Induction of Apoptosis, G0/G1 Phase Arrest and Microtubule Disassembly in K562 Leukemia Cells by Mere15, a Novel Polypeptide from Meretrix meretrix Linnaeus

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
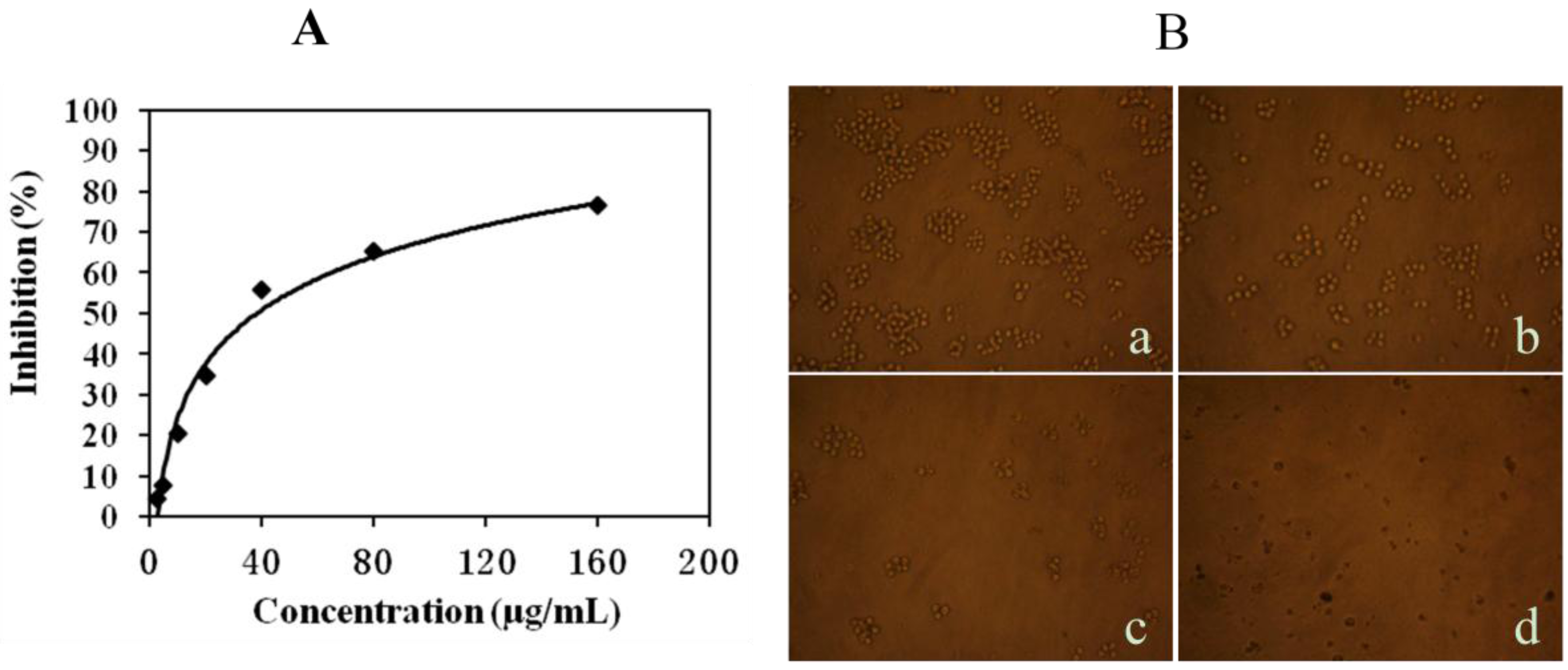
2.1. Mere15 Inhibits K562 Cell Proliferation
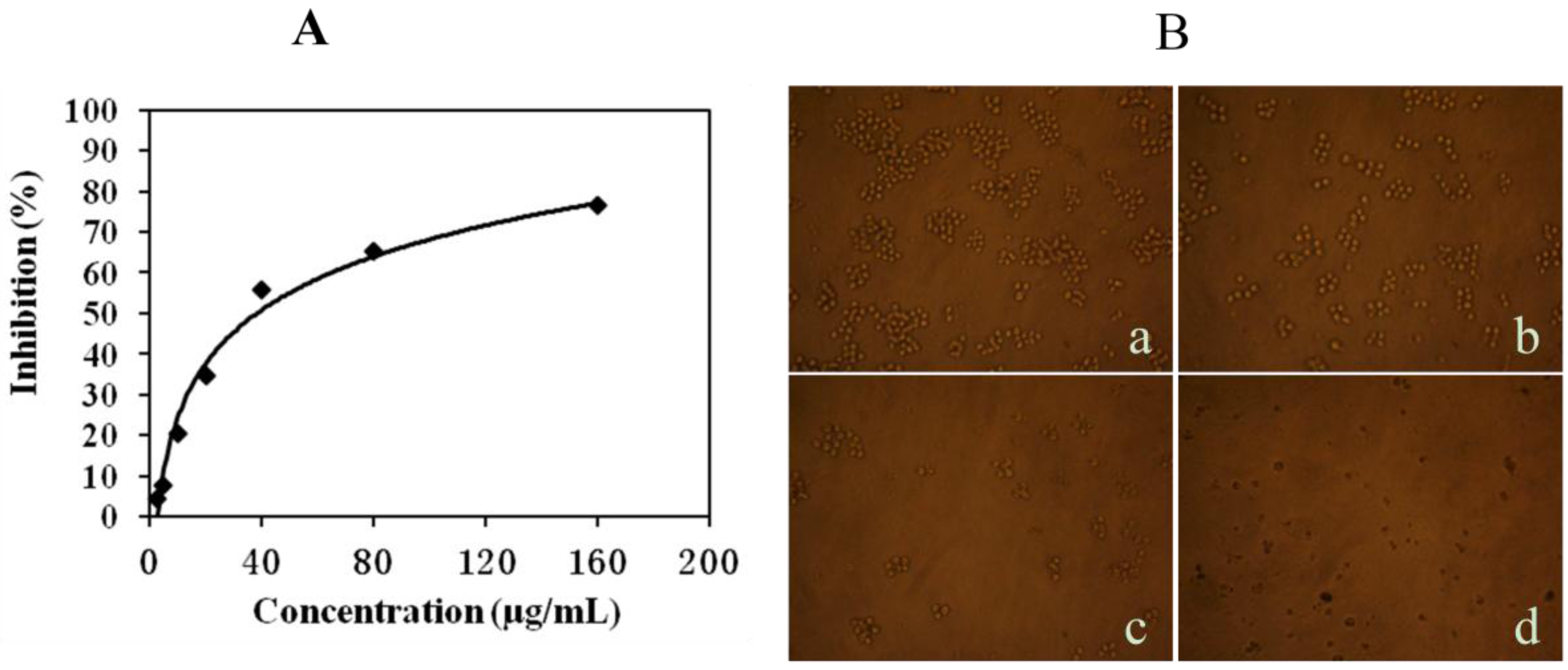
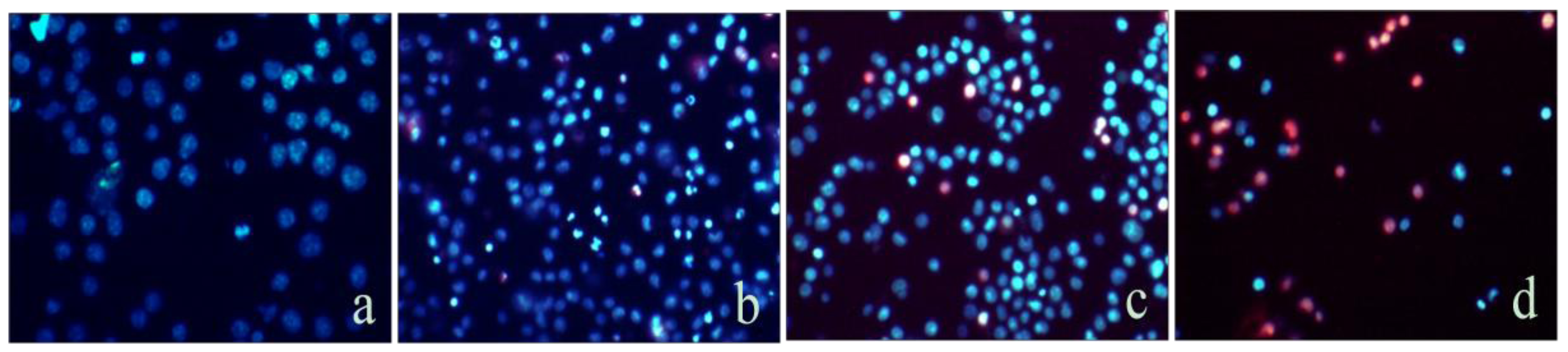
2.2. Mere15 Induces Apoptosis in K562 Cells
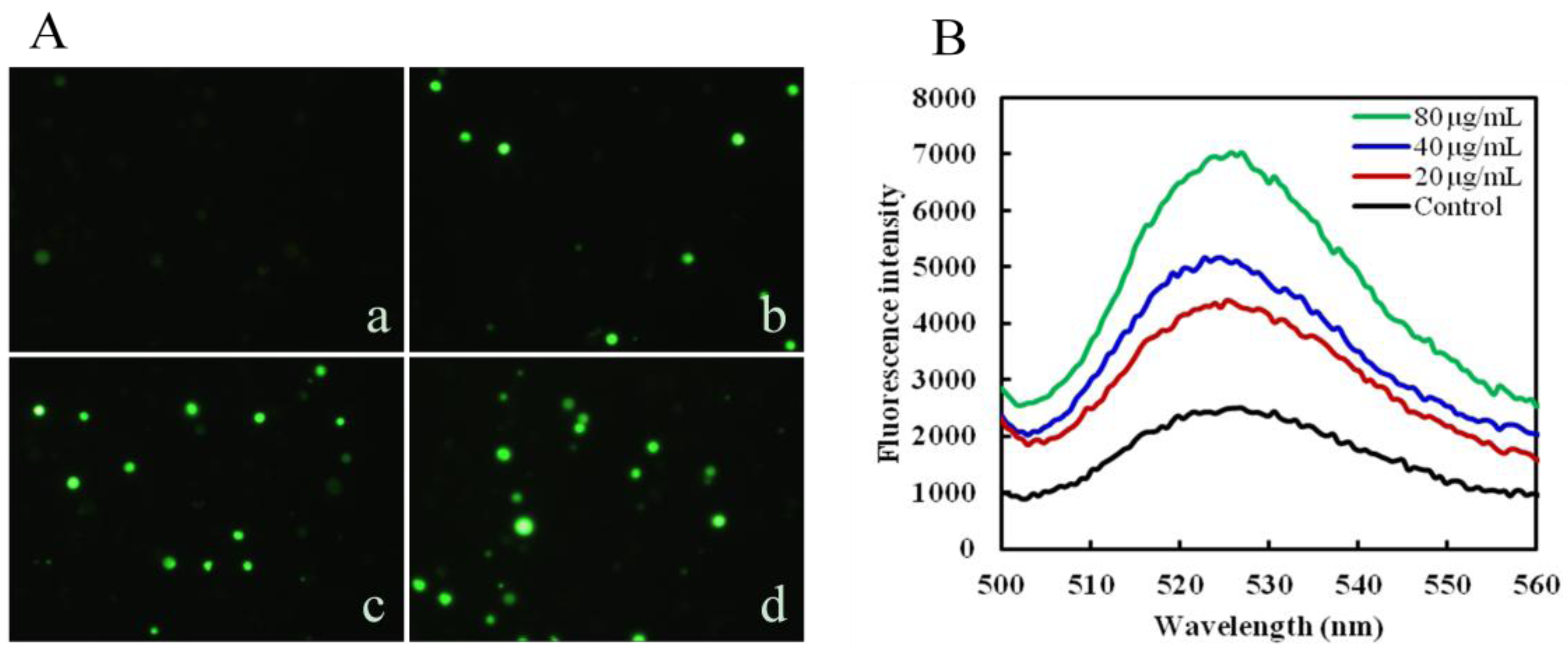
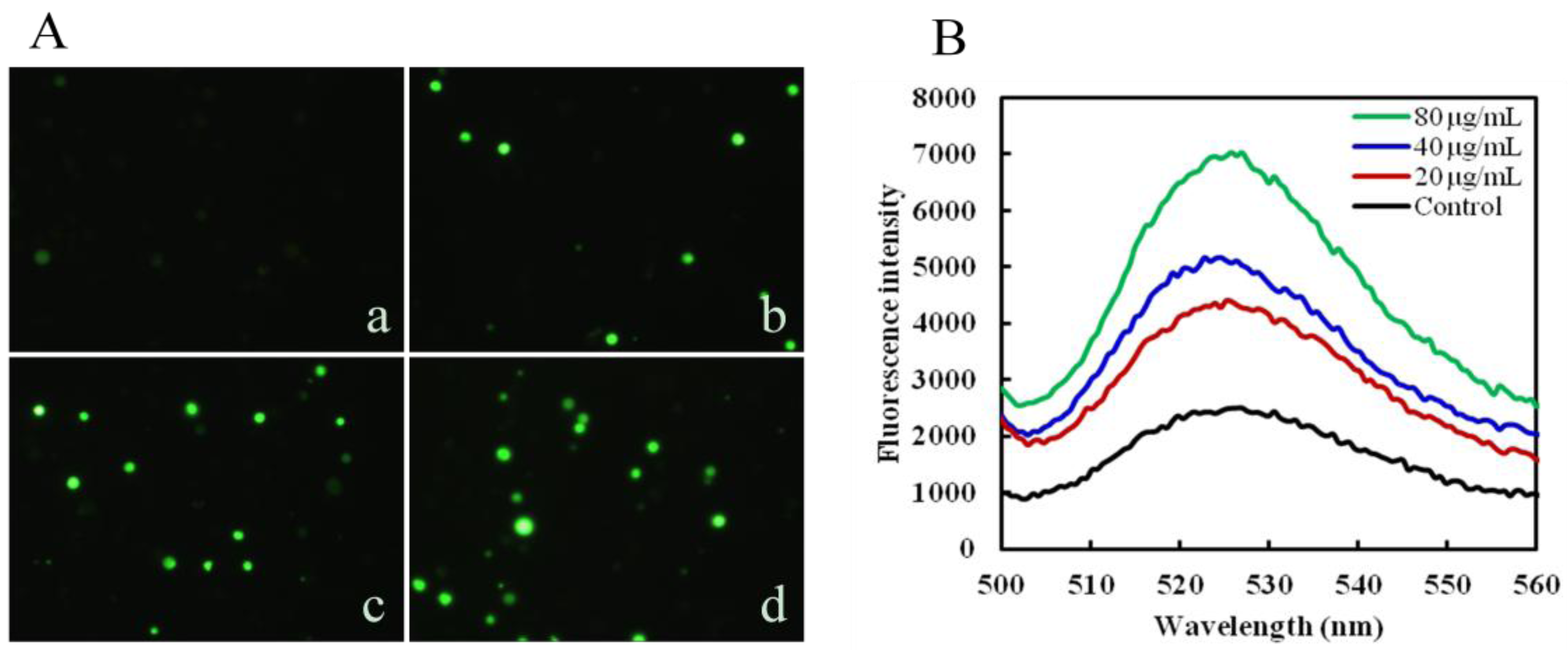
2.3. Mere15 Induces ROS Generation
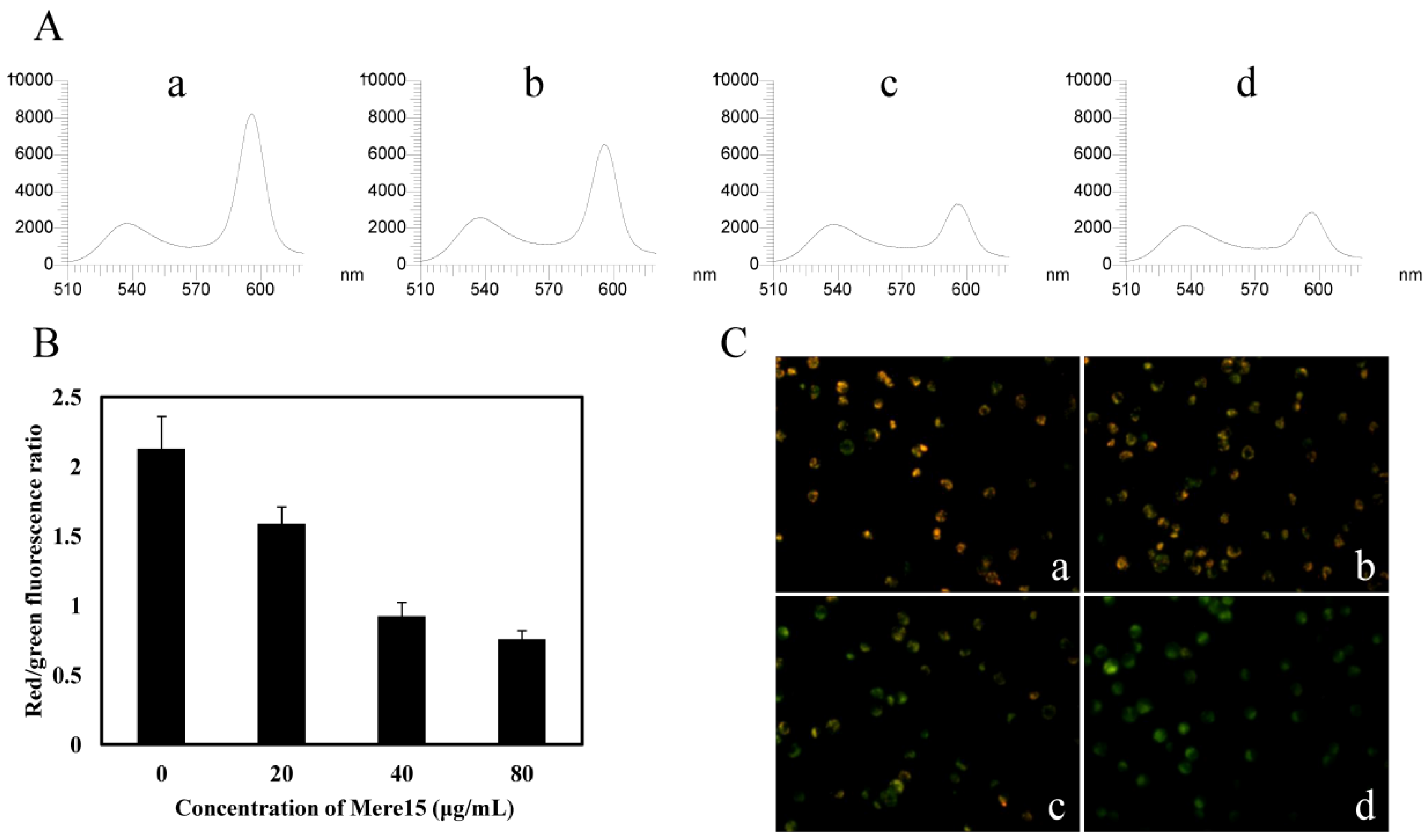
2.4. Mere15 Disturbs MMP
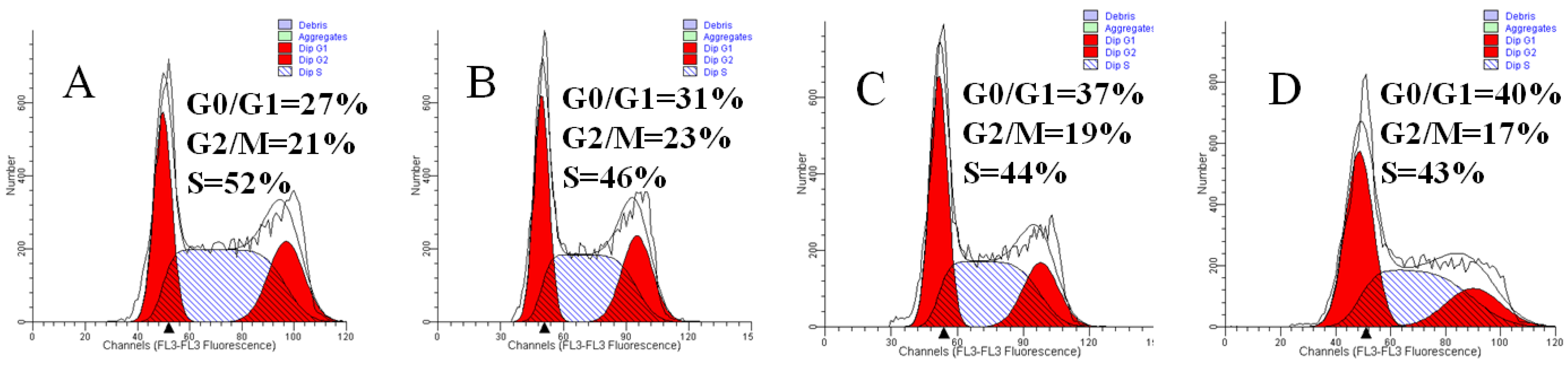
2.5. Mere15 Induces a G0/G1 Phase Arrest
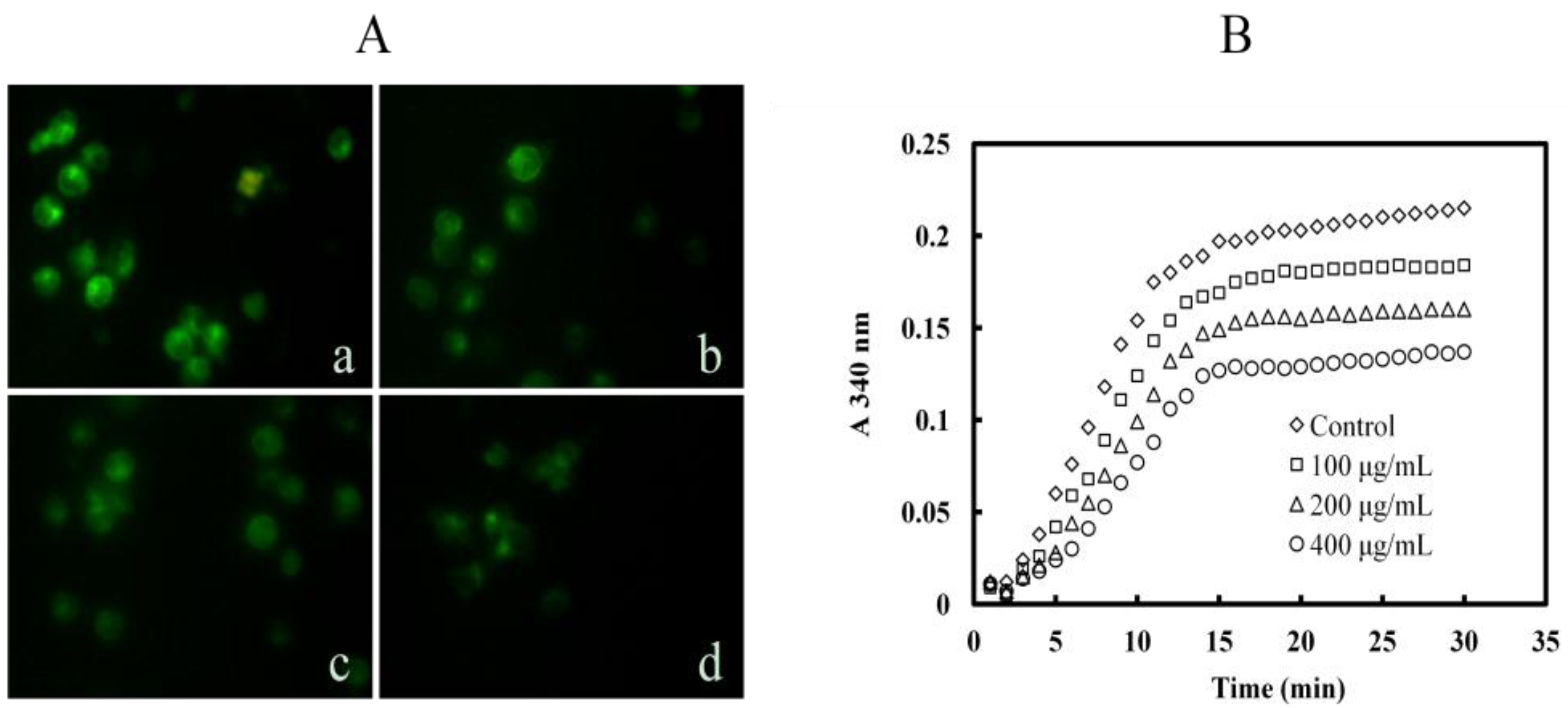
2.6. Mere15 Inhibits Tubulin Polymerization both in K562 Cells and in Cell Free System
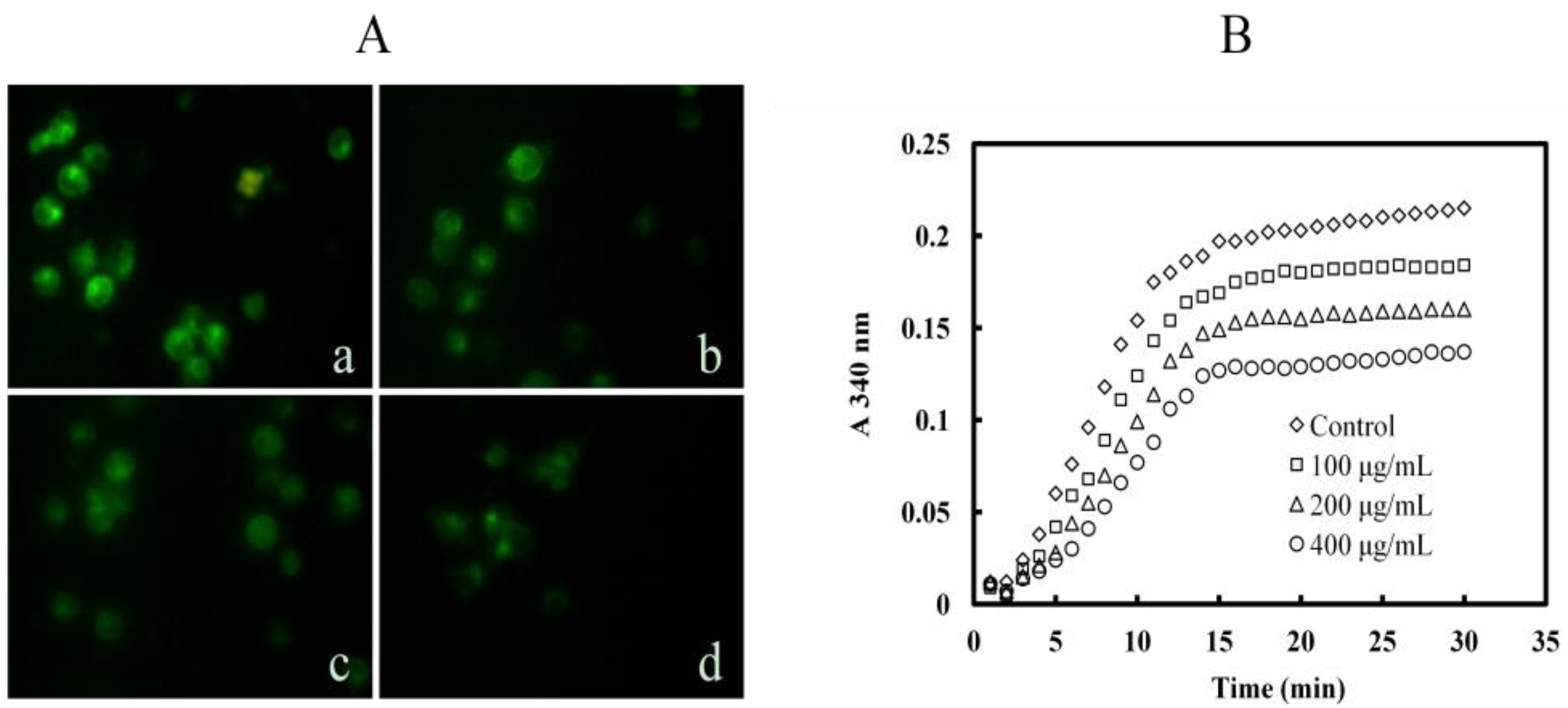
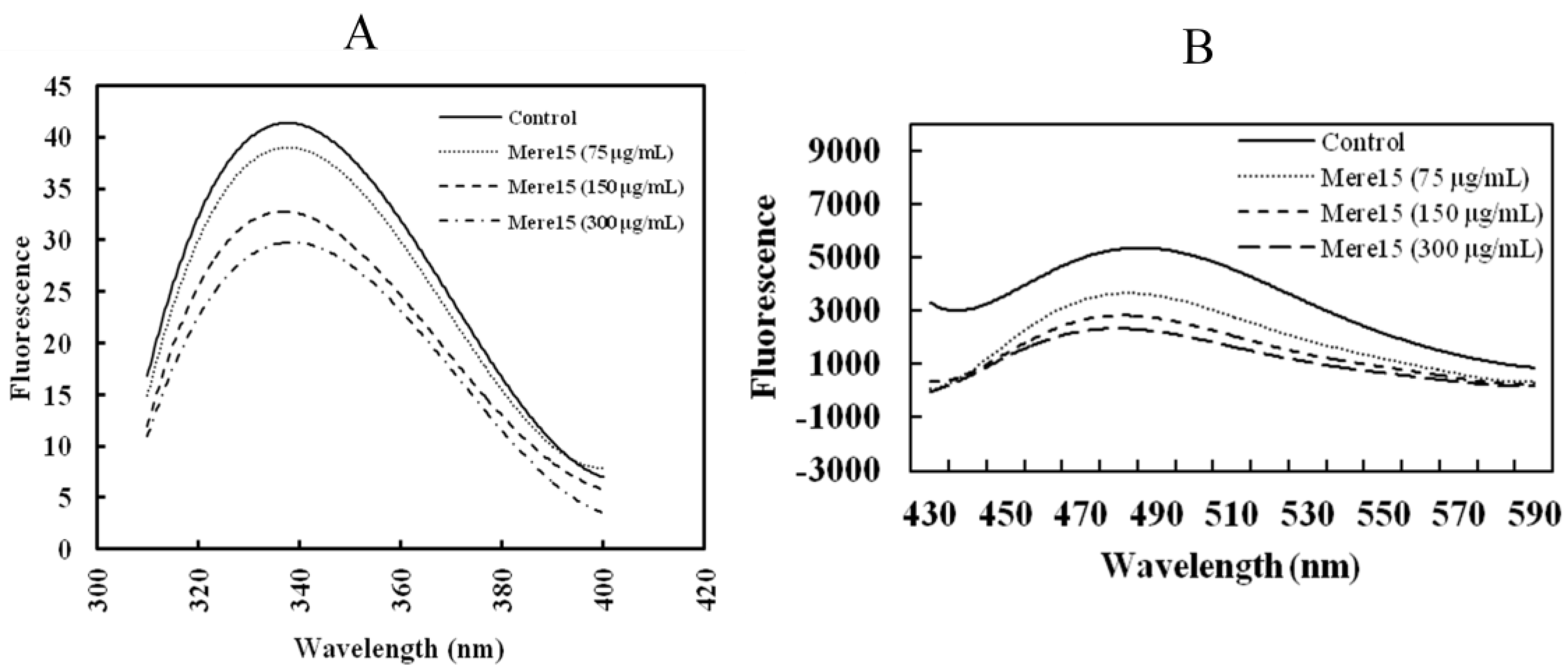
2.7. Mere15 Interacts with Tubulin in Vitro
3. Experimental Section
3.1. Materials and Reagents
3.2. Cell Lines and Cell Culture
3.3. Cell Viability Assessment
