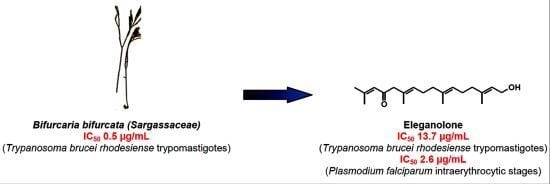
Eleganolone, a Diterpene from the French Marine Alga Bifurcaria bifurcata Inhibits Growth of the Human Pathogens Trypanosoma brucei and Plasmodium falciparum

Abstract
:
1. Introduction
2. Results and Discussion
2.1. Selected Species

{kind=link}
{kind=link}
| Species | Family | Collection site | Collection time |
|---|---|---|---|
| Chlorophyta | |||
| Codium tomentosum Stackhouse | Codiaceae | Cap Lévy (Manche) | June 2007 |
| Ulva lactuca (Linnaeus) | Ulvaceae | Luc-sur-Mer (Calvados) | October 2006 |
| Ulva clathrata (Roth) C. Agardh | Ulvaceae | Anse St Martin (Manche) | June 2007 |
| Heterokontophyta | |||
| Bifurcaria bifurcata R. Ross | Sargassaceae | Cap Lévy (Manche) | June 2007 |
| Dictyopteris polypodioides (A.P. de Candolle) J.V. Lamouroux | Dictyotaceae | Barneville (Calvados) | October 2007 |
| Dictyota dichotoma (Hudson) J.V. Lamouroux | Dictyotaceae | Anse St Martin (Manche) | June 2007 |
| Fucus serratus (Linnaeus) | Fucaceae | Luc-sur-mer (Calvados) | November 2005 |
| Himanthalia elongata (Linnaeus) | Himanthaliaceae | Cap Lévy (Manche) | June 2006 |
| Laminaria digitata (Linnaeus) J.V. Lamouroux | Laminariaceae | Langrunes-sur-Mer (Calvados) | January 2007 |
| Pelvetia canaliculata Decaisne & Thuret | Fucaceae | Cap Lévy (Manche) | June 2006 |
| Sargassum muticum (Yendo) Fensholt | Sargassaceae | Cap Lévy (Manche) | June 2006 |
| Rhodophyta | |||
| Calliblepharis jubata (Goodenough & Woodward) Kützing | Cystocloniaceae | Cap Lévy (Manche) | June 2007 |
| Chondrus crispus Stackhouse | Gigartinaceae | Cap Lévy (Manche) | June 2007 |
| Dilsea carnosa (Schmidel) Kuntze | Dumontiaceae | Langrune-sur-Mer (Calvados) | January 2007 |
| Gelidium latifolium Bornet ex Hauck | Gelidiaceae | Cap Lévy (Manche) | June 2006 |
| Gracilaria gracilis (Stackhouse) Steentoft, L.M. Irvine & Farnham | Gracilariaceae | Anse St Martin (Manche) | June 2007 |
| Grateloupia turuturu Yamada | Halymeniaceae | St Vaast-la-Hougue (Manche) | September 2007 |
| Halurus flosculosus (J. Ellis) Maggs & Hommersand | Ceramiaceae | Anse St Martin (Manche) | June 2007 |
| Mastocarpus stellatus (Stackhouse) Guiry | Phyllophoraceae | Cap Lévy (Manche) | June 2006 |
| Palmaria palmata (Linnaeus) Kuntze | Palmariaceae | Luc-sur-Mer (Calvados) | November 2005 |
2.2. In Vitro Trypanocidal Activity of the Selected Species
| IC50 (µg/mL) | Selectivity index (SI) | |||
|---|---|---|---|---|
| Antitrypanosomal activity | Cytotoxic activity | |||
| Species | Extract | T. brucei rhodesiense | L6 cells | |
| B. bifurcata | E | 29.7 | 76.0 | 2.6 |
| B. bifurcata | A | 0.5 | 6.2 | 12.4 |
