Phorbol Ester Modulation of Ca2+ Channels Mediates Nociceptive Transmission in Dorsal Horn Neurones


Abstract
:1. Introduction
2. Experimental Section
2.1. Acute spinal Cord Slice Preparation and Solutions
2.2. Electrophysiological Recording
2.3. Pharmacology
3. Results and Discussion
3.1. Effect of PMA on Synaptic Transmission in Dorsal Horn Neurones


{kind=link}
{kind=link}
| sPSC amplitude (pA) | sPSC frequency (Hz) | sPSC half-width (ms) | |
|---|---|---|---|
| WT (n = 6) | 43.5 ± 4.2 | 0.69 ± 0.16 | 6.85 ± 0.57 |
| PMA (n = 6) | 41.8 ± 5.0 | 1.40 ± 0.36 * | 6.72 ± 0.47 |
| CaV2.3(−/−) (n = 8) | 57.3 ± 9.7 | 1.00 ± 0.15 | 7.26 ± 0.42 |
| PMA (n = 8) | 47.4 ± 5.3 | 1.08 ± 0.21 | 7.27 ± 0.40 |
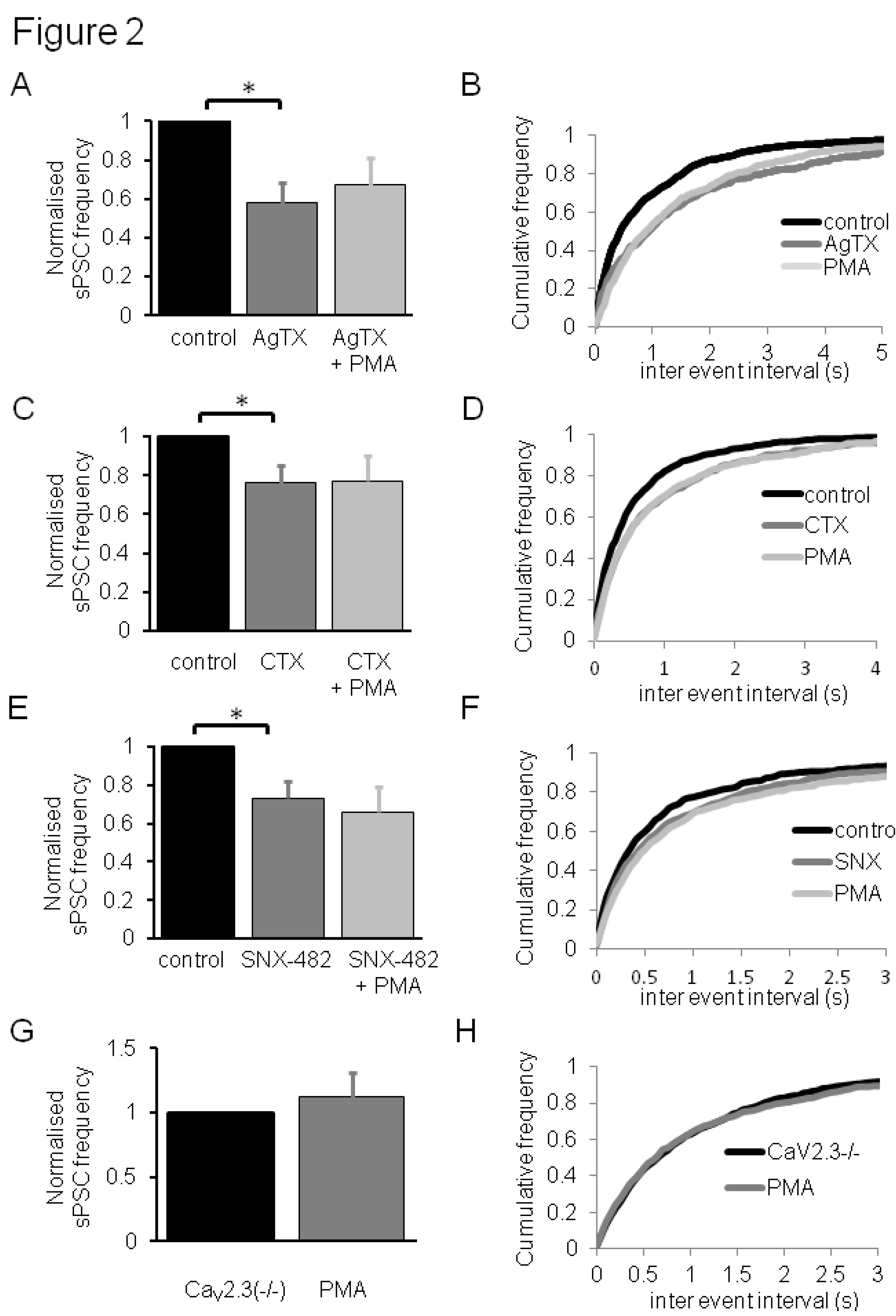
3.2. Contribution of CaV2 VDCC Subtypes to PMA Effects on Synaptic Transmission in Dorsal Horn Neurones
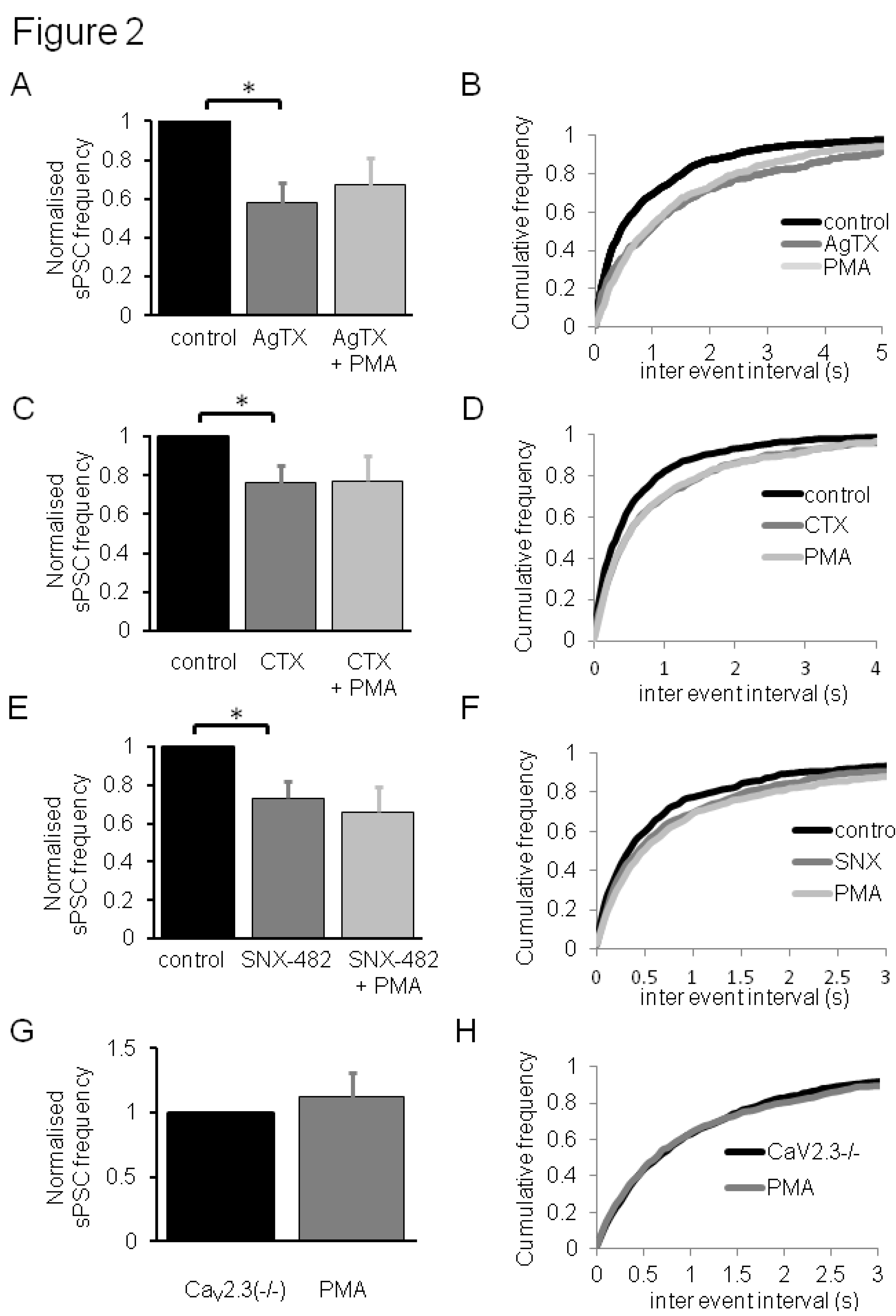
4. Conclusions
Acknowledgements
Conflict of Interest
References
- Velázquez, K.T.; Mohammad, H.; Sweitzer, S.M. Protein kinase C in pain: Involvement of multiple isoforms. Pharmacol. Res. 2007, 55, 578–589. [Google Scholar] [CrossRef]
- Zamponi, G.W.; Lewis, R.J.; Todorovic, S.M.; Arneric, S.P.; Snutch, T.P. Role of voltage-gated calcium channels in ascending pain pathways. Brain Res. Rev. 2009, 60, 84–89. [Google Scholar] [CrossRef]
- Park, J.; Luo, Z.D. Calcium channel functions in pain processing. Channels (Austin) 2010, 4, 510–517. [Google Scholar]
- Catterall, W.A. Structure and regulation of voltage-gated Ca2+ channels. Annu. Rev. Cell. Dev. Biol. 2000, 16, 521–555. [Google Scholar] [CrossRef]
- Ertel, E.A.; Campbell, K.P.; Harpold, M.M.; Hofmann, F.; Mori, Y.; Perez-Reyes, E.; Schwartz, A.; Snutch, T.P.; Tanabe, T.; Birnbaumer, L.; et al. Nomenclature of voltage-gated calcium channels. Neuron 2000, 25, 533–535. [Google Scholar] [CrossRef]
- Catterall, W.A.; Few, A.P. Calcium channel regulation and presynaptic plasticity. Neuron 2008, 59, 882–901. [Google Scholar] [CrossRef]
- Westenbroek, R.E.; Hoskins, L.; Catterall, W.A. Localization of Ca2+ channel subtypes on rat spinal motorneurones, interneurones, and nerve terminals. J. Neurosci. 1998, 18, 6319–6330. [Google Scholar]