3.4. Hoechst33342/Propidium Iodide (PI) Dual Staining Assays
3.5. Measurement of Reactive Oxygen Species (ROS) Generation
3.6. Assessment of Mitochondrial Membrane Potential (MMP)
3.7. Cell Cycle Analysis
3.8. Immunofluorescence Detection of Cellular Microtubule
3.9. Tubulin Polymerization Assay
3.10. Intrinsic Tryptophan Fluorescence of Tubulin
3.11. Hydrophobic Analysis of Tubulin Using Bis-ANS
3.12. Data Analysis
4. Conclusions
Acknowledgments
References
- Jimeno, J.; Lopez-Martin, J.A.; Ruiz-Casado, A.; Izquierdo, M.A.; Scheuer, P.J.; Rinehart, K. Progress in the clinical development of new marine-derived anticancer compounds. Anticancer Drugs 2004, 15, 321–329. [Google Scholar] [CrossRef]
- Lin, X.; Liu, M.; Hu, C.; Liao, D.J. Targeting cellular proapoptotic molecules for developing anticancer agents from marine sources. Curr. Drug Targets 2010, 11, 708–715. [Google Scholar] [CrossRef]
- Zheng, L.H.; Wang, Y.J.; Sheng, J.; Wang, F.; Zheng, Y.; Lin, X.K.; Sun, M. Antitumor peptides from marine organisms. Mar. Drugs 2011, 9, 1840–1859. [Google Scholar] [CrossRef]
