Expanding the Concept of G Protein-Coupled Receptor (GPCR) Dimer Asymmetry towards GPCR-Interacting Proteins

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
: G protein-coupled receptors (GPCRs), major targets of drug discovery, are organized in dimeric and/or oligomeric clusters. The minimal oligomeric unit, the dimer, is composed of two protomers, which can behave differently within the dimer. Several examples of GPCR asymmetry within dimers at the level of ligand binding, ligand-promoted conformational changes, conformational changes within transmembrane domains, G protein coupling, and most recently GPCR-interacting proteins (GIPs), have been reported in the literature. Asymmetric organization of GPCR dimers has important implications on GPCR function and drug design. Indeed, the extension of the “asymmetry concept” to GIPs adds a new level of specific therapeutic intervention.1. Introduction
A growing body of pharmacological, biochemical and biophysical data indicate that GPCRs form functional homo- and hetero-dimers and most likely higher-order oligomers [1,2]. Formation of such oligomeric structures has been shown to provide shielding from the quality-control checkpoints during the biosynthetic pathway [3] and to have an important role in modulating GPCR function and signaling [4]. In addition, this oligomeric organization raises a fundamental question: does each protomer within a GPCR oligomer operate as an independent functional unit or do the different protomers intercommunicate to insure the signaling response of the cell? Communication within dimers has indeed been observed for several GPCRs at different levels. Ligand binding to one protomer has been shown to modify the properties of the ligand binding pocket of the second protomer. The latter observation most likely involves ligand-induced conformational changes that are transmitted from one protomer to the other. Intercommunication among the different protomers results in dimer asymmetry and asymmetric recruitment of GPCR-interacting proteins (GIPs) including heterotrimeric G proteins. This mini-review summarizes the evidence supporting the emerging concept of asymmetric behavior within a GPCR complex. We will briefly discuss asymmetry in G protein coupling, ligand binding, conformational changes within transmembrane domains, and binding of GIPs.
2. Asymmetry at the Level of G Protein Binding
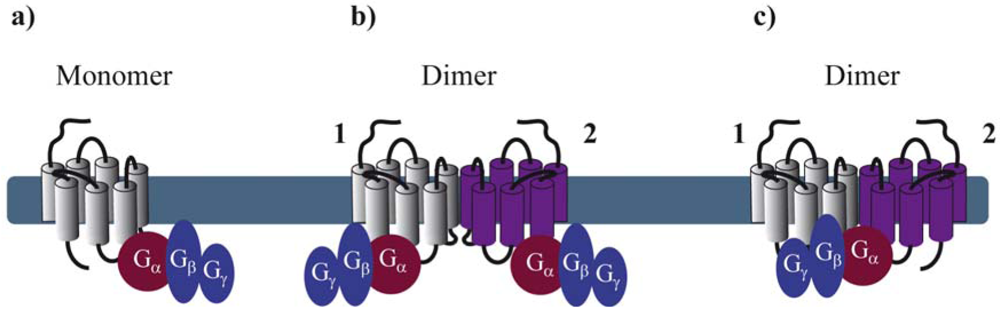
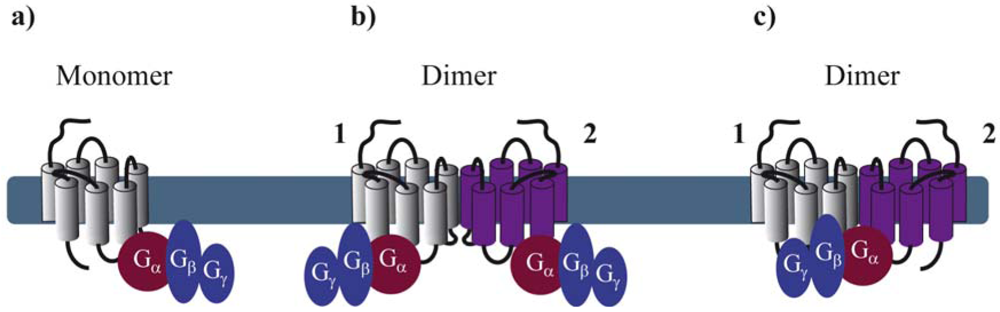
Reconstitution of purified monomeric GPCRs, in high-density lipoprotein phospholipid bilayer particles, showed that monomeric GPCRs are able to couple to G proteins. These and similar results clearly demonstrate that the minimal functional unit consists of 1 receptor monomer activating its cognate G protein (Figure 1a) [5-7]. When rod outer segment membranes were solubilized with different detergents to obtain preparations enriched in monomeric, dimeric and oligomeric rhodopsin structures, monomeric rhodopsin was indeed able to activate its G protein, the heterotrimeric transducin (Gt) confirming studies in reconstituted systems [8]. However, preparations containing mostly dimeric and oligomeric rhodopsin appeared to be more active.
