzini, Y.; Kaplan, A. Enslavement in the water body by toxic Aphanizomenon ovalisporum, inducing alkaline phosphatase in phytoplanktons. Curr. Biol. 2010, 20, 1557–1561. [Google Scholar] [CrossRef]
- Cirés, S.; Wörmer, L.; Timón, J.; Wiedner, C.; Quesada, A. Cylindrospermopsin production and release by the potentially invasive cyanobacterium Aphanizomenon ovalisporum under temperature and light gradients. Harmful Algae 2011, 10, 668–675. [Google Scholar] [CrossRef]
- Dyble, J.; Tester, P.A.; Litaker, R.W. Effects of light intensity on cylindrospermopsin production in the cyanobacterial HAB species Cylindrospermopsis raciborskii. Afr. J. Mar. Sci. 2006, 28, 309–312. [Google Scholar] [CrossRef]
- Kaebernick, M.; Neilan, B.A.; Borner, T.; Dittmann, E. Light and the transcriptional response of the microcystin biosynthesis gene cluster. Appl. Environ. Microbiol. 2000, 66, 3387–3392. [Google Scholar] [CrossRef]
- Rapala, J.; Sivonen, K. Assessment of environmental conditions that favor hepatotoxic and neurotoxic Anabaena spp. strains cultured under light limitation at different temperatures. Microb. Ecol. 1998, 36, 181–192. [Google Scholar] [CrossRef]
- Van der Westhuizen, A.J.; Eloff, J.N.; Kruger, G.H.J. Effect of temperature and light (fluence rate) on the composition of the toxin of the cyanobacterium, Microcystis aeruginosa. Arch. Hydrobiol. 1986, 108, 145–154. [Google Scholar]
- Jaiswal, P.; Singh, P.K.; Prasanna, R. Cyanobacterial bioactive molecules—An overview of their toxic properties. Can. J. Microbiol. 2008, 54, 701–717. [Google Scholar] [CrossRef]
- Leflaive, J.P.; Ten-Hage, L. Algal and cyanobacterial secondary metabolites in freshwaters: A comparison of allelopathic compounds and toxins. Freshw. Biol. 2007, 52, 199–214. [Google Scholar] [CrossRef]
- Kearns, K.D.; Hunter, M.D. Green algal extracellular products regulate antialgal toxin production in a cyanobacterium. Environ. Microbiol. 2000, 2, 291–297. [Google Scholar] [CrossRef]
- Jang, M.H.; Jung, J.M.; Takamura, N. Changes in microcystin production in cyanobacteria exposed to zooplankton at different population densities and infochemical concentrations. Limnol. Oceanogr. 2007, 52, 1454–1466. [Google Scholar] [CrossRef]
- Jang, M.-H.; Ha, K.; Joo, G.-J.; Takamura, N. Toxin production of cyanobacteria is increased by exposure to zooplankton. Freshw. Biol. 2003, 48, 1540–1550. [Google Scholar] [CrossRef]
- Jang, M.-H.; Ha, K.; Lucas, M.C.; Joo, G.-J.; Takamura, N. Changes in microcystin production by Microcystis aeruginosa exposed to phytoplanktivorous and omnivorous fish. Aquat. Toxicol. 2004, 68, 51–59. [Google Scholar] [CrossRef]
- Nogueira, I.C.G.; Saker, M.L.; Pflugmacher, S.; Wiegand, C.; Vasconcelos, V.M. Toxicity of the cyanobacterium Cylindrospermopsis raciborskii to Daphnia magna. Environ. Toxicol. 2004, 19, 453–459. [Google Scholar] [CrossRef]
- Gilbert, J.J. Effect of food availability on the response of planktonic rotifers to a toxic strain of the cyanobacterium Anabaena flos-aquae. Limnol. Oceanogr. 1996, 41, 1565–1572. [Google Scholar] [CrossRef]
- Demott, W.R.; Qing-Xue, Z.; Carmichael, W.W. Effects of toxic cyanobacteria and purified toxins on the survival and feeding of a copepod and three species of Daphnia. Limnol. Oceanogr. 1991, 36, 1346–1357. [Google Scholar] [CrossRef]