- Rawat, D.S.; Joshi, M.C.; Joshi, P.; Atheaya, H. Marine peptides and related compounds in clinical trial. Anticancer Agents Med. Chem. 2006, 6, 33–40. [Google Scholar] [CrossRef]
- Janmaat, M.L.; Rodriguez, J.A.; Jimeno, J.; Kruyt, F.A.E.; Giaccone, G. Kahalalide F induces necrosis-like cell death that involves depletion of ErbB3 and inhibition of Akt signaling. Mol. Pharmacol. 2005, 68, 502–510. [Google Scholar]
- Younes, A.; Yasothan, U.; Kirkpatrick, P. Brentuximab vedotin. Nat. Rev. Drug. Discov. 2012, 11, 19–20. [Google Scholar]
- Leng, B.; Liu, X.D.; Chen, Q.X. Inhibitory effects of anticancer peptide from Mercenaria on the BGC-823 cells and several enzymes. FEBS Lett. 2005, 579, 1187–1190. [Google Scholar] [CrossRef]
- Zhang, B.; Wu, J.L. Isolation and characterization of glycopeptide MGP0405 from Meretrix meretrix. Chin. J. Nat. Med. 2006, 4, 230–233. [Google Scholar]
- Wu, J.L.; Zhang, B.; Huang, C.H.; Zhu, X.C.; Wu, W.T. The antitumor activity of glycopeptide (MGP0501) from Meretrix meretrix in vitro. Pharm. Biotechnol. 2006, 13, 260–264. [Google Scholar]