| C. jubata | A | 23.3 | 71.5 | 3.1 |
| C. crispus | A | 13.6 | 84.3 | 6.2 |
| D. dichotoma | A | 5.8 | 27.8 | 4.8 |
| D. carnosa | A | 15.3 | 74.0 | 4.8 |
| G. latifolium | A | 20.5 | 62.1 | 3.0 |
| G. gracilis | A | 21.5 | 71.3 | 3.3 |
| G. turuturu | A | 10.8 | 71.2 | 6.6 |
| H. flosculosus | A | 22.4 | 58.7 | 2.6 |
| H. elongata | A | 30.3 | 88.3 | 2.9 |
| M. stellatus | A | 19.5 | 69.1 | 3.5 |
| P. canaliculata | A | 7.8 | 86.7 | 11.1 |
| S. muticum | A | 5.8 | 27.8 | 4.8 |
| Standards | ||||
| Melarsoprol | 0.004 | |||
| Podophyllotoxin | 0.007 | |||
2.3. Bio-Guided Fractionation of the Most Active Extract Obtained from B. bifurcata
| Antiprotozoal activity | Cytotoxic activity | ||||||
|---|---|---|---|---|---|---|---|
| IC50 (µg/mL) | IC50 (µg/mL) | SI | |||||
| T. brucei rhodesiense | T. cruzi | P. falciparum | L6 cells | SI a | SI b | SI c | |
| Ethyl acetate extract | 0.5 | 4.1 * | >5 * | 6.2 * | 12.4 | 1.5 * | >1 * |
| Fractions | |||||||
| Fraction 1 | 12.9 | 32.4 | 8.5 | 40.2 | 3.1 | 1.2 | 4.7 |
| Fraction 2 | 0.5 | 9.7 | 3.8 | 7.4 | 15.4 | 0.8 | 1.9 |
| Fraction 3 | 4.3 | 11.7 | 3.7 | 16.9 | 3.9 | 1.4 | 4.6 |
| Fraction 4 | 14.7 | 25.0 | 3.0 | 55.1 | 3.7 | 2.2 | 18.4 |
| Fraction 5 | 14.1 | 32.1 | 4.8 | 38.7 | 2.7 | 1.2 | 8.0 |
| Standard drugs | |||||||
| Melarsoprol | 0.004 | ||||||
| Benznidazole | 0.536 | ||||||
| Chloroquine | 0.069 | ||||||
| Artemisinin | 0.002 | ||||||
| Podophyllotoxin | 0.007 | ||||||
| Antiprotozoal activity | Cytotoxic activity | |||||
|---|---|---|---|---|---|---|
| IC50 in µM (µg/mL) | IC50 (µg/mL) | SI | ||||
| T. brucei rhodesiense | T. cruzi | P. falciparum | L6 cells | SI a | SI b | |
| Eleganolone | 45 (13.7) | 58 (17.7) | 7.9 (2.6) | 184 (56.1) | 4.0 | 21.6 |
| Standard drugs | ||||||
| Melarsoprol | 0.005 (0.004) | |||||
| Benznidazole | 1.69 (0.536) | |||||
| Chloroquine | 0.19 (0.069) | |||||
| Artemisinin | 0.007 (0.002) | |||||
| Podophyllotoxin | 0.048 (0.007) | |||||
2.3.1. Fractions
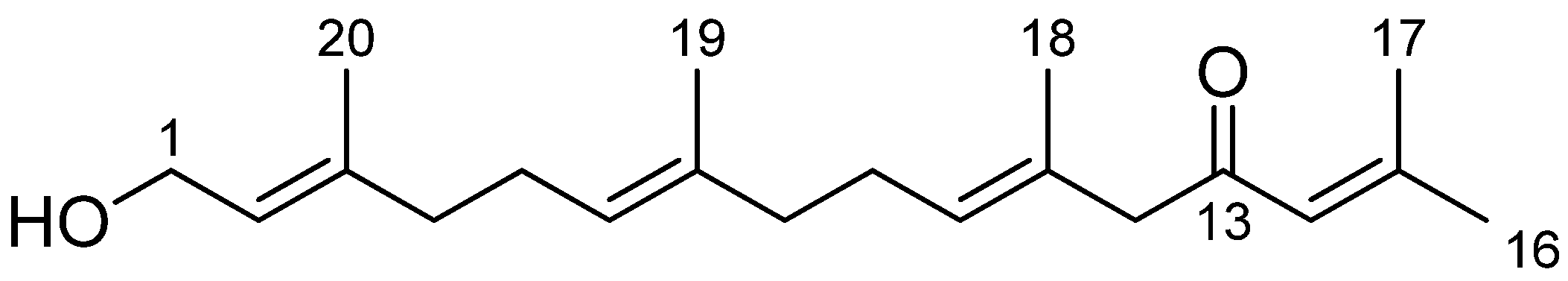
2.3.2. Pure Compound

3. Experimental Section
3.1. General Experimental Procedures
3.2. Algae Collection and Identification
3.3. Extraction and Compound Isolation
3.4. In Vitro Antiprotozoal Assays
3.4.1. Activity against P. falciparum
3.4.2. Activity against Trypanosoma cruzi
3.4.3. Activity against Trypanosoma brucei rhodesiense
3.5. Cytotoxicity against L6 Cells
3.6. Calculation of IC50
4. Conclusions
Acknowledgments
Conflict of Interest
References
- World Health Organization, Working to Overcome the Global Impact of Neglected Tropical Diseases; First WHO Report on Neglected Tropical Diseases; WHO: Geneva, Switzerland, 2010.