- Metcalf, J.S.; Beattie, K.A.; Saker, M.L.; Codd, G.A. Effects of organic solvents on the high performance liquid chromatographic analysis of the cyanobacterial toxin cylindrospermopsin and its recovery from environmental eutrophic waters by solid phase extraction. FEMS Microbiol. Lett. 2002, 216, 159–164. [Google Scholar] [CrossRef]
- Lindsay, J.; Metcalf, J.S.; Codd, G.A. Protection against the toxicity of microcystin-LR and cylindrospermopsin in Artemia salina and Daphnia spp. by pre-treatment with cyanobacterial lipopolysaccharide (LPS). Toxicon 2006, 48, 995–1001. [Google Scholar] [CrossRef]
- Van Gremberghe, I.; Vanormelingen, P.; van der Gucht, K.; Mancheva, A.; D’Hondt, S.; de Meester, L.; Vyverman, W. Influence of Daphnia infochemicals on functional traits of Microcystis strains (cyanobacteria). Hydrobiologia 2009, 635, 147–155. [Google Scholar] [CrossRef]
- Wilken, S.; Wiezer, S.; Huisman, J.; van Donk, E. Microcystins do not provide anti-herbivore defence against mixotrophic flagellates. Aquat. Microb. Ecol. 2010, 59, 207–216. [Google Scholar] [CrossRef]
- Murray, S.A.; Mihali, T.K.; Neilan, B.A. Extraordinary conservation, gene loss, and positive selection in the evolution of an ancient neurotoxin. Mol. Biol. Evol. 2011, 28, 1173–1182. [Google Scholar]
- Rantala, A.; Fewer, D.P.; Hisbergues, M.; Rouhiainen, L.; Vaitomaa, J.; Börner, T.; Sivonen, K. Phylogenetic evidence for the early evolution of microcystin synthesis. Proc. Natl. Acad. Sci. USA 2004, 101, 568–573. [Google Scholar]
- Rantala, A. Evolution and Detection of Cyanobacterial Hepatotoxin Synthetase Genes; University of Helsinki: Helsinki, Finland, 2007. [Google Scholar]
- Sergeev, V.N.; Gerasimenko, L.M.; Zavarzin, G.A. The proterozoic history and present state of cyanobacteria. Microbiology 2002, 71, 623–637. [Google Scholar] [CrossRef]
- Buskey, E.J. How does eutrophication affect the role of grazers in harmful algal bloom dynamics? Harmful Algae 2008, 8, 152–157. [Google Scholar] [CrossRef]
- Rasmussen, J.P.; Cursaro, M.; Froscio, S.M.; Saint, C.P. An examination of the antibiotic effects of cylindrospermopsin on common gram-positive and gram-negative bacteria and the protozoan Naegleria lovaniensis. Environ. Toxicol. 2008, 23, 36–43. [Google Scholar] [CrossRef]
- Maršálek, B.; Bláha, L. Comparison of 17 biotests for detection of cyanobacterial toxicity. Environ. Toxicol. 2004, 19, 310–317. [Google Scholar] [CrossRef]
- Fabbro, L.; Baker, M.; Duivenvoorden, L.; Pegg, G.; Shiel, R. The effects of the ciliate Paramecium cf. caudatum Ehrenberg on toxin producing Cylindrospermopsis isolated from the Fitzroy River, Australia. Environ. Toxicol. 2001, 16, 489–497. [Google Scholar] [CrossRef]
- Kinnear, S.H.W.; Duivenvoorden, L.J.; Fabbro, L.D. Sublethal responses in Melanoides tuberculata following exposure to Cylindrospermopsis raciborskii containing cylindrospermopsin. Harmful Algae 2007, 6, 642–650. [Google Scholar] [CrossRef]
- Singh, D.P.; Tyagi, M.B.; Kumar, A.; Thakur, J.K.; Kumar, A. Antialgal activity of a hepatotoxin-producing cyanobacterium, Microcystis aeruginosa. Word J. Microbiol. Biotechnol. 2001, 17, 15–22. [Google Scholar] [CrossRef]
- Kearns, K.D.; Hunter, M.D. Toxin-producing Anabaena flos-aquae induces settling of Chlamydomonas reinhardtii, a competing motile alga. Microb. Ecol. 2001, 42, 80–86. [Google Scholar]