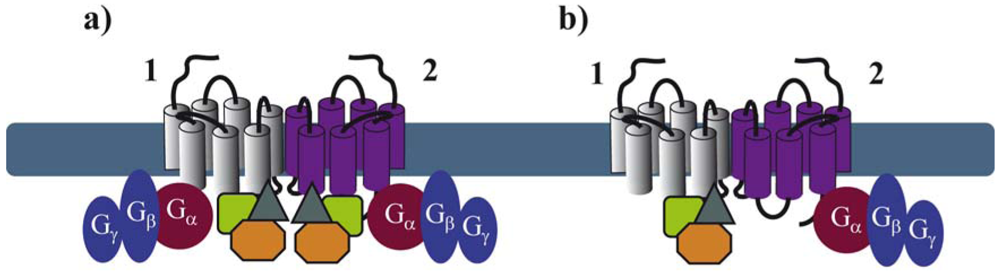
By extrapolating the idea that a GPCR monomer couples to a single G protein, one might anticipate that GPCR dimers, composed of two protomers, bind to two G proteins (Figure 1b). Nevertheless, to our knowledge, there is no clear experimental evidence today confirming such an arrangement, except one study published in 2008 by Parker et al. on the neuropeptide Y (NPY) Y2 receptor. Using radiolabeled-ligand binding assays, GTPγS binding and gradient ultracentrifugation experiments, the authors show that Y2 receptors and G proteins could be solubilized in complexes of 2:2, 2:1 and 1:1 stoichiometry, depending on the detergent and the concentration of agonist used [9]. Cumulative evidence from the literature is, however, rather consistent with a 2:1 stoichiometry with two receptor protomers coupling only one heterotrimeric G protein (Figure 1c). Such a stoichiometry implies that GPCR dimers have a radically different mode of function than monomers therefore introducing the concept of asymmetry.
GPCR asymmetry was intensely debated when the first high-resolution structure of rhodopsin, the archetypal class A GPCR, was solved by Palczewski and associates [10]. Computational docking simulations and molecular modeling of rhodopsin and its Gt protein illustrated that the receptor-facing surface of Gt is too large with respect to the rhodopsin cytoplasmic surface. These results suggest that GPCR dimer formation is a pre-requisite to fully accommodate the binding of the G protein [11,12]. Concomitantly, a 2:1 stoichiometry was proposed for another class A GPCR, the leukotriene B4 receptors BLT1 and its cognate G protein, by using purified proteins and chemical cross-linking [13].
These pioneering studies raised the fundamental question of the respective role of each protomer within a dimer in regards to G protein activation. Indeed, there are at least two ways for the G protein to contact the dimer (through protomer 1 or 2). Based on this assumption, asymmetry of the receptor dimer might be dictated by the position of the Gα subunit relative to the agonist-occupied monomer. A possible model would be that activation of one of the protomers favors an oriented interaction of the receptor dimer with the G protein in such a way that the agonist-occupied protomer directly interacts with the G protein and is therefore stabilized in a fully active, high affinity conformation, whereas the other protomer remains in an uncoupled, low affinity conformation [14]. Further important progress on how the receptor dimer interacts with the G protein has been made using BLT1 and dopamine D2 homodimers as models. By using the intrinsic fluorescence properties of the BLT1 receptor to monitor its activation, Damian et al. showed that a receptor dimer with only a single agonist-occupied subunit can trigger G protein activation. Interestingly, the two subunits of the dimer in the G protein-coupled state differ in their conformation, even when both protomers are occupied by agonists. In the absence of G protein coupling, no such asymmetric conformational changes were observed [15]. These results suggest that the interaction of the G protein with the receptor dimer would bring specific constraints, which prevent a symmetric functioning of the dimer and that the G protein itself might be partly responsible for the asymmetric functioning in a context where ligand binding cis-activates G protein binding to the same protomer [14]. On the other hand, using an original approach based on a functional complementation assay with the D2 receptor, allosteric communication between protomers of GPCR dimers was proposed [16]. This study confirmed that agonist binding to a single protomer within the dimer can maximally activate the G protein. Moreover, activation of the second protomer inhibits the functional response whereas inverse agonist binding enhances signaling, definitely proving that the way the two protomers contribute to the activated complex with the G protein is not symmetric, and that activation requires different conformational changes in each protomer [16]. Such asymmetry also applies to class C GPCRs as exemplified by the GABAB receptor, which is an obligatory heteromer composed of GB1 and GB2 subunits, with GB1 binding the ligand and GB2 activating the G protein [17,18].
Collectively, the most convincing examples in the literature favor a 2:1 stoichiometry between the receptor and the G protein. Several studies using purified and reconstituted GPCRs suggest a 1:1 stoichiometry. However, one should be careful when extrapolating these in vitro observations into a more physiological context; i.e. the plasma membrane of mammalian cells. Asymmetric behavior of GPCR dimers is further supported by observations at the level of ligand binding and ligand-promoted conformational changes within the heptahelical transmembrane domain as discussed in the next paragraphs.
3. Asymmetry at the Level of Ligand Binding
The phenomenon of allostery has been observed in many biological processes, and refers to different, albeit related, mechanisms by which protein function can be regulated and fine-tuned in either a positive or negative direction [19]. In pharmacology, allosteric modulation of a receptor results from the binding of allosteric molecules at a different site (allosteric site) from that of the endogenous ligand (orthosteric site). Allosteric modulators normally induce a conformational change in the receptor resulting in a positive or negative effect on binding affinity and efficacy of ligand binding to the orthosteric site. In the context of GPCR oligomerization, each protomer in the complex possesses an orthosteric binding pocket and one can expect that the orthosteric site of one protomer could exert allosteric effects on the orthosteric site of another protomer [20].