- Wang, C.; Liu, M.; Cheng, L.; Wei, J.; Wu, N.; Zheng, L.; Lin, X. A novel polypeptide from Meretrix meretrix Linnaeus inhibits the growth of human lung adenocarcinoma. Exp. Biol. Med. 2012, 237, 442–450. [Google Scholar] [CrossRef]
- Wang, H.; Wei, J.T.; Wu, N.; Liu, M.; Wang, C.C.; Liu, H.Z.; Lin, X.K. Mere15, a novel polypeptide from Meretrix meretrix, inhibits adhesion, migration and invasion of human lung cancer A549 cells via down-regulating MMPs. Pharm. Biol. 2012, in press. [Google Scholar]
- Roychowdhury, S.; Talpaz, M. Managing resistance in chronic myeloid leukemia. Blood Rev. 2011, 25, 279–290. [Google Scholar] [CrossRef]
- Simon, H.U.; Haj-Yehia, A.; Levi-Schaffer, F. Role of reactive oxygen species (ROS) in apoptosis induction. Apoptosis 2000, 5, 415–418. [Google Scholar] [CrossRef]
- Higuchi, M.; Honda, T.; Proske, R.J.; Yeh, E.T. Regulation of reactive oxygen species-induced apoptosis and necrosis by caspase 3-like proteases. Oncogene 1998, 17, 2753–2760. [Google Scholar]