- Hu, Z.Q.; Liu, Y.D.; Li, D.H. Physiological and biochemical analyses of microcystin-RR toxicity to the cyanobacterium Synechococcus elongatus. Environ. Toxicol. 2004, 19, 571–577. [Google Scholar] [CrossRef]
- Pflugmacher, S. Promotion of oxidative stress in the aquatic macrophyte Ceratophyllum demersum during biotransformation of the cyanobacterial toxin microcystin-LR. Aquat. Toxicol. 2004, 70, 169–178. [Google Scholar] [CrossRef]
- Pflugmacher, S.; Codd, G.A.; Steinberg, C.E.W. Effects of the cyanobacterial toxin microcystin-LR on detoxication enzymes in aquatic plants. Environ. Toxicol. 1999, 14, 111–115. [Google Scholar] [CrossRef]
- Romanowska-Duda, Z.; Tarczyńska, M. The influence of microcystin-LR and hepatotoxic cyanobacterial extract on the water plant Spirodela oligorrhiza. Environ. Toxicol. 2002, 17, 434–440. [Google Scholar] [CrossRef]
- Mitrovic, S.M.; Pflugmacher, S.; James, K.J.; Furey, A. Anatoxin-a elicits an increase in peroxidase and glutathione S-transferase activity in aquatic plants. Aquat. Toxicol. 2004, 68, 185–192. [Google Scholar] [CrossRef]
- Figueredo, C.C.; Giani, A.; Bird, D.F. Does allelopathy contribute to Cylindrospermopsis raciborskii (cyanobacteria) bloom occurence and geographic expansion? J. Phycol. 2007, 43, 256–265. [Google Scholar] [CrossRef]
- Silva, P.; Vasconcelos, V. Allelopathic effect of Cylindrospermopsis raciborskii extracts on the germination and growth of several plant species. Chem. Ecol. 2010, 26, 263–271. [Google Scholar] [CrossRef]
- Legrand, C.; Rengefors, K.; Fistarol, G.O.; Granéli, E. Allelopathy in phytoplankton—Biochemical, ecological and evolutionary aspects. Phycologia 2003, 42, 406–419. [Google Scholar] [CrossRef]
- Pinheiro, C.; Azevedo, J.; Campos, A.; Loureiro, S.; Vasconcelos, V. Absence of negative allelopathic effects of cylindrospermopsin and microcystin-LR on selected marine and freshwater phytoplankton species. Hydrobiologia 2013, 705, 27–42. [Google Scholar] [CrossRef]
- Kinnear, S.H.W.; Fabbro, L.D.; Duivenvoorden, L.J. Variable growth responses of water thyme (Hydrilla verticillata) to whole-cell extracts of Cylindrospermopsis raciborskii. Arch. Environ. Contam. Toxicol. 2008, 54, 187–194. [Google Scholar]
- Babica, P.; Bláha, L.; Maršálek, B. Exploring the natural role of Microcystins—A review of effects on photoautotrophic organisms. J. Phycol. 2006, 42, 9–20. [Google Scholar] [CrossRef]
- Berry, J.P.; Gantar, M.; Perez, M.H.; Berry, G.; Noriega, F.G. Cyanobacterial toxins as allelochemicals with potential applications as algaecides, herbicides and insecticides. Mar. Drugs 2008, 6, 117–146. [Google Scholar] [CrossRef]
- Jonsson, P.R.; Pavia, H.; Toth, G. Formation of harmful algal blooms cannot be explained by allelopathic interactions. Proc. Natl. Acad. Sci. USA 2009, 106, 11177–11182. [Google Scholar] [CrossRef]
- Leão, P.N.; Vasconcelos, M.T.S.D.; Vasconcelos, V.M. Allelopathy in freshwater cyanobacteria. Crit. Rev. Microbiol. 2009, 35, 271–282. [Google Scholar] [CrossRef]
- Hadas, O.; Pinkas, R.; Delphine, E.; Vardi, A.; Kaplan, A.; Sukenik, A. Limnological and ecophysiological aspects of Aphanizomenon ovalisporum bloom in Lake Kinneret, Israel. J. Plankton Res. 1999, 21, 1439–1453. [Google Scholar] [CrossRef]