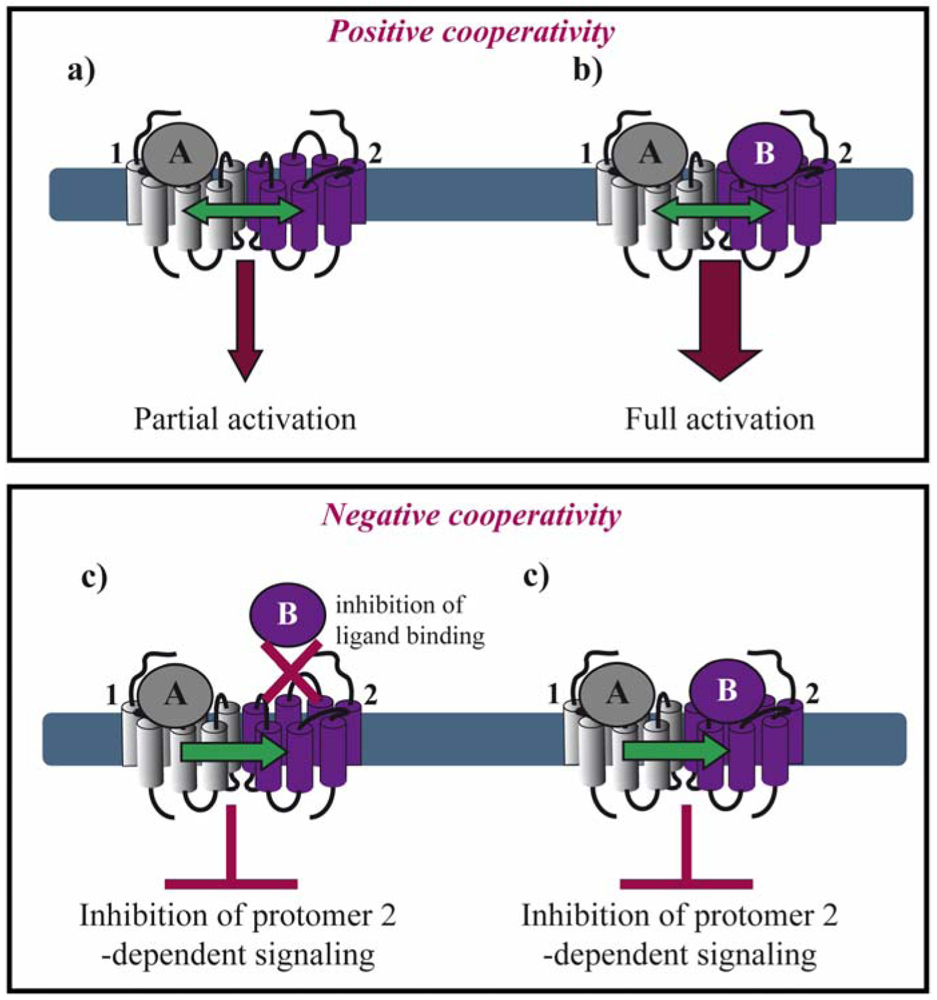
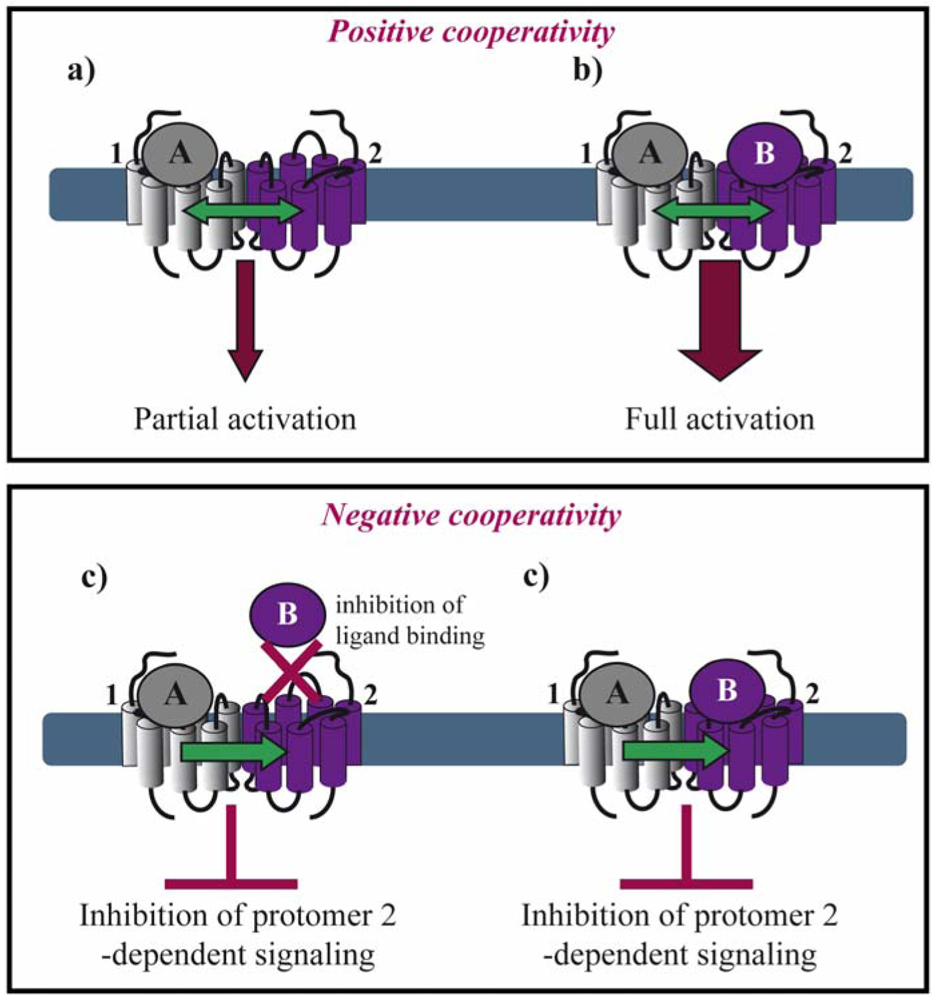
Allosteric interactions between ligand binding sites within a dimeric complex result in either positive or negative cooperativity. This fact raises the question of stoichiometry concerning the number of ligand molecules binding a dimer. The majority of the examples in the literature suggest that negative cooperativity is due to a single ligand molecule binding one protomer of the dimer, whereas positive cooperativity results from two ligand molecules binding, each, one protomer in a dimer. However, exceptions from this rule exist as mentioned below.