- Ozben, T. Oxidative stress and apoptosis: Impact on cancer therapy. J. Pharm. Sci. 2007, 96, 2181–2196. [Google Scholar] [CrossRef]
- Efferth, T.; Giaisi, M.; Merling, A.; Krammer, P.H.; Li-Weber, M. Artesunate induces ROS-mediated apoptosis in doxorubicin-resistant T leukemia cells. PLoS One 2007, 2, e693. [Google Scholar]
- Luo, M.; Liu, X.; Zu, Y.; Fu, Y.; Zhang, S.; Yao, L.; Efferth, T. Cajanol, a novel anticancer agent from Pigeonpea [Cajanus cajan (L.) Millsp.] roots, induces apoptosis in human breast cancer cells through a ROS-mediated mitochondrial pathway. Chem. Biol. Interact. 2010, 188, 151–160. [Google Scholar] [CrossRef]
- Martín, R.; Ibeas, E.; Carvalho-Tavares, J.; Hernández, M.; Ruiz-Gutierrez, V.; Nieto, M.L. Natural triterpenic diols promote apoptosis in astrocytoma cells through ROS-mediated mitochondrial depolarization and JNK activation. PLoS One 2009, 4, e5975. [Google Scholar]
- Cyr, L.; Langler, R.; Lavigne, C. Cell cycle arrest and apoptosis responses of human breast epithelial cells to the synthetic organosulfur compound p-Methoxyphenyl p-Toluenesulfonate. Anticancer Res. 2008, 28, 2753–2763. [Google Scholar]