- Kurmayer, R. The toxic cyanobacterium Nostoc sp. strain 152 produces highest amounts of microcystin and nostophycin under stress conditions. J. Phycol. 2011, 47, 200–207. [Google Scholar] [CrossRef]
- Oh, H.-M.; Lee, S.J.; Jang, M.-H.; Yoon, B.-D. Microcystin production by Microcystis aeruginosa in a Phosphorus-Limited Chemostat. Appl. Environ. Microbiol. 2000, 66, 176–179. [Google Scholar] [CrossRef]
- Granéli, E.; Flynn, K. Chemical and Physical Factors Influencing Toxin Content. In Ecology of Harmful Algae; Granéli, E., Turner, J.T., Eds.; Springer Berlin Heidelberg: New York, NY, USA, 2006; Volume 189, pp. 229–241. [Google Scholar]
- Glibert, P.; Burkholder, J. Harmful algal blooms and eutrophication: “Strategies” for nutrient uptake and growth outside the Redfield comfort zone. Chin. J. Oceanol. Limnol. 2011, 29, 724–738. [Google Scholar] [CrossRef]
- Wang, J.; Liu, B.; Guo, N.; Xie, P. Alkaline phosphatase activity in four Microcystis aeruginosa species and their responses to nonylphenol stress. Bull. Environ. Contam. Toxicol. 2006, 76, 999–1006. [Google Scholar] [CrossRef]
- Lyck, S.; Gjølme, N.; Utkilen, H. Iron starvation increases toxicity of Microcystis aeruginosa CYA 228/1 (Chroococcales, Cyanophyceae). Phycologia 1996, 35, 120–124. [Google Scholar] [CrossRef]
- Alexova, R.; Fujii, M.; Birch, D.; Cheng, J.; Waite, T.D.; Ferrari, B.C.; Neilan, B.A. Iron uptake and toxin synthesis in the bloom-forming Microcystis aeruginosa under iron limitation. Environ. Microbiol. 2011, 13, 1064–1077. [Google Scholar] [CrossRef]
- Sevilla, E.; Martin-Luna, B.; Vela, L.; Bes, M.T.; Fillat, M.F.; Peleato, M.L. Iron availability affects mcyD expression and microcystin-LR synthesis in Microcystis aeruginosa PCC7806. Environ. Microbiol. 2008, 10, 2476–2483. [Google Scholar] [CrossRef]
- Zilliges, Y.; Kehr, J.-C.; Meissner, S.; Ishida, K.; Mikkat, S.; Hagemann, M.; Kaplan, A.; Börner, T.; Dittmann, E. The cyanobacterial hepatotoxin microcystin binds to proteins and increases the fitness of Microcystis under oxidative stress conditions. PLoS One 2011, 6, e17615. [Google Scholar] [CrossRef]
- Borner, T.; Dittman, E. Molecular Biology of Cyanobacterial Toxins Genetic Basis of Microcystin Production. In Harmful Cyanobacteria; Huisman, J., Matthijs, H.C.P., Visser, P., Eds.; Aquatic Ecology Series; Springer: Dordrecht, The Netherlands, 2005; Volume 3, pp. 25–40. [Google Scholar]
- Alexova, R.; Haynes, B.; Ferrari, C.; Neilan, B. Comparative protein expression in different strains of the bloom-forming cyanobacterium Microcystis aeruginosa. Mol. Cell. Proteomics 2007, 10, 1–16. [Google Scholar]
- Jahnichen, S.; Ihle, T.; Petzoldt, T.; Benndorf, J. Impact of inorganic carbon availability on microcystin production by Microcystis aeruginosa PCC 7806. Appl. Environ. Microb. 2007, 73, 6994–7002. [Google Scholar] [CrossRef]
- Van de Waal, D.B.; Verspagen, J.M.H.; Finke, J.F.; Vournazou, V.; Immers, A.K.; Kardinaal, W.E.A.; Tonk, L.; Becker, S.; van Donk, E.; Visser, P.M.; et al. Reversal in competitive dominance of a toxic versus non-toxic cyanobacterium in response to rising CO2. ISME J. 2011, 5, 1438–1450. [Google Scholar] [CrossRef]
- El-Shehawy, R.; Gorokhova, E.; Fernández-Piñas, F.; del Campo, F.F. Global warming and hepatotoxin production by cyanobacteria: What can we learn from experiments? Water Res. 2012, 46, 1420–1429. [Google Scholar] [CrossRef]