Positive cooperativity has early on been reported for several GPCRs. In the case of agonist binding to the μ/δ opioid receptor (OR) heteromer, both agonists are required for efficient MAP kinase activation (Figures 2a,b) [21]. Similarly, activation of both protomers of a muscarinic M3 homomer is required for beta-arrestin recruitment [22]. Positive cooperative binding was also documented for class C GPCRs. Binding of a single agonist is sufficient for the activation of the homomeric metabotropic glutamate receptor (mGluR) 5. However, full activation requires binding of two agonist molecules and the closure of both agonist-binding domains of the homodimer, also called venus fly trap (VFT) [23]. Another type of positive allosterism is seen in the CXCR2/δOR heteromer, where CXCR2 antagonists enhance the function of both peptide and alkaloid-based agonists at the δOR when the two receptors are co-expressed [24].
Negative binding cooperativity of allosteric nature has been observed in other cases such as homo-and heteromers of glycoprotein hormone receptors. Ligand binding to the large extracellular ectodomain triggers activation of constitutive receptor dimers that are mainly stabilized by interactions between the transmembrane domains. Using competition, saturation and dissociation kinetics binding experiments, Urizar et al. showed for the first time for class A GPCRs that the two orthosteric sites of each dimer are implicated in negative allosteric interactions with one another (Figure 2c) [25]. Similarly, an extensive pharmacological characterization of the dopamine D2 receptor by performing saturation binding assays and dissociation kinetics with three different radioligands showed that some D2 receptor antagonists exhibit negative binding cooperativity within a D2 homodimer [26]. Furthermore, chemokine receptor heteromers display allosteric modulation in ligand binding and dissociation kinetic experiments strongly suggesting negative cooperativity for several different heteromers. Trans-inhibition of ligand binding was observed between CCR5 and CCR2b co-expressed in the same cell line and in cells endogenously expressing both receptors [27,28]. CCR5-specific ligands, unable to compete for the binding of CCR2-specific ligands on cells expressing CCR2 alone, inhibited this binding when both receptors were co-expressed. Similar results were obtained with CCR2-selective ligands.
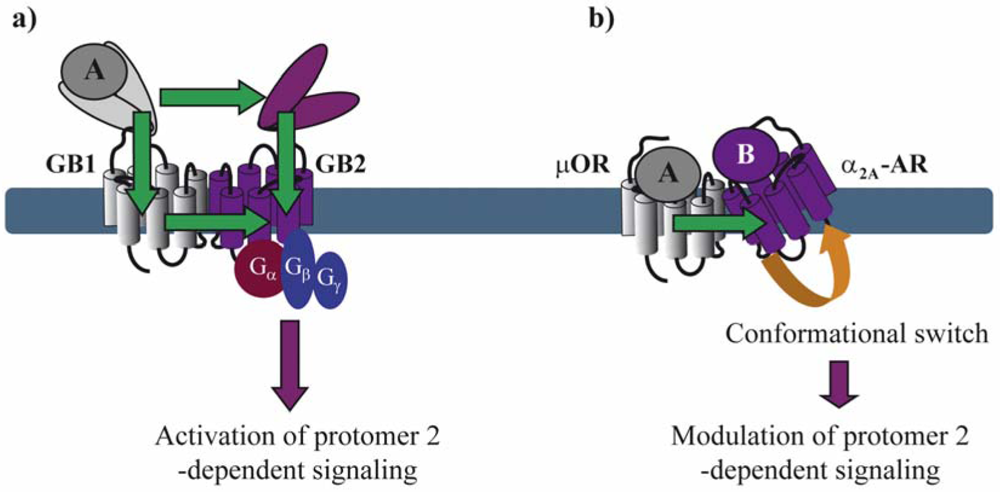
Moreover, in the context of the μOR/α2A-adrenoceptor (AR) heteromer, Vilardaga et al. showed that simultaneous binding of the two respective agonists, norepinephrine and morphine, inhibits α2A-AR-dependent Gi signaling and the downstream MAP kinase cascade [29]. These results interestingly show that negative cooperativity may occur in a dimer where both binding pockets are occupied by their respective agonists (Figure 2d).
Recently, Albizu et al. used fluorescent-labeled ligands to study oxytocine receptor dimerization in native tissues. A mixture of two antagonists, either labeled with an energy donor or acceptor, generated a strong FRET signal. However, only marginal FRET signals were obtained when two labeled agonists were used indicating that agonist binding to one protomer suppresses agonist binding to the second protomer whereas no such inhibitory effect is seen for antagonist binding [30]. Both positive and negative cooperativity have also been reported within the same heteromer as illustrated by the dopamine D2 receptor and somatostatin sst5 receptor heteromer, for which the positive cooperativity was observed for D2 agonists and negative cooperativity for D2 antagonists [31].
Collectively, different types of negative and positive allosteric modulations can be observed and may involve binding of one or two agonists or antagonists to the respective GPCR homo- or heteromer. In 2005, Durroux proposed mathematical models attempting to explain the complex allosteric interactions observed between ligand binding sites within dimeric complexes. [32].
4. Asymmetry at the Level of Ligand-Induced Conformation Changes
With the exception of neutral antagonists, binding of all other types of ligands ranging from full inverse agonists to full agonists induce conformational changes in GPCRs. Negative and positive cooperativity at the level of ligand binding is further propagated to the subsequent conformational changes within the dimer. Such cross-conformational switches between the two protomers of GPCR dimers have been observed in some cases (Figure 3). In the μOR/α2A-AR heteromer, morphine binding to the μOR triggers an inhibitory conformational change with subsecond kinetics in the norepinephrine-occupied α2A-AR as monitored directly by FRET in living cells [29].