- Huang, F.; Yang, Z.; Yu, D.; Wang, J.; Li, R.; Ding, G. Sepia ink oligopeptide induces apoptosis in prostate cancer cell lines via Caspase-3 activation and elevation of Bax/Bcl-2 ratio. Mar. Drugs 2012, 10, 2153–2165. [Google Scholar] [CrossRef]
- Wei, S.-Y.; Li, M.; Tang, S.-A.; Sun, W.; Xu, B.; Cui, J.-R.; Lin, W.-H. Induction of apoptosis accompanying with G1 phase arrest and microtubule disassembly in human hepatoma cells by jaspolide B, a new isomalabaricane-type triterpene. Cancer Lett. 2008, 262, 114–122. [Google Scholar]
- Hawe, A.; Sutter, M.; Jiskoot, W. Extrinsic fluorescent dyes as tools for protein characterization. Pharm. Res. 2008, 25, 1487–1499. [Google Scholar] [CrossRef]
- Yue, Q.X.; Liu, X.; Guo, D.A. Microtubule-binding natural products for cancer therapy. Planta Med. 2010, 76, 1037–1043. [Google Scholar]
- Tim, M. Rapid colorimetric assay for cellular growth and survival: Application to proliferation and cytotoxicity assays. J. Immunol. Methods 1983, 65, 55–63. [Google Scholar] [CrossRef]