- Jonasson, S.; Vintila, S.; Sivonen, K.; El-Shehawy, R. Expression of the nodularin synthetase genes in the Baltic Sea bloom-former cyanobacterium Nodularia spumigena strain AV1. FEMS Microbiol. Ecol. 2008, 65, 31–39. [Google Scholar] [CrossRef]
- Pomati, F.; Rossetti, C.; Calamari, D.; Neilan, B.A. Effects of saxitoxin (STX) and veratridine on bacterial Na+-K+ fluxes: A prokaryote-based STX bioassay. Appl. Environ. Microb. 2003, 69, 7371–7376. [Google Scholar] [CrossRef]
- Pomati, F.; Neilan, B.A.; Suzuki, T.; Manarolla, G.; Rossetti, C. Enhancement of intracellular saxitoxin accumulation by lidocaine hydrochloride in the cyanobacterium Cylindrospermopsis raciborskii T3 (Nostocales). J. Phycol. 2003, 39, 535–542. [Google Scholar] [CrossRef]
- Soto-Liebe, K.; Méndez, M.A.; Fuenzalida, L.; Krock, B.; Cembella, A.; Vásquez, M. PSP toxin release from the cyanobacterium Raphidiopsis brookii D9 (Nostocales) can be induced by sodium and potassium ions. Toxicon 2012, 60, 1324–1334. [Google Scholar] [CrossRef]
- Bownik, A. Harmful algae: Effects of alkaloid cyanotoxins on animal and human health. Toxin Rev. 2010, 29, 99–114. [Google Scholar] [CrossRef]
- Schatz, D.; Keren, Y.; Vardi, A.; Sukenik, A.; Carmeli, S.; Börner, T.; Dittmann, E.; Kaplan, A. Towards clarification of the biological role of microcystins, a family of cyanobacterial toxins. Environ. Microbiol. 2007, 9, 965–970. [Google Scholar] [CrossRef]
- Kehr, J.C.; Zilliges, Y.; Springer, A.; Disney, M.D.; Ratner, D.D.; Bouchier, C.; Seeberger, P.H.; Marsac, N.T.D.; Dittmann, E. A mannan binding lectin is involved in cell-cell attachment in a toxic strain of Microcystis aeruginosa. Mol. Microbiol. 2006, 59, 893–906. [Google Scholar] [CrossRef]
- Zilliges, Y.; Kehr, J.C.; Mikkat, S.; Bouchier, C.; Marsac, N.T.D.; Borner, T.; Dittmann, E. An extracellular GLYCOPROTEIN is implicated in cell-cell contacts in the toxic cyanobacterium Microcystis aeruginosa PCC 7806. J. Bacteriol. 2008, 190, 2871–2879. [Google Scholar] [CrossRef]
- Kurmayer, R.; Christiansen, G.; Chorus, I. The abundance of microcystin-producing genotypes correlates positively with colony size in Microcystis sp. and determines its microcystin net production in Lake Wannsee. Appl. Environ. Microb. 2003, 69, 787–795. [Google Scholar] [CrossRef]
- Ohtani, I.; Moore, R.E.; Runnegar, M.T.C. Cylindrospermopsin: A potent hepatotoxin from the blue green alga Cylindrospermopsis raciborskii. J. Am. Chem. Soc. 1992, 114, 7941–7942. [Google Scholar] [CrossRef]
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Holland, A.; Kinnear, S. Interpreting the Possible Ecological Role(s) of Cyanotoxins: Compounds for Competitive Advantage and/or Physiological Aide? Mar. Drugs 2013, 11, 2239-2258. https://doi.org/10.3390/md11072239
Holland A, Kinnear S. Interpreting the Possible Ecological Role(s) of Cyanotoxins: Compounds for Competitive Advantage and/or Physiological Aide? Marine Drugs. 2013; 11(7):2239-2258. https://doi.org/10.3390/md11072239
Chicago/Turabian StyleHolland, Aleicia, and Susan Kinnear. 2013. "Interpreting the Possible Ecological Role(s) of Cyanotoxins: Compounds for Competitive Advantage and/or Physiological Aide?" Marine Drugs 11, no. 7: 2239-2258. https://doi.org/10.3390/md11072239