Asymmetric ligand-induced conformational changes within GPCR dimers have been fully characterized in the GABAB receptor. Within this obligatory heterodimer, GB1 and GB2 subunits accomplish different tasks. The role of the GB2 is to traffic GB1 to the membrane where the VFT of GB1 binds to GABA and trans-activates GB2, which in turn couples to Gi proteins [33]. By analyzing chimeric and deletion constructs of GB1 and GB2, the authors show that several different conformational changes are necessary for full G protein activation. GABA-induced changes in the relative position of the VFTs of GB1 and GB2 activate the transmembrane (TM) portion of GB2 most likely through two allosteric pathways: (1) one direct from the GB2 VFT to the GB2 TM portion and (2) a second one that interconnects the GB1 VFT to GB1 TM portion, which, in turn, trans-activates GB2 TM portion [17,34].
Recently, Damian et al. monitored conformational changes within purified BLT1 receptor dimers, which were labeled with a single 5-hydroxytryptophan [15]. They showed that a receptor dimer with only a single agonist-occupied subunit can trigger G protein cis-activation. Interestingly, asymmetric conformational changes were observed even when both protomers were occupied by the same agonist. This asymmetry was highly dependent on the coupling of the dimer to purified G proteins. Altogether, asymmetric ligand-induced conformational changes have been monitored with several different techniques using different GPCRs either in living cells or as purified proteins.
5. Asymmetry at the Level of GPCR-Interacting Proteins (GIPs)
In addition to heterotrimeric G proteins, most GPCRs also bind to other intracellular proteins either directly or indirectly forming large protein complexes as exemplified by the melatonin MT1 and MT2 receptors [35,36]. Among those proteins that directly interact with GPCRs, GPCR kinases (GRKs) and arrestins are probably the most studied. GPCR signaling is believed to involve the sequential interaction of the activated receptor with the G protein, GRKs and arrestin. In contrast to this current model, recent observations made with energy transfer techniques however suggest that these different events might be interconnected and might overlap in time [37]. Recruitment of GRK2 to the α2A-AR, for instance, was shown to occur before dissociation of the G protein, therefore raising the question of the exact molecular organization of this transient protein complex. A first hint of the putative organization of such a complex came from crystallographic data of the GRK2/Gαq/Gβ1γ2 complex. These data showed that GRK2 can bind Gβγ and Gαq subunits simultaneously while maintaining a surface for a possible interaction with the receptor [38].
Arrestin is believed to be recruited after the first wave of G protein-dependent signaling events and to compete with G proteins and other GIPs for receptor binding. However, recent data on the parathyroid hormone receptor (PTHR) suggest that at least part of the arrestin binding might be faster than anticipated and occur within a pre-associated complex depending on the GIP Na/H exchange regulatory factor-1 (NHERF1). These results raise once again the question of the composition of GPCR-associated complexes [39].
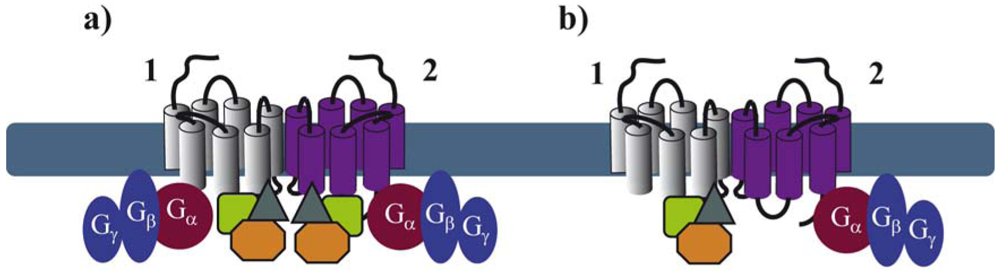
Recent advances in proteomic approaches for the identification of GPCR-associated protein complexes provided a number of further GIPs that directly or indirectly interact with intracellular domains of GPCRs [40,41]. Among these domains, the C-terminal tail and the third intracellular (i3) loop of the receptor have been shown to be the major interacting domains [42]. Many of these proteins appear to bind constitutively to receptors [43,44]. In order to better understand the exact molecular architecture of these complexes, their precise composition, which might be dynamically regulated in space and time, needs to be determined. It is quite logical to anticipate that GPCRs are likely to bind simultaneously, in a constitutive or a ligand-dependent manner, several GIPs. Binding of these proteins to the extended surface of receptor dimers, offers many possibilities for the spatiotemporal interaction of several GIPs with the same receptor. Some of these interactions may depend on receptor domains that are sufficiently distinct to allow simultaneous binding to the receptor, others are likely to overlap. As illustrated above for G proteins, asymmetric binding of GIPs to GPCR dimers might be a solution to accommodate simultaneous binding of several GIPs (Figure 4). This has been recently illustrated in the ternary complex between the melatonin MT1 receptor, Gi proteins and the regulator of G protein signaling (RGS) 20 [44]. This study shows that Gi and RGS20 are both constitutively binding to overlapping, membrane-proximal domains of MT1. These data strongly imply that Gi and RGS20 bind to two different protomers within the homodimer.
Similar observations were made for heteromeric complexes of MT1 with MT2, the second melatonin receptor, suggesting the formation of an asymmetric quaternary complex with MT1 binding to RGS20 and MT2 binding to Gi. This example suggests that a receptor, which is not regulated by a specific GIP, could become sensitive to the action of this GIP when heterodimerizing with a GIP-binding receptor. This latter observation thus provides additional options in fine-tuning GPCR function in a context-dependent manner.