- Ning, X.; Zhao, J.; Zhang, Y.; Cao, S.; Liu, M.; Ling, P.; Lin, X. A novel anti-tumor protein extracted from Meretrix meretrix Linnaeus induces cell death by increasing cell permeability and inhibiting tubulin polymerization. Int. J. Oncol. 2009, 35, 805–812. [Google Scholar]
- Kumar, S.; Sitasawad, S.L. N-acetylcysteine prevents glucose/glucose oxidase-induced oxidative stress, mitochondrial damage and apoptosis in H9c2 cells. Life Sci. 2009, 84, 328–336. [Google Scholar]
- Wang, Z.; Tang, X.; Li, Y.; Leu, C.; Guo, L.; Zheng, X.; Zhu, D. 20-Hydroxyeicosatetraenoic acid inhibits the apoptotic responses in pulmonary artery smooth muscle cells. Eur. J. Pharm. 2008, 588, 9–17. [Google Scholar] [CrossRef]
- Samples Availability: Available from the authors.
© 2012 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Liu, M.; Zhao, X.; Zhao, J.; Xiao, L.; Liu, H.; Wang, C.; Cheng, L.; Wu, N.; Lin, X. Induction of Apoptosis, G0/G1 Phase Arrest and Microtubule Disassembly in K562 Leukemia Cells by Mere15, a Novel Polypeptide from Meretrix meretrix Linnaeus. Mar. Drugs 2012, 10, 2596-2607. https://doi.org/10.3390/md10112596