- Murakami, M.; Nakagawasai, O.; Suzuki, T.; Mobarakeh, I.I.; Sakurada, Y.; Murata, A.; Yamadera, F.; Miyoshi, I.; Yanai, K.; Tan-No, K.; et al. Antinociceptive effect of different types of calcium channel inhibitors and the distribution of various calcium channel α1 subunits in the dorsal horn of spinal cord in mice. Brain Res. 2004, 1024, 122–129. [Google Scholar] [CrossRef]
- Castro, A.; Andrade, A.; Vergara, P.; Segovia, J.; Aguilar, J.; Felix, R.; Delgado-Lezama, R. Involvement of R-type Ca2+ channels in neurotransmitter release from spinal dorsolateral funiculus terminals synapsing motoneurones. J. Comp. Neurol. 2009, 513, 188–196. [Google Scholar] [CrossRef]
- Stea, A.; Soong, T.W.; Snutch, T.P. Determinants of PKC-dependent modulation of a family of neuronal calcium channels. Neuron 1995, 15, 929–940. [Google Scholar] [CrossRef]
- Hamid, J.; Nelson, D.; Spaetgens, R.; Dubel, S.; Snutch, T.P.; Zamponi, G.W. Identification of an integration center for cross-talk between protein kinase C and G protein modulation of N-type calcium channels. J. Biol. Chem. 1999, 274, 6195–6202. [Google Scholar]
- Fang, H.; Franke, R.; Patanavanich, S.; Lalvani, A.; Powell, N.K.; Sando, J.J.; Kamatchi, G.L. Role of alpha1 2.3 subunit I-II linker sites in the enhancement of CaV 2.3 current by phorbol 12-myristate 13-acetate and acetyl-beta-methylcholine. J. Biol. Chem. 2005, 280, 23559–23565. [Google Scholar]
- Fang, H.; Patanavanich, S.; Rajagopal, S.; Yi, X.; Gill, M.S.; Sando, J.J.; Kamatchi, G.L. Inhibitory role of Ser-425 of the alpha1 2.2 subunit in the enhancement of CaV 2.2 currents by phorbol-12-myristate, 13-acetate. J. Biol. Chem. 2006, 281, 20011–20017. [Google Scholar]
- Tedford, H.W.; Zamponi, G.W. Direct G protein modulation of CaV2 calcium channels. Pharmacol. Rev. 2006, 58, 837–862. [Google Scholar] [CrossRef]
- Díaz-Cárdenas, A.F.; Arenas, I.; García, D.E. PMA counteracts G protein actions on CaV2.2 channels in rat sympathetic neurones. Arch. Biochem. Biophys. 2008, 473, 1–7. [Google Scholar] [CrossRef]
- Kazanietz, M.G.; Caloca, M.J.; Eroles, P.; Fujii, T.; García-Bermejo, M.L.; Reilly, M.; Wang, H. Pharmacology of the receptors for the phorbol ester tumor promoters: multiple receptors with different biochemical properties. Biochem. Pharmacol. 2000, 60, 1417–1424. [Google Scholar] [CrossRef]