6. Conclusions and Perspectives
GPCR dimerization is a topic of great interest due to the potential importance of the functional consequences of such interactions on signal transduction pathways. Emerging evidence indicates asymmetric function of GPCR dimers and communication between the two protomers of the dimer at the level of ligand binding, ligand-induced conformational changes and interaction with intracellular signaling proteins. Negative and positive allosteric interactions between the two orthosteric binding sites of GPCR homo- and heterodimers may have major consequences on the pharmacology and functional drug outcomes.
Although monomeric GPCRs are able to bind and activate G proteins, formation of receptor dimers clearly offers additional opportunities for regulating G protein activation and signal transduction by accommodating the simultaneous binding of other regulatory GIPs to the receptor.
Finally, formation of higher-order oligomeric clusters needs to be considered in the future. Despite the existence of an extensive literature on GPCR dimerization, the precise oligomerization state of GPCRs remains in most cases unknown. Atomic force microscopy studies provided convincing evidence that rhodopsin is organized as long oligomeric arrays of dimers in rod outer segment membranes [45]. This idea was further extended in molecular docking studies based on the resolved structures of rhodopsin and Gt by proposing an hexameric complex composed of two Gt proteins and a rhodopsin tetramer [46]. These modeling studies obviously warrant experimental confirmation but provide already an interesting framework for the organization and asymmetric binding of different GIPs to GPCR oligomers. Formation of oligomeric complexes might not be restricted to rhodopsin as formation of tetrameric complexes has been proposed for the GABAB receptors, which are composed of two obligatory GABAB heterodimers [47]. Finally, the putative dynamic regulation of GPCR oligomerization might introduce a further level of regulation for GPCR function.
Acknowledgements
This work was supported by grants from SERVIER, the Fondation Recherche Médicale (“Equipe FRM”), Institut National de la Santé et de la Recherche Médicale (INSERM), Centre National de la Recherche Scientifique (CNRS). We thank Nathalie Clément (Institut Cochin, Paris) for help in figure preparation.
References
- Bouvier, M. Oligomerization of G-protein-coupled transmitter receptors. Nat. Rev. Neurosci. 2001, 2, 274–286. [Google Scholar]
- Milligan, G. G protein-coupled receptor hetero-dimerization: contribution to pharmacology and function. Br. J. Pharmacol. 2009, 158, 5–14. [Google Scholar]
- Bulenger, S.; Marullo, S.; Bouvier, M. Emerging role of homo- and heterodimerization in G-protein-coupled receptor biosynthesis and maturation. Trends Pharmacol. Sci. 2005, 26, 131–137. [Google Scholar]
- Rozenfeld, R.; Devi, L.A. Receptor heteromerization and drug discovery. Trends Pharmacol. Sci. 2010, 31, 124–130. [Google Scholar]
- Whorton, M.R.; Bokoch, M.P.; Rasmussen, S.G.; Huang, B.; Zare, R.N.; Kobilka, B.; Sunahara, R.K. A monomeric G protein-coupled receptor isolated in a high-density lipoprotein particle efficiently activates its G protein. Proc. Natl. Acad. Sci. USA 2007, 104, 7682–7687. [Google Scholar]
- Bayburt, T.H.; Leitz, A.J.; Xie, G.; Oprian, D.D.; Sligar, S.G. Transducin activation by nanoscale lipid bilayers containing one and two rhodopsins. J. Biol. Chem. 2007, 282, 14875–14881. [Google Scholar]
- Ernst, O.P.; Gramse, V.; Kolbe, M.; Hofmann, K.P.; Heck, M. Monomeric G protein-coupled receptor rhodopsin in solution activates its G protein transducin at the diffusion limit. Proc. Natl. Acad. Sci. USA 2007, 104, 10859–10864. [Google Scholar]
- Jastrzebska, B.; Maeda, T.; Zhu, L.; Fotiadis, D.; Filipek, S.; Engel, A.; Stenkamp, R.E.; Palczewski, K. Functional characterization of rhodopsin monomers and dimers in detergents. J. Biol. Chem. 2004, 279, 54663–54675. [Google Scholar]
- Parker, M.S.; Sah, R.; Balasubramaniam, A.; Sallee, F.R.; Sweatman, T.; Park, E.A.; Parker, S.L. Dimers of the neuropeptide Y (NPY) Y2 receptor show asymmetry in agonist affinity and association with G proteins. J. Recept. Signal. Transduct. Res. 2008, 28, 437–451. [Google Scholar]
- Palczewski, K.; Kumasaka, T.; Hori, T.; Behnke, C.A.; Motoshima, H.; Fox, B.A.; LeTrong, I.; Teller, D.C.; Okada, T.; Stenkamp, R.E.; Yamamoto, M.; Miyano, M. Crystal structure of rhodopsin: A G protein-coupled receptor. Science 2000, 289, 739–745. [Google Scholar]
- Hamm, H.E. How activated receptors couple to G proteins. Proc. Natl. Acad. Sci. USA 2001, 98, 4819–4821. [Google Scholar]