- Brun, R.; Blum, J.; Chappuis, F.; Burri, C. Human African trypanosomiasis. Lancet 2010, 375, 148–159. [Google Scholar] [Green Version]
- Chakraborty, K.; Lipton, A.P.; Paulraj, R.; Chakraborty, R.D. Guaiane sesquiterpenes from seaweed Ulva fasciata Delile and their antibacterial properties. Eur. J. Med. Chem. 2010, 45, 2237–2244. [Google Scholar] [CrossRef]
- Moo-Puc, R.; Robledo, D.; Freile-Pelegrin, Y. Evaluation of selected tropical seaweeds for in vitro anti-trichomonal activity. J. Ethnopharmacol. 2008, 120, 92–97. [Google Scholar] [CrossRef]
- Kubanek, J.; Jensen, P.R.; Keifer, P.A.; Sullards, M.C.; Collins, D.O.; Fenical, W. Seaweed resistance to microbial attack: A targeted chemical defense against marine fungi. Proc. Natl. Acad. Sci. USA 2003, 100, 6916–6921. [Google Scholar]
- Vonthron-Sénécheau, C.; Kaiser, M.; Devambez, I.; Vastel, A.; Mussio, I.; Rusig, A.M. Antiprotozoal activities of organic extracts from French marine seaweeds. Mar. Drugs 2011, 9, 922–933. [Google Scholar] [CrossRef]
- Spavieri, J.; Allmendinger, A.; Kaiser, M.; Casey, R.; Hingley-Wilson, S.; Lalvani, A.; Guiry, M.D.; Blunden, G.; Tasdemir, D. Antimycobacterial, antiprotozoal and cytotoxic potential of twenty-one brown algae (Phaeophyceae) from British and Irish waters. Phytother. Res. 2010, 24, 1724–1729. [Google Scholar] [CrossRef]
- Freile-Pelegrin, Y.; Robledo, D.; Chan-Bacab, M.J.; Ortega-Morales, B.O. Antileishmanial properties of tropical marine algae extracts. Fitoterapia 2008, 79, 374–377. [Google Scholar] [CrossRef]
- Orhan, I.; Sener, B.; Atici, T.; Brun, R.; Perozzo, R.; Tasdemir, D. Turkish freshwater and marine macrophyte extracts show in vitro antiprotozoal activity and inhibit FabI, a key enzyme of Plasmodium falciparum fatty acid biosynthesis. Phytomedicine 2006, 13, 388–393. [Google Scholar] [CrossRef]
- Nara, T.; Kamei, Y.; Tsubouchi, A.; Annoura, T.; Hirota, K.; Iizumi, K.; Dohmoto, Y.; Ono, T.; Aoki, T. Inhibitory action of marine algae extracts on the Trypanosoma cruzi dihydroorotate dehydrogenase activity and on the protozoan growth in mammalian cells. Parasitol. Int. 2005, 54, 59–64. [Google Scholar] [CrossRef]
- Spavieri, J.; Kaiser, M.; Casey, R.; Hingley-Wilson, S.; Lalvani, A.; Blunden, G.; Tasdemir, D. Antiprotozoal, antimycobacterial and cytotoxic potential of some British green algae. Phytotherapy Res. 2010, 24, 1095–1098. [Google Scholar]
- Moreau, D.; Thomas-Guyon, H.; Jacquot, C.; Jugé, M.; Culioli, G.; Ortalo-Magné, A.; Piovetti, L.; Roussakis, C. An extract from the brown alga Bifurcaria bifurcata induces irreversible arrest of cell proliferation in a non-small-cell bronchopulmonary carcinoma line. J. Appl. Phycol. 2006, 18, 87–93. [Google Scholar] [CrossRef]
- Patel, A.V.; Wright, D.C.; Romero, M.A.; Blunden, G.; Guiry, M.D. Molluscicidal polyphenols from species of Fucaceae. Nat. Product Commun. 2008, 3, 245–249. [Google Scholar]