- Yang, L.; Stephens, G.J. Effects of neuropathy on high-voltage-activated Ca2+ current in sensory neurones. Cell. Calcium 2009, 46, 248–256. [Google Scholar] [CrossRef]
- Matthews, E.A.; Dickenson, A.H. Effects of spinally delivered N- and P-type voltage-dependent calcium channel antagonists on dorsal horn neuronal responses in a rat model of neuropathy. Pain 2001, 92, 235–246. [Google Scholar] [CrossRef]
- Matthews, E.A.; Bee, L.A.; Stephens, G.J.; Dickenson, A.H. The CaV2.3 calcium channel antagonist SNX-482 reduces dorsal horn neuronal responses in a rat model of chronic neuropathic pain. Eur. J. Neurosci. 2007, 25, 3561–3569. [Google Scholar] [CrossRef]
- Pereverzev, A.; Mikhna, M.; Vajna, R.; Gissel, C.; Henry, M.; Weiergräber, M.; Hescheler, J.; Smyth, N.; Schneider, T. Disturbances in glucose-tolerance, insulin-release and stress-induced hyperglycemia upon disruption of the CaV2.3 (α1E) subunit of voltage-gated Ca2+ channels. Mol. Endocrinol. 2002, 16, 884–895. [Google Scholar] [CrossRef]
- Yashpal, K.; Pitcher, G.M.; Parent, A.; Quirion, R.; Coderre, T.J. Noxious thermal and chemical stimulation induce increases in 3H-phorbol 12,13-dibutyrate binding in spinal cord dorsal horn as well as persistent pain and hyperalgesia, which is reduced by inhibition of protein kinase C. J. Neurosci. 1995, 15, 3263–3272. [Google Scholar]
- Kamatchi, G.L.; Tiwari, S.N.; Chan, C.K.; Chen, D.; Do, S.H.; Durieux, M.E.; Lynch, C., III. Distinct regulation of expressed calcium channels 2.3 in Xenopus oocytes by direct or indirect activation of protein kinase C. Brain Res. 2003, 968, 227–237. [Google Scholar] [CrossRef]
- Lu, C.R.; Willcockson, H.H.; Phend, K.D.; Lucifora, S.; Darstein, M.; Valtschanoff, J.G.; Rustioni, A. Ionotropic glutamate receptors are expressed in GABAergic terminals in the rat superficial dorsal horn. J. Comp. Neurol. 2005, 486, 169–178. [Google Scholar] [CrossRef]
- Sikand, P.; Premkumar, L.S. Potentiation of glutamatergic synaptic transmission by protein kinase C-mediated sensitization of TRPV1 at the first sensory synapse. J. Physiol. 2007, 581, 631–647. [Google Scholar] [CrossRef]