Liu M, Zhao X, Zhao J, Xiao L, Liu H, Wang C, Cheng L, Wu N, Lin X. Induction of Apoptosis, G0/G1 Phase Arrest and Microtubule Disassembly in K562 Leukemia Cells by Mere15, a Novel Polypeptide from Meretrix meretrix Linnaeus. Marine Drugs. 2012; 10(11):2596-2607. https://doi.org/10.3390/md10112596
Chicago/Turabian StyleLiu, Ming, Xiangzhong Zhao, Jin Zhao, Lin Xiao, Haizhou Liu, Cuicui Wang, Linyou Cheng, Ning Wu, and Xiukun Lin. 2012. "Induction of Apoptosis, G0/G1 Phase Arrest and Microtubule Disassembly in K562 Leukemia Cells by Mere15, a Novel Polypeptide from Meretrix meretrix Linnaeus" Marine Drugs 10, no. 11: 2596-2607. https://doi.org/10.3390/md10112596
APA StyleLiu, M., Zhao, X., Zhao, J., Xiao, L., Liu, H., Wang, C., Cheng, L., Wu, N., & Lin, X. (2012). Induction of Apoptosis, G0/G1 Phase Arrest and Microtubule Disassembly in K562 Leukemia Cells by Mere15, a Novel Polypeptide from Meretrix meretrix Linnaeus. Marine Drugs, 10(11), 2596-2607. https://doi.org/10.3390/md10112596