- Arimoto, R.; Kisselev, O.G.; Makara, G.M.; Marshall, G.R. Rhodopsin-transducin interface: studies with conformationally constrained peptides. Biophys. J. 2001, 81, 3285–3293. [Google Scholar]
- Baneres, J.L.; Parello, J. Structure-based analysis of GPCR function: evidence for a novel pentameric assembly between the dimeric leukotriene B4 receptor BLT1 and the G-protein. J. Mol. Biol. 2003, 329, 815–829. [Google Scholar]
- Damian, M.; Mary, S.; Martin, A.; Pin, J.P.; Baneres, J.L. G protein activation by the leukotriene B4 receptor dimer. Evidence for an absence of trans-activation. J. Biol. Chem. 2008, 283, 21084–21092. [Google Scholar]
- Damian, M.; Martin, A.; Mesnier, D.; Pin, J.P.; Baneres, J.L. Asymmetric conformational changes in a GPCR dimer controlled by G-proteins. EMBO J. 2006, 25, 5693–5702. [Google Scholar]
- Han, Y.; Moreira, I.S.; Urizar, E.; Weinstein, H.; Javitch, J.A. Allosteric communication between protomers of dopamine class A GPCR dimers modulates activation. Nat. Chem. Biol. 2009, 5, 688–695. [Google Scholar]
- Galvez, T.; Duthey, B.; Kniazeff, J.; Blahos, J.; Rovelli, G.; Bettler, B.; Prezeau, L.; Pin, J.P. Allosteric interactions between GB1 and GB2 subunits are required for optimal GABA(B) receptor function. EMBO J. 2001, 20, 2152–2159. [Google Scholar]
- Duthey, B.; Caudron, S.; Perroy, J.; Bettler, B.; Fagni, L.; Pin, J.P.; Prezeau, L. A single subunit (GB2) is required for G-protein activation by the heterodimeric GABA(B) receptor. J. Biol. Chem. 2002, 277, 3236–3241. [Google Scholar]
- Changeux, J.P. Allosteric receptors: from electric organ to cognition. Annu. Rev. Pharmacol. Toxicol. 2010, 50, 1–38. [Google Scholar]
- Schwartz, T.W.; Holst, B. Ago-allosteric modulation and other types of allostery in dimeric 7TM receptors. J. Recept. Signal. Transduct. Res. 2006, 26, 107–128. [Google Scholar]
- Jordan, B.A.; Devi, L.A. G-protein-coupled receptor heterodimerization modulates receptor function. Nature 1999, 399, 697–700. [Google Scholar]
- Novi, F.; Stanasila, L.; Giorgi, F.; Corsini, G.U.; Cotecchia, S.; Maggio, R. Paired activation of two components within muscarinic M3 receptor dimers is required for recruitment of beta-arrestin-1 to the plasma membrane. J. Biol. Chem. 2005, 280, 19768–19776. [Google Scholar]
- Kniazeff, J.; Bessis, A.S.; Maurel, D.; Ansanay, H.; Prezeau, L.; Pin, J.P. Closed state of both binding domains of homodimeric mGlu receptors is required for full activity. Nat. Struct. Mol. Biol. 2004, 11, 706–713. [Google Scholar]
- Parenty, G.; Appelbe, S.; Milligan, G. CXCR2 chemokine receptor antagonism enhances DOP opioid receptor function via allosteric regulation of the CXCR2-DOP receptor heterodimer. Biochem. J. 2008, 412, 245–256. [Google Scholar]
- Urizar, E.; Montanelli, L.; Loy, T.; Bonomi, M.; Swillens, S.; Gales, C.; Bouvier, M.; Smits, G.; Vassart, G.; Costagliola, S. Glycoprotein hormone receptors: link between receptor homodimerization and negative cooperativity. EMBO J. 2005, 24, 1954–1964. [Google Scholar]
- Vivo, M.; Lin, H.; Strange, P.G. Investigation of cooperativity in the binding of ligands to the D(2) dopamine receptor. Mol. Pharmacol. 2006, 69, 226–235. [Google Scholar]
- Springael, J.Y.; Le Minh, P.N.; Urizar, E.; Costagliola, S.; Vassart, G.; Parmentier, M. Allosteric modulation of binding properties between units of chemokine receptor homo- and hetero-oligomers. Mol. Pharmacol. 2006, 69, 1652–1661. [Google Scholar]
- El-Asmar, L.; Springael, J.Y.; Ballet, S.; Andrieu, E.U.; Vassart, G.; Parmentier, M. Evidence for negative binding cooperativity within CCR5-CCR2b heterodimers. Mol. Pharmacol. 2005, 67, 460–469. [Google Scholar]
- Vilardaga, J.P.; Nikolaev, V.O.; Lorenz, K.; Ferrandon, S.; Zhuang, Z.; Lohse, M.J. Conformational cross-talk between alpha2A-adrenergic and mu-opioid receptors controls cell signaling. Nat. Chem. Biol. 2008, 4, 126–131. [Google Scholar]
- Albizu, L.; Cottet, M.; Kralikova, M.; Stoev, S.; Seyer, R.; Brabet, I.; Roux, T.; Bazin, H.; Bourrier, E.; Lamarque, L.; Breton, C.; Rives, M.L.; Newman, A.; Javitch, J.; Trinquet, E.; Manning, M.; Pin, J.P.; Mouillac, B.; Durroux, T. Time-resolved FRET between GPCR ligands reveals oligomers in native tissues. Nat. Chem. Biol. 2010, 6, 587–594. [Google Scholar]
- Rocheville, M.; Lange, D.C.; Kumar, U.; Patel, S.C.; Patel, R.C.; Patel, Y.C. Receptors for dopamine and somatostatin: formation of hetero-oligomers with enhanced functional activity. Science 2000, 288, 154–157. [Google Scholar]
- Durroux, T. Principles: a model for the allosteric interactions between ligand binding sites within a dimeric GPCR. Trends Pharmacol. Sci. 2005, 26, 376–384. [Google Scholar]