- Biard, J.F.; Verbist, J.F.; Letourneux, Y.; Floch, R. Antimicrobially active diterpene ketols from Bifurcaria bifurcata. Planta Med. 1980, 40, 288–294. [Google Scholar] [CrossRef]
- Glombitza, K.W.; Roesener, H.U.; Koch, M.L. Antibiotics from algae. Part 16. Polyhydroxyoligophenyls and phenyl ether from Bifurcaria bifurcata. Phytochemistry 1976, 15, 1279–1281. [Google Scholar]
- Glombitza, K.W.; Roesener, H.U. Antibiotics from algae. XI. Bifuhalol. Diphenylether from Bifurcaria bifurcata. Phytochemistry 1974, 13, 1245–1247. [Google Scholar]
- Valls, R.; Piovetti, L.; Deffo, P. Analysis of sterols and diterpenoids of brown algae (Cystoseiraceae). Oceanis 1991, 17, 305–307. [Google Scholar]
- Göthel, Q.; Lichte, E.; Köck, M. Further eleganolone-derived diterpenes from the brown alga Bifurcaria bifurcata. Tetrahedron Lett. 2012, 53, 1873–1877. [Google Scholar] [CrossRef]
- Ortalo-Magne, A.; Culioli, G.; Valls, R.; Pucci, B.; Piovetti, L. Polar acyclic diterpenoids from Bifurcaria bifurcata (Fucales, Phaeophyta). Phytochemistry 2005, 66, 2316–2323. [Google Scholar] [CrossRef]
- Culioli, G.; Ortalo-Magne, A.; Daoudi, M.; Thomas-Guyon, H.; Valls, R.; Piovetti, L. Trihydroxylated linear diterpenes from the brown alga Bifurcaria bifurcata. Phytochemistry 2004, 65, 2063–2069. [Google Scholar]
- Culioli, G.; Daoudi, M.; Ortalo-Magne, A.; Valls, R.; Piovetti, L. (S)-12-Hydroxygeranyl geraniol-derived diterpenes from the brown alga Bifurcaria bifurcate. Phytochemistry 2001, 57, 529–535. [Google Scholar]
- Culioli, G.; Mesguiche, V.; Piovetti, L.; Valls, R. Geranylgeraniol and geranylgeraniol-derived diterpenes from the brown alga Bifurcaria bifurcata (Cystoseiraceae). Biochem. Syst. Ecol. 1999, 27, 665–668. [Google Scholar] [CrossRef]
- Hougaard, L.; Anthoni, U.; Christophersen, C.; Nielsen, P.H. Eleganolone-derived diterpenes from Bifurcaria bifurcata. Phytochemistry 1991, 30, 3049–3051. [Google Scholar]
- Hougaard, L.; Anthoni, U.; Christophersen, C.; Nielsen, P.H. Two new diterpenoid dihydroxy-γ-butyrolactones from Bifurcaria bifurcata (Cystoseiraceae). Tetrahedron Lett. 1991, 32, 3577–3578. [Google Scholar] [CrossRef]
- Semmak, L.; Zerzouf, A.; Valls, R.; Banaigs, B.; Jeanty, G.; Francisco, C. Acyclic diterpenes from Bifurcaria bifurcata. Phytochemistry 1988, 27, 2347–2349. [Google Scholar]
- Göthel, Q.; Muñoz, J.; Köck, M. Formyleleganolone and bibifuran, two metabolites from the brown alga Bifurcaria bifurcata. Phytochem. Lett. 2012, 5, 693–695. [Google Scholar] [CrossRef]
- Biard, J.F.; Verbist, J.F.; Floch, R.; Letourneux, Y. Epoxyeleganolone and eleganediol, two new diterpenes from Bifurcaria bifurcata Ross (Cystoseiraceae). Tetrahedron Lett. 1980, 21, 1849–1852. [Google Scholar] [CrossRef]
- Valls, R.; Piovetti, L.; Banaig, B.; Archavlis, A.; Pellegrini, M. (S)-13-hydroxygeranyl geraniol-derived furanoditerpenes from Bifurcaria bifurcata. Phytochemistry 1995, 39, 145–149. [Google Scholar]