- Cao, D.S.; Yu, S.Q.; Premkumar, L.S. Modulation of transient receptor potential vanilloid 4-mediated membrane currents and synaptic transmission by protein kinase C. Mol. Pain 2009, 5, 5. [Google Scholar] [CrossRef]
- Silinsky, E.M.; Searl, T.J. Phorbol esters and neurotransmitter release: more than just protein kinase C? Br. J. Pharmacol. 2003, 138, 1191–1201. [Google Scholar] [CrossRef]
- Lou, X.; Korogod, N.; Brose, N.; Schneggenburger, R. Phorbol esters modulate spontaneous and Ca2+-evoked transmitter release via acting on both Munc13 and protein kinase C. J. Neurosci. 2008, 28, 8257–8267. [Google Scholar] [CrossRef]
- Saegusa, H.; Kurihara, T.; Zong, S.; Kazuno, A.; Matsuda, Y.; Nonaka, T.; Han, W.; Toriyama, H.; Tanabe, T. Suppression of inflammatory and neuropathic pain symptoms in mice lacking the N-type Ca2+ channel. EMBO J. 2001, 20, 2349–2356. [Google Scholar] [CrossRef]
- Bao, J.; Li, J.J.; Perl, E.R. Differences in Ca2+ channels governing generation of miniature and evoked excitatory synaptic currents in spinal laminae I and II. J. Neurosci. 1998, 18, 8740–8750. [Google Scholar]
- Heinke, B.; Balzer, E.; Sandkühler, J. Pre- and postsynaptic contributions of voltage-dependent Ca2+ channels to nociceptive transmission in rat spinal lamina I neurones. Eur. J. Neurosci. 2004, 19, 103–111. [Google Scholar] [CrossRef]
- Rycroft, B.K.; Vikman, K.S.; Christie, M.J. Inflammation reduces the contribution of N-type calcium channels to primary afferent synaptic transmission onto NK1 receptor-positive lamina I neurones in the rat dorsal horn. J. Physiol. 2007, 580, 883–894. [Google Scholar] [CrossRef]
- Heinke, B.; Gingl, E.; Sandkühler, J. Multiple targets of μ-opioid receptor-mediated presynaptic inhibition at primary afferent Aδ- and C-fibers. J. Neurosci. 2011, 31, 1313–1322. [Google Scholar]
- Jacus, M.O.; Uebele, V.N.; Renger, J.J.; Todorovic, S.M. Presynaptic Cav3.2 channels regulate excitatory neurotransmission in nociceptive dorsal horn neurones. J. Neurosci. 2012, 32, 9374–9382. [Google Scholar] [CrossRef]
- Santafé, M.M.; Lanuza, M.A.; Garcia, N.; Tomàs, J. Calcium inflow-dependent protein kinase C activity is involved in the modulation of transmitter release in the neuromuscular junction of the adult rat. Synapse 2005, 57, 76–84. [Google Scholar] [CrossRef]