- Rondard, P.; Goudet, C.; Kniazeff, J.; Pin, J.P.; Prezeau, L. The complexity of their activation mechanism opens new possibilities for the modulation of mGlu and GABA(B) class C G protein-coupled receptors. Neuropharmacology 2011, 60, 82–92. [Google Scholar]
- Monnier, C.; Tu, H.; Bourrier, E.; Vol, C.; Lamarque, L.; Trinquet, E.; Pin, J.P.; Rondard, P. Trans-activation between 7TM domains: implication in heterodimeric GABA(B) receptor activation. EMBO J. 2010, 30, 32–42. [Google Scholar]
- Daulat, A.M.; Maurice, P.; Froment, C.; Guillaume, J.L.; Broussard, C.; Monsarrat, B.; Delagrange, P.; Jockers, R. Purification and identification of G protein-coupled receptor protein complexes under native conditions. Mol. Cell. Proteomics 2007, 6, 835–844. [Google Scholar]
- Maurice, P.; Daulat, A.M.; Broussard, C.; Mozo, J.; Clary, G.; Hotellier, F.; Chafey, P.; Guillaume, J.L.; Ferry, G.; Boutin, J.A.; Delagrange, P.; Camoin, L.; Jockers, R. A generic approach for the purification of signaling complexes that specifically interact with the carboxy-terminal domain of G protein-coupled receptors. Mol. Cell. Proteomics 2008, 7, 1556–1569. [Google Scholar]
- Breton, B.; Lagace, M.; Bouvier, M. Combining resonance energy transfer methods reveals a complex between the {alpha}2A-adrenergic receptor, G{alpha}i1{beta}1{gamma}2, and GRK2. FASEB J. 2010, 24, 4733–4743. [Google Scholar]
- Tesmer, V.M.; Kawano, T.; Shankaranarayanan, A.; Kozasa, T.; Tesmer, J.J. Snapshot of activated G proteins at the membrane: the Galphaq-GRK2-Gbetagamma complex. Science 2005, 310, 1686–1690. [Google Scholar]
- Klenk, C.; Vetter, T.; Zurn, A.; Vilardaga, J.P.; Friedman, P.A.; Wang, B.; Lohse, M.J. Formation of a ternary complex among NHERF1, beta-arrestin, and parathyroid hormone receptor. J. Biol. Chem. 2010, 285, 30355–30362. [Google Scholar]
- Daulat, A.M.; Maurice, P.; Jockers, R. Recent methodological advances in the discovery of GPCR-associated protein complexes. Trends Pharmacol. Sci. 2009, 30, 72–78. [Google Scholar]
- Bockaert, J.; Perroy, J.; Becamel, C.; Marin, P.; Fagni, L. GPCR interacting proteins (GIPs) in the nervous system: Roles in physiology and pathologies. Annu. Rev. Pharmacol. Toxicol. 2010, 50, 89–109. [Google Scholar]
- Bockaert, J.; Fagni, L.; Dumuis, A.; Marin, P. GPCR interacting proteins (GIP). Pharmacol. Ther. 2004, 103, 203–221. [Google Scholar]
- Guillaume, J.L.; Daulat, A.M.; Maurice, P.; Levoye, A.; Migaud, M.; Brydon, L.; Malpaux, B.; Borg-Capra, C.; Jockers, R. The PDZ protein mupp1 promotes Gi coupling and signaling of the Mt1 melatonin receptor. J. Biol. Chem. 2008, 283, 16762–16771. [Google Scholar]
- Maurice, P.; Daulat, A.M.; Turecek, R.; Ivankova-Susankova, K.; Zamponi, F.; Kamal, M.; Clement, N.; Guillaume, J.L.; Bettler, B.; Gales, C.; Delagrange, P.; Jockers, R. Molecular organization and dynamics of the melatonin MT receptor/RGS20/G(i) protein complex reveal asymmetry of receptor dimers for RGS and G(i) coupling. EMBO J. 2010, 29, 3646–3659. [Google Scholar]
- Fotiadis, D.; Liang, Y.; Filipek, S.; Saperstein, D.A.; Engel, A.; Palczewski, K. Atomic-force microscopy: Rhodopsin dimers in native disc membranes. Nature 2003, 421, 127–128. [Google Scholar]
- Filipek, S.; Krzysko, K.A.; Fotiadis, D.; Liang, Y.; Saperstein, D.A.; Engel, A.; Palczewski, K. A concept for G protein activation by G protein-coupled receptor dimers: the transducin/rhodopsin interface. Photochem. Photobiol. Sci. 2004, 3, 628–638. [Google Scholar]
- Maurel, D.; Comps-Agrar, L.; Brock, C.; Rives, M.L.; Bourrier, E.; Ayoub, M.A.; Bazin, H.; Tinel, N.; Durroux, T.; Prezeau, L.; Trinquet, E.; Pin, J.P. Cell-surface protein-protein interaction analysis with time-resolved FRET and snap-tag technologies: application to GPCR oligomerization. Nat. Methods 2008, 5, 561–567. [Google Scholar]
© 2011 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Kamal, M.; Maurice, P.; Jockers, R. Expanding the Concept of G Protein-Coupled Receptor (GPCR) Dimer Asymmetry towards GPCR-Interacting Proteins. Pharmaceuticals 2011, 4, 273-284. https://doi.org/10.3390/ph4020273
Kamal M, Maurice P, Jockers R. Expanding the Concept of G Protein-Coupled Receptor (GPCR) Dimer Asymmetry towards GPCR-Interacting Proteins. Pharmaceuticals. 2011; 4(2):273-284. https://doi.org/10.3390/ph4020273
Chicago/Turabian StyleKamal, Maud, Pascal Maurice, and Ralf Jockers. 2011. "Expanding the Concept of G Protein-Coupled Receptor (GPCR) Dimer Asymmetry towards GPCR-Interacting Proteins" Pharmaceuticals 4, no. 2: 273-284. https://doi.org/10.3390/ph4020273
APA StyleKamal, M., Maurice, P., & Jockers, R. (2011). Expanding the Concept of G Protein-Coupled Receptor (GPCR) Dimer Asymmetry towards GPCR-Interacting Proteins. Pharmaceuticals, 4(2), 273-284. https://doi.org/10.3390/ph4020273