- Zee, O.P.; Kim, D.K.; Choi, S.U.; Lee, C.O.; Lee, K.R. A new cytotoxic acyclic diterpene from Carpesium divaricatum. Arch. Pharm. Res. 1999, 2, 225–227. [Google Scholar]
- Hellio, C.; de la Broise, D.; Dufossé, L.; le Gal, Y.; Bourgougnon, N. Inhibition of marine bacteria by extracts of macroalgae: Potential use for environmentally friendly antifouling paints. Mar. Environ. Res. 2001, 52, 231–247. [Google Scholar] [CrossRef]
- Hellio, C.; Thomas-Guyon, H.; Culioli, G.; Piovetti, L.; Bourgougnon, N.; le Gal, Y. Marine antifoulants from Bifurcaria bifurcata (Phaeophyceae, Cystoseiraceae) and other brown macroalgae. Biofouling J. Bioadhes. Biofilm Res. 2001, 17, 189–201. [Google Scholar] [CrossRef]
- Geraci, S.; Faimali, M.; Piovetti, L.; Cimino, G. Antifouling from Nature: Laboratory Test with Balanus Amphitrite Darwin on Algae and Sponges; DSTO Technical Report No. DSTO-GD-0287; Defence Science and Technology Organisation: Melbourne, Australia, 2001. [Google Scholar]
- Scala, F.; Fattorusso, E.; Menna, M.; Taglialatela-Scafati, O.; Tierney, M.; Kaiser, M.; Tasdemir, D. Bromopyrrole alkaloids as lead compounds against protozoan parasites. Mar. Drugs 2010, 8, 2162–2174. [Google Scholar] [CrossRef]
- Thaithong, S.; Beale, G.H. Resistance of ten Thai isolates of Plasmodium falciparum to chloroquine and pyrimethamine by in vitro tests. Trans. R. Soc. Trop. Med. Hyg. 1981, 75, 271–273. [Google Scholar] [CrossRef]
- Buckner, F.S.; Verlinde, C.L.; la Flamme, A.C.; van Voorhis, W.C. Efficient technique for screening drugs for activity against Trypanosoma cruzi using parasites expressing beta-galactosidase. Antimicrob. Agents Chemother. 1996, 40, 2592–2597. [Google Scholar]
- Freiburghaus, F.; Kaminsky, R.; Nkunya, M.H.H.; Brun, R. Evaluation of African medicinal plants for their in vitro trypanocidal activity. J. Ethnopharmacol. 1996, 55, 1–11. [Google Scholar] [CrossRef]
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Gallé, J.-B.; Attioua, B.; Kaiser, M.; Rusig, A.-M.; Lobstein, A.; Vonthron-Sénécheau, C. Eleganolone, a Diterpene from the French Marine Alga Bifurcaria bifurcata Inhibits Growth of the Human Pathogens Trypanosoma brucei and Plasmodium falciparum. Mar. Drugs 2013, 11, 599-610. https://doi.org/10.3390/md11030599
Gallé J-B, Attioua B, Kaiser M, Rusig A-M, Lobstein A, Vonthron-Sénécheau C. Eleganolone, a Diterpene from the French Marine Alga Bifurcaria bifurcata Inhibits Growth of the Human Pathogens Trypanosoma brucei and Plasmodium falciparum. Marine Drugs. 2013; 11(3):599-610. https://doi.org/10.3390/md11030599
Chicago/Turabian StyleGallé, Jean-Baptiste, Barthélémy Attioua, Marcel Kaiser, Anne-Marie Rusig, Annelise Lobstein, and Catherine Vonthron-Sénécheau. 2013. "Eleganolone, a Diterpene from the French Marine Alga Bifurcaria bifurcata Inhibits Growth of the Human Pathogens Trypanosoma brucei and Plasmodium falciparum" Marine Drugs 11, no. 3: 599-610. https://doi.org/10.3390/md11030599