- Santafé, M.M.; Lanuza, M.A.; Garcia, N.; Tomàs, J. Muscarinic autoreceptors modulate transmitter release through protein kinase C and protein kinase A in the rat motor nerve terminal. Eur. J. Neurosci. 2006, 23, 2048–2056. [Google Scholar] [CrossRef]
- Harvey, V.L.; Stephens, G.J. Mechanism of GABA receptor-mediated inhibition of spontaneous GABA release onto cerebellar Purkinje cells. Eur. J. Neurosci. 2004, 20, 684–700. [Google Scholar] [CrossRef]
- Stephens, G.J.; Morris, N.P.; Fyffe, R.E.; Robertson, B. The CaV2.1/α1A (P/Q-type) voltage-dependent calcium channel mediates inhibitory neurotransmission onto mouse cerebellar Purkinje cells. Eur. J. Neurosci. 2001, 13, 1902–1912. [Google Scholar] [CrossRef]
- Newcomb, R.; Szoke, B.; Palma, A.; Wang, G.; Chen, Xh.; Hopkins, W.; Cong, R.; Miller, J.; Urge, L.; Tarczy-Hornoch, K.; et al. Selective peptide antagonist of the class E calcium channel from the venom of the tarantula Hysterocrates gigas. Biochemisrty 1998, 37, 15353–15362. [Google Scholar] [CrossRef]
- Arroyo, G.; Aldea, M.; Fuentealba, J.; Albillos, A.; García, A.G. SNX482 selectively blocks P/Q Ca2+ channels and delays the inactivation of Na+ channels of chromaffin cells. Eur. J. Pharmacol. 2003, 475, 11–18. [Google Scholar] [CrossRef]
- Saegusa, H.; Kurihara, T.; Zong, S.; Minowa, O.; Kazuno, A.; Han, W.; Matsuda, Y.; Yamanaka, H.; Osanai, M.; Noda, T.; et al. Altered pain responses in mice lacking 1E subunit of the voltage-dependent Ca2+ channel. Proc. Natl. Acad. Sci. USA 2000, 97, 6132–6137. [Google Scholar] [CrossRef]
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Yang, L.; Topia, I.; Schneider, T.; Stephens, G.J. Phorbol Ester Modulation of Ca2+ Channels Mediates Nociceptive Transmission in Dorsal Horn Neurones. Pharmaceuticals 2013, 6, 777-787. https://doi.org/10.3390/ph6060777
Yang L, Topia I, Schneider T, Stephens GJ. Phorbol Ester Modulation of Ca2+ Channels Mediates Nociceptive Transmission in Dorsal Horn Neurones. Pharmaceuticals. 2013; 6(6):777-787. https://doi.org/10.3390/ph6060777
Chicago/Turabian StyleYang, Li, Iqbal Topia, Toni Schneider, and Gary J. Stephens. 2013. "Phorbol Ester Modulation of Ca2+ Channels Mediates Nociceptive Transmission in Dorsal Horn Neurones" Pharmaceuticals 6, no. 6: 777-787. https://doi.org/10.3390/ph6060777